

Abstract
Small-molecule antagonists of Hedgehog-pathway signaling, such as cyclopamine, have been known for some time. Now, small-molecule agonists of the Hedgehog pathway have also been identified. The finding that both antagonists and agonists target the protein Smoothened supports the emerging hypothesis that Smoothened may be regulated by endogenous small molecules.
Thirty years before genetic experiments in Drosophila created the hedgehog mutant [1], a naturally occurring 'chemical genetic' experiment had produced sheep with an even more disturbing phenotype: cyclopia. The lack of midline facial structures in the offspring of grazing sheep in the western United States was attributed to ingestion of the lily Veratrum californicum, and subsequent work identified the jervine family of steroidal alkaloids, including the compound cyclopamine, as the teratogens responsible for the striking effects [2]. Insights into a possible mechanism of action did not emerge until the mid 1990s, when it was discovered that mutation of the Sonic hedgehog (Shh) gene in mice [3] or humans [4] could produce defects that resembled those caused by administration of cyclopamine to animals. Shh is a secreted protein ligand that, like other members of the Hedgehog (Hh) family, activates the Hh signal transduction pathway, and plays an important role in patterning many tissues[5]. The similarity in phenotypes suggested that teratogens might induce cyclopia by antagonizing the Hh pathway, and this hypothesis was confirmed when it was found that cyclopamine could directly block the response of tissues to Shh without interfering with the generation or processing of the Shh ligand [6,7].
How does cyclopamine block the response of cells to Shh? An exciting answer to this question has just emerged from the Beachy lab [8], which has identified the target of cyclopamine as the protein Smoothened (Smo), a protein with seven transmembrane domains that is distantly related to G-protein coupled receptors (GPCRs) [5]. In unstimulated cells, the activity of Smo is somehow repressed by the protein Patched (Ptc), which appears to be the receptor for the Shh ligand. When Ptc is engaged by Shh, Smo is activated and stimulates transcription factors of the Cubitus interruptus (Ci) or Gli family to induce the expression of specific genes. How Smo activates these transcription factors also remains unclear: although Smo has distant homology to GPCRs, no G protein has yet been identified as essential for Hh-pathway signaling.
Like other GPCRs, however, it now appears that Smo can be activated by small molecules. In this issue of the Journal of Biology, Jeff Porter and colleagues [9] at Curis Inc. report the identification of a class of synthetic small molecules (see Figure 1a) that potently activate the Hh-signaling pathway by binding to the Smo protein. I will refer to this chemical class of agonists as 'leiosamines' (from the Greek leios, meaning smooth), to reflect their ability to target the Smo protein. In addition to having important therapeutic implications (see the article by Stecca and Ruiz i Altaba in this issue [10]), the discovery of leiosamine also supports the exciting new idea that endogenous small molecules may regulate Smo activity in vivo[11].
Figure 1.
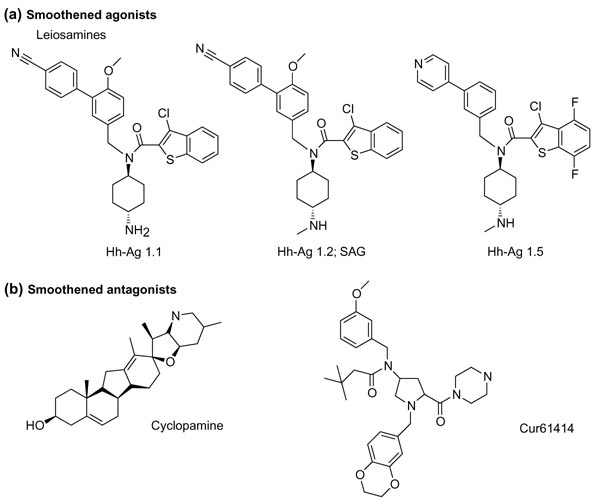
The structures of small molecules that activate or inhibit Hedgehog (Hh) signaling. (a) The leiosamine family of compounds that activate Hh signaling by binding to Smoothened. Hh-Ag (Hedgehog agonist) 1.1 was the original compound identified in the high-throughput screen by Frank-Kamenetsky et al.[9], with an EC50 of 3 μM in their luciferase reporter assay. Hh-Ag 1.2 is a more potent derivative that is also characterized by Chen et al.[17], who refer to it as SAG for 'synthetic Hh agonist'. Hh-Ag 1.5 is the most potent Hh agonist reported [9], with an E50 of 1 nM. (b) The structures of two compounds that bind to Smoothened to inhibit Hh signaling, cyclopamine and Cur61414. Structurally distinct classes of Smoothened antagonists have also been reported [17] but are not shown here.
The distinct routes by which cyclopamine and leiosamine were discovered provide an interesting glimpse into the changing nature of small-molecule discovery in biology and medicine. Cyclopamine was discovered through a natural accident, whereas leiosamine was discovered by systematic screening of chemical libraries using a cell-based assay that monitored Hh-pathway activation. The goal of screening against an entire pathway, rather than taking a more targeted approach, is to let the pathway reveal which components are most sensitive to pharmacological perturbation. Because this type of broad phenotypic screening emulates traditional genetics it has been referred to as "chemical genetics" [12-14]. The approach is ambitious and not without risk, because real biological insight (and perhaps approval by the US Food and Drug Administration of any resulting drug) requires identification of the target protein. The challenging nature of target identification is a key reason that many drug companies stick to screening isolated target proteins. But the recent success in identifying Smo as the target of cyclopamine and leiosamine suggests that the challenge of target identification can be overcome, and that pathway-based screening strategies can yield valuable dividends. In the remainder of this article, I will summarize how the Smo protein was identified as a receptor for small molecules, and the implications of this finding for understanding the mechanism of Hh signal transduction.
The target of cyclopamine was not discovered blindly; instead, several clues pointed to Smo as a possible target. The Beachy group had previously shown that cyclopamine remained competent to block Hh signaling even in cell lines in which signaling was constitutively activated by loss-of-function mutations in Ptc, placing the site of cyclopamine action downstream of Ptc [15]. Cyclopamine could also block signaling by constitutively activated oncogenic mutant forms of Smo, although higher concentrations were required, highlighting the possibility that Smo might be a target. To test this hypothesis, Chen et al.[8] synthesized a radiolabeled version of cyclopamine containing a photo-activatable crosslinker. Transfection of cells with a tagged version of Smo allowed detection of specific crosslinking to Smo that could be specifically competed by unlabeled cyclopamine. Cells transfected with Smo were also shown to bind specifically to a fluorescent derivative of cyclopamine, and by expressing truncation mutants of Smo, Chen et al. showed that cyclopamine binding requires only the seven transmembrane-spanning domains of Smo. Taken together, these findings strongly suggest that cyclopamine inhibits Hh signal transduction by binding to Smo.
The Curis group was interested in identifying small molecules that could activate the Hh pathway in the absence of the Shh ligand. Frank-Kamenetsky et al.[9] first established a mouse cell line that upregulated luciferase expression in response to addition of Shh protein. Screening of 140,000 compounds for the ability to activate luciferase expression in the absence of Shh protein led to the identification of the leiosamine family of compounds. Medicinal chemistry efforts improved the potency of this class of compounds by a thousand-fold, with significant enhancement of activity after alkylation of a basic nitrogen. The authors demonstrated that leiosamine is capable of stimulating the expression of Hh-specific genes in cells and also of activating appropriate Hh-dependent biological responses, such as proliferation of neonatal cerebellar granule neurons. In an assay using an explant of the developing chick neural plate, leiosamine was shown to induce dose-dependent changes in cell fate, as would be expected for a Hh-pathway agonist.
Even more excitingly, one leiosamine derivative was able to activate Ptc expression (a target of Hh-pathway activation) in mouse embryos after oral administration to pregnant females [9]. In an elegant set of experiments, the Curis group took advantage of this activity of leiosamine in vivo to narrow the range of possible targets. Mouse embryos with homozygous mutations in the Shh gene normally fail to express Hh-pathway target genes such as Ptc. Treatment of pregnant mothers carrying Shh-/- embryos with leiosamine was shown to restore Ptc expression, indicating that leiosamine can activate the Hh pathway in the absence of Shh ligand. Signaling was not restored in Smo-/- embryos, however, indicating that Smo is essential for the signaling that is activated by leiosamine.
Additional experiments in cell culture also hinted that Smo might be a target of leiosamine. Forskolin, a downstream inhibitor of the Hh pathway, was found to inhibit leiosamine-stimulated signaling as well as signaling activated by addition of Shh protein. But cyclopamine inhibited leiosamine-stimulated signaling less than it inhibited Shh-driven signaling, suggesting that cyclopamine and leiosamine might target the same receptor. To test the hypothesis that Smo is the biochemical target of leiosamine, the authors prepared a tritiated version of leiosamine and used it to treat cells that overexpressed an epitope-tagged version of Smo. The authors were able to immunoprecipitate a substantial fraction of the input radioactive counts only in cell lines in which Smo was overexpressed, and this binding could be competed by addition of unlabeled leiosamine or cyclopamine. Another novel antagonist of the Hh pathway, Cur61414 ([16]; Figure 1b), was also found to compete with leiosamine for binding, suggesting that it also targets Smo. Equilibrium binding measurements using membranes containing Smo protein indicated that the best leiosamine derivative bound to wild-type Smo with a dissociation constant (Kd) of 0.37 nM. Interestingly, when this experiment was repeated using membranes containing the activated mutant of Smo, the strength of leiosamine binding was not affected, whereas binding of antagonists was substantially weakened. To explain this observation, the authors propose that leiosamine and cyclopamine bind to separate sites on Smo, but that negative cooperativity between the sites results in antagonistic binding.
The Beachy lab has also recently characterized leiosamine and has confirmed the Smo protein as its target using photo-affinity crosslinking experiments with a radiolabeled version of leiosamine [17]. Competition experiments with fluorescently labeled cyclopamine suggest that the transmembrane heptahelical bundle of Smo is sufficient for leiosamine binding. Interestingly, higher concentrations of leiosamine are inhibitory to Hh signaling, suggesting that the compound may also interact with a downstream effector protein that becomes titrated away from Smo at high concentrations of ligand. The identity of this effector is unknown, but perhaps leiosamine will be a useful tool in itsidentification.
The susceptibility of Smo to activation or inhibition by synthetic small molecules suggests the intriguing possibility that endogenous small molecules may also regulate Smo activity. But how does this idea fit with current thinking about Hh-pathway signal transduction? Early models proposed that Ptc might regulate Smo by forming a stoichiometric complex. But recent work indicates that Ptc can inhibit Smo at sub-stoichiometric levels [11]. Ptc has homology to a family of bacterial transmembrane proteins that can transport small molecules, and inactivating mutations in Ptc are similar to those known to abolish transport activity in bacteria. It has therefore been proposed that Ptc may transport a small molecule that binds Smo and regulates its activity [11]. Now that we know that Smo can be either activated or inhibited by synthetic small molecules, the discovery of the natural endogenous counterparts is eagerly anticipated.
References
- Nusslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature. 1980;287:795–801. doi: 10.1038/287795a0. [DOI] [PubMed] [Google Scholar]
- Keeler RF, Binns W. Teratogenic compounds of Veratrum californicum V. Comparison of cyclopian effects of steroidal alkaloids from the plant and structurally related compounds from other sources. Teratology. 1968;1:5–10. doi: 10.1002/tera.1420010103. [DOI] [PubMed] [Google Scholar]
- Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, Beachy PA. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature. 1996;383:407–413. doi: 10.1038/383407a0. [DOI] [PubMed] [Google Scholar]
- Roessler E, Belloni E, Gaudenz K, Jay P, Berta P, Scherer SW, Tsui LC, Muenke M. Mutations in the human Sonic Hedgehog gene cause holoprosencephaly. Nat Genet. 1996;14:357–360. doi: 10.1038/ng1196-357. [DOI] [PubMed] [Google Scholar]
- Ingham PW, McMahon AP. Hedgehog signaling in animal development: paradigms and principles. Genes Dev. 2001;15:3059–3087. doi: 10.1101/gad.938601. [DOI] [PubMed] [Google Scholar]
- Cooper MK, Porter JA, Young KE, Beachy PA. Teratogen-mediated inhibition of target tissue response to Shh signaling. Science. 1998;280:1603–1607. doi: 10.1126/science.280.5369.1603. [DOI] [PubMed] [Google Scholar]
- Incardona JP, Gaffield W, Kapur RP, Roelink H. The teratogenic Veratrum alkaloid cyclopamine inhibits sonic hedgehog signal transduction. Development. 1998;125:3553–3562. doi: 10.1242/dev.125.18.3553. [DOI] [PubMed] [Google Scholar]
- Chen JK, Taipale J, Cooper MK, Beachy PA. Inhibition of Hedgehog signaling by direct binding of cyclopamine to Smoothened. Genes Dev. 2002;16:2743–2748. doi: 10.1101/gad.1025302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank-Kamenetsky M, Zhang XM, Bottega S, Guicherit O, Wichterle H, Dudek H, Bumcrot D, Wang FY, Jones S, Shulok J, Rubin LL, Porter JA. Small molecule modulators of hedgehog signaling: identification and characterization of smoothened agonists and antagonists. J Biol. 2002;1:10. doi: 10.1186/1475-4924-1-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stecca B, Ruiz i Altaba A. The therapeutic potential of modulators of the Hedgehog-Gli signaling pathway. J Biol. 2002;1:9. doi: 10.1186/1475-4924-1-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taipale J, Cooper MK, Maiti T, Beachy PA. Patched acts catalytically to suppress the activity of Smoothened. Nature. 2002;418:892–897. doi: 10.1038/nature00989. [DOI] [PubMed] [Google Scholar]
- Mitchison TJ. Towards a pharmacological genetics. Chem Biol. 1994;1:3–6. doi: 10.1016/1074-5521(94)90034-5. [DOI] [PubMed] [Google Scholar]
- Crews CM, Splittgerber U. Chemical genetics: exploring and controlling cellular processes with chemical probes. Trends Biochem Sci. 1999;24:317–320. doi: 10.1016/S0968-0004(99)01425-5. [DOI] [PubMed] [Google Scholar]
- Stockwell BR. Chemical genetics: ligand-based discovery of gene function. Nat Rev Genet. 2000;1:116–125. doi: 10.1038/35038557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taipale J, Chen JK, Cooper MK, Wang B, Mann RK, Milenkovic L, Scott MP, Beachy PA. Effects of oncogenic mutations in Smoothened and Patched can be reversed by cyclopamine. Nature. 2000;406:1005–1009. doi: 10.1038/35023008. [DOI] [PubMed] [Google Scholar]
- Williams JA, Guicherit OM, Zaharian BI, Xu Y, Chai L, Gatchalian C, Porter JA, Rubin LL, Wang FY. Identification of novel inhibitors of the hedgehog signaling pathway: Effects on basal cell carcinoma-like lesions. Proc Natl Acad Sci USA. [DOI] [PMC free article] [PubMed]
- Chen JK, Taipale J, Young KE, Maiti T, Beachy PA. Small molecule modulation of Smoothened activity. Proc Natl Acad Sci USA. 2002;99:14071–14076. doi: 10.1073/pnas.182542899. [DOI] [PMC free article] [PubMed] [Google Scholar]