

Abstract
Most words in natural language are polysemous; i.e., they can be used in more than one way. For example, paper can be used to refer to a substance made out of wood pulp or to a daily publication printed on that substance. Even though virtually every sentence contains polysemy, there is little agreement as to how polysemy is represented in the mental lexicon. Do different uses of polysemous words involve access to a single representation or do our minds store distinct representations for each different sense? Here we investigated priming between senses with a combination of behavioral and magnetoencephalographic (MEG) measures in order to test whether different senses of the same word involve identity or mere formal and semantic similarity. Our results show that polysemy effects are clearly distinct from similarity effects bilaterally. In the left hemisphere, sense-relatedness elicited shorter latencies of the M350 source, which has been hypothesized to index lexical activation. Concurrent activity in the right hemisphere, on the other hand, peaked later for sense-related than for unrelated target stimuli, suggesting competition between related senses. The obtained pattern of results supports models in which the representation of polysemy involves both representational identity and difference: Related senses connect to same abstract lexical representation, but are distinctly listed within that representation.
INTRODUCTION
How the human parser deals with the ambiguity of natural language is a key problem in the investigation of the cognitive architecture of language processing. For lexical items, two different types of ambiguity are traditionally distinguished. In homonymy, a single phonological word form has two or more semantically unrelated meanings; for example, the form bank can be used to refer to the edge of a river (as in river bank) or a financial institution (as in savings bank). In polysemy, on the other hand, a single word form is associated with two or more meanings, traditionally called “senses,” that are distinct but semantically related. For instance, the word paper can be used to refer to a material, as in shredded paper, or to the content of a publication, as in Today's paper was engaging. Notice that, in the age of the internet, reading the paper does not necessarily involve any paper (the material). Different senses of a word can be rather different from one another, as when the word paper is used to refer to a newspaper company (The paper fired its editors), which has little in common with writing material. Nevertheless, language users perceive connections between the different uses of paper, which in fact are historically related.
Although homonymy has been studied in countless psychological experiments (see Simpson, 1995, for a review), polysemy has been much less studied, even though it is far more frequent. Indeed, most content words are polysemous to some degree, and the more frequent a word is, the more polysemous it tends to be (Zipf, 1935). One can verify this by looking in a dictionary and noticing that the longest entries tend to be for very frequent words. Caramazza and Grober (1976) identified 26 distinct senses for the word line and 40 senses for run. Thus, speakers and listeners must solve the problem of polysemous ambiguity in almost every sentence they utter and hear.
The question we address here is how the different but related senses in polysemy are psychologically represented in the mental lexicon (dictionary). Although most models agree that homonyms are represented as separate words, i.e., are listed as their own entries in the mental lexicon, the representation of polysemy has been highly controversial. Is polysemy just homonymy, or a qualitatively different phenomenon? There are no widely accepted criteria for distinguishing between these two types of ambiguity (Geeraerts, 1993). In this work we used magnetoencephalography (MEG) to tease apart predictions of different representational hypotheses about polysemy. In what follows, we first summarize behavioral evidence pertaining to the representation of polysemy and then describe contrasting neurophysiological predictions of different hypotheses.
Behavioral Evidence for Sense Representations
Many researchers hold that polysemous senses are not stored as different words, for a number of reasons (Cruse, 1986; Fellbaum, 2000). First, the senses are clearly related to one another and are sometimes extremely similar (e.g., paper as a kind of material and as that material formed into sheets used for writing). Second, the senses are historically derived from other senses, suggesting a close relationship that is absent in homonyms. Third, polysemy often forms patterns across the lexicon, as explained below, which is not true of homonyms.
Indeed, one proposal is that polysemous words are represented as a single, core meaning. The different senses are not explicitly represented in the lexicon at all, but are derived by speakers through lexical rules (e.g., Caramazza & Grober, 1976; Nunberg, 1979). For example, the word paper might be represented as a single sense of sheets of writing material. When hearing the word, a listener retrieves that sense, and, if it does not fit the present context, attempts to derive the intended meaning. This is possible because some polysemous senses follow lexical patterns or rules (Lehrer, 1990). For example, the use of paper to refer to a physical object like a newspaper as well as the content of a newspaper (That paper is dull) is also found in book, CD, video, film, and other words referring to objects that have content. Another rule is that a word referring to an animal can generally also refer to the meat of that animal (I saw a salmon. I ate some salmon.). Such rules might permit the lexicon to store only one sense, with other senses derived from it.
However, this core meaning proposal has some serious problems, including the fact that many of the senses of polysemous words do not follow these patterns (e.g., paper used to refer to a newspaper company) and the fact that the different senses are sometimes extremely different in meaning. Klein and Murphy (2002) demonstrated this empirically, by showing that speakers did not perceive the different senses of polysemous words to refer to the same kinds of things. Furthermore, when people process a polysemous word in one sense, this interferes with their later processing of the word in a different sense (Klein & Murphy, 2001). Surprisingly, the size of this interference effect is very similar to that found for the two meanings of a homonym. This clearly challenges the hypothesis that all senses derive from the same core meaning. Finally, some researchers have argued that polysemous senses are unpredictable in detail, and so they must be explicitly represented in the lexicon, because they cannot be fully derived by a linguistic rule (Cruse, 1986; Lehrer, 1990; Rice, 1992).
In order to account for inhibitory effects between distinct senses of the same word (henceforth polysemes), one needs to assume either that sense representations exist or that sense-deriving rules can inhibit each other. The former hypothesis is significantly more straightforward, since similarity-based inhibition between lexical representations is a well-known phenomenon (Allen & Badecker, 1999; Grainger, Cole, & Segui, 1991; Laudana, Badecker, & Caramazza, 1989; Marslen-Wilson, 1987; Soto-Faraco, Sebastián-Gallés, & Cutler, 2001). In contrast, so far as we know, there is no evidence for inhibition between rules. However, while the behavioral data summarized above suggest that senses are represented in the lexicon, they leave open the question of what these representations are like. Two possibilities suggest themselves. First, the senses could be represented as separate lexical items, just as homonyms are. So, one entry could be paper1: sheets of writing material, and another could be paper2: substance consisting of wood pulp, while another is paper3: the content of an article, and so on. All these entries would have the same pronunciation. Alternatively, the senses could all be listed as part of the same lexical item. This is how most dictionaries handle the problem, giving homonyms like bank two entries, but placing the different senses of paper or chicken under the same entry. On this theory, then, bank is really two different words, but paper is a single word with a number of distinct but related meanings.
The choice between these two alternatives has important consequences for the structure of the mental lexicon and the very basic question of what a word is. On one account, most “words” are in fact multiple entries in the mental lexicon; on the other, most words are a single lexical entry. However, distinguishing between these alternatives is extremely difficult, because there is no obvious behavioral test by which one can tell whether chicken has two different meanings associated to the same word or is two different words that have the same spelling, sound, and part of speech. In this work we used the MEG M350 component that has been argued to reflect initial stages of lexical activation as a tool to attempt to discover whether chicken and other polysemous words are represented as one word or as several.
Are Polysemy Effects Special? Hypotheses and Predictions
If senses are represented as their own lexical entries, like homonyms, then different senses of the same word should prime and inhibit each other in ways that are predictable from the way that phonologically and semantically related lexical items generally affect each other in processing. Semantic relatedness is known to be facilitory in a wide range of paradigms. Phonological relatedness, on the other hand, is often inhibitory, as similar-sounding words compete with each other in recognition. Thus, under the ‘separate lexical entries’ hypothesis, we might expect different senses of a polysemous word (such as green book – interesting book) to prime each other's activation less than purely semantic controls (such as green novel – interesting book), due to form-based inhibition. This prediction parallels those sometimes made in the derivational morphology literature. For example, Seidenberg & Gonnerman (2000) argue that if affixed words, such as teacher, do not decompose into their stems and affixes, then affixed words should prime their stems in ways that are predictable from general effects of semantic and phonological similarity, rather than showing identity priming (due to stem identity). Similarly, in the domain of polysemy, if green book and interesting book do not activate the same morpheme book, but rather two phonologically similar competitors, we would expect the two books to reflect both semantic priming and phonological inhibition.
In contrast, if senses are listed within the same lexical entry, sense-relatedness effects can be “special” and not necessarily explainable in terms of general effects of sound and meaning relatedness. Under this hypothesis, sense representations are part of the representations linking sound to meaning, called morphological roots (e.g., Halle & Marantz, 1993). For example, the lexical entry of paper links the phonological form of paper to separate sense representations. This hypothesis draws a crucial difference between polysemy and homonymy: In polysemy, senses share a morphological root, whereas homonyms have separate lexical representations. Consequently, under the shared root hypothesis, polysemy priming/inhibition effects should not show the effects of sound and meaning relatedness that are found in competing words, because the different senses are not different words. Instead, polysemy should rather show repetition priming (of the morphological root) and, potentially, sense competition. That is, seeing the word paper used in two different senses should activate the same lexical entry and so reveal repetition effects.
In this research we employed a priming paradigm and MEG to test these two hypotheses. As shown in Table 1, polysemy was contrasted with homonymy and semantic relatedness. The semantic condition modeled the kind of semantic relatedness that occurs in polysemy as closely as possible. Semantically related prime-target pairs were created by changing the target nouns of polysemous prime-target pairs to synonyms or near-synonyms. Priming was assessed relative to unrelated controls, and the subjects' task was to decide whether the two-word phrases make sense or not (a sensicality judgment). The structure of the experimental trials is shown in Fig. 1.
Table 1.
Design of stimuli.
| related prime | unrelated prime | target | |
|---|---|---|---|
| homonymous | river bank | salty dish | savings bank |
| polysemous | lined paper | military post | liberal paper |
| semantic | lined paper | clock tick | monthly magazine |
Figure 1.
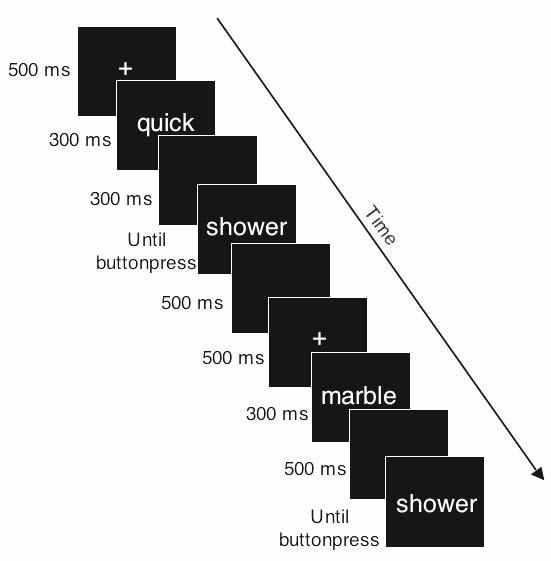
Timing of stimulus presentation.
Prior to the MEG study we also performed a purely behavioral study with the materials described in Table 1 as well congruous controls where there was no meaning or sense change between the prime and target (as in insightful book–difficult book). Space precludes a detailed report of the behavioral study but, importantly, we replicated the results of Klein and Murphy (2002), finding that using a word in the same sense twice led to faster judgments than switching senses. This pattern was identical for homonyms and polysemes. Thus, in purely behavioral terms, we do not have strong evidence for different lexical representations of these two forms of ambiguity. This underlines the importance of using more fine-grained measures, such as electrophysiology, to investigate how these two forms of ambiguity are represented.
As our main MEG dependent measure, we used the M350, a response component generated in the left superior temporal cortex at 300-400 ms after the onset of a visual word. The M350 constitutes an appropriate dependent measure for examining priming of morphological roots, as it shows many of the properties that one would expect of a neural index of lexical activation (Pylkkänen & Marantz, 2003). First, the M350 is the first MEG component in response to visual words that is sensitive to factors such as lexical frequency (Embick, Hackl, Schaeffer, Kelepir & Marantz, 2001) and repetition (Pylkkänen, Stringfellow, Flagg, & Marantz, 2001). Crucially, however, the M350 is not sensitive to late, decision-related factors and hence is not likely to reflect task-related processes (Pylkkänen, Stringfellow & Marantz, 2002). Finally, M350 latencies have been shown to track morphological constituent frequency rather than whole word frequency in compound processing (Fiorentino & Poeppel, 2004), which directly supports the hypothesis that the M350 is an index of morphological root access.
Given the evidence that the M350 reflects morphological root access, we hypothesized that if polysemy involves distinct sense representations but morphological root sharing, M350 sources should show shorter peak latencies for targets that are preceded by sense-related primes, as compared to unrelated controls. In contrast, if polysemy is represented like homonymy, M350 effects elicited in the polysemy comparison should be explainable as a combination of the effects elicited in the homonymy and semantic comparisons. While our hypotheses pertained to the M350 only, we also applied a multiple source model to the MEG data, in order to discover potential neural sources showing an inhibitory effect of sense-competition.
RESULTS
Sensicality Judgment Data
Consistent with earlier results, the behavioral data collected during the MEG experiment revealed very similar effects of polysemy and homonymy. Related targets (lined paper - liberal paper) were 50 ms faster than controls in the polysemy comparison (660 ms vs. 710 ms; F(1,16) = 6.84, p = .01) and 60 ms faster in the homonymy comparison (705 ms vs. 765 ms; F(1,16) = 8.32, p = .01). In the polysemy comparison, related targets were also responded to more accurately (83% vs. 70%; F(1,16) = 31.22, p < .001), whereas homonym targets did not differ from their unrelated controls in accuracy (both were 73%).
Semantic relatedness (e.g., green book - interesting novel) was facilitory, as related targets were responded to with 90% accuracy, compared to 80% on the unrelated controls (F(1,16) = 24, p < .001). Response speed did not show semantic priming (F < 1).
In sum, the behavioral data collected during the MEG experiment did not distinguish polysemy from homonymy. This necessitates investigation of earlier processing stages, allowed by the millisecond temporal resolution of MEG.
MEG Multiple Source Models
In order to obtain a maximally complete characterization of the neural sources activated by our stimuli, a multiple source model BESA (Brain Electrical Source Analysis, 5.0) was applied to all MEG data elicited at 0-500 ms. Source solutions were created from the grandaveraged responses of each individual subject to all critical items, and then kept constant across experimental conditions within a subject. First, the dipole sources of all major response components were modeled, as shown in Fig. 2. Unlike previous M350 studies, which have used sensors-of-interest analyses, we used data from all 148 sensors in all the source models. Although this method may increase between-subjects variance in localization, it has the advantage of removing any subjectivity associated with sensor selection. The magnetic field maps associated with major response components were used to guide the number of dipoles entered into source modeling. For example, if sensors covering the left and right hemispheres showed separate dipolar field patterns, two dipoles would be entered into the source model. Prior to statistical analysis, the source solutions of the all the major response components were combined into a single multidipole solution for each subject, where the sources were allowed to affect each other in intensity (but not in location or orientation). After this, peak latency and approximate source location were used as criteria for dividing the sources into the groups shown in Fig. 3, for the purposes of statistical analysis.
Figure 2.
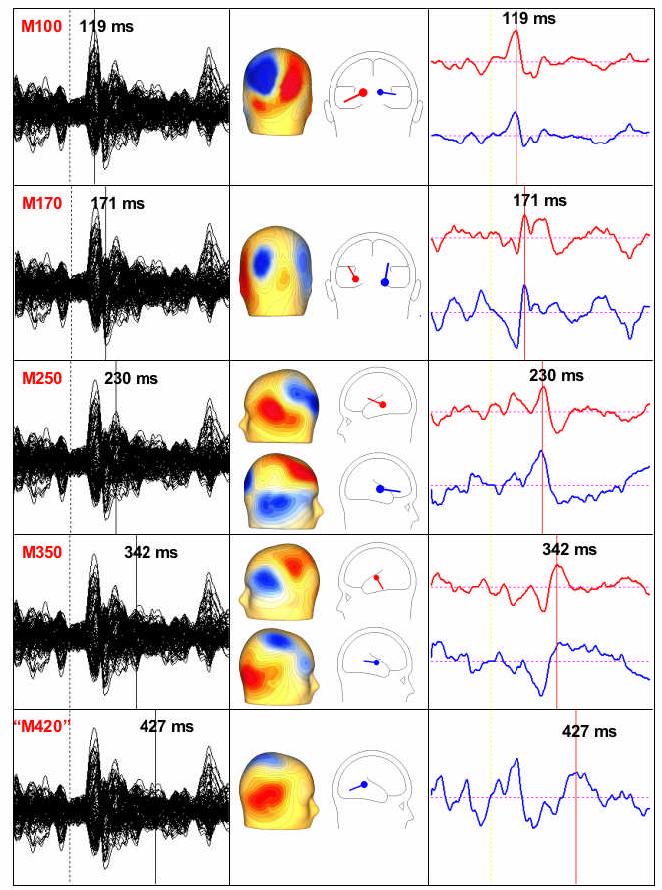
Sample data illustrating one participant's response to all critical trials and source solutions time-window by time-window. The left column shows activity in all 148 sensors. Component-specific source solutions were created for all time-points indicated by the cursor. The middle column shows the magnetic field maps at the timepoints of the cursor and the dipole localizations of the activity. The right column shows activity over time in the modeled sources.
Figure 3.

Results of multisource modeling for all 17 subjects, broken down into the groups that were entered into statistical analysis. Red dipoles represent all individual data points. Further, in order to visualize average activity in each time-window, mean dipole locations and orientations are plotted in black. Dipoles are plotted inside one average-sized participant's spherical head model. Since MRIs were not available for our subjects, we collected auditory M100 data at the end of the experiment to serve as a functional landmark representing primary auditory cortex (see Method). Although localizations of temporal activity at 200-500ms showed larger between-subjects differences than auditory M100 or early visual dipoles, statistical analyses of the midlatency temporal sources showed them to be statistically separable from each other (See Method).
While the M350 is generated by a left hemisphere source, some subjects, such as the one shown in Fig. 2, show a similar concurrent right-hemisphere source. Right hemisphere activity in the M350 time window often shows relatively little constancy across subjects. Consequently, previous M350 studies have not discussed it. However, our multidipole method (with no sensors of interest) did not allow us to ignore these right hemisphere sources when they were present. Therefore, this study serves as the first systematic investigation of right hemisphere activity in the M350 time-window during lexical processing.
Early visual responses
For 15 out of the 17 subjects, the first major activation peak occurred at 90-150 ms and was associated with a right-lateralized outgoing magnetic field and a left-lateralized re-entering field over the occipital sensors. This activity, the so-called visual M100, was explained best with a midline single dipole solution in 14 subjects and with a two-dipole solution in one subject. These localizations are consistent with previous MEG results where activity in this time window has localized in areas surrounding the V1 cortex (Type 1 activity in Tarkiainen, Helenius, Hansen, Cornelissen, & Salmelin, 1999).
The visual M100 was always followed by the M170 response, which exhibits a polarity reversal compared to the visual M100. M170 activity occurred at 150-220 ms and was best explained by a bilateral two-dipole solution in ten subjects and by a single mid-line dipole in seven subjects. Left hemisphere M170 activity has been shown to be specifically involved in letter-string processing (Tarkiainen et al., 1999).
Temporal/temporoparietal activity at 200-400 ms
Early visual responses were followed by a more complex pattern of activity, consisting of up to four major activation peaks. Left-hemisphere M350 field distributions (shown in Fig. 3) were first identified and the underlying activity modeled, including concurrent activity in the right hemisphere. After this, remaining activity was modeled.
The M350 (or “N400m”) is characterized by a downward oriented current in the left hemisphere at 300-400ms. Consistent with previous MEG studies on word processing (e.g., Helenius et al., 2002; Pylkkänen et al., 2002, 2004), we classified all dipoles meeting these criteria as M350 sources. Sixteen out of our 17 subjects had a characteristic M350 source. For one subject, activity at 300-400 ms localized to the midline of the back of the head; this activity could not be grouped with any other activity and was therefore not entered into statistical analysis.
Midlatency components at 300-400 ms typically exhibit more individual differences in localization than early components, such as the visual M100, the M170 or the auditory M100 (Cornelissen et al., 2003; Helenius, Salmelin, Service, & Connolly, 1999). In this dataset, the majority of M350 sources localized in superior and middle temporal areas, with four exceptions. The four sources outside the main cluster localized either in parietal cortex or in inferior temporal cortex (Fig. 3). Since these sources nevertheless exhibited the typical M350 orientation and timing, they were included in the M350 statistics. Results from electrical interference studies on transcortical sensory aphasia show that the localization of lexical access can in fact vary from parietal cortex to inferior temporal cortex (Boatman et al., 2000). Thus there is independent evidence from the functional lesion methodology that the variance obtained here is likely to represent true differences between individuals.
For nine of the subjects showing a left-hemisphere M350 field distribution (2 male), the left-hemisphere M350 localized in a physiologically plausible way only if a second dipole was added to the model. This second source always localized to the right hemisphere, either in temporal or parietal cortex.
Activity between the M170 and the M350 localized to the temporal regions, either bilaterally (n = 7), or to the left hemisphere only (n = 6). For one subject, activity at 200-300 ms localized to the right hemisphere only, and two subjects showed no prominent activation peaks between the M170 and the M350. Four subjects also showed temporal sources that peaked after the M350, which were clearly separate from the M350 in field distribution and dipole location. Three of these sources were in the left hemisphere, and four in the right (2 bilateral).
In order to test whether the various temporal sources were statistically separable from each other, analyses of variance were performed on all temporal source locations and orientations, including those of the auditory M100. These tests showed a reliable difference in location and/or orientation between all the components, with the exception of the M350 and the M420 sources (bilaterally), the locations and/or orientations of which differed within individuals, but not systematically enough to result in reliable effects in the group analysis (see Method). Black dipoles in Fig 3. depict mean dipole locations and orientations.
Late frontomedial activity
In addition to late activity in temporal regions, the majority of the subjects (n = 11) also showed a clear peak shortly after the M350, at 350-450 ms, which consistently localized to inferior frontal regions, close to midline (see Fig. 3). This finding is consistent with the results of previous MEG language studies, where medial prefrontal regions have been found to be activated both by visual words in a sentential context (Halgren et al., 2002; Pylkkänen, Llinás & McElree, 2005) as well as by a semantic judgment task on visual and auditory words with no sentential context (Marinkovic et al., 2003).
MEG Time Course Analysis: Bilateral Effects at 300-400 ms
Given that the main question of this research has to do with the lexical access of polysemous and homonymous words, the most important analyses concern the effect of priming condition on the left-hemisphere M350, a measure of lexical access. Relatedness had no reliable effects on the amplitudes or latencies of early visual sources, M250 activity, or late post-M350 activity, including temporal and frontal sources. However, relatedness did affect source latency (but not amplitude) at 300-400 ms in all comparisons.
As shown in Fig. 4, homonym targets elicited later left hemisphere M350 latencies (M = 355 ms) than unrelated controls (M = 334 ms; F(1,15) = 11.99, p < .01), suggesting inhibition. Semantic targets, on the other hand, elicited priming, M350 sources peaking earlier for semantically related (M = 345) than for unrelated targets (M = 367; F(1,15) = 9.04, p < .01). Given these effects of complete phonological overlap and semantic relatedness, what should we expect of polysemy effects if senses are stored as separate lexical entries? Since semantic relatedness elicited reduced left-hemisphere M350 latencies, one might expect some semantic priming for related senses. However, since homonymy elicited an M350 delay, and since related senses would be represented like homonyms, the amount of left hemisphere M350 priming should be less for polysemes than for the purely semantic targets. In contrast, under the hypothesis that polysemy involves morphological root identity, related senses should prime each other at least as much purely semantic controls.
Figure 4.
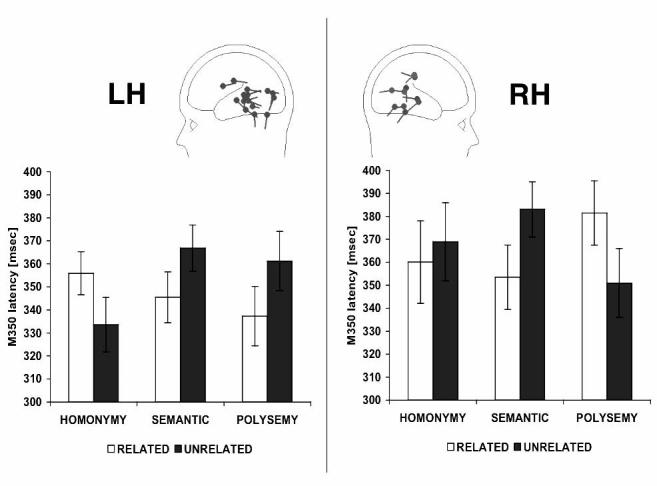
Effect of relatedness on M350 latencies. Error bars are SDs. The critical data are the priming effects (differences between related and unrelated conditions) for each word type.
Our data pattern in favor of the single lexical entry hypothesis. No indication of a homonym delay was present in the left hemisphere M350 data for polysemes. The amount of priming for related senses was comparable to the semantic priming effect. Polysemous related targets peaked at 337 ms on average, as compared to a mean latency of 361 ms for the unrelated targets (F(1,15) = 11.62, p < .01). In order to maintain to the separate lexical entries hypothesis, one would need to argue that phonologically similar lexical representations compete only when they are semantically unrelated. As far as we are aware, no model predicts this. In contrast, the single lexical entry hypothesis explains these data straightforwardly, with no additional assumptions.
As mentioned, under the single lexical entry hypothesis, processing might involve selection between separately listed senses within a lexical entry. We had no predictions as to what such an effect should look like, but our right hemisphere data at 300-400 ms offers suggestive evidence of a potential sense competition effect, as shown in Fig. 4. As explained above, nine of our subjects had right hemisphere sources at 300-400ms. Homonymy did not affect these sources. However, they showed a marginal semantic priming effect, sources peaking earlier for semantically related targets (M = 353) than for unrelated controls (F(1,8) = 4.14, p = .07). Thus this activity plausibly plays a role in semantic activation. Given that semantic relatedness was facilitory, one might expect some positive priming for polysemy as well, especially since the polysemous and semantic stimuli were closely matched in their semantic relations. Surprisingly, however, the polysemous stimuli showed exactly the opposite effect: related targets peaked reliably later (M = 381) than unrelated targets (M = 351; F(1,8) = 10.12, p < .05).
DISCUSSION
In this research we used magnetoencephalography to test whether distinct senses of the same word are represented as their own lexical entries. Since different senses of the same word have identical phonologies as well as related meanings, we aimed to first find out how phonological identity and semantic relatedness affect responses independently of polysemy, and then to investigate whether the effects of sense-relatedness can be explained in terms of combined effects of sound and meaning relatedness. Such a result would strongly support the hypothesis that different senses of the same word are represented as their own lexical entries, as there would be nothing special about polysemy effects that would not simply follow from the kinds of priming and inhibition that generally occur across lexical entries.
Our results are inconsistent with the separate lexical entries hypothesis. Related polysemous targets elicited a reliable left-hemisphere M350 latency reduction, which showed no indication of homonym competition. Similar results have recently been reported by Beretta, Fiorentino & Poeppel (in press), who contrasted polysemy and homonymy in single word lexical decision and found a facilitory effect of polysemy and an inhibitory effect of homonymy in left hemisphere M350 latency. We also obtained a special effect of polysemy in mid-latency right hemisphere sources: Polysemous targets elicited delayed latencies even though homonyms showed no effect, and purely semantic targets showed priming.
Thus, contrary to the predictions of the separate lexical entries hypothesis, polysemy effects are “special” and cannot be explained as mere combined effects of sound and meaning similarity. They can, however, be explained by the hypothesis that related senses share a morphological root, or “live” in the same lexical entry. On this hypothesis, different senses of the same word should activate an identical morphological root, and this was here evidenced by left hemisphere M350 priming. However, unlike (any straightforward versions of) sense-generation hypotheses, the shared morphological root hypothesis is also compatible with possible competition effects between senses, as senses stored within lexical entries could potentially stand in inhibitory relations to each other. Indeed, we found tentative evidence for such competition, as activity in the right hemisphere at 300-400 ms was delayed for polysemous sense-related targets.
In order to articulate our hypotheses in the context of morphological theory and previous M350 research, we have described the representational identity between related senses as identity of a morphological root. Frisson & Pickering (1999) formulate a representational hypothesis similar to ours except that what is identical between related, stored, senses is an abstract core meaning of a word. Putting aside doubts about whether there are such abstract core meanings (Klein & Murphy, 2002), this type of hypothesis could easily be construed as entailing morphological root sharing. Under such a formulation, it should make many of the same empirical predictions as the root-sharing hypothesis.
Left Hemisphere M350 as an Index of Morphological Root Access
Our MEG findings add to the increasing body of evidence that the M350 is a neural index of lexical access, and more specifically, an index of access to morphological root representations (Embick et al., 2001; Fiorentino & Poeppel, 2004; Pylkkänen & Marantz, 2003). If these morphological roots are the mental entities where the sound-meaning pairings of language are established, then the M350 should be sensitive not only to morphological identity, but also to sound and meaning relatedness. The present findings support this hypothesis directly: The M350 is not only affected by morphological identity but also by homonymy and semantic relatedness.
Effect of Homonymy
Ample behavioral evidence suggests that in lexical access a wide range of phonologically similar lexical representations are activated and compete for the selection of the optimal match to the input (Allopenna, Magnuson, & Tanenhaus, 1998; Goldinger, Luce, Pisoni, & Marcario, 1992; Grainger, O'Regan, Jacobs, & Segui, 1989; Luce & Large, 2001; Norris, McQueen, & Cutler, 1995; Soto-Faraco et al., 2001; Vitevitch & Luce, 1998, 1999; Vroomen & de Gelder, 1995). In cases of homonymy, this competition clearly cannot be solved on the basis of phonological-formal evidence alone; rather, semantic context must be used to disambiguate the stimulus. In these studies we disambiguated the homonym prior to its presentation (as in savings bank), and consequently, the present results bear on the mechanisms of homonym interpretation in unambiguous contexts. Our left-hemisphere M350 results show that the presentation of one homonym meaning delays the activation of a competing meaning on a subsequent trial. This suggests that homonym processing involves deactivation of a competing, context-incompatible representation.
A few previous electrophysiological studies have investigated homonym processing, using the N400 ERP as a dependent measure. The N400 is the closest ERP correlate to the magnetic M350 (Pylkkänen & Marantz, 2003; although see Friedrich, Kotz, Friederici & Gunter, 2004). In a semantic priming paradigm, Van Petten and Kutas (1987) showed that at long SOAs of 700 ms, contextually inappropriate homonym meanings are no longer active. This result conforms straightforwardly to the results of the present study, where SOA between prime and target nouns was always long as it included both the sensicality judgments to the prime as well as the presentation of the modifier of the target. Similar results were obtained by Hagoort and Brown (1994), who found larger N400 amplitudes for disambiguating words which occured two words after a homonym and were associated with the subordinate meaning of a homonym. Finally, Gunter et al. (2003) reported N400 evidence that working memory (WM) span affects homonym processing. Subjects with a high WM span inhibited nondominant homonym meanings, whereas for subjects with a low WM span, the alternative interpretation was still active after five subsequent words. Thus the time-course and/or strength of homonym inhibition may be modulated by WM span, a factor not manipulated here.
Effect of Semantic Relatedness
While phonological priming is still relatively understudied with the methods of cognitive neuroscience, semantic priming has been heavily researched. One of the best established results pertaining to the N400 ERP is that it is sensitive to various semantic properties of the stimulus; N400 amplitudes are systematically smaller for words that semantically “fit” the previous context, regardless of whether the context is sentential or simply an individual, semantically related word (e.g., Bentin, 1987; Besson & Macar, 1986; Holcomb & Neville, 1991; Kutas & Hillyard, 1980a, 1980b; Van Berkum, Hagoort, & Brown, 1999; Weisbrod et al., 1999).
The study reported here tested whether the types of semantic associative relations that occur in polysemy affect processing independently of polysemy. In other words, we aimed to find out how book primes novel when the prime and target mismatch in sense, i.e., priming from the concrete, object sense of book (green book) to the abstract, content sense of novel (interesting novel). We found that book reliably primes the activation of novel, as evidenced here by left-hemisphere M350 priming.
Right-Lateralized Temporal Activity at 300-400 ms
While our predictions pertained only to the left hemisphere M350, we also found priming and anti-priming effects for right-lateralized temporal activity at 300-400 ms. For semantically related targets, the right hemisphere effect paralleled the one found in the left hemisphere: decreased latencies for related targets. In contrast, polysemous related targets elicited a right-hemisphere effect that was opposite to the one found in the left-hemisphere: longer latencies for related than for unrelated targets. Homonyms elicited no effects in the right hemisphere.
How should we understand this mid-latency right-lateralized activity? Since only slightly more than half of our subjects showed this activity, and since our study was not designed to test hypotheses about activity other than the LH M350, any conclusions must be tentative. RH effects were found only when the prime and target were semantically related (i.e., in the semantic and polysemy comparisons), so it is likely that the RH activity reflects semantic rather than phonological processing. One possibility is that the RH activity performs some type of conceptual selection. However, no simple version of this hypothesis can explain our data since the polysemous and semantic targets showed opposite RH effects even though their semantic relations were matched. This suggests the intriguing possibility that semantic representations in the RH interact with each other in a qualitatively different way when they form part of the same lexical representation (as in polysemy) than when they do not. When the representations belong to the same lexical entry as in polysemy, they compete with each other, but when they belong to separate lexical entries, they prime each other. Clearly, further experimentation is needed to narrow down on the interpretation of the RH activity. However, given the hemispheric asymmetry obtained here, our results are broadly compatible with previous work suggesting that lexical-semantic processing is bilateral, with somewhat different functions in the left and right hemispheres (see Beeman, 1998; and Chiarello, 1998, for reviews).
CONCLUSION
This study investigated the basic question of how the brain represents the sound-meaning connections of natural language. Specifically, we addressed the question of polysemy, asking whether multiple related meanings of a single phonological/formal code are represented as one word or as many. Our results support representational identity in polysemy: Multiple related meanings with identical sound representations form part of a single lexical entry. Further, while semantic relatedness elicited a bilateral priming effect, sense-relatedness showed opposite effects in the left and right hemispheres. Thus our results add to the existing body of research suggesting that a model of the neurobiology of the mental lexicon crucially requires an understanding of the precise interplay between the left and right hemispheres.
METHOD
Participants
17 healthy right-handed native English speakers, aged between 18 and 32 (5 male) participated in the study. All participants gave their informed consent and had normal or corrected-to-normal vision.
Materials
180 prime-target pairs, divided into the six conditions shown in Table 1 (30 stimuli per condition), served as materials to investigate the effects of polysemy, homonymy and semantic relatedness. Priming was assessed with respect to unrelated controls. In order to examine the contribution of semantic priming to polysemy priming, semantically related prime-target pairs were created pairwise from sense-related prime-target pairs by changing the noun of the target phrase into a synonym or a near-synonym. For example, a semantically related pair was created from the polysemous pair green book - insightful book by replacing the target noun with novel (i.e., green book - insightful novel). All two-word phrases were either adjective-noun combinations (such as green book) or noun-noun compounds (baby bat). In order to equate any effects of syntactic priming, experimental conditions were matched on this variable.
In order to ensure that phonological repetition of the sort that occurred in the polysemous and homonymous prime-target pairs was not predictive of target phrase sensicality, our filler materials included 60 pairs where the noun repeated and the prime was sensible but the target was not (history lecture - yellow lecture), 60 pairs in which the prime and target were both nonsensical (pudding lamp – bullet lamp), and 40 pairs in which the prime was nonsensical and the target sensible (manual fish - saltwater fish). In addition, each participant saw 100 filler items that did not involve phonological repetition of the noun and which had either a nonsensical prime (n = 20), a nonsensical target (n = 40) or both (n = 40). Altogether, each participant saw 880 phrases, of which 360 were nonsensical. Since participants saw each target phrase twice, the critical stimuli were divided into two lists so that repetition was balanced across conditions. Further, the order of list presentation was counterbalanced across subjects. Stimulus presentation within lists was randomized.
The participants' task was to judge whether the two-word phrases made sense or not. The presentation of each phrase started with a fixation point (500 ms), followed by the modifier (300 ms), a brief blank (300 ms), and the noun, which stayed on the screen until the button press response. Participants were given the opportunity to rest after every 40 pairs.
Stimuli were presented in nonproportional Courier font (font size = 90). The timing of trials is shown in Fig. 1. Participants took approximately 50 minutes to complete the experiment.
Procedure and Apparatus
For the purposes of source localization, small electromagnetic coils were attached to the participant's head prior to the MEG measurement. Using a 3D digitizer, the locations of these coils were calculated with respect to three anatomical landmarks (the nasion and points just anterior to the ear canals), which established the head coordinate system for each participant. Once the participant was positioned in the MEG instrument, the coils were also localized with respect to the sensors. Thus MEG measurements could be transformed into each participant's individual head coordinate system. Since structural MRIs were not available for our participants, the shape of each participant's head was recorded during digitization. The head shapes were later used to estimate a spherical head model for each participant for the purposes of source localization.
During the experiment, participants viewed the experimental stimuli via fiber optic goggles (Avotec, FL). Participants indicated their sensicality judgments by button presses performed with the middle and index fingers of their left hand. Neuromagnetic fields were recorded with a whole-head, 148-channel neuromagnetometer array (4-D Neuroimaging, Magnes WH 2500) in epoch mode at a sampling rate of 678 Hz in a band between 1 and 200 Hz. After completing the sensicality judgment task, participants performed an auditory baseline test in which they listened to 100 1 kHz tones. Source locations of auditory M100 responses were later used as functional landmarks in source modeling.
Data Analysis
MEG data were averaged according to stimulus category by using an epoch length of 800 ms, preceded by a prestimulus interval of 300 ms. Artifact rejection excluded all trials that contained signals exceeding ± 2pT in amplitude and resulted in the exclusion of 15% of the trials. Prior to data analysis, MEG averages were low-pass filtered at 40Hz.
Although our experimental hypotheses were focused on the left temporal M350 response, a multiple-source model BESA was applied to all MEG activity elicited at 0-500 ms in order to obtain a fuller picture of the cortical regions activated by the task in both hemispheres. In order to use data with a maximally high signal-to-noise ratio in the source modeling, the grandaverages of each participant's responses to all critical target stimuli as well as responses to the primes in the homonym and polysemy comparisons were used in localization. The mean number of trials contained in these grandaverages was 252 (SD = 20). In the data analysis, the source models of the grandaverages were kept constant across experimental conditions within subjects.
Visual inspection of the magnetic field maps in the individual grandaverages revealed a pattern of responses familiar from previous MEG language studies (Embick, et al., 2001; Helenius et al., 1998, 1999; Pylkkänen, et al., 2001, 2002, 2004; Stockall, Stringfellow & Marantz, 2004). Sample data are depicted in Fig. 2 (for ease of visualization, all source wave peaks are plotted as positive). Early visual responses at ∼100 ms and ∼170-200 ms were generally followed by two major response components in the temporal regions, first one at around 250 ms (M250), and the second around 350 ms (M350). In some subjects, further temporal sources were found between 400-500 ms (as shown in Fig. 3), but in most subjects (in 11 out of 17), activity at 400-500 ms localized to anterior medial prefrontal regions.
MEG signals were first modeled time window by window, as shown in Fig. 2, and then introduced into a multiple source model for each subject. Sources were fit at the peaks of prominent response components, where dipolar field distributions were the clearest. Magnetic field maps guided the initial hypotheses about the number of dipoles used to model the activity in each time window. Only dipoles whose locations and orientations were consistent with the magnetic field patterns observed were entered into the multiple source solutions and further statistical analysis. The resulting multiple source models explained 85% (SD = 5%) of all the MEG activity in the grandaverages between 0 and 500 ms post stimulus-onset and 75% (SD = 8%) of the averages of the individual experimental conditions. Fig. 3 depicts all source localizations across subjects time window by time window, plotted inside one participant's spherical head model.
Statistical Analysis of Temporal Source Locations and Orientations
To test whether the various temporal sources were statistically separable from each other, all temporal source locations and orientations, including those of the auditory M100, were entered into a one-way ANOVA with Component as the independent factor with four levels: auditory M100, M250 (standing for activity between the M170 and the M350), M350, and M420 (standing for any post-M350 temporal/temporoparietal activity). Given that the number of sources in the left and right hemispheres was quite different for most time windows, left and right hemisphere sources were analyzed separately. With the exception of the M350 and the M420 sources (bilaterally), post-hoc pair-wise comparisons (Scheffé) showed a statistical separation between the dipole sources of all the response components either in location or in orientation, as described below. M420 sources were distinct in location and/or orientation from M350 sources within individual subjects, but there was little systematicity to this difference, resulting in no reliable localization differences in the group analysis.
In our coordinate system, the x-axis runs from left to right, the y-axis from posterior to anterior and the z-axis from inferior to superior, as is conventional. In the left hemisphere, Component had a reliable effect on y-coordinates (F(3,42) = 7.95, p < .001), both the M250 and the M350 localizing into areas posterior to the sources of the auditory M100 (p < .001 and p < .05, respectively). Also, Component had a reliable effect on z-coordinates (F(3,42) = 3.3; p < .05), M250 sources localizing into areas reliably inferior to the auditory M100 sources (p < .05). Left hemisphere dipole orientations were reliably affected by Component along all axes (Fs for x, y and z orientations all > 10). M350 and auditory M100 sources did not differ in orientation, but M350 source orientations did differ from those of M250 orientations (see Fig. 3; p's for all orientations < .001). Finally, while the locations of M420 sources were not statistically separable from any of the other sources, M420 sources did differ reliably from the auditory M100 sources in z-orientation (p < .01) as well as from M250 sources in y-orientation (p < .001). In the right hemisphere, Component had a reliable effect on x-coordinates (F(3,31) = 8.48; p < .001), auditory M100 sources localizing more right-laterally than both M250 sources (p = .001) and M420 sources (p < .01). Component also affected y-coordinates (F(3,31) = 5.82, p < .01), the M350 and the M420 both localizing to areas posterior to the auditory M100 sources (p < .05 for both). Similarly to the left hemisphere, right hemisphere dipole orientations were affected by Component along all axes (Fs > 6, p < .01). Again, M350 orientations were not statistically separable from auditory M100 orientations, but M250 and M420 sources were (p's < .001). Further, M350 and M250 y-orientations differed reliably (p < .001), as did M250 and M420 x-orientations (p < .05) and y-orientations (p < .001).
REFERENCES
- Allen M, Badecker W. Stem homograph inhibition and stem allomorphy: Representing and processing inflected forms in a multilevel lexical system. Journal of Memory and Language. 1999;41:105–123. [Google Scholar]
- Allopenna PD, Magnuson JS, Tanenhaus K. Tracking the time course of spoken word recognition using eye movements: Evidence for continuous mapping models. Journal of Memory and Language. 1998;38:419–439. [Google Scholar]
- Beeman M. Coarse semantic coding and discourse comprehension. In: Beeman M, Chiarello C, editors. Right hemisphere language comprehension: Perspectives from cognitive neuroscience. Erlbaum; Mahwah, NJ: 1998. pp. 255–284. [Google Scholar]
- Beretta A, Fiorentino R, Poeppel D. The effect of homonymy and polysemy on lexical access: an MEG study. Cognitive Brain Research. doi: 10.1016/j.cogbrainres.2004.12.006. in press. [DOI] [PubMed] [Google Scholar]
- Besson M, Macar M. Visual and auditory event-related potentials elicited by linguistic and non-linguistic incongruities. Neuroscience Letters. 1986;63:109–114. doi: 10.1016/0304-3940(86)90045-5. [DOI] [PubMed] [Google Scholar]
- Boatman D, Gordon B, Hart J, Selnes O, Miglioretti D, Lenz F. Transcortical sensory aphasia: revisited and revised. Brain. 2000;123:1634–42. doi: 10.1093/brain/123.8.1634. [DOI] [PubMed] [Google Scholar]
- Caramazza A, Grober E. Polysemy and the structure of the subjective lexicon. In: Rameh C, editor. Georgetown University roundtable on languages and linguistics. Semantics: Theory and application. Georgetown University Press; Washington, DC: 1976. pp. 181–206. [Google Scholar]
- Chiarello C. On codes of meaning and the meaning of codes: Semantic access and retrieval within and between hemispheres. In: Beeman M, Chiarello C, editors. Right hemisphere language comprehension: Perspectives from cognitive neuroscience. Erlbaum; Mahwah, NJ: 1998. pp. 141–160. [Google Scholar]
- Cornelissen P, Tarkiainen A, Helenius P, Salmelin R. Cortical effects of shifting letter position in letter strings of varying length. Journal of Cognitive Neuroscience. 2003;15:731–46. doi: 10.1162/089892903322307447. [DOI] [PubMed] [Google Scholar]
- Cruse DA. Lexical semantics. Cambridge University Press; Cambridge: 1986. [Google Scholar]
- Embick D, Hackl M, Schaeffer J, Kelepir M, Marantz A. A magnetoencephalographic component whose latency reflects lexical frequency. Cognitive Brain Research. 2001;10:345–348. doi: 10.1016/s0926-6410(00)00053-7. [DOI] [PubMed] [Google Scholar]
- Fellbaum C. Autotroponomy. In: Ravin Y, Leacock C, editors. Polysemy: Theoretical and computational approaches. Oxford University Press; Oxford: 2000. pp. 52–67. [Google Scholar]
- Fiorentino R, Poeppel D. Decomposition of compound words: An MEG measure of early access to constituents. In: Alterman R, Kirsch D, editors. Proceedings of the 25th Annual Conference of the Cognitive Science Society. Erlbaum; Mahwah, NJ: 2004. p. 1342. [Google Scholar]
- Friedrich CK, Kotz SA, Friederici AD, Gunter TC. ERPs reflect lexical identification in word fragment priming. Journal of Cognitive Neuroscience. 2004;16:541–52. doi: 10.1162/089892904323057281. [DOI] [PubMed] [Google Scholar]
- Frisson S, Pickering M,J. The Processing of metonymy: Evidence from eye movements. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1999;25:1366–1383. doi: 10.1037//0278-7393.25.6.1366. [DOI] [PubMed] [Google Scholar]
- Geeraerts D. Vagueness's puzzles, polysemy's vagaries. Cognitive Linguistics. 1993;4:223–272. [Google Scholar]
- Grainger J, Cole P, Segui J. Masked morphological priming in visual word recognition. Journal of Memory and Language. 1991;30:370–384. [Google Scholar]
- Grainger J, O'Regan JK, Jacobs AM, Segui J. On the role of competing word units in visual word recognition: The neighborhood frequency effect. Perception & Psychophysics. 1989;45:189–195. doi: 10.3758/bf03210696. [DOI] [PubMed] [Google Scholar]
- Gunter TC, Wagner S, Friederici AD. Working memory and lexical ambiguity resolution as revealed by ERPs: A difficult case for activation theories. Journal of Cognitive Neuroscience. 2003;15:643–567. doi: 10.1162/089892903322307366. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Brown C. Brain responses to lexical ambiguity resolution and parsing. In: Clifton C, Frazier L, Rayner K, editors. Perspectives on sentence processing. Erlbaum; Hillsdale: 1994. pp. 45–80. [Google Scholar]
- Halgren E, Dhond RP, Christensen N, VanPetten C, Marinkovic K, Lewine JD, Dale AM. N400-like MEG responses modulated by semantic context, word frequency, and lexical class in sentences. NeuroImage. 2002;17:1101–16. doi: 10.1006/nimg.2002.1268. [DOI] [PubMed] [Google Scholar]
- Halle M, Marantz A. Distributed morphology and the pieces of inflection. In: Hale K, Keyser SJ, editors. A View from Building. Vol. 20. MIT Press; Cambridge, MA: 1993. pp. 111–176. [Google Scholar]
- Helenius P, Salmelin R, Service E, Connolly JF. Distinct time courses of word and context comprehension in the left temporal cortex. Brain. 1998;121:1133–1142. doi: 10.1093/brain/121.6.1133. [DOI] [PubMed] [Google Scholar]
- Helenius P, Salmelin R, Service E, Connolly JF. Semantic cortical activation in dyslexic readers. Journal of Cognitive Neuroscience. 1999;11:535–550. doi: 10.1162/089892999563599. [DOI] [PubMed] [Google Scholar]
- Helenius P, Salmelin R, Service E, Connolly JF, Leinonen S, Lyytinen H. Cortical activation during spoken-word segmentation in nonreading-impaired and dyslexic adults. Journal of Neuroscience. 2002;22:2936–44. doi: 10.1523/JNEUROSCI.22-07-02936.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb O, Neville H. Natural speech processing: An analysis using event-related brain potentials. Psychobiology. 1991;19:286–300. [Google Scholar]
- Klein DE, Murphy GL. The representation of polysemous words. Journal of Memory and Language. 2001;45:259–282. [Google Scholar]
- Klein DE, Murphy GL. Paper has been my ruin: Conceptual relations of polysemous senses. Journal of Memory and Language. 2002;47:548–570. [Google Scholar]
- Kutas M, Hillyard SA. Event-related potentials to semantically inappropriate and surprisingly large words. Biological Psychology. 1980a;11:99–116. doi: 10.1016/0301-0511(80)90046-0. [DOI] [PubMed] [Google Scholar]
- Kutas M, Hillyard SA. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science. 1980b;207:203–205. doi: 10.1126/science.7350657. [DOI] [PubMed] [Google Scholar]
- Laudanna A, Badecker W, Caramazza A. Priming homographic stems. Journal of Memory and Language. 1989;28:531–546. [Google Scholar]
- Lehrer A. Polysemy, conventionality, and the structure of the lexicon. Cognitive Linguistics. 1990;1:207–246. [Google Scholar]
- Luce PA, Large N. Phonotactics, neighborhood density, and entropy in spoken word recognition. Language and Cognitive Processes. 2001;16:565–581. [Google Scholar]
- Marinkovic K, Dhond RP, Dale AM, Glessner M, Carr V, Halgren E. Spatiotemporal dynamics of modality-specific and supramodal word processing. Neuron. 2003;8:487–97. doi: 10.1016/s0896-6273(03)00197-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marslen-Wilson WD. Functional parallelism in spoken word-recognition. Cognition. 1987;25:71–102. doi: 10.1016/0010-0277(87)90005-9. [DOI] [PubMed] [Google Scholar]
- Norris D, McQueen JM, Cutler A. Competition and segmentation in spoken-word recognition. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1995;21:1209–1228. doi: 10.1037//0278-7393.21.5.1209. [DOI] [PubMed] [Google Scholar]
- Nunberg G. The non-uniqueness of semantic solutions: Polysemy. Linguistics and Philosophy. 1979;3:143–184. [Google Scholar]
- Pylkkänen L, Feintuch S, Hopkins E, Marantz A. Neural correlates of the effects of morphological family frequency and family size: an MEG study. Cognition. 2004;91:B35–B45. doi: 10.1016/j.cognition.2003.09.008. [DOI] [PubMed] [Google Scholar]
- Pylkkänen L, Llinás R, McElree B. Distinct effects of semantic plausibility and semantic composition in MEG. 2005 Submitted. [Google Scholar]
- Pylkkänen L, Marantz A. Tracking the time course of word recognition with MEG. Trends in Cognitive Sciences. 2003;7:187–189. doi: 10.1016/s1364-6613(03)00092-5. [DOI] [PubMed] [Google Scholar]
- Pylkkänen L, Stringfellow A, Flagg E, Marantz A. Proceedings of Biomag 2000, 12th International Conference on Biomagnetism. Helsinki University of Technology; Espoo, Finland: 2001. A neural response sensitive to repetition and phonotactic probability: MEG investigations of lexical access; pp. 363–367. [Google Scholar]
- Pylkkänen L, Stringfellow A, Marantz A. Neuromagnetic evidence for the timing of lexical activation: An MEG component sensitive to phonotactic probability but not to neighborhood density. Brain and Language. 2002;81:666–678. doi: 10.1006/brln.2001.2555. [DOI] [PubMed] [Google Scholar]
- Rice SA. Proceedings of the Fourteenth Annual Conference of the Cognitive Science Society. Erlbaum; Hillsdale, NJ: 1992. Polysemy and lexical representation: The case of three English prepositions; pp. 89–94. [Google Scholar]
- Seidenberg MS, Gonnerman L. Explaining derivational morphology as the convergence of codes. Trends in Cognitive Sciences. 2000;4:353–361. doi: 10.1016/s1364-6613(00)01515-1. [DOI] [PubMed] [Google Scholar]
- Simpson GB. Context and the processing of ambiguous words. In: Gernsbacher MA, editor. Handbook of psycholinguistics. Academic Press; San Diego: 1994. pp. 359–374. [Google Scholar]
- Soto-Faraco S, Sebastián-Gallés N, Cutler A. Segmental and suprasegmental mismatch in lexical access. Journal of Memory and Language. 2001;45:412–432. [Google Scholar]
- Stockall L, Stringfellow A, Marantz A. The precise time course of lexical activation: MEG measurements of the effects of frequency, probability, and density in lexical decision. Brain and Language. 2004;90:88–94. doi: 10.1016/S0093-934X(03)00422-X. [DOI] [PubMed] [Google Scholar]
- Tarkiainen A, Helenius P, Hansen PC, Cornelissen PL, Salmelin R. Dynamics of letter string perception in the human occipitotemporal cortex. Brain. 1999;22(Pt 11):2119–32. doi: 10.1093/brain/122.11.2119. [DOI] [PubMed] [Google Scholar]
- Van Berkum JA, Hagoort P, Brown CM. Semantic integration in sentences and discourse: Evidence from the N400. Journal of Cognitive Neuroscience. 1999;11:657–671. doi: 10.1162/089892999563724. [DOI] [PubMed] [Google Scholar]
- Van Petten C, Kutas M. Ambiguous words in context: An event-related potential analysis of the time course of meaning activation. Journal of Memory and Language. 1987;26:188–208. [Google Scholar]
- Vitevitch MS, Luce PA. When words compete: Levels of processing in spoken word recognition. Psychological Science. 1998;9:325–329. [Google Scholar]
- Vitevitch MS, Luce PA. Probabilistic phonotactics and neighborhood activation in spoken word recognition. Journal of Memory and Language. 1999;40:374–408. [Google Scholar]
- Vroomen J, de Gelder B. Proceedings of the Fourth European Conference on Speech Communication and Technology. Madrid, Spain: 1995. Lexical inhibition in spoken word recognition; pp. 1711–1714. [Google Scholar]
- Weisbrod M, Kiefer M, Winkler S, Maier S, Hill H, Roesch-Ely D, Spitzer M. Electrophysiological correlates of direct versus indirect semantic priming in normal volunteers. Cognitive Brain Research. 1999;8:289–298. doi: 10.1016/s0926-6410(99)00032-4. [DOI] [PubMed] [Google Scholar]
- Zipf GK. The meaning-frequency relationship of words. Journal of General Psychology. 1945;33:251–256. doi: 10.1080/00221309.1945.10544509. [DOI] [PubMed] [Google Scholar]