

Abstract
Although 1,25-dihydroxyvitamin D3 (1,25D3) and retinoic acid (RA) have distinct developmental and physiological roles, both regulate the cell cycle. We provide molecular and genomic evidence that their cognate nuclear receptors regulate common genes through everted repeat TGA(C/T)TPyN8PuG(G/T)TCA (ER8) response elements. ER8 motifs were found in the promoters of several target genes of 1,25D3 and/or RA. Notably, an element was characterized in the cyclin-dependent kinase (CDK) inhibitor p19ink4d gene, and 1,25D3- or RA-induced p19INK4D expression. P19ink4d knockdown together with depletion of p27kip1, another CDK inhibitor regulated by 1,25D3 and RA, rendered cells resistant to ligand-induced growth arrest. Remarkably, p19INK4D-deficient cells showed increased autophagic cell death, which was markedly enhanced by 1,25D3, but not RA, and attenuated by loss of p27KIP1. These results show a limited crosstalk between 1,25D3 and RA signalling by means of overlapping nuclear receptor DNA binding specificities, and uncover a role for p19INK4D in control of cell survival.
Keywords: vitamin D, retinoic acid, nuclear receptors, autophagy, cross-talk, cyclin-dependent kinase inhibitors
Introduction
Retinoic acid (RA) and the hormonal form of vitamin D3 (1,25-dihydroxyvitamin D3; 1,25D3) signal through related nuclear receptors (Chawla et al, 2001). Both retinoic acid receptors (RARs) and the vitamin D receptor (VDR) heterodimerize with retinoid X receptors (RXRs). RAR/RXRs bind to retinoic acid response elements (RAREs) composed of direct repeats of PuG(G/T)TCA motifs separated by 1, 2 or 5 bp (DR1, DR2 or DR5), whereas vitamin D response elements (VDREs) are arranged as DR3 or weaker everted repeat (ER6) elements (Thompson et al, 2002). RAR signalling controls many aspects of embryonic development (Chambon 1996), whereas 1,25D3 is primarily known for its role in controlling mineral ion homeostasis. However, both 1,25D3 and RA regulate cell differentiation, proliferation and apoptosis (Freemantle et al, 2003; Lin & White, 2004), and show overlapping effects on the immune system (Stephensen, 2001; Lin & White, 2004).
A range of data has provided evidence for chemopreventive actions of 1,25D3 and its analogues, which show potent anticancer activities in in vitro and in vivo models (Lin & White, 2004). Similarly, vitamin A deficiency leads to squamous metaplasia of epithelia resembling early stages of carcinogenesis, and retinoids show chemotherapeutic and chemopreventive effects in cancers of the breast, liver and aerodigestive tract, and in acute promyelocytic leukaemia (Hong & Sporn, 1997; Freemantle et al, 2003).
We show here that ER8 elements function as VDREs and RAREs in vitro and in vivo, thus serving as a point of convergence of 1,25D3 and RA signalling. Scanning the human genome for promoter-proximal ER8 motifs showed elements in several previously identified 1,25D3 or RA target genes, including those encoding interleukin-10, the genotoxic stress-inducible factor DDIT3/GADD153/CHOP and the cyclin-dependent kinase (CDK) inhibitor p19INK4D, which are responsive to both 1,25D3 and RA. Moreover, our results indicate that induction of p19INK4D contributes to the cell-cycle regulatory properties of 1,25D3 and RA, and, remarkably, protects cells against autophagic cell death.
Results and Discussion
Identification of ER8 motifs as VDREs and RAREs
We identified 1,25D3 target genes in human SCC25 head and neck squamous cell carcinoma cells, the proliferation of which is arrested in G0/G1 by 1,25D3 and RA (Akutsu et al, 2001; Lin et al, 2002). In an analysis of 1,25D3 target genes, an everted repeat of PuG(G/T)TCA motifs separated by 8 bp (ER8) was identified in gene encoding transcription factor NRF2 (Fig 1A). Chromatin immunoprecipitation (ChIP) assays showed that the VDR bound to ER8 region of the nfr2 promoter, but not to adjacent DNA (Fig 1A). Significantly, ER8 motifs also function as RAREs (Tini et al, 1993), and ligand-dependent binding of RARs to the ER8 region of the nrf2 promoter was also detected by ChIP assay (Fig 1A).
Figure 1.
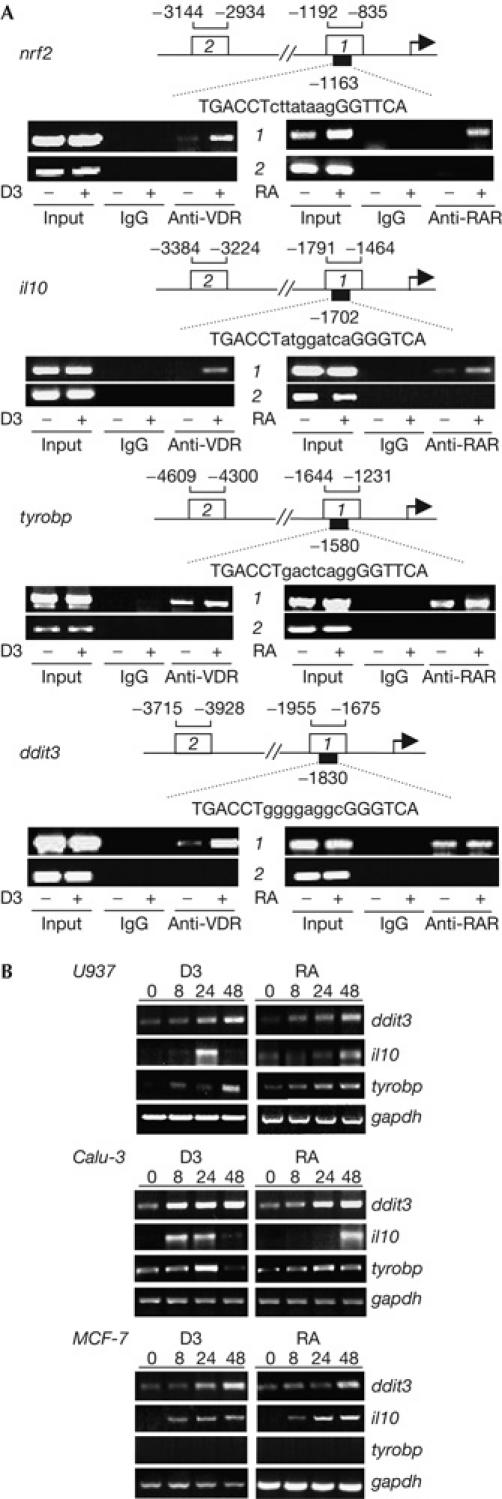
Identification of ER8 elements in 1,25-dihydroxyvitamin D3 and RA target genes. (A) Chromatin immunoprecipitation (ChIP) analysis of receptor binding to the ER8 element (region 1) of the nrf2 promoter. Immunoprecipitation with nonspecific IgG and PCR amplification of an adjacent region (2) served as controls. ChIP assays were also carried out to analyse binding to ER8s in the il10, ddit3 and tyrobp genes. RAR, retinoic acid receptor; VDR, vitamin D receptor. (B) Reverse transcription–PCR analysis of 1,25-dihydroxyvitamin D3 (1,25D3)- or retinoic acid (RA)-regulated expression of ddit3, il10, and tyrobp in U937, Calu-3 and MCF-7 cells.
We screened the human genome for ER8 motifs (Bourdeau et al, 2004; supplementary Table 1 online), and identified elements in several genes regulated by 1,25D3 and/or RA (supplementary Table 2 online). Apart from nrf2 (nfe2l2; element at −1163), these include genes encoding thioredoxin reductase 1 (txnrd1; −1378), ETS variant 4 (etv4/e1a-f; −4233), TYROBP/DAP12 (tyrobp; −1580) and E4F1 (e4f1; +3102), identified as 1,25D3-regulated on microarrays (Lin et al, 2002), and il10, which is induced by 1,25D3 in vivo (Cobbold et al, 2003). The gene encoding CDK inhibitor p19INK4D (cdkn2d; −245) was identified as RA-regulated on microarrays (Liu et al, 2000). RA is also an inducer of RUNX1 (AML1; runx1; −1662; Tanaka et al, 1995) and human cholesteryl ester transfer protein (cetp; +418; Florentin et al, 1996), and both RA and 1,25D3 induce expression of keratin 13 (krt13; −3339; van Rossum et al, 2000; Palmer et al, 2003) and DDIT3 (CHOP, GADD153, C/EBPζ; ddit3; −1830; Kim et al, 2002; Lin et al, 2002).
Notably absent are hox genes, which are regulators of cell fate and embryonic patterning. RA, but not 1,25D3, regulates several hox genes, and DR2- or DR5-type RAREs are present in several hox promoters (Gellon & McGinnis, 1998). The absence of ER8 motifs in hox genes is consistent with the notion that flexibility in response element recognition would permit partially overlapping DNA binding specificities of different receptors, and thus limited crosstalk between receptor signalling pathways.
We further examined il10 and tyrobp, which are not known to be regulated by RA, and ddit3, which is induced during growth arrest and in response to genotoxic and endoplasmic reticulum stress (Oyadomari & Mori, 2004). Interleukin-10, like 1,25D3, promotes immune system tolerance (Cobbold et al, 2003), and TYROBP is a transmembrane adaptor expressed throughout the immune system (Colonna, 2003). VDRs and RARs bind to ER8 regions of the il10, tyrobp and ddit3 promoters in a ligand-dependent manner (Fig 1A). Regulation of gene expression was tested in ligand-responsive U937 myelomonocytic, Calu-3 lung carcinoma and MCF-7 breast cancer cells. With the exception of tyrobp, which was not detected in MCF-7 cells, expression of il10, tyrobp and ddit3 was induced by 1,25D3 and RA in all cells (Fig 1B).
An ER8 element in the p19INK4D promoter
P19INK4D interacts selectively with CDK4, and its overexpression arrests cells in G0/G1 (Hirai et al, 1995). As we were interested in cell-cycle arrest by 1,25D3 and RA, we further analysed the regulation of p19ink4d and the function of its ER8 motif. Binding of VDR/RXRs to ER8 in vitro was analysed by electrophoretic mobility shift assay (EMSA) using the mouse osteopontin (mop) DR3 or the weaker cyp3A4 ER6 VDRE as controls (Fig 2A). VDR-specific complex formation on the ER8 motif was supershifted or partially disrupted by antibodies against VDRs and RXRs, respectively. VDR binding to the ER8 element was weaker than to the DR3 VDRE, but comparable with binding to the ER6 VDRE. Similar analysis of RAR binding using the rarβ DR5 RARE as a control (Fig 2B) confirmed that RAR/RXRs bound strongly to the ER8 motif. Ligand-dependent binding of VDRs and RARs to the p19ink4d promoter was confirmed in vivo, and treatment with either ligand enhanced the interaction of RNA polymerase II with the initiation site of the p19ink4d promoter (Fig 2C). ReChIP experiments showed that receptors bound as heterodimers with RXRs, whereas no evidence was found for binding of VDR/RAR heterodimers to ER8 (Fig 2D). Moreover, the integrity of the ER8 motif was essential for 1,25D3- or RA-dependent induction of a p19ink4d promoter-luciferase reporter (Fig 2E–G), confirming the function of the element. Finally, transfection of RARs α, β or γ produced similar fold inductions of reporter activity (supplementary Fig 1 online), and no evidence for synergism between RA and 1,25D3 was found (data not shown).
Figure 2.
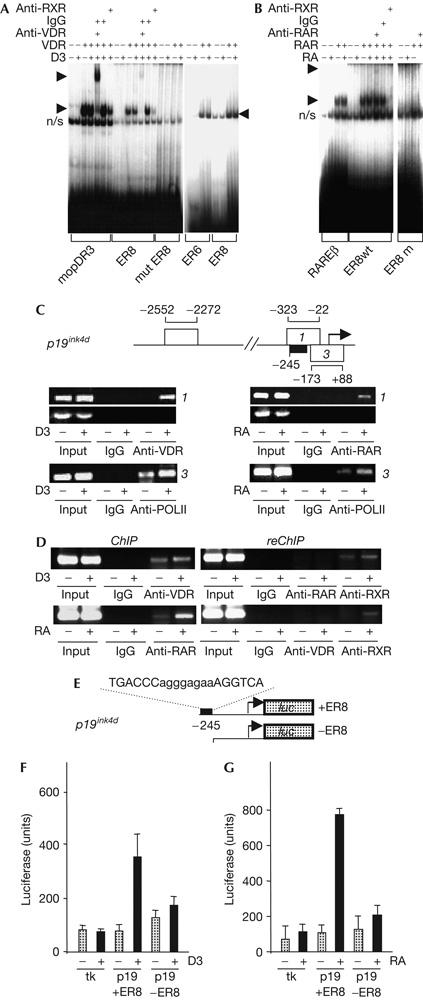
Characterization of in vitro and in vivo p19ink4d ER8 element function. (A) Electrophoretic mobility shift assay (EMSA) of binding of vitamin D receptors (VDRs)/retinoid X receptors (RXRs) to the p19ink4d ER8, with binding to the mouse osteopontin (mop) DR3 vitamin D responsive element as a control, along with a comparison of binding to ER6 and ER8 motifs (right-hand panel). (B) EMSA of retinoic acid receptor (RAR)/RXR binding to the p19ink4d ER8, and the rarβ DR5 retinoic acid response element, as a control. (C) Chromatin immunoprecipitation (ChIP) analysis of binding of VDRs and RARs to the p19ink4d ER8 (region 1) and RNA polymerase II (POLII) binding to the transcription start site. (D) ReChIP analysis confirms the presence of VDR/RXRs and RAR/RXRs but not of VDR/RARs on the p19ink4d ER8. (E) Cloning of p19ink4d promoter containing or lacking ER8 upstream of a luciferase reporter. 1,25-Dihydroxyvitamin D3 (1,25D3)- (F) and retinoic acid (RA)-regulated (G) luciferase expression is dependent on the ER8 element in the p19ink4d promoter. Luciferase expression driven by a thymidine kinase (tk) control promoter is shown.
Rapid induction of p19ink4d messenger RNA was observed in four 1,25D3- and RA-sensitive cell lines (Fig 3A), and treatment with either ligand increased p19INK4D protein levels in U937, SCC25 or Calu-3 cells in 48 h (Fig 3B; data not shown). This coincided with a transient increase in the association of p19INK4D with CDK4 in U937 and SCC25 cells, although levels of CDK4 were not affected (Fig 3C; data not shown). In contrast, a more gradual and sustained increase in p27KIP1/CDK4 complexes was observed in 1,25D3-treated cells (Fig 3D), which suggested that the two CDK inhibitors function cooperatively.
Figure 3.
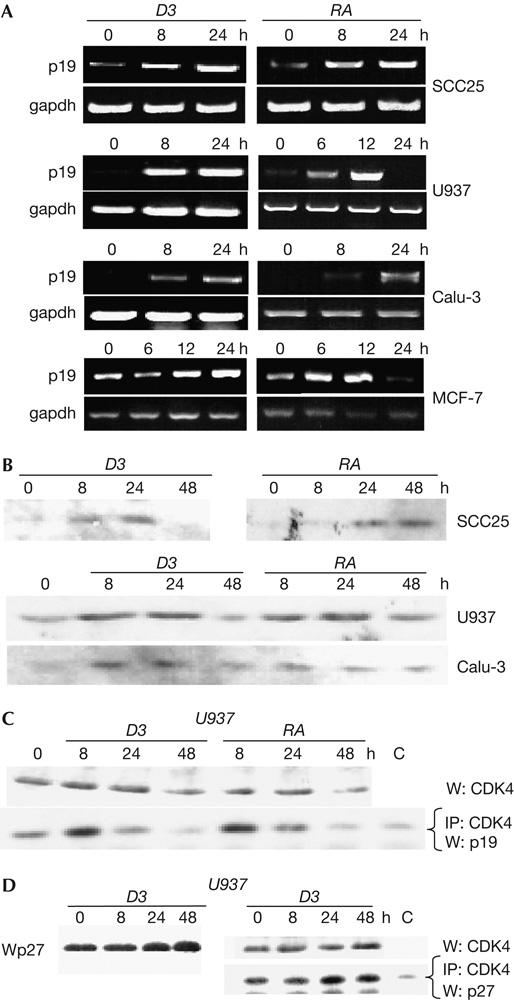
1,25-dihydroxyvitamin D3 and retinoic acid induce p19ink4d expression. Expression of p19ink4d messenger RNA by reverse transcription–PCR (A) or protein by western blotting (B) was carried out on extracts of cells treated with 1,25-dihydroxyvitamin D3 (1,25D3) or retinoic acid (RA), as indicated. (C) Treatment with 1,25D3 or RA transiently enhances co-immunoprecipitation (IP) of p19INK4D with cyclin-dependent kinase (CDK)4 in U937 cells. Similar results were obtained in SCC25 cells (not shown). A control western blot (W) of CDK4 levels in ligand-treated cells is shown below. (D) Treatment with 1,25D3 enhances co-immunoprecipitation of p27KIP1 with CDK4 in U937 cells. Extracts of U937 cells were probed for p27KIP1 and CDK4, and immunoprecipitated with an anti-CDK antibody and probed for p27KIP1 co-immunoprecipitation.
SiRNA knockdown of p19INK4D and p27KIP1
We were interested in determining whether induction of p19INK4D contributed to cell-cycle regulation by 1,25D3 and RA, and whether it cooperated with p27KIP1, another 1,25D3- and RA-induced CDK inhibitor. P19INK4D expression was knocked down by short interfering RNAs (siRNAs) to test its role in 1,25D3- and RA-regulated SCC25 cell proliferation (Akutsu et al, 2001). SCC25 cells were transfected with siRNAs with almost 100% efficiency (supplementary Fig 2 online), and complete p19INK4D knockdown was achieved, whereas no effect of either scrambled RNA or cyclophilin-specific siRNA was observed (Fig 4A; data not shown). Complete knockdown of p27KIP1 also occurred, alone and in combination with p19INK4D (Fig 4B). 1,25D3 treatment of cells transfected with scrambled siRNA reduced cells in S phase and increased cells in sub-G0/G1 (Fig 4C), suggestive of cytotoxicity. Remarkably, p19INK4D knockdown substantially reduced cell survival (supplementary Fig 3A online) and increased the sub-G0/G1 fraction (Fig 4C), an effect that was markedly enhanced by 1,25D3, indicating that p19INK4D expression protects cells from (1,25D3-induced) cell death. Surviving p19INK4D-depleted cells treated with 1,25D3 showed enhanced Trypan blue uptake, consistent with decreased viability (supplementary Fig 3B online). In contrast, p27KIP1 knockdown reduced the G0/G1 fraction, slightly enhanced cells in S and substantially increased cells in G2/M (Fig 4C), effects that were modestly reversed by 1,25D3. Combined loss of p19INK4D and p27KIP1 produced a cell-cycle distribution that was similar to that of p27KIP1-depleted cells (Fig 4C), indicating that p27KIP1 loss reverses the sensitivity to cell death caused by p19INK4D depletion. Although 1,25D3 treatment somewhat reduced cells in G2/M, the proportion of p19INK4D/p27KIP1-depleted cells in S phase remained elevated, consistent with a role for the two CDK inhibitors in 1,25D3-induced G0/G1 arrest. In contrast, RA treatment of p19INK4D-depleted cells substantially reduced the sub-G0/G1 fraction (Fig 4D). However, there was double the proportion of cells in S phase relative to controls, consistent with p19ink4d induction contributing to RA-induced cell-cycle arrest. Similarly, the proportion of cells in S phase remained elevated in RA-treated cells depleted for p27KIP1, or both p19INK4D and p27KIP1 (Fig 4D).
Figure 4.
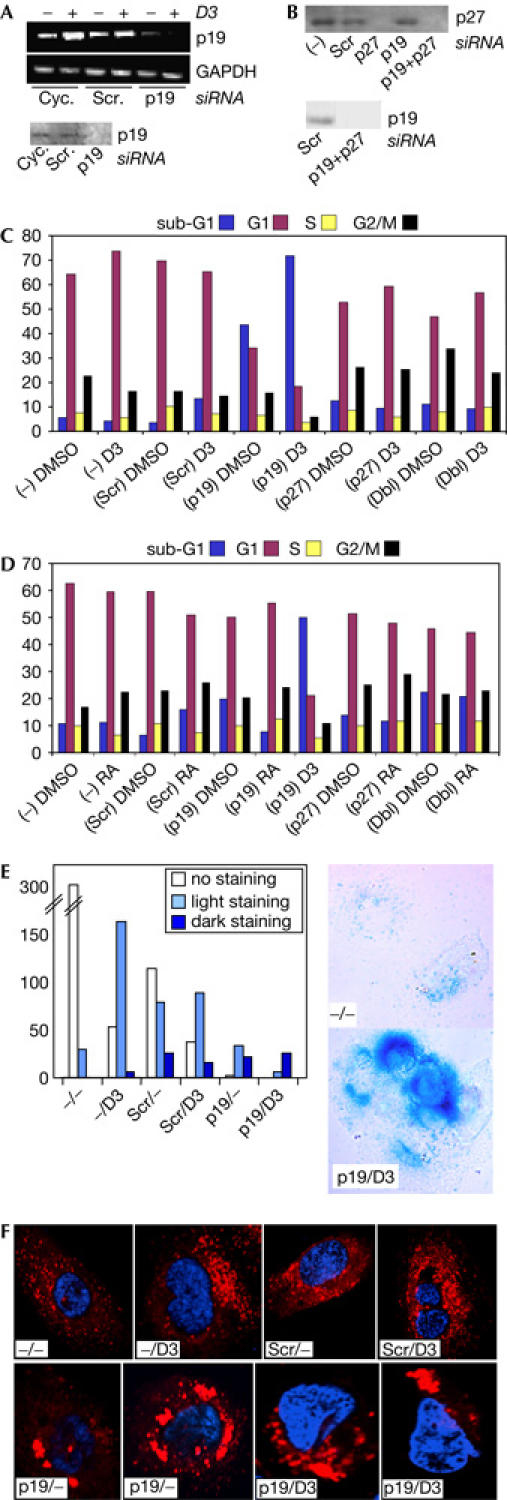
Knockdown of p19ink4d and p27kip1 in SCC25 cells. (A) Short interfering RNAs (siRNA) knockdown of p19ink4d messenger RNA and protein in SCC25 cells. SiRNAs were directed against p19ink4d, scrambled p19ink4d (Scr.) or cyclophilin (cyc.) controls. (B) Combined knockdown of p27KIP1 and p19INK4D in SCC25 cells analysed by western blotting for p27KIP1 (top) and p19INK4D (bottom). (C,D) Fluorescence-activated cell sorting analysis of cell-cycle distribution of control SCC25 cells, or cells transfected with scrambled (Scr), p19ink4d- or p27kip1-directed siRNAs, individually or together (Dbl). (C) Cells were treated with vehicle or 1,25-dihydroxyvitamin D3 (1,25D3), as indicated. (D) Cells were treated with DMSO vehicle, retinoic acid (RA) or 1,25D3, as indicated. (E) Loss of p19INK4D enhances lysosomal β-galactosidase activity in SCC25 cells. (Left) Histogram of numbers of cells staining for β-galactosidase activity in the absence (−) or presence of 1,25D3 in control cells (−) or in cells transfected with scrambled (Scr) or p19INK4D siRNAs. (Right) Bright field images of untransfected cells (−/−) or p19INK4D-depleted cells treated with 1,25D3 (p19/D3). (F) Loss of p19INK4D induces lysosomal fusion. Analysis of lysosomal fusion (autophagosomes) in SCC25 cells treated as in (E), stained for incorporation of lysotracker red. Nuclei were counterstained with Hoechst (blue).
Cell death observed in p19INK4D-depleted cells was not accompanied by enhanced caspase-3 cleavage or annexin V staining (supplementary Fig 4 online; data not shown), indicating that cell death was not apoptotic. No β-galactosidase activity characteristic of senescence (pH 6.0; Dimri et al, 1995) was detected (data not shown). However, as cell shrinkage and cytoplasmic vacuolization were observed, we investigated whether cells were dying by autophagy. The combination of p19INK4D loss and 1,25D3 treatment markedly enhanced the proportion of cells expressing high levels of lysosomal β-galactosidase activity (pH 4.0; Fig 4E). Moreover, although 1,25D3 treatment or siRNA transfection tended to increase lysosomal numbers in SCC25 cells, as judged by lysotracker red incorporation, vacuolization was only observed in p19INK4D-depleted cells (Fig 4F). The effect, however, is cell specific, as p19INK4D knockdown in U937 cells neither significantly affected cell viability (data not shown) nor ligand-induced U937 differentiation (supplementary Fig 5 online), an unexpected result given the induction of macrophage differentiation by p19INK4D overexpression in immature myeloid cells (Adachi et al, 1997).
Conclusions
We found that 1,25D3 and RA regulate common target genes, including p19ink4d, by recognizing ER8 motifs as response elements. Induction of p19ink4d contributes to cell-cycle regulation by 1,25D3 and RA, and emphasizes the overlapping effects of 1,25D3 and RA on cell-cycle regulators. Both enhance expression of p19INK4D and p27KIP1, although by distinct mechanisms. 1,25D3 or RA treatment increases p27KIP1 protein expression by repressing levels of transcripts encoding p45SKP2, a ubiquitin ligase that targets KIP1 for proteasomal degradation (Dow et al, 2001; Lin et al, 2003). We found that cells lacking both p19INK4D and p27KIP1 are refractory to 1,25D3 and RA, and that the two CDK inhibitors cooperate in ligand-mediated cell-cycle regulation. Cooperation between p19INK4D and p27KIP1 is supported by genetic studies, where they were combined to control mouse postnatal neuronal (Zindy et al, 1999) and retinal cell proliferation (Cunningham et al, 2002).
Remarkably, loss of p19ink4d rendered cells more sensitive to autophagic cell death, an effect that was markedly enhanced by 1,25D3, but not by RA. These results are consistent with autophagic cell death observed in 1,25D3-treated MCF-7 cells (Hoyer-Hansen et al, 2005), and show a role for p19INK4D induction in protecting cells from death induced by autophagy. These results are also noteworthy in the light of observations that induction of p19INK4D confers resistance to UV-induced apoptosis, and, importantly, enhances DNA repair (Ceruti et al, 2005). Induction of p19INK4D may thus be one of the components of the protective effects of 1,25D3 and RA against UV damage in epithelia, and would, in part, explain their chemopreventive properties. We also found that p19INK4D-depleted SCC25 cells were hypersensitive to UV-induced apoptosis, as measured by annexin V and propidium iodide co-staining (unpublished results).
The autophagic phenotype of p19INK4D depletion is striking, given the contrast with the hyperproliferative response in cells lacking other INK4 proteins (Bond et al, 2004). Gene knockouts have not supported a role for p19ink4d in tumour suppression. However, combined loss of p19INK4D and p27KIP1 in our study did not result in autophagy, but led to resistance to ligand-induced cell-cycle arrest. Combined loss of p19INK4D and p27KIP1 in mice led to postnatal death associated with neuronal defects (Zindy et al, 1999), thus preventing analyses of long-term effects of combined depletion on cancer susceptibility. It therefore remains possible that loss of both CDKs could promote aberrant cell proliferation during tumorigenesis.
Methods
Details of cell culture, plasmids, transfections, ChIP assays, cell-cycle analysis, primers and antibodies used are found in the supplementary information online.
Response element screening. Screening was carried out as described by Bourdeau et al (2004) using NCBI fasta and gbs files of the Human genome reference assembly (Build 35 version 1; 26 August 2004) to search for specified sequences and extract the positions of matching motifs in the genome contigs, as well as the coordinates of the surrounding genes within a preset cutoff distance of each motif.
Short interfering RNA knockdowns. SCC25 cells, grown in 6 cm wells to a density of 2 × 105 cells/well, were transfected in OPTI-MEM (Invitrogen, Burlington, Ontario, Canada) with Oligofectamine (Invitrogen) and 100 nM of scrambled, p19ink4d or p27kip1 siRNAs (Dharmacon, Lafayette, CO, USA). DMEM/F12 with 30% fetal bovine serum was added 6 h after transfection. After 24 h, the medium was replaced by DMEM/F12 with 10% fetal bovine serum for 24 h, followed by a medium containing charcoal-stripped serum (10%) and DMSO, 1,25D3 (100 nM) or RA (1 μM) for 48 h. Cells were collected for fluorescence-activated cell sorting (FACS) analysis.
Microscopy. Cells were stained for lysosomal β-galactosidase (Dimri et al, 1995), and bright field digital images were captured using a Zeiss Axioplan 2 microscope, equipped with motorized stage and focus, and Zeiss AxioCamHRC digital camera coupled to AxioVision 4 software (Zeiss, Canada). Confocal microscopy was carried out using a Zeiss LSM 510 microscope with a two-photon titanium:sapphire laser. Samples were stained with lysotracker red DND99 and counterstained with Hoechst dye (Molecular Probes, Eugene, OR, USA).
Supplementary information is available at EMBO reports online (http://www.nature.com/embor/journal/vaop/ncurrent/extref/7400594-s1.pdf).
Supplementary Material
Supplementary Information
Acknowledgments
We are grateful to Dr D. Maysinger, J. Laliberté and K. Bell for help with cytochemistry and confocal microscopy, and to K. MacDonald for help with FACS. This work was supported by grants from the Canadian Institutes of Health Research (CIHR) to J.H.W. and from the Natural Sciences and Engineering Research Council to S.M. J.H.W. and S.M. are Chercheurs-Boursier of the Fonds de Recherché en Santé du Québec. L.T.M. was supported by a training grant from the CIHR. V.B. was supported by Genome Canada and the CIHR. S.M. is the holder of a Canadian Imperial Bank of Commerce Breast Cancer Research Chair.
References
- Adachi M, Roussel MF, Havenith K, Sherr CJ (1997) Features of macrophage differentiation induced by p19INK4D, a specific inhibitor of cyclin D-dependent kinases. Blood 90: 126–137 [PubMed] [Google Scholar]
- Akutsu N, Lin R, Bastien Y, Bestawros A, Enepekides DJ, Black MJ, White JH (2001) Regulation of gene expression by 1α,25-dihydroxyvitamin D3 and its analog EB1089 under growth inhibitory conditions in squamous carcinoma cells. Mol Endocrinol 15: 1127–1139 [DOI] [PubMed] [Google Scholar]
- Bond J, Jones C, Haughton M, DeMicco C, Kipling D, Wynford-Thomas D (2004) Direct evidence from siRNA-directed ‘knock down' that p16(INK4a) is required for human fibroblast senescence and for limiting ras-induced epithelial cell proliferation. Exp Cell Res 292: 151–156 [DOI] [PubMed] [Google Scholar]
- Bourdeau V, Deschenes J, Metivier R, Nagai Y, Nguyen D, Bretschneider N, Gannon F, White JH, Mader S (2004) Genome-wide identification of high affinity estrogen response elements in human and mouse. Mol Endocrinol 18: 1411–1427 [DOI] [PubMed] [Google Scholar]
- Ceruti JM, Scassa ME, Flo JM, Varone CL, Canepa ET (2005) Induction of p19INK4d in response to ultraviolet light improves DNA repair and confers resistance to apoptosis in neuroblastoma cells. Oncogene 24: 4065–4080 [DOI] [PubMed] [Google Scholar]
- Chambon P (1996) A decade of molecular biology of retinoic acid receptors. FASEB J 10: 940–954 [PubMed] [Google Scholar]
- Chawla A, Repa J, Evans RM, Mangelsdorf DJ (2001) Nuclear receptors and lipid physiology: opening the X-files. Science 294: 1866–1870 [DOI] [PubMed] [Google Scholar]
- Cobbold SP et al. (2003) Regulatory T cells and dendritic cells in transplantation tolerance: molecular markers and mechanisms. Immunol Rev 196: 109–124 [DOI] [PubMed] [Google Scholar]
- Colonna M (2003) Trems in the immune system and beyond. Nat Rev Immunol 3: 445–453 [DOI] [PubMed] [Google Scholar]
- Cunningham JJ, Levine EM, Zindy F, Goloubeva O, Roussel MF, Smeyne RJ (2002) The cyclin-dependent kinase inhibitors p19(Ink4d) and p27(Kip1) are coexpressed in select retinal cells and act cooperatively to control cell cycle exit. Mol Cell Neurosci 19: 359–374 [DOI] [PubMed] [Google Scholar]
- Dimri GP et al. (1995) A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci USA 92: 9363–9367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dow R, Hendley J, Pirkmaier A, Musgrove EA, Germain D (2001) Retinoic acid-mediated growth arrest requires ubiquitylation and degradation of the F-box protein Skp2. J Biol Chem 276: 45945–45951 [DOI] [PubMed] [Google Scholar]
- Florentin E, Athias A, Lagrost L (1996) Modulation of the activity of the human cholesteryl ester transfer protein by carboxylated derivatives—evidence for 13-cis-retinoic acid as a potent activator of the protein's activity in plasma. Eur J Biochem 240: 699–706 [DOI] [PubMed] [Google Scholar]
- Freemantle SJ, Spinella MJ, Dmitrovsky E (2003) Retinoids in cancer therapy and chemoprevention: promise meets resistance. Oncogene 22: 7305–7315 [DOI] [PubMed] [Google Scholar]
- Gellon G, McGinnis W (1998) Shaping animal body plans in development and evolution by modulation of Hox expression patterns. Bioessays 20: 116–125 [DOI] [PubMed] [Google Scholar]
- Hirai H, Roussel M, Kato JY, Ashmun RA, Sherr CJ (1995) Novel INK4 proteins p18 and p19 are specific inhibitors of the cyclin D-dependent kinases CDK4 and CDK6. Mol Cell Biol 15: 2762–2781 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong WK, Sporn MB (1997) Recent advances in chemoprevention of cancer. Science 278: 1073–1077 [DOI] [PubMed] [Google Scholar]
- Hoyer-Hansen M, Bastholm L, Mathiasen IS, Elling F, Jaattela M (2005) Vitamin D analog EB1089 triggers dramatic lysosomal changes and beclin 1-mediated autophagic cell death. Cell Death Differ 12: 1297–1309 [DOI] [PubMed] [Google Scholar]
- Kim DG, You KR, Liu MJ, Choi YK, Won YS (2002) GADD153-mediated anticancer effects of N-(4-hydroxyphenyl)retinamide on human hepatoma cells. J Biol Chem 277: 38930–38938 [DOI] [PubMed] [Google Scholar]
- Lin R, White JH (2004) The pleiotropic actions of vitamin D. Bioessays 26: 21–28 [DOI] [PubMed] [Google Scholar]
- Lin R, Nagai Y, Sladek R, Bastien Y, Ho J, Petrecca K, Sotiropoulou G, Diamandis EP, Hudson TJ, White JH (2002) Expression profiling in squamous carcinoma cells reveals pleiotropic effects of vitamin D3 signaling on cell proliferation, differentiation and immune system regulation. Mol Endocrinol 16: 1243–1256 [DOI] [PubMed] [Google Scholar]
- Lin R, Wang TT, Miller WH, White JH (2003) Inhibition of F-box protein p45SKP2 expression and stabilization of cyclin-dependent kinase inhibitor p27KIP1 in vitamin D analogue-treated cancer cells. Endocrinology 144: 749–753 [DOI] [PubMed] [Google Scholar]
- Liu TX et al. (2000) Gene expression networks underlying retinoic acid-induced differentiation of acute promyelocytic leukemia cells. Blood 96: 1496–1504 [PubMed] [Google Scholar]
- Oyadomari S, Mori M (2004) Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ 11: 381–389 [DOI] [PubMed] [Google Scholar]
- Palmer HG, Sanchez-Carbayo M, Ordonez-Moran P, Larriba MJ, Cordn-Cardo C, Munoz A (2003) Genetic signatures of differentiation induced by 1α,25-dihydroxyvitamin D-3 in human colon cancer cells. Cancer Res 63: 7799–7806 [PubMed] [Google Scholar]
- Stephensen CB (2001) Vitamin A, infection, and immune function. Annu Rev Nutr 21: 167–192 [DOI] [PubMed] [Google Scholar]
- Tanaka K, Tanaka T, Ogawa S, Kurokawa M, Mitani K, Yazaki Y, Hirai H (1995) Increased expression of aml1 during retinoic-acid-induced differentiation of U937 cells. Biochem Biophys Res Commun 211: 1023–1030 [DOI] [PubMed] [Google Scholar]
- Thompson PD, Jurutka PW, Whitfield GK, Myskowski SM, Eichhorst KR, Dominguez CE, Haussler CA, Haussler MR (2002) Liganded VDR induces CYP3A4 in small intestinal and colon cancer cells via DR3 and ER6 vitamin D responsive elements. Biochem Biophys Res Commun 299: 730–738 [DOI] [PubMed] [Google Scholar]
- Tini M, Otulakowski G, Breitmen ML, Tsui LC, Giguère V (1993) An everted repeat mediates retinoic acid induction of the γ-F-crystallin gene—evidence of a direct role for retinoids in lens development. Genes Dev 7: 295–307 [DOI] [PubMed] [Google Scholar]
- van Rossum MM, Mommers JM, van de Kerkhof PCM, van Erp PEJ (2000) Coexpression of keratins 13 and 16 in human keratinocytes indicates association between hyperproliferation-associated and retinoid-induced differentiation. Arch Dermatol Res 292: 16–20 [DOI] [PubMed] [Google Scholar]
- Zindy F, Cunningham JJ, Sherr CJ, Jogal S, Smeyne RJ, Roussel MF (1999) Postnatal neuronal proliferation in mice lacking Ink4d and Kip1 inhibitors of cyclin-dependent kinases. Proc Natl Acad Sci USA 96: 13462–13467 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Information