

Summary
Cell surface CD147 protein promotes production of matrix metalloproteinases (MMPs) and hyaluronan, associates with monocarboxylate transporters (MCTs) and integrins, and is involved in reproductive, neural, inflammatory, and tumor functions. Here we combine covalent cross-linking, mass spectrometric protein identification and co-immunoprecipitation to show selective CD147 association with three major types of transporter (CD98hc-LAT1, ASCT2, and MCTs), as well as a regulator of cell proliferation (EpCAM). In the assembly of these multicomponent complexes, CD147 and CD98hc play a central organizing role. RNAi knockdown experiments establish a strong connection between CD147 and CD98hc expression and a strong positive association of CD147 (and CD98hc) with cell proliferation. As the CD147-CD98hc complex and proliferation diminished, AMP-activated protein kinase (AMPK, a cellular ‘fuel gauge’), became activated, indicating a disturbance of cellular energy metabolism. Our data point to a CD147-CD98 cell surface super-complex that plays a critical role in energy metabolism, likely by coordinating transport of lactate and amino acids. Furthermore, we show how covalent crosslinking, together with mass spectrometry, can be used to identify closely associated transmembrane proteins. This approach should also be applicable to many other types of transmembrane proteins besides those associated with CD98hc and CD147.
Abbreviations: AMPK: AMP-activated protein kinase, BS3: bis(sulfosuccinimidyl)suberate, DSP: dithiobis(succinimidylpropionate), EMMPRIN: extracellular matrix metalloproteinase inducer, EpCAM: epithelial cell adhesion molecule, LAT1: L-type amino acid transporter 1, MCTs: monocarboxylate transporters, MMPs: matrix metalloproteinases, mTOR: mammalian target of rapamycin, SDFR1: stromal cell derived factor receptor 1, TCSF: tumor cell-derived collagenase-stimulatory factor
Introduction
CD147 (EMMPRIN, basigin, neurothelin, TCSF, OX-47, 5A11, CE9, gp42, M6) is a cell surface protein with multiple glycosylated forms (1;2). CD147 is ubiquitously expressed, with highest levels on metabolically active cells, such as lymphoblasts (3), inflammatory cells (4), brown adipocytes (5), and malignant tumors (1). CD147 promotes production of matrix metalloproteinases (MMPs) (1) and hyaluronan (6), and is involved in reproduction, neural function, inflammation, tumor invasion, and HIV infection (7;1;8). As a chaperone for monocarboxylate transporters (MCT1 and MCT4), CD147 enables insertion of MCT1 and MCT4 into cell membranes, which facilitates import and/or export of lactate and pyruvate (9;10). CD147 also interacts with β1 integrin (11), cyclophilin A (7), and caveolin-1 (12). Caveolin-1 association appears to restrict CD147 glycosylation and function (2). Consistent with the functional importance of CD147, CD147−− mice are sterile, have reduced body weight, and show impaired spermatogenesis, sensory, learning and memory functions. Half of the surviving mice subsequently die of pneumonia (13). In some genetic backgrounds, the majority of mouse embryos lacking CD147 die at around the time of implantation (14).
Emerging proteomics technologies are helping to elucidate cellular protein-protein interaction networks (15;16). However elucidation of cell surface transmembrane protein interactions has lagged behind, often due to issues involving detergent solubilization. To understand better the remarkably diverse functions of CD147, we sought to identify its major cell surface protein partners. First we used homobifunctional cross-linking agents to stabilize protein interaction networks on intact cells, and then we lysed cells using relatively harsh detergent conditions to disrupt non-crosslinked complexes. Next we immuno-isolated crosslinked CD147 complexes, and used nanoscale LC-MS/MS to identify all transmembrane proteins in these complexes. Our data point to CD147 interacting not only with monocarboxylate transporters, but also with amino acid transporters (CD98hc-LAT1, ASCT2) and a regulator of cell proliferation (EpCAM).
Like CD147, CD98hc (CD98 heavy chain, 4F2, FRP-1) also is a multifunctional glycoprotein with a single-transmembrane domain, is highly expressed on proliferating cells, and functions as a chaperone for transporters (17;18). Indeed, CD98hc forms disulfide-bonded heterodimers with at least six different light chains (LAT1, LAT2, y+LAT1, y+LAT2, Asc-1, xCT) that serve as amino acid transporters (18). In addition, CD98hc may regulate cell fusion (19), and integrin-dependent adhesion functions (20;21), while associating either indirectly (22) or directly (23) with β1 integrins. Our detailed analysis of CD98hc complexes confirmed CD98 association with CD147, monocarboxylate transporters, amino acid transporters (LAT1, ASCT2) and EpCAM. Furthermore, RNAi depletion of either CD147 or CD98hc diminished cell surface expression of both molecules and diminished cell proliferation. Together these results point to CD147 and CD98hc playing a central organizing role within a “super-complex” that is critical for cellular energy metabolism.
Experimental Procedures
Antibodies
We used antibodies to CD147 [mAb 8G6; pAb B10 (11)], CD98hc [mAb 4F2 (24); pAb C-20, Santa Cruz], EpCAM [mAb KS1/4, BD Biosciences; pAb H-70, Santa Cruz)], β1 integrin [mAb TS2/16], MHC class I [mAb W6/32], Na+/K+ ATPase-α1 [mAb C464.4, Santa Cruz], AMPK-α and phospho-AMPK-α [Cell signaling], MCT1 [pAb from Alpha Diagnostics], and β tubulin [mAb Tub2.1, Sigma)]. Anti-GFP [mAb 3E6, Qbiogene] was used for immunoprecipitation (IP), and anti-FLAG (mAb M2 and M2-agarose, Sigma-Aldrich) was used for IP and immunoblotting.
Cross-linking and LC-MS/MS
Intact cells were grown to 90% confluence in five 150-mm plates and then were washed three times with PBS. Cross-linkers DSP or BS3 (Pierce) were added to 1 mM (final concentration) in 20 mM Hepes buffer, pH7.5, 150 mM NaCl, and 5 mM MgCl2 for 30 min at RT, or for 1 hour at 4 ºC, before termination with 25 mM Glycine (pH 7.5). After washing with PBS, cells were lysed with lysis buffer [25 mM Hepes, pH 7.5, 150 mM NaCl, and 5 mM MgCl2 supplemented with 1% Triton X-100 (Roche) with proteinase inhibitor cocktail (Roche)] at 4°C for 1hr. Lysate was centrifuged at 20,000 g for 30 min, and the supernatant was precleared with 2 ml of protein A-agarose for 2 hrs at 4°C. After centrifugation (6000g, 20 min), the supernatant was mixed with 1 ml of anti-FLAG M2-agarose (Sigma) or mAb 4F2-conjugated beads, and shaken overnight at 4 ºC overnight. Beads were then washed six times with 30 ml of lysis buffer. Complexes were eluted with 2.5 ml of 100mM glycine, pH 2.5, and then neutralized with 200 μl of 1 M Tris-HCl, pH 7.5. Concentrated eluate (Centricon, Millipore) was resolved by SDS-PAGE, using reducing conditions for BS3 crosslinked samples and nonreducing conditions for DSP crosslinked samples. All coomassie blue stained bands larger than 40 kD were excised and sent to the Taplin Biological Mass Spectrometry Facility (Harvard Medical School). For protein identification, excised SDS-PAGE gel bands were chopped into 1 mm3 pieces and in-gel digestion with trypsin was performed as described (25). All data were acquired by nanoscale microcapillary liquid chromatography coupled to a linear ion trap mass spectrometer (ThermoElectron, San Jose, CA) as described (26). Briefly, a gradient of increasing organic modifier eluted peptides into the mass spectrometer. The instrument was set to cycle between collecting one survey scan followed by five MS/MS scans on the five most abundant ions with dynamic exclusion of ions previously selected. MS/MS spectra were extracted using Bioworks 3.1 and searched with the Sequest (version 27) algorithm against the nonredundant human database from NCBI, which contained 237,384 sequences. Typical ion trap parameters were used including a peptide tolerance of 2.0 Da, default fragment ion tolerance, variable modification of methionine (+16), no enzyme specificity and up to 3 missed cleavages allowed. Peptide matches were deemed correct when two or more high-scoring, fully-tryptic peptides matched to a protein. Xcorr values of 1.8, 1.8, and 3.0 were used for 1+, 2+, and 3+ peptides, respectively, and no dCorr threshold was used. Peptide matches with three or fewer tryptic matches were manually verified. Also, there was no smoothing of data, signal to noise criteria, charge state determination or peak de-isotoping utilized for these experiments.
Immunoprecipitation and Western blotting
Cells grown in 100-mm plates were lysed with 1 ml of lysis buffer for 1 h at 4 °C. Centrifugation and preclearing were carried out as above. For each immunoprecipitation, 2–4 μg antibody with 50 μl protein A (or protein G)-agarose beads or 50 μl of FLAG-agarose beads were used, with shaking at 4º overnight. Beads were then washed three times and immunecomplexes were eluted using 60 μl of 1 x Laemmli sample buffer (non-reducing) at 100°C for 2 min. Sample aliquots (20 μl) were resolved using SDS-PAGE, and immunoblotting and flow cytometry were carried out as described previously (11). In some experiments, after blotting, relative band densities were assessed quantitatively using GeneTools version 3 (Syngene Laboratories, Frederick, MD).
Cell lines
Human cell lines HT1080 (fibrosarcoma), SW480 (colorectal adenocarcinoma) and MCF7 (breast adenocarcinoma) from ATCC were maintained in DMEM medium supplemented with 10% fetal calf serum (FCS), and (for MCF7 cells) an additional 10 μg/ml insulin. CD147 and SDFR1 (GenBank GI:20552516, stromal cell derived factor receptor 1, with 40% sequence identity with CD147) were C-terminally FLAG-tagged, and CD98hc and LAT1 were N-terminally FLAG-tagged and inserted into pcDNA3.1(+) or pLXIZ vectors. Quantitation of immunoblotted CD147 was carried out using pAb B10. Ectopic CD147-FLAG expression was 1.4-fold higher than endogenous CD147 in HT1080 cells; 1.8-fold higher in HEK293 cells and 2.1-fold higher in MCF7 cells. Similarly, we blotted with anti-FLAG antibody to show that ectopic CD147-FLAG and SDFR1-FLAG were expressed at comparable levels to each other in HT1080, HEK293 and MCF7 cells. We also prepared C-terminal GFP-tagged CD147 and SDFR1 and N-terminal GFP-tagged LAT1, using pEGFP-C3 or pEGFP-N1 vectors (Clontech). Cells were either stably transfected using FuGENE 6 (Roche) or infected, and then selected using G418 or Zeocin (Invitrogen) for 4–6 weeks.
Oligonucleotides for CD147, CD98hc RNAi
Sense and antisense oligonucleotides were cut with BamHI and Hind III, and purified fragments were ligated into RNAi plasmid pSilencer 3.1 H1 Hygro (Ambion), and confirmed by DNA sequencing. The RNAi negative plasmid is from the same kit (Ambion). Cells were transfected with FuGENE 6 (Roche). Stable cells were selected for about 4 to 8 weeks in 200 μg/ml Hygomycin (Roche). Oligonucleotide sequences used for CD147 and CD98hc RNAi experiments were as follows: CD147 RNAi Oligonucleotide, Pair 1: sense: (BamHI ) 5’-CG↓GATCCCGACCTTGGCTCCAAGATACTTCAAGAGAGTATCTTGGAGCCAAGGTCTTTTTGGAAA↓AGCTTGGGC-3’ (Hind III); antisense: (Hind III) 5’-GCCCA↓AGCTTTTCCAAAAAGACCTTGGCTCCAAGATACTCTCTTGAAGTATCTTGGAGCCAAGGTCGG↓GATCCGG-3’(BamHI). Pair 2: sense: (BamHI) 5’-CCG↓GATCCCGGCCGTGAAGTCGTCAGAATTCAAGAGATTCTGACGACTTCACGGCCTTTTTGGAAA↓AGCTTGGGC-3’ (Hind III); antisense: (HindIII) 5’-GCCCA↓AGCTTTTCCAAAAAGGCCGTGAAGTCGTCAGAATCTCTTGAATTCTGACGACTTCACGGCCGG↓GATCCGG (BamHI). CD98hc RNAi Oligonucleotide sequences, pair 1: sense: (BamHI) 5’-CCG↓GATCCCGTGAGTAGAGCCCGAGAAGTTCAAGAGACTTCTCGGGCTCTAACTCATTTTTTGGAAA↓AGCTTGGG C-3’ (Hind III); antisense: (HindIII) 5’-GCCCA↓AGCTTTTCCAAAAAATGAGTTAGAGCCCGAGAAGTCTCTTG AACTTCTCGGGCTCTAACTCACGG↓GATCCGG (BamHI). Pair 2: sense: (BamHI) 5’-CCG↓GATCCCGGATGCATCCTCATTCTTGTTCAAGAGACAAGAATGAGGATG CATCCTTTT TTGGAAA↓AGCTTGGGC-3’ (Hind III); antisense: (HindIII) 5’-GCCCA↓AGCTTTTCCAAAAAAGGATGCATCCTCATTCTTGTCTCTTGAACAAGAATGAGGATGCATCCGG↓GATCCGG-3’ (BamHI).
Cellular proliferation assay and statistical analysis
Cellular proliferation/survival was measured using the MTT (Roche) colorimetric method. Cell suspensions (1 × 104 cells/100 μl) were plated into 96-well plates with 4 wells for each cell clone in complete medium. Control wells contained either mitomycin C (Sigma)(1, 2.5 or 5 μg/ml) or DMEM without serum. The plates were incubated for 3 or 4 days at 37°C prior to the addition of MTT solution, and measurement of absorbance at 560 nm. Each experiment was repeated three times. The relative proliferation was calculated compared with mean of RNAi control. Data on CD147 and CD98hc expression levels were assessed by flow cytometry. Each experiment was repeated three times. The correlation between CD147 expression and relative proliferation was analyzed using VassarStats statistics (http://faculty.vassar.edu/lowry/VassarStats.html).
Results
CD147 protein partners identified by LC-MS/MS
FLAG-tagged CD147 was expressed in HT1080 and MCF7 cells at levels only a little higher (1.4–2.1-fold) than endogenous CD147. Intact HT1080 cells (Fig. 1, left lane) and MCF7 cells (Fig. 1, right lane) were then treated with chemical crosslinkers (non-reducible BS3 or reducible DSP respectively), cells were lysed in Triton X-100, CD147 complexes were isolated, and then proteins within the indicated gel regions were identified using LC-MS/MS analysis. A summary of results is shown in Table I, for CD147 complexes isolated from MCF7 epithelial cells (Exp. I), HT1080 fibrosarcoma cells (Exp. II) or HT1080 co-cultured with MCF7 cells (Exp. III). Proteins identified in all three experiments, besides CD147 itself, were CD98hc, and the monocarboxylate transporters MCT1 and MCT4. Proteins appearing in at least one experiment were ASCT2 (neutral amino acid transporter), LAT1 (L-type amino acid transporter 1, a CD98 light chain), EpCAM (epithelial cell adhesion molecule), an additional MCT family protein (MCT8), β1 integrin, plus others listed in Table I. Use of covalent crosslinkers allowed isolation of CD147 partners under relatively stringent (1% Triton X-100) conditions. For comparison, an additional experiment was carried out using a mild detergent (1% Brij 58) with no crosslinking (Table I, Exp IV). CD147 complexes again contained CD98hc, ASCT2, and the other proteins listed in Table I, Exp. IV, plus a few additional proteins (not listed) that appeared only under these less stringent conditions. Results such as seen in Table 1 were consistently obtained when we isolated CD147 complexes while using anti-FLAG antibody to C-terminal FLAG-tagged CD147. In contrast, monoclonal and polyclonal antibodies to CD147 extracellular domains failed to yield many associated proteins (e.g. see Fig. 5 below, and not shown), presumably because the associated proteins were blocking access of the antibodies to CD147.
Fig. 1.
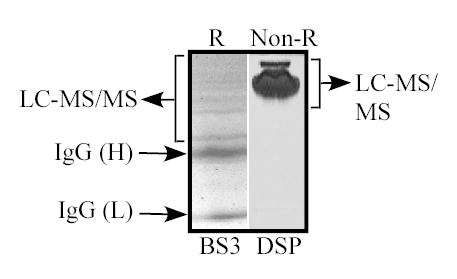
Representative samples of CD147-associated proteins. HT1080 cells were treated with non-reducible crosslinker BS3, CD147-FLAG complexes were isolated from 1% Triton X-100 lysates, and then samples were resolved using reducing conditions (left lane). MCF7 cells were treated with reducible crosslinker DSP, CD147-FLAG complexes were isolated from 1% Triton X-100 lysates, and samples were resolved using non-reducing conditions (right lane). After staining with Coomassie Blue, the indicated regions were excised for LC-MS/MS analysis.
Table 1.
Membrane proteins identified in CD147 and CD98 complexes by LC-MS/MS
| CD147 | CD98hcd | ||||||
|---|---|---|---|---|---|---|---|
| Name (GenBank GI No) | I | II | III | IV | V | VI | VII |
| CD147 (34448) | +(7)* | +(10) | +(8) | +(8) | - | - | - |
| CD98hc (182864) | +(13) | +(6) | + | +(6) | +(23) | +(14) | +(20) |
| ASCT2 (1478280) | +(5) | +(3) | - | +(4) | + | - | + |
| LAT1 (3639057)b | +(3) | - | - | - | - | + (3) | + (3) |
| EpCAM (186775) | + | - | - | - | + | + | + |
| MCT1 (561742)a | + | +(3) | +(2) | - | - | - | - |
| MCT4 (2463633)a | + | +(4) | + | - | - | - | - |
| MCT8 (458253) | + | - | - | - | - | - | - |
| CD29 (Integrin β 1) (31441)ab | - | +(3) | - | - | - | - | - |
| Na/K ATPase, α 1 (219941)c | +(2) | +(5) | - | +(2) | - | + | + |
| Na/K ATPase, β 3 (1522634)c | - | - | - | - | + | - | - |
| Glycopr. Trop-2 (31590) | + | - | - | - | - | - | + |
| I3 bp (17985370) | + | - | - | - | - | - | - |
| CD71 (37432)c | + | +(2) | - | +(2) | - | - | - |
| Sel-1 homolog (6683457) | +(2) | - | - | +(2) | - | - | - |
| CD166 (886257) | - | +(4) | - | - | - | - | - |
| CD49e (Int. α 5) (31437) | - | + | - | - | - | - | - |
| CD44 (337955) | - | + | - | - | - | - | - |
| Leuk. virus recpt.1 (306769) | - | + | - | - | - | - | - |
| HLA-I, A-25 (187794) | - | + | - | - | - | - | - |
I. MCF7/CD147-FLAG cells, DSP cross-linker; II. HT1080/CD147-FLAG cells, BS3 cross-linker; III. HT1080/CD147-FLAG cells were co-cultured with MCF7 cells, and then treated with BS3 cross-linker; IV. MCF7/CD147-FLAG cells, IP: FLAG in 1% Brij 58, no crosslinker. V. MCF7/CD98hc-FLAG cells, BS3 cross-linking, IP FLAG in 1% Triton X-100. VI. MCF7 cells, DSP cross-linking, IP 4F2-beads in 1% Triton X-100. VII. MCF7/CD98hc-FLAG cells were co-cultured with HT1080 cells. BS3 cross-linking, IP FLAG in RIPA. Molecules with names in bold letter were validated in independent biochemical co-immunoprecipitation experiments.
number of peptides from LC-MS/MS if >1. The sequences of all peptides are listed in Supplemental Table S1.
These results were not consistently observed in further biochemical experiments.
CD98 experiments were carried out using cells (MCF7) with little CD147, to allow assessment of CD98hc partners independent of CD147.
Fig. 5.
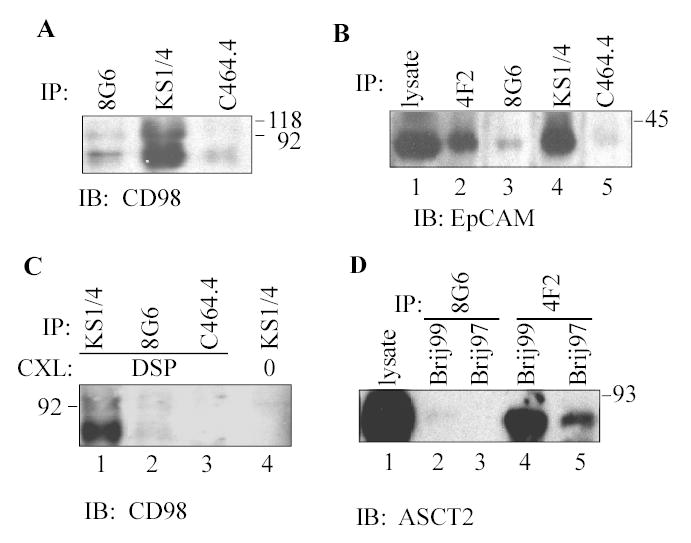
Association of CD98hc with EpCAM and ASCT2. A) SW480 cells were lysed in 1% Brij 99. Endogenous EpCAM (mAb KS1/4), CD147 (mAb 8G6) or Na/K ATPase α1 (mAb C464.4) were immunoprecipitated and then blotted for endogenous associated CD98hc using pAb C-20. B) SW480 cells were lysed in 1% Brij 99, endogenous CD98hc (lane 2), CD147 (lane 3), EpCAM (lane 4) and Na/K ATPase-α1 (lane 5) were immunoprecipitated, and samples were blotted for associated EpCAM. C) Intact MCF7 cells were treated with crosslinker (1 mM DSP), and lysed in 1% Triton X-100. Endogenous EpCAM, CD147, and Na/K ATPase, were immunoprecipitated and blotted for CD98hc. D) HEK293 cells were lysed in 1% Brij 99 or Brij 97, CD147 (mAb 8G6) and CD98hc (mAb 4F2) were immunoprecipitated, and associated ASCT2-FLAG was detected by blotting with anti-FLAG antibody.
Confirmation of CD147 association with CD98hc
While suggestive, results in Table 1 are neither definitive nor quantitative. To firmly establish direct CD147-CD98hc association, we used covalent crosslinking and co-immunoprecipitation. Following either DSP (thiol cleavable) or BS3 (uncleavable) cross-linking, immunoprecipitation of CD147 yielded endogenous CD98hc from MCF7 (Fig. 2A; lanes 2, 3), HT1080 (Fig. 2B; lanes 2, 3) and HEK293 (Fig. 2C, lanes 4, 6) cells. No CD98hc was obtained when crosslinker was omitted (Figs. 2A, B, lanes 1, 4; Fig. 2C, lanes 1,2), or antibodies to control SDFR1 protein were utilized (Figs 2A, B, lanes 4–6; Fig. 2C, lanes 3,5). Monomeric CD98hc appears as two bands (~85 and 92 kDa) likely due to variable glycosylation. The larger size of CD98hc (140–230 kDa) captured in CD147 complexes from uncleavable BS3-crosslinked lanes is consistent with endogenous CD98hc being covalently crosslinked to CD147 (~50 kD) plus additional components. One likely additional component is the known CD147 partner MCT1. As indicated in Fig. 2D (lane 3), MCT1 (45 kDa) crosslinked to CD147 (50 kDa) yielded a 95 kDa complex, plus a larger complex (~140–230 kDa) similar in size to that containing CD98hc. In the absence of crosslinker, MCT1 (~45,000) associated with CD147 in Brij 97 lysate (Fig. 2E), but not in Triton X-100 (not shown).
Fig. 2.
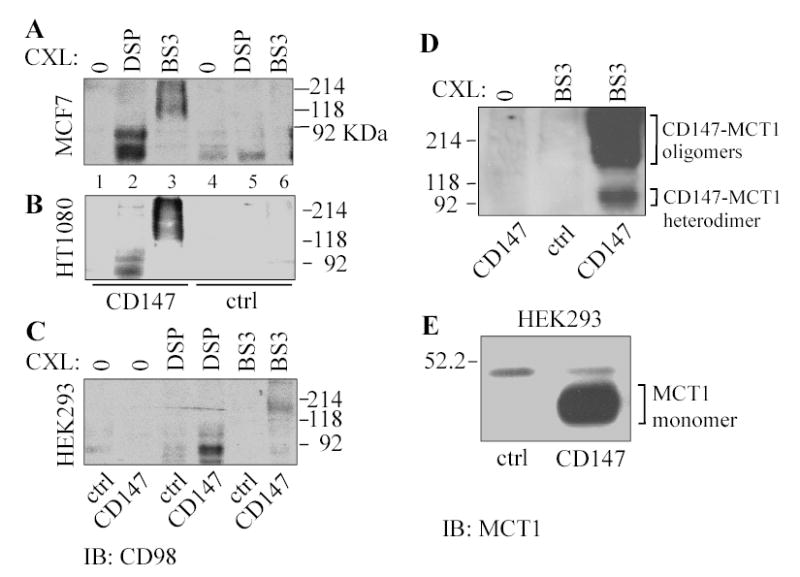
Crosslinked CD147 complexes contain CD98hc. A) Transfected MCF7 cells were treated with no cross-linker (lanes 1, 4), DSP (lanes 2, 5) or BS3 (lanes 3, 6), and lysed using 1% Triton X-100. CD147 (lanes 1-3) and control SDFR1 (lanes 4–6) were immunoprecipitated using anti-FLAG antibody, and samples were blotted for endogenous CD98hc using pAb C-20. B) Transfected HT1080 cells were treated with or without crosslinker (as in Part A), CD147-FLAG, or negative control (SDFR1-FLAG) was immunoprecipitated, and samples were blotted for endogenous CD98hc. C) Transfected HEK293 cells were treated with or without crosslinker (as indicated), CD147-FLAG or control SDFR1-FLAG was immunoprecipitated, and samples were blotted blot for endogenous CD98hc. For cells used in panels A–C, CD147-FLAG and SDFR1-FLAG were expressed a comparable levels, as indicated by anti-FLAG blotting (not shown). D) Intact HEK293 cells were crosslinked (1 mM BS3), lysed in 1% Triton X-100, and then CD147-FLAG was immunoprecipitated and samples were blotted for MCT1. E) HEK293 cells were lysed in 1% Brij 97, CD147-FLAG was immunoprecipitated, and samples were blotted for MCT1.
In the absence of crosslinker, CD147-CD98hc complexes were also stable in Brij 97, as well as CHAPS and Brij 99, but were almost completely disrupted in Triton X-100 (Fig. 3A). Immunoprecipitation of CD147 yielded CD98hc when the two molecules were co-expressed together in HEK293 cells (Fig. 3B, lane 8), but not when expressed separately and then the cells (Fig. 3B, lane 5) or lysates (lane 4) were mixed. Hence, CD147 forms a cis-interacting complex with CD98hc. Negative control SDFR1 did not associate with CD98hc, whereas positive control LAT1 (an established CD98 light chain) did associate with CD98hc (Fig. 3B, lanes 7, 6).
Fig. 3.
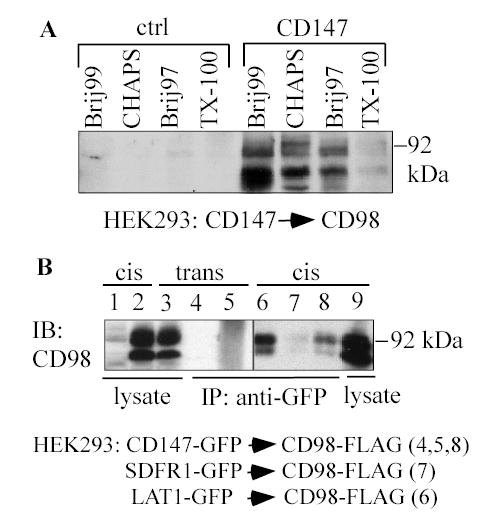
Further characterization of CD147-CD98hc complexes. A) HEK293 cells were lysed in the indicated detergents (each at 1%), CD147-FLAG, or negative control SDFR1-FLAG was immunoprecipitated, and samples were blotted for endogenous CD98 using pAb C-20. B) HEK293 cells were lysed 1% Brij 99, CD147-GFP (lane 8), SDFR1-GFP (lane 7) or LAT1-GFP (lane 6) was immunoprecipitated, and CD98-FLAG (from the same cell lysate) was detected using anti-FLAG antibody. For trans experiments, HEK293 cells (lane 5) or lysates (lane 4) containing CD147-GFP were mixed with cells or lysates containing CD98hc-FLAG. Then CD147-GFP was immunoprecipitated and CD98-FLAG was detected by blotting (lanes 4, 5). Also, CD98hc-FLAG is blotted from control lysates of HEK293-CD147-GFP cells (lane 1), HEK293-CD98-FLAG cells (lane 2), mixed HEK293-CD147-GFP and HEK-CD98-FLAG cells (lane 3) and HEK293 co-expressing both CD147-GFP and CD98hc-FLAG (lane 9).
In a reciprocal experiment, CD98hc immunoprecipitation yielded a large complex (170–230 kDa) containing endogenous CD147 when covalent cross-linker (BS3) was added to HEK293 cells prior to Triton X-100 lysis (Fig. 4A, lane 9), but not when BS3 was omitted (lane 8). In mild Brij 99 or CHAPS detergent, CD147 was obtained even without cross-linking (Fig. 4A, lanes 5, 6). Additional data, involving CD147 domain deletion and swapping, point to CD147-Ig domain 1 being critical for CD98 association (not shown). Immunoprecipitation of CD98hc also yielded MCT1 (Fig. 4C, lanes 5, 6), which associates directly with CD147 (9), and as we confirmed in Figs. 2D and 2E. The association between CD98hc and MCT1 was seen in mild detergents (1% Brij 99, CHAPS; Fig. 4C, lanes 5, 6), but not in more stringent detergent conditions (Brij 97; lane 7) under which MCT1 still associated with CD147 (Fig. 2E). Moreover, MCT1 was only minimally cross-linked with CD98hc (Fig. 4C, lane 9). Rather, most of the MCT1 (45 kDa) was directly crosslinked to CD147 (50 kDa) to yield a heterodimer of ~ 95 KD (Fig. 4A, lane 2; Fig. 2D, lane 3).
Fig. 4.
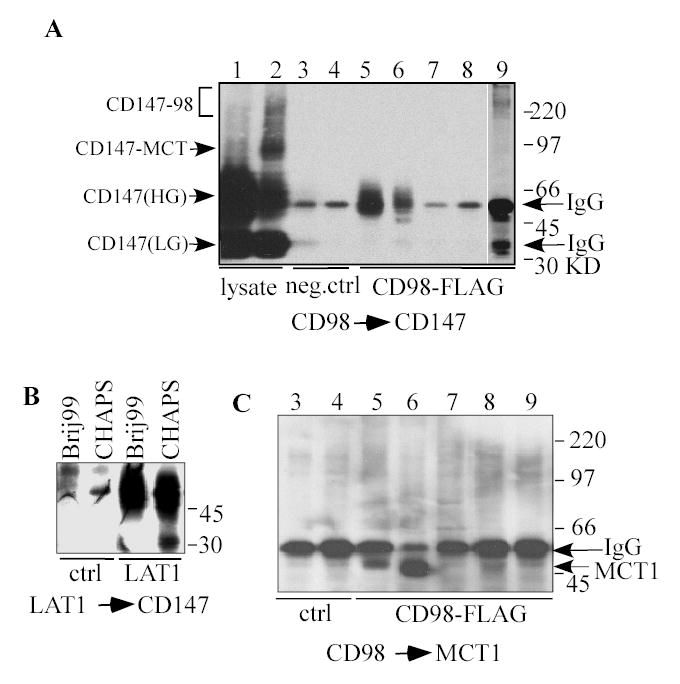
CD98hc (and LAT1) co-precipitate CD147 (and MCT1). A) HEK293 cells expressing CD98hc-FLAG (lanes 1, 2, 5–9) or FLAG vector alone (lanes 3,4) were lysed using Triton X-100 (lanes 1, 2, 4, 8, 9), Brij 99 (lanes 3, 5), CHAPS (lane 6) or Brij 97 (lane 7). A portion of cells (used for lanes 2, 9) was treated with BS3 crosslinker. Following immunoprecipitation of CD98hc-FLAG or vector control-FLAG using anti-FLAG antibody, samples were blotted for CD147. CD147(HG) = high glycosylated form; CD147(LG) = low glycosylated form (2). Note: the membrane used in part A was first used in part C and then stripped and re-probed with anti-CD147 antibody B10. B) LAT1-FLAG was immunoprecipitated from HEK293 cells (lysed in Brij 99 or CHAPS), and samples were blotted for CD147. C) CD98hc immunoprecipitates (prepared as in panel A) were blotted for MCT1.
Associations mediated through CD98hc
It seemed possible that some potential CD147 partners from Exp I–IV of Table 1 (e.g.LAT1, EpCAM, ASCT2) could be more proximal to CD98 than CD147. To explore this further, we identified CD98hc partners when CD147 expression was minimal. To achieve this, we used CD98hc-FLAG-expressing MCF7 cells with low endogenous levels of CD147, as detected by flow cytometry and by Western Blotting (not shown). After cross-linking of intact cells with either BS3 or DSP, cells were lysed in Triton X-100, and we isolated CD98hc-FLAG complexes (Table I, Exp. V, VII) or endogenous CD98hc complexes (Exp. VI) from MCF7 cells alone (Exps. V, VI) or after co-culture with HT1080 cells (Exp. VII). Again LC-MS/MS analyses revealed the presence of LAT1, EpCAM and ASCT2, plus CD98hc itself. Despite elevated CD147 expression in HT1080 cells, HT1080-CD147 was not identified among the protein partners of MCF7-CD98hc (Exp. VII), consistent with CD98hc and CD147 not interacting in trans (as established in Fig. 3B). For some proteins listed in Table I (CD71, Na+/K+ ATPase), we could not confirm CD98hc (or CD147) association in subsequent biochemical experiments (e.g. see Fig. 5A–C). These and other proteins appearing twice or less in 7 experiments (Table I, bottom) were not studied further.
LAT1, a Na+-independent transporter for large, neutral amino acids, naturally and directly associates with CD98hc via a disulfide bond (27). Although LAT1 immunoprecipitation did yield CD147 from mild Brij 99 and CHAPS lysates (Fig. 4B), LAT1 did not co-immunoprecipitate CD147 in more stringent detergent conditions (Triton X-100, Brij 97) either with or without crosslinking (not shown). These results are consistent with a CD147-CD98hc-LAT1 linkage. Confirming results in Table I (Exps. V-VII), CD98hc from SW480 cells also co-immunoprecipitated (Fig. 5A, lane 2) with EpCAM, an epithelial cell surface protein involved in tumor proliferation (28;29). Conversely, immunoprecipitation of endogenous CD98hc from SW480 cells yielded endogenous EpCAM (Fig. 5B, lane 2). EpCAM-CD98hc complexes were stable in 1% Brij 99 (Figs 5A, 5B), and CHAPS and Brij 97 (data not shown), but not in Triton X-100 (Fig. 5C, lane 4) unless captured using the covalent cross-linker DSP (Fig. 5C, lane 1). In each of these experiments, antibody (mAb 8G6) to endogenous CD147 failed to yield much CD98 (Fig. 5A, lane 1; 5C lane 2) or EpCAM (Fig. 5B, lane 3), at least partly because the 8G6 epitope (on extracellular Ig domain 1) is blocked by CD147-associated proteins (not shown). Although Na/K ATPase α1 was prominently expressed in SW480 cell lysates, an antibody (mAb C464.4) to that protein yielded minimal CD98 or EpCAM, and hence was used as a negative control in Fig. 5A-C. As an additional negative control, anti-β1 integrin mAb TS2/16 also failed to yield EpCAM from SW480 cells.
Another protein in Table I, associating with both CD147 and CD98hc, is the neutral amino acid transporter, ASCT2 (30). Confirming this result, CD98hc immunoprecipitation from HEK293 cells yielded ASCT2 in both Brij 99 and Brij 97 cell lysates (Fig. 5D, lanes 4, 5). However, immunoprecipitation with anti-CD147 mAb 8G6 failed to yield ASCT2 (Fig. 5D, lanes 2, 3), again because the 8G6 epitope is likely blocked. Treatment of HEK293 cells with DSP crosslinker prior to lysis allowed immunoprecipitation of endogenous CD98 and recovery of ASCT2, even in 1% Triton X-100 (not shown).
Correlation between CD147, CD98hc expression and cell proliferation
To explore further the CD147-CD98hc super-complex, we used RNAi to knock down either CD147 or CD98hc. CD147 was reduced up to 80% in HEK293 cells as seen by immunoblotting (Fig. 6A) and flow cytometry (Fig. 6B, top panel). The same RNAi that diminished CD147, also reduced CD98hc expression (Fig. 6B, lower panel). Conversely, knockdown of CD98hc caused a loss of CD147. Indeed, cell surface expression data for 14 different CD147-depleted clones and 13 different CD98hc-depleted clones each showed strikingly parallel effects on both CD147 and CD98hc expression (Fig. 7A). Levels of other prominent cell surface proteins (β1 integrin, MHC Class I), were unaffected by CD147 or CD98hc depletion (not shown).
Fig. 6.
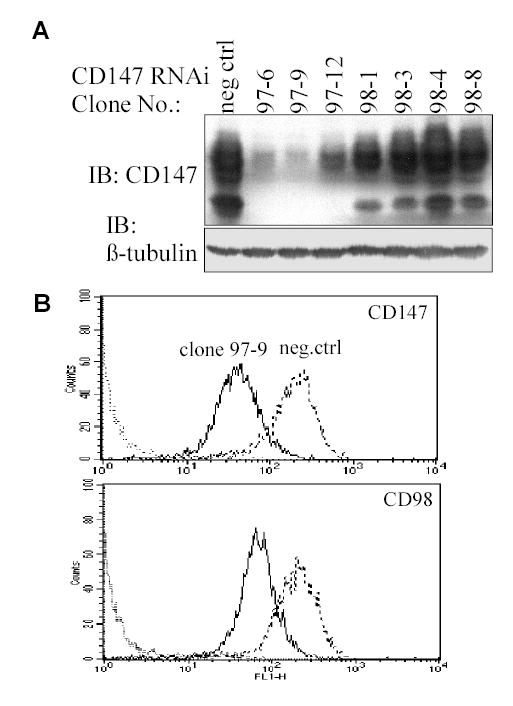
Interdependent expression of CD147 and CD98hc. A) Lysates from CD147 RNAi-depleted HEK293 clones were blotted for CD147 and β tubulin. B) CD147-depleted HEK293 cells (clone 97-9) were analyzed by flow cytometry for CD147 (mAb 8G6, upper panel) and CD98 (mAb 4F2, lower panel). Negative control cells (dotted lines) were treated with non-depleting control RNAi.
Fig. 7.
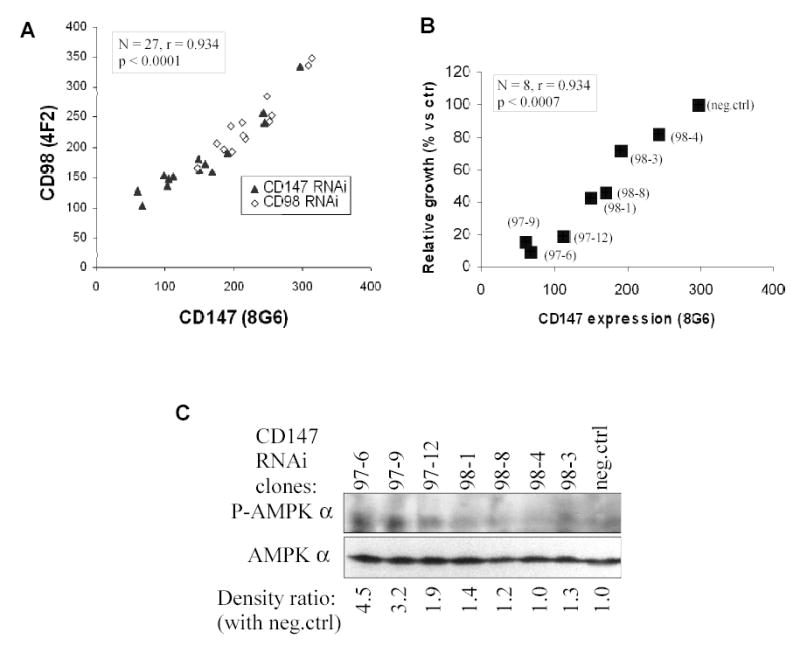
Functional co-regulation of CD147 and CD98hc. A) Cell surface expression levels (in MFI units) are plotted for CD98hc (y axis) and CD147 (x axis) for each clone of CD147- and CD98-RNAi depleted cells. B) Relative proliferation rates for CD147-depleted clones are plotted vs cell surface CD147 expression levels. For each result, N = 3, and SEM ≤5% of mean proliferation rate. Relative proliferation rates are shown relative to RNAi negative control rates. C) CD147-depleted HEK293 clones were lysed in RIPA with proteinase inhibitor cocktail supplemented with 2 mM PMSF, 1 mM Na3VO4, NaF, and then detected for expression of phosphorylated AMPKα and total AMPKα. Band densities were determined using GeneTools version 3 ( Syngene Laboratories, Frederick, MD). Each band in the P-AMPK blot (upper panel) was normalized relative to total AMPK (lower panel). These values were then standardized relative to the negative control lane, to yield the numbers shown below the lower panel.
While there was no obvious morphological abnormality in CD147 and CD98hc-depleted HEK293 cells, they did show markedly decreased proliferation in proportion to diminished CD147 expression (Fig. 7B). Serum starvation and mitomycin C treatment likewise impaired HEK293 cell proliferation, but without altering CD147 and CD98hc expression levels (not shown). Hence, diminished CD147 and CD98hc levels are a cause, rather than a consequence of diminished proliferation. While cell proliferation and CD147-CD98hc complex levels diminished in parallel, activation of AMP-activated protein kinase (AMPK) showed an inversely proportional increase (Fig. 7C). Activation of this cellular energy sensor, which is triggered by a high AMP/ATP ratio (29), is indicative of impaired energy metabolism.
Discussion
The CD147-CD98 complex
Striking similarities between the MCT-CD147 and CD98hc-LAT1 complexes had previously been noted. Both CD147 and CD98hc 1) are cell surface glycoproteins with a single transmembrane domain, 2) are highly expressed on activated or proliferating cells, 3) associate directly with transporters containing several transmembrane domains, and 4) are required to bring those transporters (MCT’s or LAT1 and related molecules) to the cell surface (27;9;10;18). Now our results reveal the existence of a novel MCT-CD147-CD98hc-LAT1 transporter complex, which also includes another amino acid transporter (ASCT2) as well as a protein (EpCAM) previously linked to epithelial cell proliferation. At the core of this complex is the interaction between CD147 and CD98hc. This interaction is direct (as seen using two different covalent crosslinking agents), was reciprocally demonstrated, was captured on the surface of multiple intact cell lines, and was shown to occur in cis rather than in trans.
The close association between CD147 and MCT proteins (10) is confirmed by our covalent crosslinking of CD147 and MCT1 (Fig. 2D). However, although we did see association of MCT with CD98hc under mild detergent conditions (1% Brij 97, CHAPS), we did not observe direct MCT-CD98hc crosslinking. CD98hc does associate directly, via a disulfide linkage, with LAT1 (31). However, although we did see association of LAT1 with CD147 under mild detergent conditions, we did not observe direct CD147-LAT1 crosslinking. Furthermore, a CD98hc-C109S mutant, which lacks LAT1 light chain association (32;22), retained CD147 association (not shown). These results support an MCT-CD147-CD98hc-LAT1 arrangement, in which the MCT-CD147 and CD98-LAT1 heterodimers are linked via direct CD147-CD98hc contact (Fig. 8).
Fig. 8.
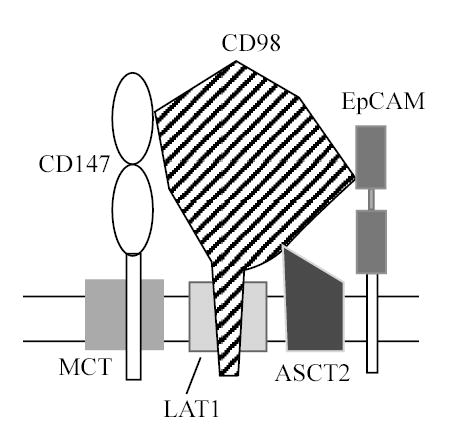
Model of the CD147-CD98hc super-complex. Results in Figs. 2 and 4 support a direct association between CD147 and CD98hc. Also, CD147 associates directly with MCT1 and MCT4 [(10) and Fig. 2D], CD98hc associates directly with LAT1 (31), and EpCAM associates directly with CD98hc (see text). Although Exp. I (in Table 1) suggested that LAT1 and EpCAM could also directly contact CD147, this possibility was not confirmed in further experiments. Experiments in Table 1 suggested that ASCT2 could contact both CD147 and CD98. Results from subsequent experiments currently favor a CD98hc contact, but an ASCT2-CD147 contact remains a possbility.
Functions of CD147-CD98hc complexes
RNAi-mediated depletion of either CD147 or CD98hc resulted in parallel diminution of both components, as well as activation of AMPK (AMP kinase) and decreased cell proliferation. We suggest that decreased cell proliferation is likely achieved through a regulatory pathway involving the mTOR (mammalian target of rapamycin) kinase (33;34). Without CD147, cellular metabolism of lactate and pyruvate should be impaired, since MCT1 and MCT4 do not insert into cell membranes properly (9;10). Without CD98hc, LAT1 family transporters are mostly trapped inside of cells, and amino acid transport should be impaired (27;35). Diminished intracellular levels of nutrients such as amino acids, glucose, pyruvate and lactate should lead to impaired energy metabolism, and a high AMP/ATP ratio, thereby triggering phosphorylation and activation of AMPK (AMP kinase) (36), which we did indeed observe. Decreased cellular nutrients and activated AMPK are both known to cause diminished activity of the serine/threonine kinase mTOR. This key regulatory “rheostat” in cells responds to environmental nutrient signals by regulating cell growth and proliferation, at least in part by affecting protein biosynthesis (37).
Why are expression levels of CD147 and CD98hc regulated in parallel? One possibility is that expression of one component may facilitate expression of the other, in the context of a MCT-CD147-CD98hc-LAT1 complex. For example, co-expression could boost the overall biosynthetic assembly of these complexes and/or could exert a stabilizing effect on each component once they reach the cell surface. Consistent with this “facilitated co-expression” idea, CD147 (1;3;4;5), and CD98hc (38;39;40;41) are each upregulated on tumor cell lines, and other types of metabolically activated cells. Another possibility is that CD147 and CD98hc proteins are co-regulated at the transcriptional or translational level. In the preceding paragraph, we emphasized how decreased levels of CD147 and CD98hc could cause changes in AMPK and mTOR pathways, leading to diminished proliferation. However, diminished expression of CD147 and CD98hc could also be a consequence of altered AMPK and mTOR pathways. In this regard, downregulation of mTOR activity can lead to decreased cell surface expression of CD98hc/4F2, and glucose transporter Glut1 (42), which is a molecule that can be co-regulated with MCT1 (43). Hence, RNAi-dependent depletion of either CD147 or CD98hc could lead to decreased mTOR phosphorylation, resulting in decreased expression of both molecules. This regulation must be somewhat specific, since levels of other highly expressed cell surface molecules (β1 integrins, MHC class I) were not affected by removal of either CD147 or CD98hc.
With the discovery of MCT-CD147-CD98hc-LAT1 complexes, we are now better able to understand a few observations that previously suggested a functional link between CD147 and CD98hc. In studies involving HIV gp160-mediated cell fusion and monocyte cell fusion, antibodies to CD98hc and CD147 had cross-regulatory effects (44). In studies of T lymphocyte costimulation by U937 cells (45) and dendritic cells (46), antibodies to CD147 and CD98hc were among the few antibodies to have inhibitory effects. In another study involving U937 cells, anti-CD147 antibodies inhibited homotypic aggregation that was stimulated by an anti-CD98 antibody (47). Our results might now also help to explain how CD98hc/4F2hc could unexpectedly facilitate transport of pyruvate (48), which is typically a substrate for MCT1 and MCT4.
Other molecules associated with CD147 and CD98hc
Although EpCAM had not previously been shown to associate with either CD147 or CD98hc, we observed such an association in four separate mass spectrometry experiments, and confirmed association with CD98hc in reciprocal co-immunoprecipitation and covalent crosslinking experiments. Like CD147 and CD98hc, EpCAM is also a cell surface glycoprotein that is highly expressed on most malignant epithelial cells. In this regard, EpCAM is being tested as a therapeutic target in anti-tumor clinical trials (28;29). Another molecule, ASCT2 (ALC1A5, AAAT, hATB0), was also found to associate with CD147 and CD98hc, as seen in five mass spectrometry experiments (Table 1). ASCT2 association with CD98hc was confirmed by co-immunoprecipitation and covalent crosslinking experiments. ASCT2 contains 8 transmembrane domains and acts as transporter for neutral amino acids (49). As seen for CD98hc (50;51), ASCT2 also may contribute to cancer progression (52) consistent with tumors having an increased need for amino acid transport. Both EpCAM and ASCT2 showed preferential association with CD98hc over CD147, and thus are placed in contact with CD98hc in Fig. 8. However, it is possible that EpCAM and ASCT2 could also contact CD147, as suggested by results in Table 1. Further experiments will be needed to resolve this issue.
Various β1 integrins have previously been shown to associate with CD147 (11) and with CD98 either directly (23) or indirectly (22). Indeed, we did obtain evidence for β1 integrin association, but in only 1 of 7 mass spectrometry experiments. Also, RNAi-mediated knockdown of CD147 and CD98hc did not decrease the levels of β1 integrins on the cell surface. Hence, integrins were not emphasized in this paper. Nonetheless, it is possible that independently identified integrin-EpCAM (53), integrin-CD147 (11), and integrin-CD98hc complexes (22;23) may be at least partially overlapping.
Association of CD147 with caveolin-1 has a negative effect on CD147 multimerization, glycosylation, and MMP induction. The smaller, less glycosylated form of CD147 preferentially associated with caveolin-1, with Ig domain 2 of CD147 being required (2;12). In contrast, the larger, highly glycosylated form of CD147 associated with CD98hc, with Ig domain 1 of CD147 being required. Hence, the CD147-CD98hc complex is distinct from complexes containing CD147 and caveolin-1. It is notable that the highly glycosylated form of CD147, which associates with CD98hc, is also involved in induction of MMP’s (54;55). However, a functional link between CD98hc and MMP induction has yet to be established.
The appearance of MCT1 and MCT4 in Table 1 was not unexpected, since both were previously shown to associate with CD147 (10). MCT8, a thyroid hormone transporter and another member of the MCT family, also appeared as a potential partner for CD147 in Table 1. This result still needs further biochemical confirmation. However, if an MCT8-CD147-CD98hc-LAT1 connection does truly occur, it would be highly relevant towards understanding of how thyroid hormone might be transported by both the CD98hc-LAT1 complex (56;57), and the CD147-MCT8 complex (58;59). Among the six known light chains that associate with CD98hc (35;18), only the neutral amino acid transporter LAT1 appeared in Table 1. It remains to be seen whether any of the other light chains might also appear in complexes with CD147.
Technical considerations
Comprehensive proteomics technologies have produced a remarkable compilation of networks of potential protein-protein interactions (15;16). However, within such networks, interactions among transmembrane proteins tend to be under-represented, since it is difficult to recapitulate hydrophobic interaction conditions on a massive scale in vitro. Furthermore, commonly used detergents such as Triton X-100 disrupt many functionally significant interactions. On the other hand, use of milder detergents may increase the recovery of functionally important complexes, but at the same time can yield an unacceptable level of non-specific, background interactions due to incomplete solubilization. To solve this problem, here we have treated intact cells with covalent crosslinkers, to capture existing interactions among transmembrane proteins. Thus we were able to use relatively stringent (1% Triton X-100) lysis and washing conditions, to decrease levels of background proteins, while retaining protein associations that otherwise would have been disrupted. In this way, we were able to discover several new interactions involving CD147 and CD98hc that had not been seen previously.
Antibodies to CD98hc and CD147 extracellular domains were of little use in isolating MCT-CD147-CD98hc-LAT1 complexes, presumably because extracellular epitopes on these molecules were blocked by associated proteins. In this regard, the anti-CD147 mAb 8G6 epitope maps to the same region of CD147 (Ig domain-1) needed for CD98hc association. Instead we relied on antibodies to intracellular, C-terminal FLAG- and GFP-tagged forms of CD147 and CD98hc. One disadvantage to this approach is that anti-FLAG detection is extremely non-linear, thus making estimation of stoichiometry very imprecise. Nonetheless, we did estimate (after correction for antibody detection efficiency) that at least 20–30% of CD147, and 20–30% of CD98 may be complexed with each other. In the case of EpCAM association, the epitope on CD98hc recognized by mAb 4F2 was not blocked, and we were able to recover ~30% of EpCAM in a complex with CD98hc.
Does our technical approach lead to the identification of proteins, captured by covalent crosslinking, simply because they are very abundant on the cell surface? Indeed, we suspect that some highly abundant proteins listed at the bottom of Table 1 (e.g. transferrin receptor/CD71, CD44, HLA class I) might appear because they were trapped among the others. Consistent with this interpretation, subsequent co-immunoprecipitation experiments did not support specific interactions with either CD147 or CD98hc. In contrast, interactions involving proteins listed at the top of Table I were confirmed in separate biochemical experiments. Furthermore, to minimize concerns regarding high expression levels, ectopic CD147 was expressed at levels only 1.4–2.1-fold above endogenous CD147. Also, many experiments utilized endogenous CD98hc, CD147, EpCAM, ASCT2, LAT1 and MCT proteins rather than ectopically over-expressed proteins. As further evidence of specificity, we have carried out several additional mass spectrometry experiments, aimed at isolating protein partners for other abundant cell surface proteins (laminin-binding integrins, EWI-2 protein, tetraspanins). The dozens of potential protein partners identified in those experiments did not include CD147, MCT proteins, CD98hc, amino acid transporters, or EpCAM (not shown).
In conclusion, we have combined covalent crosslinking and mass spectrometry to discover novel physical associations among molecules involved in cell proliferation and transport. Functional co-regulation of cell surface CD98hc-LAT1 and CD147-MCT protein complexes is consistent with their physical association, and demonstrates their joint role in cellular energy metabolism. The presence of additional transporter (ASCT2) and proliferation-related (EpCAM) components suggests the existence of a transmembrane protein super-complex, which provides a novel physical framework for understanding functional connections among these diverse molecules.
Supplemental Table S1.
Peptide sequences obtained by LC-MS/MS
| CD147: | |
| Exp. I (7) | Exp. II (10) |
| FFVSSSQGR | WLKGGVVLK |
| GSDQAIITLR | FFVSSSQGR |
| GGVVLKEDALPGQK | GSDQAIITLR |
| SESVPPVTDWAWYK | GGVVLKEDALPGQK |
| KPEDVLDDDDAGSAPLK | SESVPPVTDWAWYK |
| RKPEDVLDDDDAGSSAPLK | KPEDVLDDDDAGSAPLK |
| SELHIENLNMEADPGQYR | RKPEDVLDDDDAGSSAPLK |
| RRKPEDVLDDDDAGSSAPLK | |
| SELHIENLNMEADPGQYR | |
| RKPEDVLDDDDAGSSAPLKSSGQHQNDK | |
| Exp. III (8) | Exp. IV (8) |
| FFVSSSQGR | FFVSSSQGR |
| GSDQAIITLR | GSDQAIITLR |
| GGVVLKEDALPGQK | GGVVLKEDALPGQK |
| SESVPPVTDWAWYK | SESVPPVTDWAWYK |
| KPEDVLDDDDAGSAPLK | SSEHINEGETAMLLVCK |
| RKPEDVLDDDDAGSSAPLK | KPEDVLDDDDAGSAPLK |
| RRKPEDVLDDDDAGSSAPLK | RKPEDVLDDDDAGSAPLK |
| SELHIENLNMEADPGQYR | SELHIENLNMEADPGQYR |
| CD98hc: | |
| Exp. 1 (13) | Exp. II (6) |
| EEGSPLELER | EEGSPLELER |
| VILDLTPNYR | ADLLLSTQPGR |
| EDFDSLLQSAK | EDFDSLLQSAK |
| FTGLSKEELLK | LKLEPHEGLLLR |
| LLTSFLPAQLLR | IKVAEDEAEAAAAAK |
| IKVAEDEAEAAAAAK | DDVAQTDLLQIDPNFGSK |
| GQSEDPGSLLSLFR | |
| GENSWFSTQVDTVATK | Exp. III |
| IGDLQAFQGHGAGNLAGLK | GQSEDPGSLLSLFR |
| DDVAQTDLLQIDPNFGSK | |
| SLLHGDFHAFSAGPGLFSYIR | |
| NQKDDVAQTDLLQIDPNFGSK | |
| NQKDDVAQTDLLQIDPNFGSKEDFDSLLQSAK | |
| Exp. IV (6) | Exp. V (23) |
| VILDLTPNYR | GLVLGPIHK |
| LLTSFLPAQLLR | LDYLSSLK |
| GQSEDPGSLLSLFR | GRLDYLSSLK |
| DDVAQTDLLQIDPNFGSK | EEGSPLELER |
| DLLLTSSYLSDSGSTGEHTK | ADLLLSTQPGR |
| NQKDDVAQTDLLQIDPNFGSK | LEPHEGLLLR |
| WWHTGALYR | |
| VILDLTPNYR | |
| WCSWSLSQAR | |
| VAEDEAEAAAAAK | |
| EDFDSLLQSAK | |
| EVELNELEPEK | |
| LLTSFLPAQLLR | |
| LKLEPHEGLLLR | |
| IKVAEDEAEAAAAAK | |
| GQSEDPGSLLSLFR | |
| GENSWFSTQVDTVATK | |
| IGDLQAFQGHGAGNLAGLK | |
| DDVAQTDLLQIDPNFGSK | |
| DLLLTSSYLSDSGSTGEHTK | |
| ADLLLSTQPGREEGSPLELER | |
| NQKDDVAQTDLLQIDPNFGSK | |
| EVELNELEPEKQPMNAASGAAMSLAGAEK | |
| Exp. VI (14) | Exp VII (20) |
| LDYLSSLK | VAGSPGWVR |
| EEGSPLELER | GLVLGPIHK |
| ADLLLSTQPGR | LDYLSSLK |
| LEPHEGLLLR | GRLDYLSSLK |
| WWHTGALYR | EEGSPLELER |
| VILDLTPNYR | ADLLLSTQPGR |
| EDFDSLLQSAK | WWHTGALYR |
| EVELNELEPEK | VILDLTPNYR |
| LLTSFLPAQLLR | EDFDSLLQSAK |
| LKLEPHEGLLLR | LLTSFLPAQLLR |
| GQSEDPGSLLSLFR | LKLEPHEGLLLR |
| GENSWFSTQVDTVATK | IKVAEDEAEAAAAAK |
| DLLLTSSYLSDSGSTGEHTK | GQSEDPGSLLSLFR |
| ADLLLSTQPGREEGSPLELER | GENSWFSTQVDTVATK |
| IGDLQAFQGHGAGNLAGLK | |
| DDVAQTDLLQIDPNFGSK | |
| DLLLTSSYLSDSGSTGEHTK | |
| SLLHGDFHAFSAGPGLFSYIR | |
| NQKDDVAQTDLLQIDPNFGSK | |
| EVELNELEPEKQPMNAASGAAMSLAGAEK | |
| ASCT2: | |
| Exp. I (5) | Exp. II (3) |
| LGPEGELLIR | STEPELIQVK |
| STEPELIQVK | EVLDSFLDLAR |
| EVLDSFLDLAR | SELPLDPLPVPTEEGNPLLK |
| NIFPSNLVSAAFR | |
| SELPLDPLPVPTEEGNPLLK | |
| Exp IV (4) | Exp. V |
| EVLDSFLDLAR | SELPLDPLPVPTEEGNPLLK |
| NIFPSNLVSAAFR | |
| IVEMEDVGLLFAR | Exp. VII |
| SELPLDPLPVPTEEGNPLLK | SELPLDPLPVPTEEGNPLLK |
| LAT1: | |
| Exp. I (3) | Exp. VI (3) |
| ALAAPAAEEKEEAP | SADGSAPAGEGEGVTLQR |
| SADGSAPAGEGEGVTLQR | GDVSNLDPNFSFEGTK |
| GDVSNLDPNFSFEGTK | VQDAAAAKLLALALIILLGFVQIGK |
| Exp. VII (3) | |
| ALAAPAAEEKEEAP | |
| SADGSAPAGEGEGVTLQR | |
| GDVSNLDPNFSFEGTK | |
| EpCAM: | |
| Exp. I TQNDVDIADVAYYFEK | |
| Exp. V TQNDVDIADVAYYFEK | |
| Exp. VI TQNDVDIADVAYYFEK | |
| Exp. VII TQNDVDIADVAYYFEK | |
| MCT1: | |
| Exp. I | Exp.II (3) |
| DLHDANTDLIGR | DLHDANTDLIGR |
| DLHDANTDLIGRHPK | |
| Exp. III (2) | EEETSIDVAGKPNEVTK |
| DLHDANTDLIGR | |
| EEETSIDVAGKPNEVTK | |
| MCT4: | |
| Exp. I | Exp. II (4) |
| LHKPPADSGVDLR | EVEHFLK |
| NGEVVHTPETSV | |
| Exp. III | LHKPPADSGVDLR |
| LHKPPADSGVDLR | AEPEKNGEVVHTPETSV |
| MCT 8: | |
| Exp. I VHEPEPTPTVETR | |
| CD29: | |
| Exp. II | |
| IGFGSFVEK | |
| LRSGEPQTFTLK | |
| LSENNIQTIFAVTEEFQPVYK | |
| Na/K ATPase, α 1: | |
| Exp. I (2) | Exp. II (5) |
| QGAIVAVTGDGVNDSPALK | ADIGVAMGIAGSDVSK |
| GVGIISEGNETVEDIAAR | RAVAGDASESALLK |
| AVFQANQENLPILK | |
| Exp. IV (2) | LNIPVSQVNPR |
| LNIPVSQVNPR | SPDFTNENPLETR |
| AVFQANQENLPILK | |
| Exp. VI | Exp. VII |
| GVGIISEGNETVEDIAAR | GVGIISEGNETVEDIAAR |
| Na/K ATPase, β3: | |
| Exp. V LFIYNPTTGEFLGR | |
| Glycopr. Trop-2: | |
| Exp. I AAGDVDIGDAAYYFER | |
| Exp. VII AAGDVDIGDAAYYFER | |
| I3 Bp: | |
| Exp. I TVNTFSQSVSSLFGEDNVR | |
| CD71: | |
| Exp. I GFVEPDHYVVVGAQR | |
|
Exp. II (2) SSGLPNIPVQTISR
VEYHFLSPYVSPK |
|
|
Exp. IV (2) VSASPLLYTLIEK
GFVEPDHYVVVGAQR |
|
| Sel-1 homolog: | |
| Exp. I (2) | Exp. IV (2) |
| LVANHVASDISLTGGSVVQR | LVANHVASDISLTGGSVVQR |
| VSYALLFGDYLPQNIQAAR | VSYALLFGDYLPQNIQAAR |
| CD166: | |
| Exp. II (4) | |
| ALFLETEQLK | |
| SSNTYTLMDVR | |
| ALFLETEQLKK | |
| SVQYDDVPEYK | |
| CD49e: | |
| Exp. II LLESSLSSSEGEEPVEYK | |
| CD44: | |
| Exp. II SQEMVHLVNK | |
| Leuk. virus recept.1: | |
| Exp. II LSVGDIENKHPVSEVGPATVPLQAVVEER | |
| HLA-I,A-25: | |
| Exp. II APWIEQEGPEYWDR | |
Footnotes
This work was supported by a grant (CA102034) from the National Institutes of Health. We thank Dr. Tatiana Kolesnikova for antibodies and cDNA’s for wild type and mutant CD98. We thank Dr. Wei Tang for CD147 cDNA and antibodies.
References
- 1.Toole BP. Emmprin (CD147), a cell surface regulator of matrix metalloproteinase production and function. Curr Top Dev Biol. 2003;54:371–389. doi: 10.1016/s0070-2153(03)54015-7. [DOI] [PubMed] [Google Scholar]
- 2.Tang W, Chang SB, Hemler ME. Links between CD147 Function, Glycosylation, and Caveolin-1. Mol Biol Cell. 2004;15:4043–4050. doi: 10.1091/mbc.E04-05-0402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Koch C, Staffler G, Huttinger R, Hilgert I, Prager E, Cerny J, Steinlein P, Majdic O, Horejsi V, Stockinger H. T cell activation-associated epitopes of CD147 in regulation of the T cell response, and their definition by antibody affinity and antigen density. Int Immunol. 1999;11:777–786. doi: 10.1093/intimm/11.5.777. [DOI] [PubMed] [Google Scholar]
- 4.Betsuyaku T, Tanino M, Nagai K, Nasuhara Y, Nishimura M, Senior RM. Extracellular matrix metalloproteinase inducer is increased in smokers' bronchoalveolar lavage fluid. Am J Respir Crit Care Med. 2003;168:222–227. doi: 10.1164/rccm.200301-103OC. [DOI] [PubMed] [Google Scholar]
- 5.Nehme CL, Fayos BE, Bartles JR. Distribution of the integral plasma membrane glycoprotein CE9 (MRC OX-47) among rat tissues and its induction by diverse stimuli of metabolic activation. Biochem J. 1995;310 ( Pt 2):693–698. doi: 10.1042/bj3100693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Marieb EA, Zoltan-Jones A, Li R, Misra S, Ghatak S, Cao J, Zucker S, Toole BP. Emmprin promotes anchorage-independent growth in human mammary carcinoma cells by stimulating hyaluronan production. Cancer Res. 2004;64:1229–1232. doi: 10.1158/0008-5472.can-03-2832. [DOI] [PubMed] [Google Scholar]
- 7.Pushkarsky T, Zybarth G, Dubrovsky L, Yurchenko V, Tang H, Guo H, Toole B, Sherry B, Bukrinsky M. CD147 facilitates HIV-1 infection by interacting with virus-associated cyclophilin A. Proc Natl Acad Sci USA. 2001;98:6360–6365. doi: 10.1073/pnas.111583198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dumitrescu AM, Liao XH, Best TB, Brockmann K, Refetoff S. A novel syndrome combining thyroid and neurological abnormalities is associated with mutations in a monocarboxylate transporter gene. Am J Hum Genet. 2004;74:168–175. doi: 10.1086/380999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kirk P, Wilson MC, Heddle C, Brown MH, Barclay AN, Halestrap AP. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. EMBO J. 2000;19:3896–3904. doi: 10.1093/emboj/19.15.3896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Halestrap AP, Meredith D. The SLC16 gene family-from monocarboxylate transporters (MCTs) to aromatic amino acid transporters and beyond. Pflugers Arch. 2004;447:619–628. doi: 10.1007/s00424-003-1067-2. [DOI] [PubMed] [Google Scholar]
- 11.Berditchevski F, Chang S, Bodorova J, Hemler ME. Generation of monoclonal antibodies to integrin-associated proteins: Evidence that α3β1 complexes with EMMPRIN/basigin/OX47/M6. J Biol Chem. 1997;272:29174–29180. doi: 10.1074/jbc.272.46.29174. [DOI] [PubMed] [Google Scholar]
- 12.Tang W, Hemler ME. Caveolin-1 regulates matrix metalloproteinases-1 induction and CD147/EMMPRIN cell surface clustering. J Biol Chem. 2004;279:11112–11118. doi: 10.1074/jbc.M312947200. [DOI] [PubMed] [Google Scholar]
- 13.Naruhashi K, Kadomatsu K, Igakura T, Fan QW, Kuno N, Muramatsu H, Miyauchi T, Hasegawa T, Itoh A, Muramatsu T, Nabeshima T. Abnormalities of sensory and memory functions in mice lacking Bsg gene. Biochem Biophys Res Commun. 1997;236:733–737. doi: 10.1006/bbrc.1997.6993. [DOI] [PubMed] [Google Scholar]
- 14.Chen S, Kadomatsu K, Kondo M, Toyama Y, Toshimori K, Ueno S, Miyake Y, Muramatsu T. Effects of flanking genes on the phenotypes of mice deficient in basigin/CD147. Biochem Biophys Res Commun. 2004;324:147–153. doi: 10.1016/j.bbrc.2004.08.232. [DOI] [PubMed] [Google Scholar]
- 15.Giot L, Bader JS, Brouwer C, Chaudhuri A, Kuang B, Li Y, Hao YL, Ooi CE, Godwin B, Vitols E, Vijayadamodar G, Pochart P, Machineni H, Welsh M, Kong Y, Zerhusen B, Malcolm R, Varrone Z, Collis A, Minto M, Burgess S, McDaniel L, Stimpson E, Spriggs F, Williams J, Neurath K, Ioime N, Agee M, Voss E, Furtak K, Renzulli R, Aanensen N, Carrolla S, Bickelhaupt E, Lazovatsky Y, DaSilva A, Zhong J, Stanyon CA, Finley RL, Jr, White KP, Braverman M, Jarvie T, Gold S, Leach M, Knight J, Shimkets RA, McKenna MP, Chant J, Rothberg JM. A protein interaction map of Drosophila melanogaster. Science. 2003;302:1727–1736. doi: 10.1126/science.1090289. [DOI] [PubMed] [Google Scholar]
- 16.Li S, Armstrong CM, Bertin N, Ge H, Milstein S, Boxem M, Vidalain PO, Han JD, Chesneau A, Hao T, Goldberg DS, Li N, Martinez M, Rual JF, Lamesch P, Xu L, Tewari M, Wong SL, Zhang LV, Berriz GF, Jacotot L, Vaglio P, Reboul J, Hirozane-Kishikawa T, Li Q, Gabel HW, Elewa A, Baumgartner B, Rose DJ, Yu H, Bosak S, Sequerra R, Fraser A, Mango SE, Saxton WM, Strome S, Van Den HS, Piano F, Vandenhaute J, Sardet C, Gerstein M, Doucette-Stamm L, Gunsalus KC, Harper JW, Cusick ME, Roth FP, Hill DE, Vidal M. A map of the interactome network of the metazoan C. elegans. Science. 2004;303:540–543. doi: 10.1126/science.1091403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Deves R, Boyd CA. Surface antigen CD98(4F2): not a single membrane protein, but a family of proteins with multiple functions. J Membr Biol. 2000;173:165–177. doi: 10.1007/s002320001017. [DOI] [PubMed] [Google Scholar]
- 18.Verrey F, Closs EI, Wagner CA, Palacin M, Endou H, Kanai Y. CATs and HATs: the SLC7 family of amino acid transporters. Pflugers Arch. 2004;447:532–542. doi: 10.1007/s00424-003-1086-z. [DOI] [PubMed] [Google Scholar]
- 19.Tsurudome M, Ito Y. Function of fusion regulatory proteins (FRPs) in immune cells and virus-infected cells. Crit Rev Immunol. 2000;20:167–196. [PubMed] [Google Scholar]
- 20.Miyamoto YJ, Mitchell JS, McIntyre BW. Physical association and functional interaction between beta1 integrin and CD98 on human T lymphocytes. Mol Immunol. 2003;39:739–751. doi: 10.1016/s0161-5890(02)00255-9. [DOI] [PubMed] [Google Scholar]
- 21.Fenczik CA, Zent R, Dellos M, Calderwood DA, Satriano J, Kelly C, Ginsberg MH. Distinct domains of CD98hc regulate integrins and amino acid transport. J Biol Chem. 2001;276:8746–8752. doi: 10.1074/jbc.M011239200. [DOI] [PubMed] [Google Scholar]
- 22.Kolesnikova TV, Mannion BA, Berditchevski F, Hemler ME. Beta1 integrins show specific association with CD98 protein in low density membranes. BMC Biochem. 2001;2:10. doi: 10.1186/1471-2091-2-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zent R, Fenczik CA, Calderwood DA, Liu S, Dellos M, Ginsberg MH. Class- and splice variant-specific association of CD98 with integrin beta cytoplasmic domains. J Biol Chem. 2000;275:5059–5064. doi: 10.1074/jbc.275.7.5059. [DOI] [PubMed] [Google Scholar]
- 24.Hemler ME, Strominger JL. Characterization of antigen recognized by the monoclonal antibody (4F2): different molecular forms on human T and B lymphoblastoid cell lines. J Immunol. 1982;129:623–628. [PubMed] [Google Scholar]
- 25.Shevchenko A, Wilm M, Vorm O, Mann M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal Chem. 1996;68:850–858. doi: 10.1021/ac950914h. [DOI] [PubMed] [Google Scholar]
- 26.Peng J, Gygi SP. Proteomics: the move to mixtures. J Mass Spectrom. 2001;36:1083–1091. doi: 10.1002/jms.229. [DOI] [PubMed] [Google Scholar]
- 27.Mastroberardino L, Spindler B, Pfeiffer R, Skelly PJ, Loffing J, Shoemaker CB, Verrey F. Amino-acid transport by heterodimers of 4F2hc/CD98 and members of a permease family. Nature. 1998;395:288–291. doi: 10.1038/26246. [DOI] [PubMed] [Google Scholar]
- 28.Armstrong A, Eck SL. EpCAM: A new therapeutic target for an old cancer antigen. Cancer Biol Ther. 2003;2:320–326. doi: 10.4161/cbt.2.4.451. [DOI] [PubMed] [Google Scholar]
- 29.Di Paolo C, Willuda J, Kubetzko S, Lauffer I, Tschudi D, Waibel R, Pluckthun A, Stahel RA, Zangemeister-Wittke U. A recombinant immunotoxin derived from a humanized epithelial cell adhesion molecule-specific single-chain antibody fragment has potent and selective antitumor activity. Clin Cancer Res. 2003;9:2837–2848. [PubMed] [Google Scholar]
- 30.Utsunomiya-Tate N, Endou H, Kanai Y. Cloning and functional characterization of a system ASC-like Na+-dependent neutral amino acid transporter. J Biol Chem. 1996;271:14883–14890. doi: 10.1074/jbc.271.25.14883. [DOI] [PubMed] [Google Scholar]
- 31.Verrey F, Meier C, Rossier G, Kuhn LC. Glycoprotein-associated amino acid exchangers: broadening the range of transport specificity. Pflugers Arch. 2000;440:503–512. doi: 10.1007/s004240000274. [DOI] [PubMed] [Google Scholar]
- 32.Pfeiffer R, Spindler B, Loffing J, Skelly PJ, Shoemaker CB, Verrey F. Functional heterodimeric amino acid transporters lacking cysteine residues involved in disulfide bond. FEBS Lett. 1998;439:157–162. doi: 10.1016/s0014-5793(98)01359-3. [DOI] [PubMed] [Google Scholar]
- 33.Dennis PB, Jaeschke A, Saitoh M, Fowler B, Kozma SC, Thomas G. Mammalian TOR: a homeostatic ATP sensor. Science. 2001;294:1102–1105. doi: 10.1126/science.1063518. [DOI] [PubMed] [Google Scholar]
- 34.Rohde J, Heitman J, Cardenas ME. The TOR kinases link nutrient sensing to cell growth. J Biol Chem. 2001;276:9583–9586. doi: 10.1074/jbc.R000034200. [DOI] [PubMed] [Google Scholar]
- 35.Verrey F. System L: heteromeric exchangers of large, neutral amino acids involved in directional transport. Pflugers Arch. 2003;445:529–533. doi: 10.1007/s00424-002-0973-z. [DOI] [PubMed] [Google Scholar]
- 36.Carling D. The AMP-activated protein kinase cascade--a unifying system for energy control. Trends BiochemSci. 2004;29:18–24. doi: 10.1016/j.tibs.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 37.Fingar DC, Blenis J. Target of rapamycin (TOR): an integrator of nutrient and growth factor signals and coordinator of cell growth and cell cycle progression. Oncogene. 2004;23:3151–3171. doi: 10.1038/sj.onc.1207542. [DOI] [PubMed] [Google Scholar]
- 38.Cotner T, Williams JM, Christenson L, Shapiro HM, Strom TB, Strominger JL. Simultaneous flow cytometric analysis of human T cell activation antigen expression and DNA content. J Exp Med. 1983;157:461–472. doi: 10.1084/jem.157.2.461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yoon JH, Kim YB, Kanai Y, Endou H, Kim dK. Sequential increases in 4F2hc expression during DMBA-induced hamster buccal pouch carcinogenesis. Anticancer Res. 2003;23:3877–3881. [PubMed] [Google Scholar]
- 40.Nawashiro H, Otani N, Shinomiya N, Fukui S, Nomura N, Yano A, Shima K, Matsuo H, Kanai Y. The role of CD98 in astrocytic neoplasms. Hum Cell. 2002;15:25–31. doi: 10.1111/j.1749-0774.2002.tb00096.x. [DOI] [PubMed] [Google Scholar]
- 41.Diaz LA, Jr, Fox DA. A role for CD98 in cellular activation. J Biol Regul Homeost Agents. 1998;12:25–32. [PubMed] [Google Scholar]
- 42.Edinger AL, Thompson CB. Akt maintains cell size and survival by increasing mTOR-dependent nutrient uptake. Mol Biol Cell. 2002;13:2276–2288. doi: 10.1091/mbc.01-12-0584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Maurer MH, Canis M, Kuschinsky W, Duelli R. Correlation between local monocarboxylate transporter 1 (MCT1) and glucose transporter 1 (GLUT1) densities in the adult rat brain. Neurosci Lett. 2004;355:105–108. doi: 10.1016/j.neulet.2003.10.056. [DOI] [PubMed] [Google Scholar]
- 44.Mori, K., Nishimura, M., Tsurudome, M., Ito, M., Nishio, M., Kawano, M., Kozuka, Y., Yamashita, Y., Komada, H., Uchida, A., and Ito, Y. (2003) The functional interaction between CD98 and CD147 in regulation of virus-induced cell fusion and osteoclast formation. Med.Microbiol.Immunol.(Berl) [DOI] [PubMed]
- 45.Stonehouse TJ, Woodhead VE, Herridge PS, Ashrafian H, George M, Chain BM, Katz DR. Molecular characterization of U937-dependent T-cell co-stimulation. Immunology. 1999;96:35–47. doi: 10.1046/j.1365-2567.1999.00670.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Woodhead VE, Stonehouse TJ, Binks MH, Speidel K, Fox DA, Gaya A, Hardie D, Henniker AJ, Horejsi V, Sagawa K, Skubitz KM, Taskov H, Todd RF, III, van Agthoven A, Katz DR, Chain BM. Novel molecular mechanisms of dendritic cell-induced T cell activation. IntImmunol. 2000;12:1051–1061. doi: 10.1093/intimm/12.7.1051. [DOI] [PubMed] [Google Scholar]
- 47.Cho JY, Fox DA, Horejsi V, Sagawa K, Skubitz KM, Katz DR, Chain B. The functional interactions between CD98, beta1-integrins, and CD147 in the induction of U937 homotypic aggregation. Blood. 2001;98:374–382. doi: 10.1182/blood.v98.2.374. [DOI] [PubMed] [Google Scholar]
- 48.Yao SY, Muzyka WR, Cass CE, Cheeseman CI, Young JD. Evidence that the transport-related proteins BAT and 4F2hc are not specific for amino acids: induction of Na+-dependent uridine and pyruvate transport activity by recombinant BAT and 4F2hc expressed in Xenopus oocytes. Biochem Cell Biol. 1998;76:859–865. doi: 10.1139/bcb-76-5-859. [DOI] [PubMed] [Google Scholar]
- 49.Kanai Y, Hediger MA. The glutamate/neutral amino acid transporter family SLC1: molecular, physiological and pharmacological aspects. Pflugers Arch. 2004;447:469–479. doi: 10.1007/s00424-003-1146-4. [DOI] [PubMed] [Google Scholar]
- 50.Hara K, Kudoh H, Enomoto T, Hashimoto Y, Masuko T. Malignant transformation of NIH3T3 cells by overexpression of early lymphocyte activation antigen CD98. Biochem Biophys Res Commun. 1999;262:720–725. doi: 10.1006/bbrc.1999.1051. [DOI] [PubMed] [Google Scholar]
- 51.Hara K, Kudoh H, Enomoto T, Hashimoto Y, Masuko T. Enhanced tumorigenicity caused by truncation of the extracellular domain of GP125/CD98 heavy chain. Oncogene. 2000;19:6209–6215. doi: 10.1038/sj.onc.1204019. [DOI] [PubMed] [Google Scholar]
- 52.Li R, Younes M, Frolov A, Wheeler TM, Scardino P, Ohori M, Ayala G. Expression of neutral amino acid transporter ASCT2 in human prostate. Anticancer Res. 2003;23:3413–3418. [PubMed] [Google Scholar]
- 53.Schmidt DS, Klingbeil P, Schnolzer M, Zoller M. CD44 variant isoforms associate with tetraspanins and EpCAM. Exp Cell Res. 2004;297:329–347. doi: 10.1016/j.yexcr.2004.02.023. [DOI] [PubMed] [Google Scholar]
- 54.Guo H, Zucker S, Gordon MK, Toole BP, Biswas C. Stimulation of matrix metalloproteinase production by recombinant extracellular matrix metalloproteinase inducer from transfected Chinese hamster ovary cells. J Biol Chem. 1997;272:24–27. [PubMed] [Google Scholar]
- 55.Sun J, Hemler ME. Regulation of MMP production through homophilic CD147/EMMPRIN interaction. Cancer Res. 2001;61:2276–2281. [PubMed] [Google Scholar]
- 56.Friesema EC, Docter R, Moerings EP, Verrey F, Krenning EP, Hennemann G, Visser TJ. Thyroid hormone transport by the heterodimeric human system L amino acid transporter. Endocrinol. 2001;142:4339–4348. doi: 10.1210/endo.142.10.8418. [DOI] [PubMed] [Google Scholar]
- 57.Ritchie JW, Taylor PM. Role of the System L permease LAT1 in amino acid and iodothyronine transport in placenta. Biochem J. 2001;356:719–725. doi: 10.1042/0264-6021:3560719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Friesema EC, Ganguly S, Abdalla A, Manning Fox JE, Halestrap AP, Visser TJ. Identification of monocarboxylate transporter 8 as a specific thyroid hormone transporter. J Biol Chem. 2003;278:40128–40135. doi: 10.1074/jbc.M300909200. [DOI] [PubMed] [Google Scholar]
- 59.Friesema EC, Grueters A, Biebermann H, Krude H, von Moers A, Reeser M, Barrett TG, Mancilla EE, Svensson J, Kester MH, Kuiper GG, Balkassmi S, Uitterlinden AG, Koehrle J, Rodien P, Halestrap AP, Visser TJ. Association between mutations in a thyroid hormone transporter and severe X-linked psychomotor retardation. Lancet. 2004;364:1435–1437. doi: 10.1016/S0140-6736(04)17226-7. [DOI] [PubMed] [Google Scholar]