

Abstract
We applied a high-resolution PCR-based typing method, multiple-locus variable-number tandem repeat analysis (MLVA), for discrimination of 30 multidrug-resistant clinical isolates of Staphylococcus epidermidis. The results of MLVA were congruent with results obtained by pulsed-field gel electrophoresis (PFGE). MLVA generated discrete character data, and its discriminatory capacity was comparable to that of PFGE.
The clinical significance of Staphylococcus epidermidis is increasingly recognized. The bacterium is primarily associated with nosocomial infections such as prosthesis- and intravascular-catheter-related infections and a variety of postoperative infections (17). Moreover, nosocomial isolates of S. epidermidis frequently exhibit multidrug resistance, which limits the therapeutic arsenal (5). Despite the growing importance of S. epidermidis as a cause of hospital-acquired infections, there is still limited information available regarding the epidemiology of S. epidermidis in the hospital setting. Methods that may distinguish clinically significant strains from contaminant strains are lacking. For epidemiological studies, there is also a lack of easy-to-use, rapid typing methods with high reproducibility. Since the 1990s, pulsed-field gel electrophoresis (PFGE) of whole-genome SmaI digests has been considered the “gold standard” for molecular typing of S. epidermidis (13, 14). The method has high discriminatory power but is laborious and therefore costly. Interpretation and exchange of PFGE-typing data are not uncomplicated because they depend on banding patterns and subjective decisions regarding the true existence of discrete bands. There is a consensus that exchange of typing data between laboratories is highly desirable, and systems with a binary output (numbers or characters), rather than methods generating banding patterns such as PFGE, are therefore preferred. Multiple-locus variable-number tandem repeat analysis (MLVA) has proved efficient for high-resolution typing of several bacterial species (4, 6, 16). The method targets multiple genomic loci and relies on the detection of different copy numbers of short repeated sequences that are arranged in tandem arrays. MLVA is an easy-to-use, PCR-based method that provides simple numerical data by identifying the copy numbers of tandem repeats that are present at the assayed genomic loci. The aim of the present study was to develop and evaluate an MLVA system for molecular typing of S. epidermidis.
Collection of bacteria and epidemiological information.
Clinical isolates of multidrug-resistant S. epidermidis were obtained during November and December 2001 and October and November 2002 from clinical specimens from patients treated at different ward units at the 700-bed University Hospital of Umeå, Sweden. Epidemiological information was retrieved from admission notes of clinical specimens and by a retrospective review of hospital records for each patient. Each isolate was assigned an arbitrary isolate identification number for this study. Identification of bacterial cultures to the species level was performed by using biochemical typing as previously described (10). To evaluate the species specificity of the MLVA, one reference strain of each of 18 different staphylococcal species was obtained from the American Type Culture Collection (ATCC), Manassas, Va., or the Czech Collection of Microorganisms (CCM), Brno, Czech Republic.
Susceptibility analysis.
Antibiotic susceptibility analysis was performed by the agar dilution technique with a 25-pin multipoint inoculation device as previously described (2, 9). Briefly, 90-mm agar plates were prepared that contained dilutions of six antimicrobial agents. PDM ASM agar (AB Biodisk, Solna, Sweden) supplemented with 5% defibrinated horse blood was used with fusidic acid, cotrimoxazole, gentamicin, vancomycin, and clindamycin, while Mueller-Hinton II agar (Becton Dickinson, Sparks, Md.) supplemented with 2% NaCl was used with oxacillin (8). Bacterial suspensions for inoculation were prepared and susceptibility breakpoints applied according to the Swedish Reference Group for Antibiotics (3). Plates were read after 24 h of incubation at 35°C. All selected clinical isolates exhibited a uniform pattern of multidrug resistance with resistance to oxacillin, fusidic acid, cotrimoxazole, gentamicin, and clindamycin. All isolates were vancomycin susceptible.
Identification of variable-number tandem repeats.
The genome sequence of S. epidermidis ATCC 12228 (accession no. AE015929) was downloaded from GenBank and analyzed for the presence of tandem repeat sequences with the software program Tandem Repeats Finder (1). The analysis identified 137 genomic regions with tandem repeat sequences. Fourteen regions were selected for further analysis on the basis of genomic location, repeat length, and copy number. The selected tandem repeat sequences exhibited a minimum size of 18 bp for an individual repeat and a tandem copy number greater than two.
MLVA and PFGE.
Bacteria were grown on blood agar at 35°C overnight, and DNA was extracted with the QIAamp DNA mini kit (QIAGEN, Hilden, Germany). DNA concentration was determined with a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, Del.). Initially, selected tandem repeat loci were investigated by PCR analysis of a panel of 10 S. epidermidis isolates. Flanking primers were designed for each repeat locus based on the genome sequence of strain ATCC 12228. PCR mixtures of 50 μl contained 0.4 μM forward and reverse primers (DNA Technology A/S, Århus, Denmark), 0.05 U of AmpliTaq Gold (Applied Biosystems, Stockholm, Sweden), 0.2 mM each deoxynucleoside triphosphate (Amersham Biosciences, Buckinghamshire,England), 1.5 mM MgCl2, and 60 ng of template DNA in PCR buffer II with bovine serum albumin. DNA amplification was carried out by initial denaturation at 95°C for 10 min and then cycling at 95°C for 1 min, 55°C for 1 min, and 72°C for 1 min for 30 cycles, with a final incubation at 72°C for 5 min in a DNA Engine PTC-200 (MJ Research Inc., Watertown, Mass.).
DNA fragments from each strain at all 14 loci were successfully amplified and the sizes determined on agarose gels. Five loci, designated Se1 to Se5, that showed size polymorphism among isolates of the test panel were dispersed over the chromosome and therefore selected for further analysis in 30 clinical isolates of S. epidermidis (Fig. 1 and Table 1). PCR amplification products were analyzed for size variation on agarose gels with 2% NuSieve (FMC BioProducts) for Se1 to Se4 or 1.75% SeaKem GTG (Cambrex Bio Science) for Se5. We used the molecular weight standards GeneRuler 100 bp DNA Ladder (Fermentas International Inc., Vilnius, Lithuania) and GeneRuler 100 bp DNA Ladder Plus (Fermentas). The sizes of the PCR amplification products obtained were translated to repeat copy numbers for each isolate (Table 2). To verify repeat copy number variation, PCR products representing two different isolates were sequenced at each of the five repeat loci on the ABI 377 platform with forward and reverse amplification primers and the BigDye terminator cycle sequencing ready reaction kit (PE Applied Biosystems). Each isolate was genotyped by PFGE with SmaI digests of whole-genome DNA as previously described (9). Macrorestriction profiles were interpreted as described by Tenover et al. (15).
FIG. 1.
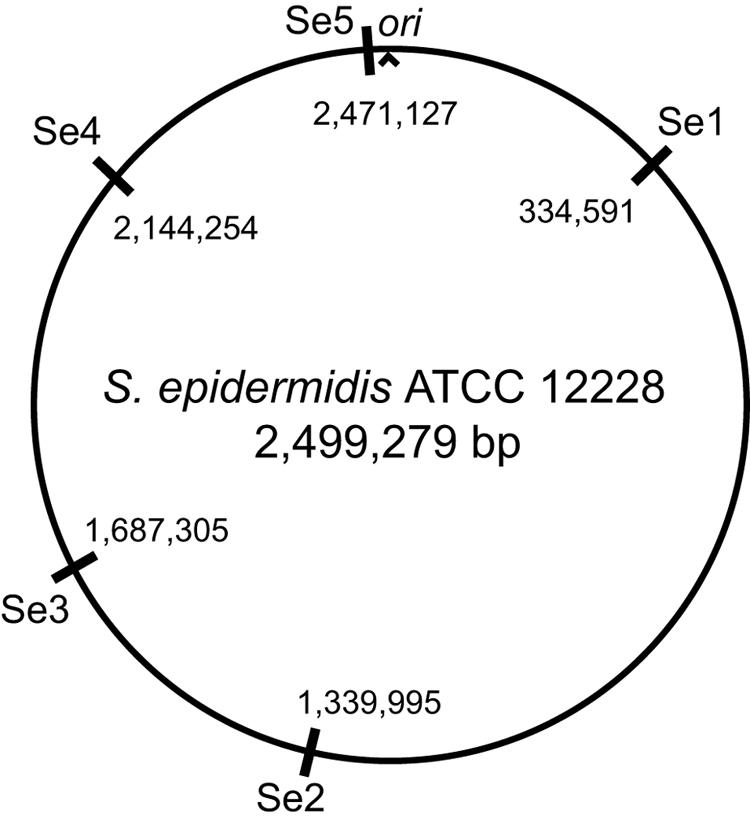
Genomic locations of investigated variable-number tandem repeats within the circular map of S. epidermidis strain ATCC 12228. Positions are given with reference to the origin of replication (ori).
TABLE 1.
Attributes of variable-number tandem repeat loci used for typing of S. epidermidis isolates
| Variable-number tandem repeat locus | Repeat motif | Repeat size (nt)a | Genomic location and gene designationb | Repeat copy no. of strain ATCC 12228 | Primer sequences 5′-3′ (forward/reverse) | Allele no.c | Dd value |
|---|---|---|---|---|---|---|---|
| Se1 | TCAGACAGCGACTCAGATGGC CCTAGTCCCAACTTGC | 18 | 334591-335113; G (SE0331) | 29 | GCTGATGGGGAAGAAGTTCA/ AACGCTCCTAAACCTGCAAA | 8 | 0.65 |
| Se2 | TCTGCCTGTTGAATTTCTTTG TGAAATTCTCTTTGTTGG | 58 | 1339995-1340177; I | 3 | AGGCCCAAATAAAAAGCAAA/ AACTGACGCTCCAGGAGAAG | 4 | 0.54 |
| Se3 | TCTGAATCACTATCTGAACCC CAACCCCAACTTGCTT | 18 | 1687305-1687643; G (SE1632) | 18 | TTTCCGGTATGTGAACCCTTA/ TGACACTAGTCGCACAGGAA | 7 | 0.67 |
| Se4 | TGTCCATGGAATTTCTTCGAA AATTCTCTCTGTTGGGG | 57 | 2144254-2144479; G (SE2101) | 4 | TTCATTGTCCCCTGTCTTCT/ TCGATCCTGGTAAAGCGATTA | 3 | 0.51 |
| Se5 | GAATCCGAGTCACTGTCT | 18 | 2471127-2472499; G (SE2395) | 75 | GGCCATATAGACCTGGCTTG/ AGATGCTGATGGGGAAGATG | 9 | 0.80 |
Indicates repeat size as number of nucleotides (nt).
G or I indicate that the repeat motif is located within an open reading frame (genic) or between two open reading frames (intergenic), respectively. Genomic locations and gene designations refer to the genome sequence of S. epidermidis ATCC 12228.
The allele number tells how many different repeat copy variants were detected among isolates.
Individual marker diversity (D) was calculated as 1 − Σ(allele frequency)2 and based upon allele frequencies of distinct genotypes among 31 S. epidermidis isolates.
TABLE 2.
Repeat copy numbers of S. epidermidis isolates at five variable-number tandem repeat loci and information on their clinical backgrounds
| Isolate | Clinical context (yr) | PFGE banding patterna | Repeat copy no. at tandem repeat locus:
|
||||
|---|---|---|---|---|---|---|---|
| Se1 | Se2 | Se3 | Se4 | Se5 | |||
| ATCC 12228 | Non-biofilm-forming reference strain | A3 | 29 | 3 | 18 | 4 | 75 |
| 1 | SSIc pancreatic surgery (2002) | A | 23 | 1 | 14 | 1 | 57 |
| 2 | Pacemaker infection, thoracic surgery (2002) | A | 23 | 1 | 14 | 1 | 57 |
| 3 | Bone tissue culture, revision of hip arthroplasty (2002) | A | 23 | 1 | 14 | 1 | 57 |
| 4 | Sternal osteomyelitis, cardiac valvular replacement (2002) | A | 23 | 1 | 14 | 1 | 57 |
| 5 | SSI, internally fixed elbow fracture (2001) | A | 23 | 1 | 14 | 1 | 57 |
| 6 | SSI, internally fixed tibial fracture (2001) | A | 23 | 1 | 14 | 1 | 57 |
| 7 | CVCd tip culture, drug-induced rhabdomyolysis (2001) | A | 23 | 1 | 14 | 1 | 57 |
| 9 | Endophthalmitis, cataract surgery (2002) | A | 23 | 1 | 14 | 1 | 57 |
| 22 | Femoral artery access site infection, acute aortic dissection (2001) | A | 23 | 1 | 14 | 1 | 57 |
| 8 | Temporary pacing lead culture, acute aortic dissection (2001) | A | 23 | 1 | 12 | 1 | 57 |
| 14 | CVC tip culture, endocarditis (2002) | A1 | 23 | 1 | 14 | 1 | 58 |
| 15 | CVC tip culture, cardiac valvular replacement (2002) | A1 | 23 | 1 | 14 | 1 | 58 |
| 17 | Leg ulcer, plastic surgery (2001) | A | 23 | 1 | 14 | 1 | 58 |
| 10 | CVC tip culture, revision of hip arthroplasty (2002) | A2 | 23 | 1 | 14 | 1 | 59 |
| 11 | CVC tip culture, coiling of cerebral aneurysm (2002) | A2 | 23 | 1 | 14 | 1 | 59 |
| 12 | SSI, failed internal fixation of hip fracture (2001) | A2 | 23 | 1 | 14 | 1 | 59 |
| 13 | Femoral artery catheter tip culture, gastroesophageal resection (2001) | A2 | 23 | 1 | 14 | 1 | 59 |
| 29 | SSI, kidney transplantation (2001) | G | 8 | 4 | 19 | 1 | 55 |
| 30 | Nose swab culture, neutropenia (2002) | G | 8 | 4 | 19 | 1 | 0b |
| 28 | Incisional SSI, hip arthroplasty (2002) | B | 8 | 1 | 13 | 4 | 0b |
| 23 | Face wound, fatal multitrauma (2001) | D | 38 | 2 | 18 | 1 | 69 |
| 18 | Skin ulcer culture, neutropenia (2001) | C | 9 | 1 | 20 | 2 | 60 |
| 16 | Nose swab culture, neutropenia (2001) | E | 39 | 2 | 12 | 2 | 59 |
| 24 | Central venous port culture, chronic liver disease (2002) | F | 39 | 2 | 11 | 2 | 59 |
| 26 | CVC tip culture, revision of hip arthroplasty (2002) | K | 34 | 2 | 13 | 4 | 74 |
| 19 | Perianal abscess, surgical incision (2001) | I | 38 | 2 | 13 | 4 | 74 |
| 21 | Perirectal abscess, fistula of Crohn's disease (2001) | I1 | 38 | 2 | 13 | 4 | 74 |
| 27 | SSI, limb amputation (2002) | I2 | 38 | 2 | 13 | 4 | 74 |
| 20 | Blood culture, sepsis, gastric resection (2001) | H | 37 | 2 | 13 | 4 | 0b |
| 25 | CVC tip culture, lung resection (2002) | J | 38 | 2 | 13 | 4 | 0b |
Banding patterns were interpreted as described by Tenover et al. (15). Banding patterns were assigned arbitrary letter codes (A, B, C…). Different letters indicate unrelated isolates (seven or more band differences). Closely or possibly related banding patterns (two to six band differences) were considered subtypes and are indicated by number codes (A, A1, A2…).
DNA of these isolates was not amplified.
SSI, surgical site infection.
CVC, central venous catheter.
Data analysis.
The discrete-character MLVA data (i.e., the repeat copy numbers found at the five repeat loci) were analyzed with Bionumerics v. 3.5 (Applied-Maths, Saint-Martens-Latem, Belgium), the categorical coefficient, and the unweighted pair group method using arithmetic averages. To estimate the discriminatory power of the typing method, allelic diversity (D) was calculated with the formula D = 1 − Σ(allele frequency)2, i.e., a function of individual allele frequencies and the number of alleles detected in the population.
Clinical and epidemiological data.
The spatial distribution of collection sites of 30 multiresistant clinical S. epidermidis isolates at Umeå University Hospital is depicted in Fig. 2. Twelve isolates were collected in a setting of suspected postoperative deep surgical site infection in the presence of implanted foreign material, and most of them were from the thoracic surgery unit or the orthopedic surgery unit (Fig. 2; also Table 2). Culture of central venous catheters yielded eight isolates. The remaining isolates were retrieved by collection of more diverse specimens including screening cultures of neutropenic patients and specimens from superficial surgical incision site infections. Collection of culture specimens was initiated by clinicians and hence not performed due to a suspected nosocomial outbreak.
FIG. 2.

Spatial distribution of hospital localities for collection of 30 multiresistant S. epidermidis isolates at Umeå University Hospital, Umeå, Sweden. Isolate identification numbers are circled. Asterisks indicate isolates of a genetic clone as defined by MLVA and PFGE typing. The data presented show that a multiresistant S. epidermidis clone was widespread at the hospital.
MLVA compared to PFGE.
The variable repeat loci used in MLVA displayed individual repeat sizes ranging from 18 bp to 58 bp (Table 1). Among the 30 isolates of S. epidermidis analyzed and control strain ATCC 12228, the number of alleles at repeat loci ranged from three for Se4 to nine for Se5. D values ranged from 0.51 for Se4 to 0.80 for Se5. The D value for the combination of five repeat loci (Se1 to Se5 treated as a single allele) was 0.87. To compare MLVA with an established typing method, PFGE with SmaI-digested DNA was performed. It should be noted that we used permissive criteria for identifying unique PFGE patterns; a two-band difference was set to identify unique patterns. The D value obtained for typing by PFGE was 0.84, a value comparable to that obtained by MLVA. The finding of relatively low D values results from the abundance of multiple isolates exhibiting identical genotypes in the bacterial population under study (Fig. 3).
FIG. 3.

Cluster analysis on the basis of MLVA data demonstrating the congruence of results obtained by MLVA and PFGE typing. Letter codes indicate PFGE banding patterns, where A, A1, A2, and A3 indicate closely or possibly related patterns. (A) Five-locus (Se1 to Se5) MLVA cluster analysis. (B) Four-locus (Se1 to Se4) MLVA cluster analysis. An asterisk indicates a cluster of isolates that exhibited extensive genetic similarity (i.e., a genetic clone).
In performing MLVA, we noted PCR amplification failure for four S. epidermidis isolates at locus Se5, while all other PCR amplifications at loci Se1 to Se5 were successful (Table 2). The amplification failure at Se5 persisted despite the use of four different flanking primer pairs. The easiest explanation for this is that the targeted gene is missing in some S. epidermidis strains. PCRs with primers for Se1 to Se5 did not amplify DNA of the following non-S. epidermidis staphylococcal strains: S. aureus ATCC 29223, S. equorum CCM 3832, S. capitis ATCC 27840, S. caseolyticus CCM 3540, S. carnosus CCM 3885, S. cohnii CCM 2736, S. gallinarum CCM 3572, S. haemolyticus ATCC 29970, S. hominis CCM 2733, S. lentus CCM 3472, S. lugdunensis ATCC 48309, S. saprophyticus ATCC 15305, S. simulans ATCC 27849, S. sciuri CCM 3473, S. schleiferi ATCC 43808, S. warneri ATCC 27836, and S. xylosus ATCC 29971. Thus, it seems that the MLVA identifies the species S. epidermidis and discriminates within the species.
It was apparent that MLVA and PFGE typing resulted in very similar clustering of isolates (Fig. 3). Hierarchical clustering of S. epidermidis isolates on the basis of MLVA data revealed 16 genotypes among the 31 isolates analyzed. This compares with the 16 unique banding patterns that were resolved by PFGE. The result of MLVA was, with very few exceptions, congruent with the results obtained by PFGE typing. Eleven isolates showed the same PFGE banding pattern, and nine of these were also identical by MLVA (Fig. 3). The 10th and 11th isolates each differed at only one variable-number tandem repeat locus, at Se3 (isolate 8) and Se5 (isolate 17), respectively (Fig. 3; also Table 2). Three isolates exhibited the same MLVA genotype, while typing by PFGE showed a two-band difference for each strain (isolates 19, 21, and 27). Taken together, our findings indicate that MLVA has a discriminatory capacity similar to that of typing by PFGE.
A multiresistant S. epidermidis clone appeared to be widespread.
Spatial analysis of the dissemination of multiresistant S. epidermidis genotypes at the hospital indicated that many isolates were closely related and widespread in the hospital environment (Fig. 2 and 3). A straightforward interpretation is that an S. epidermidis clone was maintained in the hospital environment during the study period and that this clone was highly successful in colonizing patients. The limited number of isolates makes further epidemiological interpretations difficult, although an association of the predominant genotype with deep surgical site infection and/or the presence of implanted foreign medical devices seemed to be present. Spatial analysis showed an association of the predominant genotype with the thoracic surgery unit, the ophthalmology-plastic surgery unit, and the orthopedic surgery unit. There is, however, good reason for caution due to the limited size of the present study. Previous work with PFGE typing has failed to find firm associations of a particular genotype with various clinical parameters but corroborates the finding of highly successful multiresistant S. epidermidis clones that are geographically disseminated (7).
Congruent typing results indicate robust measurements of genetic relationships.
The finding of congruence of PFGE typing with MLVA argues that the genomic targets of MLVA and PFGE share evolutionary histories although MLVA and PFGE typing measure very different evolutionary events. PFGE is based on restriction cleavage and may detect single nucleotide mutations at a few cleavage sites but is especially sensitive for measuring recombination events within a genome. In contrast, MLVA is designed to target multiple genomic regions that are prone to rapid mutation by change of copy numbers of tandem repeat sequences. The congruent results obtained by two methods measuring different mutational mechanisms at multiple genomic loci indicate that the sequences targeted by these methods were horizontally transferred infrequently or not at all among the S. epidermidis genomes included in this study. Our findings suggest that the genomic loci targeted by MLVA provide a robust measure of genetic relationships.
Tandem repeats were located in genes encoding fibrinogen-binding proteins.
Strikingly, three of five polymorphic repeat loci identified in S. epidermidis (Se1, Se3, and Se5) were located in genes predicted to encode surface binding proteins that may have a functional role in adherence (Table 1). The loci were located within open reading frames predicted to encode fibrinogen-binding proteins of the serine-aspartate (SD) repeat-containing (Sdr) family of cell wall-anchored surface proteins. This protein family is believed to play a pathogenic role in some gram-positive bacteria by enabling binding to host fibrinogen (15). The 18-bp tandem repeats identified at Se1, Se3, and Se5 are predicted to encode SD stretches of variable length in Sdr proteins. Different copy number of an 18-bp repeat at Se1, Se3, or Se5 will only affect the length of SD stretches without disrupting the open reading frames. Similar to our findings, a previous MLVA study of methicillin-resistant S. aureus found variation in genes encoding Sdr proteins among different strains (12). In another study addressing protein structure, it was shown that the SD stretch of a member of the Sdr family, SdrG of S. epidermidis, is localized just outside a cell wall-spanning region (11). The SD stretch was not shown to have a functional role for ligand binding or cell wall attachment. Our observation that three out of five targeted repeat sequences code for putative fibrinogen-binding proteins requires further exploration. First, it would be valuable to explore whether there is any correlation of MLVA genotypes with the adherence properties of S. epidermidis. Second, since adherence to host fibrinogen is considered a functional trait of pathogenic S. epidermidis, evolution at tandem repeats encoding Sdr proteins might be functionally biased. A common environmental selective pressure might cause convergent evolution, especially at a rapidly evolving tandem repeat locus, thereby biasing the estimation of the genetic distance among isolates.
Adjustment of discriminatory capacity.
The discriminatory capacity of an MLVA system can be adjusted by inclusion of different tandem repeat loci exhibiting different variabilities. We observed that the use of five repeat loci (Se1 to Se5) for MLVA resulted in a high discriminatory capacity (Fig. 3). Interestingly, the use of only four repeat loci (Se1 to Se4) resulted in a highly similar clustering of isolates while slightly decreasing the discriminatory capacity. By using four loci, isolates that were judged to be closely or possibly related by PFGE typing (showing two to six band differences) clustered as a single MLVA genotype (Fig. 3). A reduction in discriminatory capacity might be valuable in epidemiological investigations spanning larger geographical regions or time scales and in studies of population dynamics. In such studies, highly variable markers might dim the interpretation of genetic relatedness due to homoplasy effects. To further stabilize the S. epidermidis MLVA, the inclusion of additional tandem repeat loci exhibiting low variability might be considered. In local investigations over shorter time periods, highly variable markers will be important.
Concluding remarks.
MLVA of S. epidermidis is potentially cost saving compared to PFGE typing because of the speed of analysis and a limited need for sophisticated equipment. The current protocol was designed to avoid extensive use of sequencing facilities, although we have verified the sequence data of selected isolates. Limitations of the present study include a restricted number of isolates investigated and a relative homogeneity of isolates since they were collected in only one hospital and exhibited uniform resistance to multiple antimicrobial agents. This is illustrated by the relatively low discrimination indices obtained. For comparison, a PFGE study by Miragaia et al. (7) on a more diverse methicillin-resistant S. epidermidis collection from multiple hospitals showed excellent discriminatory power. The calculation criteria of the present study and their published PFGE data result in a D value of 0.98 (106 PFGE subtypes, 230 isolates). Future studies should evaluate MLVA of S. epidermidis with a larger number of isolates exhibiting greater diversity.
In conclusion, the present study demonstrates that MLVA is a promising typing approach for molecular epidemiology studies of S. epidermidis due to the speed of analysis, the low cost, and the ability to produce numerical data that can be easily exchanged among laboratories. We believe that MLVA will be a valuable tool for investigations of population dynamics, maintenance, and spread of nosocomial S. epidermidis.
Nucleotide sequence accession numbers.
The nucleotide sequences of the S. epidermidis variable-number tandem repeat loci reported in this paper (Se1 to Se5) have been deposited in the GenBank database. The sequences at each of the five loci that represent two different isolates with two different repeat copy number variants have been assigned accession no. DQ247711 to DQ247722.
Acknowledgments
This work was supported by funding from the Medical Faculty, Umeå University, Umeå, Sweden, and the Swedish MoD, project A4854.
REFERENCES
- 1.Benson, G. 1999. Tandem Repeats Finder: a program to analyze DNA sequences. Nucleic Acids Res. 27:573-580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Burman, L. G., and R. Östensson. 1978. Time- and media-saving testing and identification of microorganisms by multipoint inoculation on undivided agar plates. J. Clin. Microbiol. 8:219-227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kahlmeter, G. April 20 2005, revision date. SRGA homepage, version 3. The Swedish Reference Group for Antibiotics (SRGA) and its subcommittee on methodology (SRGA-M). [Online.] http://www.srga.org/. Accessed 24 April 2005.
- 4.Keim, P., L. B. Price, A. M. Klevytska, K. L. Smith, J. M. Schupp, R. Okinaka, P. J. Jackson, and M. E. Hugh-Jones. 2000. Multiple-locus variable-number tandem repeat analysis reveals genetic relationships within Bacillus anthracis. J. Bacteriol. 182:2928-2936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Livermore, D. M. 2000. Antibiotic resistance in staphylococci. Int. J. Antimicrob. Agents 16(Suppl. 1):S3-S10. [DOI] [PubMed] [Google Scholar]
- 6.Mazars, E., S. Lesjean, A. L. Banuls, M. Gilbert, V. Vincent, B. Gicquel, M. Tibayrenc, C. Locht, and P. Supply. 2001. High-resolution minisatellite-based typing as a portable approach to global analysis of Mycobacterium tuberculosis molecular epidemiology. Proc. Natl. Acad. Sci. USA 98:1901-1906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Miragaia, M., I. Couto, S. F. F. Pereira, K. G. Kristinson, H. Westh, J. O. Jarløv, J. Carriço, J. Almeida, I. Santos-Sanches, and H. de Lencastre. 2002. Molecular characterization of methicillin-resistant Staphylococcus epidermidis clones: evidence of geographic dissemination. J. Clin. Microbiol. 40:430-438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Monsen, T., H. Abd, K. Leonardsson, H. Edebro, and J. Wiström. 2002. Prediction of mecA-positive coagulase-negative staphylococci: assessment of different phenotypic methods, breakpoints, culture media and culture conditions. J. Antimicrob Chemother. 49:197-200. [DOI] [PubMed] [Google Scholar]
- 9.Monsen, T., C. Olofsson, M. Rönnmark, and J. Wiström. 2000. Clonal spread of staphylococci among patients with peritonitis associated with continuous ambulatory peritoneal dialysis. Kidney Int. 57:613-618. [DOI] [PubMed] [Google Scholar]
- 10.Monsen, T., M. Rönnmark, C. Olofsson, and J. Wiström. 1998. An inexpensive and reliable method for routine identification of staphylococcal species. Eur. J. Clin. Microbiol. Infect. Dis. 17:327-335. [DOI] [PubMed] [Google Scholar]
- 11.Ponnuraj, K., M. G. Bowden, S. Davis, S. Gurusiddappa, D. Moore, D. Choe, Y. Xu, M. Hook, and S. V. Narayana. 2003. A “dock, lock, and latch” structural model for a staphylococcal adhesin binding to fibrinogen. Cell 115:217-228. [DOI] [PubMed] [Google Scholar]
- 12.Sabat, A., J. Krzyszton-Russjan, W. Strzalka, R. Filipek, K. Kosowska, W. Hryniewicz, J. Travis, and J. Potempa. 2003. New method for typing Staphylococcus aureus strains: multiple-locus variable-number tandem repeat analysis of polymorphism and genetic relationships of clinical isolates. J. Clin. Microbiol. 41:1801-1804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Spare, M. K., S. E. Tebbs, S. Lang, P. A. Lambert, T. Worthington, G. W. Lipkin, and T. S. Elliott. 2003. Genotypic and phenotypic properties of coagulase-negative staphylococci causing dialysis catheter-related sepsis. J. Hosp. Infect. 54:272-278. [DOI] [PubMed] [Google Scholar]
- 14.Tenover, F. C., R. D. Arbeit, and R. V. Goering. 1997. How to select and interpret molecular strain typing methods for epidemiological studies of bacterial infections: a review for healthcare epidemiologists. Infect. Control Hosp. Epidemiol. 18:426-439. [DOI] [PubMed] [Google Scholar]
- 15.Tenover, F. C., R. D. Arbeit, R. V. Goering, P. A. Mickelsen, B. E. Murray, D. H. Persing, and B. Swaminathan. 1995. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 33:2233-2239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Titze-de-Almeida, R., R. J. Willems, J. Top, I. P. Rodrigues, R. F. Ferreira II, H. Boelens, M. C. Brandileone, R. C. Zanella, M. S. Felipe, and A. van Belkum. 2004. Multilocus variable-number tandem-repeat polymorphism among Brazilian Enterococcus faecalis strains. J. Clin. Microbiol. 42:4879-4881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.von Eiff, C., G. Peters, and C. Heilmann. 2002. Pathogenesis of infections due to coagulase-negative staphylococci. Lancet Infect. Dis. 2:677-685. [DOI] [PubMed] [Google Scholar]