

Abstract
An advanced understanding of acid–base physiology is as central to the practice of critical care medicine, as are an understanding of cardiac and pulmonary physiology. Intensivists spend much of their time managing problems related to fluids, electrolytes, and blood pH. Recent advances in the understanding of acid–base physiology have occurred as the result of the application of basic physical-chemical principles of aqueous solutions to blood plasma. This analysis has revealed three independent variables that regulate pH in blood plasma. These variables are carbon dioxide, relative electrolyte concentrations, and total weak acid concentrations. All changes in blood pH, in health and in disease, occur through changes in these three variables. Clinical implications for these findings are also discussed.
Keywords: Acid–base balance, acidosis, alkalosis, anion gap, arterial blood gases, strong ion difference, strong ion gap
Introduction
Whereas most medical and surgical subspecialists concern themselves with a specific organ (eg nephrology), region of the body (eg cardiothoracic surgery), or disease process (eg infectious disease), practitioners of critical care are more often concerned with the interaction of various organs and disease states. As such, our practice is often defined by certain syndromes (sepsis, multiorgan failure) and pathophysiologic conditions (shock) that do not confine themselves to the domains of a single subspecialty. Acid–base regulation is one of these 'areas' of medicine that crosses organ-specific boundaries, and the intensive care unit is often the place where severe derangements in this area exist. For these reasons, intensivists, and others called upon to care for critically ill patients in the intensive care unit, operating room, or emergency department, are expected to diagnose and manage complicated disorders of acid–base balance. This review provides a rather in-depth examination of chemistry and physiology of acid–base balance in health and disease.
The concentration of H+ in blood plasma and various other body solutions is among the most tightly regulated variables in human physiology. (Most of the principles discussed in this review are applicable to animal physiology as well. A complete discussion of the differences between species, however, particularly aquatic versus terrestrial species, is beyond the scope of the present review.) Acute changes in blood pH induce powerful regulatory effects at the level of the cell, organ, and organism [1]. The mechanisms responsible for local, regional, and systemic acid–base balance are incompletely understood though, and controversy exists in the literature as to what methods should be used to understand these mechanisms [2]. Much of this controversy exists only because the strict rules for causation (as opposed to correlation) have not often been applied to the understanding of acid–base balance, and methods that are useful clinically have often been used to understand physiology without being subjected to appropriate scientific rigor. The use of various laboratory variables to diagnose an acid–base disorder is analogous to the use of the electrocardiogram to diagnose a myocardial infarction. However, neither the changes in the electrocardiogram tracing nor the disturbances in electrical conduction that these changes reflect were ever considered to be the cause of a myocardial infarction. In contrast, changes in HCO3- (bicarbonate)concentration, for example, have been assumed to be responsible for metabolic acidosis or alkalosis. Failure to establish causation has lead to numerous incorrect notions of acid–base physiology and has fueled years of, often heated, debate [2,3,4]. This review analyzes what is known about the causal relationships between acid–base variables and acid–base balance in health and disease.
Quantification, classification, and causation
In order to understand acid–base physiology, we must first agree on how to describe and measure it. Since Sörensen (1868–1939) first introduced the pH notation, we have used the pH scale to quantify acid–base balance The pH scale has a tremendous advantage because it lends itself to colorimetric and electrometric techniques. There is also some physiologic relevance to the logarithmic pH scale [5]. pH is a confusing variable, however. It is a nonlinear transformation of H+ concentration – the logarithm of its reciprocal. Strictly speaking, pH can only be thought of as a dimensionless representation of H+ concentration and is not, itself, a concentration. Indeed, pH is actually the logarithmic measure of the volume required to contain 1 mol/l of H+. In blood plasma at pH 7.4, this volume is approximately 25 million liters [6].
Regardless of how we express the concentration of H +, either directly or as the pH, it is generally accepted that changes in blood H +concentration occur as the result of changes in volatile [partial carbon dioxide tension (pCO2)] and nonvolatile acids (hydrochloric, sulfuric, lactic, etc). Clinically, we refer to changes in volatile acids as 'respiratory' and changes in nonvolatile acids as 'metabolic'. There are three major methods of quantifying (describing) acid–base disorders, and each differs only in assessment of this latter, 'metabolic' component. These three methods quantify the metabolic component either by using HCO3 - (in the context of pCO2), the standard base excess (SBE),or the strong ion difference (SID). Although there has been significant debate regarding the accuracy and utility of each method compared with the others, all three yield virtually identical results when used to quantify the acid–base status of a given blood sample [7,8]. The only differences between these three approaches are conceptual (ie in how they approach the understanding of mechanisms) [9,10,11].
Beyond Henderson and Hasselbalch
Since Hasselbalch adapted the Henderson equation to the pH notation of Sörenson, we have used the following equation to understand the relationship between respiratory and metabolic acid–base variables:
pH = pK × log [HCO3 /(0.03 × pCO2)] (1)
This is the Henderson–Hasselbalch equation, and it is important to realize what this equation tells us. Anincrease in pCO2 will result in a decrease in the pH and anincrease in the HCO3- concentration. Thus, a patientfound to have a low blood pH, a condition known asacidemia, will either have an increased pCO2 or a pCO2that is 'not increased'. In the former circumstance, we classify the disorder as a 'respiratory acidosis'. We use the term 'acidosis' to describe the process resulting in acidemia and 'respiratory' because the apparent cause is an increased pCO2. This is logical, because carbonic acid results when CO2 is added to water (or blood), and the resultant decrease in pH is entirely expected. In the latter condition pCO2 is not increased, and thus there cannot be a respiratory acidosis. We therefore refer to this condition as 'metabolic' because some nonvolatile acid must be the cause of the acidemia. We can reverse the above logic and easily classify simple conditions of alkalemia as either resulting from respiratory or metabolic alkaloses. Thus, equation 1 allows us to classify disorders according to the primary type of acid being increased or decreased. Over time physiology superimposes its effects on simple chemistry and the relationship between pCO2 and HCO3- is altered in order to reduce the alterations in pH. By carefully examining the changes that occur in pCO2 and HCO3- in relationship to each, however, one can discernhighly conserved patterns. In this way rules can be established to allow one to discover mixed disorders and to separate chronic from acute respiratory derangements. For example one such rule is the convenient formula for predicting the expected pCO2 in the setting of a metabolic acidosis [12]:
pCO2 = (1.5 × HCO3-) + 8 ± 5 (2)
This rule tells us what the pCO2 should be secondary to the increase in alveolar ventilation that accompanies a metabolic acidosis. If pCO2 does not change enough or changes too much, we classify the condition as a 'mixed' disorder, with either a respiratory acidosis if the pCO2 is still too high, or a respiratory alkalosis if the change is too great. This rule, along with others (Table 1) has been recently translated to SBE terminology [7]:
Table 1.
Observational acid–base patterns
| Disorder | HCO3- (mmol/l) | pCO2 (mmHg) | SBE (mmol/I) |
| Metabolic acidosis | <22 | = (1.5 × HCO3-) + 8 | <-5 |
| = 40 + SBE | |||
| Metabolic alkalosis | >26 | = (0.7 × HCO3-) + 21 | >+5 |
| = 40 + (0.6 × SBE) | |||
| Acute respiratory acidosis | = [(pCO2 – 40)/10] + 24 | >45 | = 0 |
| Chronic respiratory acidosis | = [(pCO2 – 40)/3] + 24 | >45 | = 0.4 × (pCO2 – 40) |
| Acute respiratory alkalosis | = [(40 – pCO2)/5] + 24 | <35 | = 0 |
| Chronic respiratory alkalosis | = [(40 – pCO2)/2] + 24 | <35 | = 0.4 × (pCO2 – 40) |
Compiled from various sources. The changes in SBE were taken from Schlichtig et al [7]. pCO2, partial carbon dioxide tension; SBE, standard base excess.
pCO2 = (40 + SBE) ± 5 (3)
For example, consider the following arterial blood gas sample: pH7.31, pCO2 31, HCO3- 15, SBE-9.5. Equation 2 tells us that the expected pCO2 =(1.5×15)+8 ± 5=30.5 ± 5, and in Equation 3 the SBE added to 40 also yields 30.5. The measured pCO2 of 31 mmHg is consistent with a pure metabolic acidosis (ie no respiratory disorder).
It is also very important to understand what the Henderson-Hasselbach equation does not tell us. First, it does not allow us to discern the severity (quantity) of the metabolic derangement in a manner analogous to the respiratory component. For example, when there is a respiratoryacidosis, the increase in the pCO2 quantifies the derangement even when there is a mixed disorder. However, the metabolic component can only be approximated by the change in HCO3-. Second, Equation 1 does not tell us about any acids other than carbonic acid. The relationship between CO2 and HCO3- provides a useful clinical 'roadmap' to guide the clinician in uncovering the etiology of an acid–base disorder as described above. The total CO2concentration, and hence the HCO3- concentration, is determined by the pCO2, however, which is in turn determined by the balance between alveolar ventilation and CO2 production. HCO3- cannot be regulated independent of pCO2. The HCO3- concentration in the plasma will always increase as the pCO2 increases, but this is not an alkalosis. To understand how the pH and HCO3- concentration are altered independent of pCO2, we must look beyond Henderson and Hasselbach.
Base excess
In order to address the first 'shortcoming' of the Henderson-Hasselbach equation – the inability to quantify the metabolic component – several methods have been devised. In 1948, Singer and Hastings proposed the term 'buffer base' to define the sum of HCO3- plus the nonvolatile weak acid buffers (A-) [13]. A change in buffer base corresponds to a change in the metabolic component. The methods for calculating the change in buffer base were later refined by investigators [14,15] and refined further by others [16,17,18] to yield the base excess methodology. Base excess is the quantity of metabolic acidosis or alkalosis, defined as the amount of acid or base that must be added to a sample of whole blood in vitro in order to restore the pH of the sample to 7.40 while the pCO2 is held at 40 mmHg [15]. Although this calculation is quite accurate in vitro, inaccuracy exists when applied in vivo in that base excess changes with changes in pCO2 [19,20]. This effect is understood to be due to equilibration across the entire extracellular fluid space (whole blood + interstitial fluid). When the base excess equation is modified to account for an 'average' content of hemoglobin across this entire space, a value of 5 g/dl is used instead, and this defines the SBE. It should be pointed out that this value does not represent the true content of hemoglobin suspended in the volume of whole blood together with interstitial fluid, but rather is an empiric estimate that improves the accuracy of the base excess. It can be argued that the entire extracellular fluid space is involved in acid–base balance, because this fluid flows through blood vessles and lymphatics, mixing constantly [21]. Thus, the value of SBE is that it quantifies the change in metabolic acid–base status in vivo. It is of interest that base excess is only accurate in vivo when it assumes a constant hemoglobin concentration.
However, the base excess approach does not address the second problem associated with using the Henderson-Hasselbach equation alone (ie it still does not tell us about the mechanisms of metabolic acid–base balance). For example the body does not 'regulate' the SBE. It is not a substance that can be excreted in the feces or reabsorbed from the proximal tubule. Similarly, HCO3 - is not a strong acid or base and its addition to or removal from the plasma cannot be translated into changes in SBE. This not to say that changes in SBE and HCO3- do not correlate closely, because they do. However, correlation and causation are not the same thing. The difference has traditionally been ascribed to the effects of 'buffering', the argument being that stong acid (or base), quantified by SBE, is 'buffered' by plasma proteins, hemoglobin, and finally by HCO3-. The resulting changes in HCO3-and pH are then a result of this buffering process. These buffers are actually weak acids, however, and their addition to plamsa both decreases the pH and increases the responsiveness to pCO2 (Fig. 1). Furthermore, as explained by Stewart [6,9], the fundamental physical-chemical properties of biologic solutions dictate much of this so-called 'buffering'.
Figure 1.
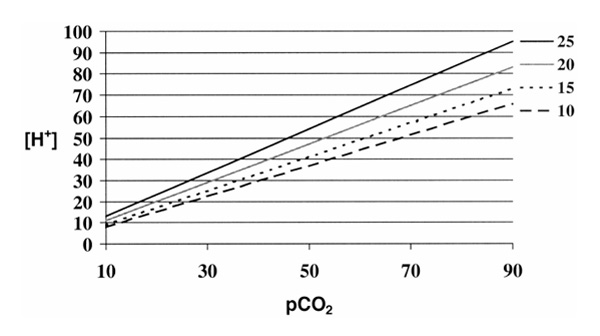
Changes in the relationship between partial carbon dioxide tension (pCO2) and H+ concentration as function of changes in 'buffer' strength. Individual curves are drawn for varying concentration of total nonvolatile buffers in mmol/l. Note that as the concentration of 'buffer' increases, the slope of the curve increases, making changes in H+ concentration more responsive to changes in CO2.
Physical-chemical properties of biologic solutions
A physical–chemical analysis of acid–base physiology requires the application of two basic principles. The first is electroneutrality, which dictates that, in aqueous solutions, the sum of all positively charged ions must equal the sum of all negatively charged ions. The second is conservation of mass, which means that the amount of a substance remains constant unless it is added to or generated, or removed or destroyed. These principles may seem very basic indeed, but they are often overlooked in the analysis of clinical acid–base physiology, leading to incorrect conclusions. For example, a hyperchloremic metabolic acidosis can only be brought about in two ways. First, Cl- ions can be added to the circulation, either via an exogenous source (eg HCl or saline) or via internal shifts (eg from the red cell). Second, Cl- ions can be retained or reabsorbed, whereas water and other ions (ie Na+) are excreted so that the relative concentration of Cl- increases. A decrease in - HCO3 or H+ concentration does not produce hyperchloremia, but rather hyperchloremia is a cause of acidosis. This distinction is not merely semantics, any more than Copernicus' observation that the Earth, rather than the sun moves [11,22].
In addition to these physical–chemical principles, almost all solutions of biologic interest share two important characteristics. First, virtually all are aqueous (composed of water), and second, most are alkaline (OH-concentration >H+ concentration). Because these characteristics are so universal in human physiology, they are often ignored in reviews of physiology, especially for clinical medicine, but they are extremely important. Aqueous solutions contain a virtually inexhaustible source of H+. Although pure water dissociates only slightly into H+ and OH-, electrolytes and CO2 produce powerful electrochemical forces that influence water dissociation. Similarly, aqueous solutions that are alkaline behave very differently compared with acidic solutions in terms of the extent to which changes in their composition influence changes in pH.
To illustrate this point, first consider a 1 l solution of pure water. Pure water contains only a small amount of H+ and OH- ions and molecular H2O. Pure water is a neutral solution by definition, because the H+ and OH- concentrations are equal. The concentration of these ions is determined solely by the extent to which water dissociates and can be defined by a constant, K'w. Water dissociation is temperature sensitive because K'w is, but, at all times, the concentrations of H+ and OH- must be equal, and H+× OH- = K'w. If we add 10 mmol/l of each Na+ and Cl- to this 1 lsolution of pure water, we would have an aqueous solution that contains H+, OH-, Na+ and Cl- ions, and molecular water. Of note, the solution does not contain any molecules of NaOH, HCl, or NaCl, because both Na+ and Cl- are strong ions and as such are completely dissociated in water. The solution we now have is still a neutral solution by definition, and at 25ºC the concentrations of both H+ and OH- are approximately 100 nmol/l, or pH7.0. If we then add 10 mmol/l HCl, we will have a solution that contains 10 mmol/l Na+ and 20 mmol/l Cl-. This solution is acidic: OH- =4.4 × 10-9 nmol/l, and H+ = ~ 10 mmol/l. Note that in this acidic solution the H+ concentration increased by the amount of H+ added (ie 10 mmol/l). However, if we were to add 10 mmol/l NaOH instead of HCl, the solution would instead contain 20 mmol/l Na+ and 10 mmol/l Cl-, and would be alkaline: H+ = 4.4 × 10-9 nmol/l and OH- = ˜ 10 mmol/l. If we then add 5 mmol/l HCl to this alkaline solution, the resulting concentration of Na+ would be 20 mmol/l and of Cl- would be 15 mmol/l. The final H+ concentration is now 8.8 × 10-9 nmol/l and OH- is approximately 5 mmol/l. Note that in this final example 5 mmol/l of H+ were added to the solution, yet the final concentration of free H+ changed by less than billionth of this amount. It should be further noted that the solution I have described contains no 'buffers', and thus what is often attributed to the power of buffering systems is merely a physical–chemical property of alkaline solutions.
Determinants of hydrogen concentration
From the preceding discussion it is apparent that, for aqueous solutions, water is the primary source of H+, and the determinants of H+concentration are the determinants of water dissociation. Fortunately, even for an aqueous solution as complex as blood plasma, there are but three independent variables that determine H+ concentration. Please note that I use the term 'determine' rather than 'describe' because, as shown by Stewart [6,9], these three variables are mathematically independent determinants of the H+ concentration. Thus, these variables are causally related to the H+ concentration rather than being merely correlated. The distinction between independent and dependent, between causation and correlation, is as important to acid–base physiology as to any other area of science. Only by the careful analysis of causal variables can mechanisms be determined. For blood plasma, these three variables are pCO2, SID, and the total weak acid concentration (ATOT).
Carbon dioxide
CO2 is an independent determinant of pH and is produced by cellular metabolism or by the titration of HCO3 - by metabolic acids. Normally, alveolar ventilation is adjusted to maintain the arterial pCO2 between 35 and 45 mmHg. When alveolar ventilation is increased or decreased out of proportion to pCO2 production, a respiratory acid–base disorder exists. CO2 production by the body (at 220 ml/min) is equal to 15000 mmol/day of carbonic acid [23]. This compares with less than 500 mmol/day for all nonrespiratory acids. The respiratory center, in response to signals from pCO2, pH, and partial oxygen tension, as well as some from exercise, anxiety, wakefulness, and others, controls alveolar ventilation. A precise match of alveolar ventilation to metabolic CO2 production attains the normal arterial pCO2 of 40 mmHg. Arterial pCO2 is adjusted by the respiratory center in response to altered arterial pH produced by metabolic acidosis or alkalosis in predictable ways.
When CO2 elimination is inadequate relative to the rate of tissue production, pCO2 will increase to a new steadystate that is determined by the new relationship between alveolar ventilation and CO2 production. Acutely, this increase in pCO2will increase both the H+ and the HCO3- concentrations according to the Henderson-Hasselbach equation (Equation 1). Thus, this change in HCO3- concentration is mediated by chemical equilibrium, and not by any systemic adaptation. Similarly, this increased HCO3- concentration does not 'buffer' H+ concentration. There is no change in the SBE. Tissue acidosis always occurs in respiratory acidosis, because CO2 diffuses into the tissues. If the pCO2 remains increased the body will attempt to compensate by altering another independent determinant of pH, namely the SID.
Electrolytes (strong ions)
Blood plasma contains numerous ions. These ions can be classified both by charge, positive 'cations' and negative 'anions', as well as by their tendency to dissociate in aqueous solutions. Some ions are completely dissociated in water, for example, Na+, K+, Ca2+, Mg2+, and Cl-. These ions are called 'strong ions' to distinguish them from 'weak ions' (eg albumin, phosphate and HCO3-), which can exist both as charged (dissociated) and uncharged forms. Certain ions such as lactate are so nearly completely dissociated that they may be considered strong ions under physiologic conditions. In a neutral salt solution containing only water and NaCl, the sum of strong cations (Na+) minus the sum of strong anions (Cl-) is zero (ie Na+ = Cl-). In blood plasma, however, strong cations (mainly Na+) outnumber strong anions (mainly Cl-). The difference between the sum of all strong cations and all strong anions is known as the SID. SID has a powerful electrochemical effect on water dissociation, and hence on H+ concentration. As SID becomes more positive, H+, a 'weak' cation, decreases (and pH increases) in order to maintain electrical neutrality (Fig. 2).
Figure 2.
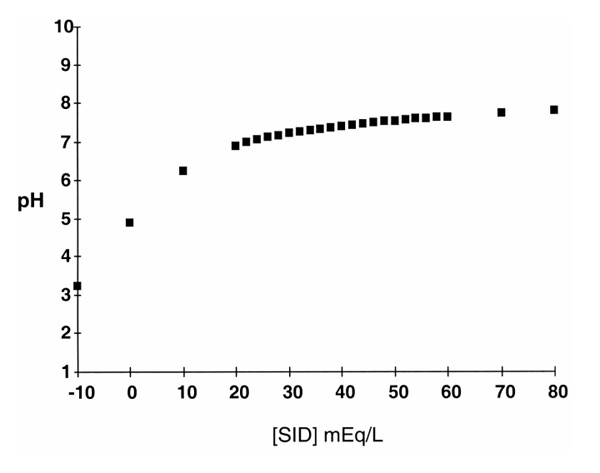
Plot of pH versus strong ion difference (SID). For this plot, total weak acid concentration (ATOT) and partial carbon dioxide tension (pCO2) were held constant at 18 mmol/l and 40 mmHg, respectively. Assumes a water dissociation constant for blood of 4.4 × 10–14 (mol/l). Note how steep the pH curve becomes at SID <20 mmol/l. Adapted from Kellum and Kellum [10].
In healthy humans, the plasma SID is between 40 and 42 mmol/l, although it is often quite different in critically ill patients. According to the principle of electrical neutrality, blood plasma cannot be charged, so the remaining negative charges balancing the SID come from CO2 and theweak acids (A-) and, to very small extent, from OH-. At physiologic pH, the contribution of OH- is so small (nmol range) that it can be ignored. The total weak acid concentration (mainly albumin and phosphate) can be considered together and for convenience is abbreviated ATOT, where AH + A- = ATOT. The SID of a blood sample can be estimated from the value of the remaining negative charge, because SID-(CO2 +A -)=0. This estimate of SID has been termed the 'effective' SID [24], but it is really no different from buffer base described over half a century ago [13]. Thus SID and buffer base are mirror images of each other. Recall that SBE is essentially the change in buffer base in vivo, and hence SBE defines the change in SID from the equilibrium point where pH=7.4 and pCO2 = 40 mmHg [8].
An alternative estimate of SID is as follows: (Na+ + K+ + Ca2++ Mg2+) – (Cl- + lactate-). This is often referred to as the 'apparent' SID with the understanding that some 'unmeasured' ions might also be present [24]. Neither effective SID nor the apparent SID are perfect estimates of the true SID. Blood samples from patients may contain unmeasured strong ions (eg sulfate, ketones) making the apparent SID an inaccurate estimate of SID. Similarly, these patients may have abnormal weak ions (eg proteins) that will make the effective SID inaccurate. In healthly humans, however, the apparent SID and the effective SID are nearly identical, and are thus valid estimates of SID [24]. Furthermore, when the apparent SID and the effective SID are not equal, a condition we have referred to as the strong ion gap (SIG), where apparent SID – effective SID = SIG, abnormal strong and/or weak ions must be present [25]. The SIG is positive when unmeasured anions exceed unmeasured cations, and negative when unmeasured cations exceed unmeasured anions. Unexplained anions, and in some cases cations, have been found in the circulation of patients with a variety of diseases [25,26,27,28] and in animals under experimental conditions [29].
The SIG is not the same as the anion gap (AG). Normally, the SIG is zero, whereas the AG is 8–12mmol/l. The AG is an estimate of the sum of SIG + A-. Thus, subtracting A- from the AG approximates the SIG. A convenient and reasonably accurate way to estimate A- is to use the following formula [30]:
2 (albumin g/dl) + 0.5 (phosphate mg/dl) (4)
or for international units:
0.2 (albumin g/l) + 1.5 (phosphate mmol/l) (5)
Note that the 'normal' AG for a person with no unmeasured anions or cations in their plasma is equal to A-, such that AG - A- = SIG = 0. This technique allows one to 'calibrate' the AG for patients with abnormal albumin and/or phosphate concentrations.
Physiologic mechanisms
In order to alter the SID, the body must affect a change in the relative concentrations of strong cations and strong anions. The kidney is the primary organ that affects this change. However, the kidney can only excrete a very small amount of strong ion into the urine each minute, and several minutes to hours are therefore required to impact significantly on the SID. The handling of strong ions by the kidney is extremely important because every Cl- ion filtered but not reabsorbed increases the SID. Because most of the human diet contains similar ratios of strong cations to strong anions, there is usually sufficient Cl-available for this to be the primary regulating mechanism. This is particularly apparent when we consider that renal Na+ and K+ handling are influenced by other priorities (eg intravascular volume and plasma K+ homeostasis). Accordingly, 'acid handling' by the kidney is generally mediated through Cl- balance. How the kidney handles Cl- is obviously very important. Traditional approaches to this problem have focused on H+ excretion and have emphasized the importance of NH3 and its add-on cation NH4+. H+ excretion per se is irrelevant, however, because water provides an essentially infinite source of free H+. Indeed the kidney does not excrete H+ any more as NH4+ than it does as H2O. The purpose of renal ammoniagenesis is to allow the excretion of Cl- without excreting Na+ or K+. This is achieved by supplying a weak cation (NH4+) to excrete with Cl-.
Thus, NH4+ is important to systemic acid–base balance,not because of its carriage of H+ or because of its direct action in the plasma (normal plasma NH4+ concentration is <0.01mmol/l), but because of its 'coexcretion' with Cl-. Of course, NH4+ is not only produced in the kidney. Hepatic ammoniagenesis (and also glutaminogenesis) is important for systemic acid–base balance and, as expected, it is tightly controlled by mechanisms that are sensitive to plasma pH [31]. Indeed this reinterpretation of the role of NH4+ in acid–base balance is supported by the evidence that hepatic glutaminogenesis is stimulated by acidosis [32]. Nitrogen metabolism by the liver can result in either urea, glutamine or NH4+. Normally, the liver does not release more than a very small amount NH4+, but rather incorporates this nitrogen into either urea or glutamine. Hepatocytes have enzymes to enable them to produce either of these end products, and both allow for the regulation for plasma NH4+ at suitably low levels. The production of urea or glutamine has significantly different effects at the level of the kidney. This is because glutamine is used by the kidney to generate NH4+ and facilitatethe excretion of Cl-. Thus, the production of glutamine can be seen as having an alkalinizing effect on plasma pH because of the way in which the kidney utilizes it.
Further support for this scenario comes from the recent discovery of an anatomic organization of hepatocytes according to their enzymatic content [33]. Hepatocytes with a propensity to produce urea are positioned closer to the portal venule, and thus have the first chance at the NH4+delivered. Acidosis inhibits ureagenesis, however, and under these conditions more NH4+ is available for the downstream hepatocytes that are predisposed to produce glutamine. Thus, the leftover NH4+ is 'packaged' as glutamine for export to the kidney, where it is used to facilitate Cl- excretion and hence increases the SID.
The gastrointestinal tract also has important effects on the SID. Along its length, the gastrointestinal tract handles strong ions quite differently. In the stomach, Cl- is pumped out of the plasma and into the lumen, reducing the SID of the gastric juice and thus reducing the pH. On the plasma side, SID is increased by the loss of Cl- and the pH is increased, producing the so-called 'alkaline tide' that occurs at the beginning of a meal when gastric acid secretion is maximal [34]. In the duodenum Cl- is reabsorbed and the plasma pH is restored. Normally, only slight changes in plasma pH are evident because Cl- is returned to the circulation almost as soon as it is being removed. If gastric secretions are removed from the patient, however, either by suction catheter or vomiting, Cl- will be progressively lost and the SID will steadily increase. It is important to realize that it is the Cl- loss, and not the H+ that is the determinant of plasma pH. Although H+ is 'lost' as HCl, it is also lost with every molecule of water removed from the body. When Cl- (a strong anion) is lost without loss of a strong cation, the SID is increased and therefore the plasma H+ concentration is decreased. When H+ is 'lost' as water (HOH) rather than HCl, there is no change in the SID and hence no change in the plasma H+ concentration.
In contrast to the stomach, the pancreas secretes fluid into the small intestine that has a SID much higher than that of plasma and is very low in Cl-. Thus, the plasma perfusing the pancreas has its SID decreased, a phenomenon that peaks about 1 h after a meal and helps counteract the alkaline tide. If large amounts of pancreatic fluid are lost, for example from surgical drainage, an acidosis will result as a consequence of the decreased plasma SID. In the large intestine, fluid also has a high SID because most of the Cl- has been removed in the small intestine and the remaining electrolytes are mostly Na+ and K+. The body normally reabsorbs much of the water and electrolytes from this fluid, but when severe diarrhea exists large amounts of cations can be lost. If this loss is persistent, the plasma SID will decrease and acidosis will result. Finally, whether the gastrointestinal tract is capable of regulating strong ion uptake in a compensatory manner has not been well studied. There is some evidence that the gut may modulate systemic acidosis in experimental endotoxemia by removing anions from the plasma [35]. The full capacity of this organ to affect acid–base balance is unknown, however.
Pathophysiologic mechanisms
Metabolic acidoses and alkaloses are categorized according to the ions that are responsible. Thus we have lactic acidosis and chloride responsive alkalosis, etc. It is important to recognize that metabolic acidosis is produced by a decrease in the SID, which produces an electrochemical force that results in an increase in free H+ concentration. A decrease in SID may be brought about by the generation of organic anions (eg lactate, ketones), the loss of cations (eg diarrhea), the mishandling of ions (eg renal tubular acidosis), or the addition of exogenous anions (eg iatrogenic acidosis, poisonings). By contrast metabolic alkaloses occur as a result of an inappropriately large SID, although the SID need not be greater than the 'normal' 40-42 mmol/l. This may be brought about by the loss of anions in excess of cations (eg vomiting, diuretics), or rarely by administration of strong cations in excess of strong anions (eg transfusion of large volumes of banked blood). Tables 2 and 3 provide a useful means to differentiate the various causes of metabolic acidosis and alkalosis.
Table 2.
Differential diagnosis for metabolic acidosis (decreased SID)
| Renal tubular acidosis: | Nonrenal: |
| urine SID (Na+ + K+ – Cl) > 0 | urine SID (Na+ + K+ – Cl-) < 0 |
| Distal (type I): urine pH >5.5 | Gastrointestinal: diarrhea, small |
| bowel/pancreatic drainage | |
| Proximal (type II): urine | |
| pH <5.5/low serum K+ | Iatrogenic: parenteral nutrition, |
| saline, anion exchange resins | |
| Aldosterone deficiency (type IV): | |
| urine pH <5.5/high serum K+ |
SID, strong ion difference.
Table 3.
Differential diagnosis of a metabolic alkalosis (increased SID)
| Chloride loss < sodium loss |
| Chloride responsive (urine Cl- concentration <10 mmol/l) |
| Gastrointestinal losses: vomiting, gastric drainage, chloride |
| wasting diarrhea (villous adenoma) |
| Postdiuretic use |
| Posthypercapnea |
| Chloride unresponsive (urine Cl- concentration >20 mmol/l) |
| Mineralocorticoid excess: primary hyperaldosteronism (Conn's |
| syndrome), secondary hyperaldosteronism, Cushing's |
| syndrome, Liddle's syndrome, Bartter's syndrome, |
| exogenous corticoids, excessive licorice intake |
| Ongoing diuretic use |
| Exogenous sodium load (>chloride) |
| Sodium salt administration (acetate, citrate): massive blood |
| transfusions, parenteral nutrition, plasma volume expanders, |
| sodium lactate (Ringer's solution) |
| Other |
| Severe deficiency of intracellular cations: Mg2+, K+ |
SID, strong ion difference.
In the intensive care unit acidosis is usually more of a problem than alkalosis, and in the critically ill the most common sources of metabolic acidosis are disorders of chloride homeostasis, lactate, and other anions. Hyperchloremic metabolic acidosis occurs either as a result of chloride administration or secondary to abnormalities in chloride handling or related to movements of chloride from one compartment to another. The effect of chloride administration on the development of metabolic acidosis has been known for many years [36,37]. Recently, new attention has been paid to this area in light of better understanding of the mechanisms responsible for this effect [38,39,40]. It has now been shown in animal models of sepsis [38] and in patients undergoing surgery [39,40] that saline causes metabolic acidosis not by 'diluting' HCO3-, but rather by its Cl- content. From a physical-chemical prospective, this is completely expected. HCO3-is a dependent variable and cannot be the cause of the acidosis. Instead, Cl- administration decreases the SID (an independent variable) and produces an increase in water dissociation and hence H+ concentration. The reason why this occurs with saline administration is that, although saline contains equal amounts of both Na+ and Cl-, the plasma does not. When large amounts of salt are added, the Cl- concentration increases much more than the sodium concentration. For example, 0.9% ('normal') saline contains 154 mmol/l of Na+ and Cl-. Administration of large volumes of this fluid will have a proportionally greater effect on total body Cl- than on total body Na+. Of note, it is the total body concentrations of these strong ions that must be considered and, although the true volume of distribution of Cl- is less, like Na+, the effective volume of distribution (after some time of equilibration) is equal to total body water [38].
There are other important causes of hyperchloremia (Table 2) and, in addition, this form of metabolic acidosis is common in critical illness, especially sepsis. Although saline resuscitation undoubtedly plays a role, there appear to be unexplained sources of Cl-, at least in animal models of sepsis [38]. We have hypothesized that this Cl- comes from intracellular and interstitial compartments as a result of the partial loss of Donnan equilibrium due to albumin exiting the intravascular space. However, this hypothesis is yet untested.
In addition to Cl-, several other strong ions may be present in the blood of critically ill patients. Lactate is perhaps the most important of these, but ketones, sulfates, and certain poisons (eg methanol, salicylate) are important in the appropriate clinical conditions. In addition, unexplained anions have been shown to be present in the blood of many critically ill patients [25,26,27,28].
Weak acids
The third and final determinant of H+ concentration is ATOT. The weak acids are mostly proteins (predominantlyalbumin) and phosphates, and they contribute the remaining charges to satisfy the principle of electroneutrality, such that SID–(CO2 +A-)=0. However, A- is not an independent variable because it changes with alterations in SID and pCO2. Rather, ATOT (AH + A-) is the independent variable, because its value is not determined by any other. The identification of ATOT as the third independent acid–base variable has lead some authors to suggest that a third 'kind' of acid–base disorder exists [41,42]. Thus, along with respiratory and metabolic acidosis and alkalosis, we would also have acidosis and alkalosis due to abnormalities in ATOT. However, mathematical, and therefore chemical independence does not necessarily imply physiologic independence. Although the loss of weak acid (ATOT) from the plasma space is an alkalinizing process,there is no evidence that the body regulates ATOT to maintain acid–base balance. Furthermore, there is no evidence that we as clinicians should treat hypoalbuminemia as an acid–base disorder.
Critically ill patients frequently have hypoalbuminemia andas such their ATOT is reduced. These patients are not often alkalemic and their SID is also reduced, however [43]. When these patients have a normal pH and a normal SBE and HCO3- concentration, it would seem most appropriate to consider this to be physiologic compensation for a decreased ATOT [44], rather than classifying this condition as a complex acid–base disorder with a mixed metabolic acidosis/hypoalbuminemic alkalosis. Thus, it seems far more likely that this 'disorder' is in fact the normal physiologic response to a decreased ATOT. Furthermore, because changes in ATOT generally occur slowly, the development of alkalemia would require the kidney to continue to excrete Cl- despite an evolving alkalosis. I would consider such a scenario to be renal-mediated hypochloremic metabolic alkalosis, the treatment for which would include fluids and/or chloride, depending on the clinical conditions. Stewart's designation of a 'normal' SID of approximately 40 mmol/l was based on a 'normal' CO2 and ATOT [6,9]. The 'normal' SID for a patient with an albumin of 2g/dl would be much lower (eg approximately 32 mmol/l).
Conclusion
Unlike many other areas in clinical medicine, the approach to acid–base physiology has not often distinguished cause from effect. Although it is perfectly reasonable to describe an alteration in acid–base status by the observed changes in H+ and HCO3-, this does not itself imply causation. The essence of the Stewart [6,9] approach is the understanding that only three variables are important in determining H+ concentration: pCO2, SID and ATOT. Neither H+ nor HCO3- can change unless one or more of these three variables change. The principle of conservation of mass makes this point more than semantics. Strong ions cannot be created or destroyed to satisfy electroneutrality but H+ ions are generated or consumed by changes in water dissociation. Hence, in order to understand how the body regulates pH, we need only consider how it regulates these three independent variables. Other approaches to acid–base physiology have ignored the distinction between independent and dependent variables, and although it is possible to describe an acid–base disorder in terms of H+ or HCO3- concentrations or SBE, it is incorrect to analyze the pathology or to plan treatment on the basis of altering these variables.
References
- Kellum JA. Diagnosis and treatment of acid–base disorders. Textbook of Critical Care. Edited by Grenvik A, Shoemaker PK, Ayers S, Holbrook PR. Philadelphia, PA: WB Saunders Co, 1999. pp. 839–853.
- Severinghaus JW. Siggard-Andersen and the 'great trans-Atlantic acid–base debate'. Scand J Clin Lab Invest. 1993;53 (suppl 214):99–104. [PubMed] [Google Scholar]
- Siggard-Andersen O, Foch-Andersen N. Base excess or buffer base (strong ion difference) as measure of a non-respiratory acid–base disturbance. Acta Anaesthiol Scand. 1995;39(suppl 107):123–128. doi: 10.1111/j.1399-6576.1995.tb04346.x. [DOI] [PubMed] [Google Scholar]
- Worthley L. Strong ion difference: a new paradigm or new clothes for the acid–base emperor. Crit Care Resusc. 1999;1:211–214. [PubMed] [Google Scholar]
- Severinghaus JW. More RipH [letter]. JAMA. 1992;267:2035–2036. doi: 10.1001/jama.1992.03480150041022. [DOI] [PubMed] [Google Scholar]
- Stewart PA. How to understand acid–base. A Quantitative acid–base Primer for Biology and Medicine. Edited by Stewart PA, New York: Elsevier, 1981. pp. 1–286.
- Schlichtig R, Grogono AW, Severinghaus JW. Human PaCO2 and standard base excess compensation for acid–base imbalance. Crit Care Med. 1998;26:1173–1179. doi: 10.1097/00003246-199807000-00015. [DOI] [PubMed] [Google Scholar]
- Schlichtig R, Kellum JA. Base excess and strong ion difference are quantitatively interchangeable. Adv Exp Med Biol 2000 (in press) 2000.
- Stewart PA. Modern quantitative acid–base chemistry. Can J Physiol Pharmacol. 1983;61:1444–1461. doi: 10.1139/y83-207. [DOI] [PubMed] [Google Scholar]
- Kellum JA, Kellum JA. Metabolic acidosis in the critically ill: lessons from physical chemistry. Kidney Int. 1998;53(suppl 66) :81–86. [PubMed] [Google Scholar]
- Kellum JA. Acid–base physiology in the post-Copernican era. Curr Opin Crit Care. 1999;5:458–463. [Google Scholar]
- Albert M, Dell R, Winters R. Quantitative displacement of acid–base equilibrium in metabolic acidosis. Ann Intern Med. 1967;66:312–322. doi: 10.7326/0003-4819-66-2-312. [DOI] [PubMed] [Google Scholar]
- Singer RB, Hastings AB. An improved clinical method for the estimation of disturbances of the acid–base balance of human blood. Medicine (Balt) 1948;27:223–242. doi: 10.1097/00005792-194805000-00003. [DOI] [PubMed] [Google Scholar]
- Astrup P, Jorgensen K, Siggaard-Andersen O, et al. Acid–base metabolism: new approach. Lancet. 1960;i:1035–1039. doi: 10.1016/s0140-6736(60)90930-2. [DOI] [PubMed] [Google Scholar]
- Siggaard-Andersen O. The pH-log pCO2 blood acid–base nomogram revised. Scand J Lab Invest. 1962;14:598–604. doi: 10.1080/00365516209051290. [DOI] [PubMed] [Google Scholar]
- Siggaard-Andersen O. The acid–base Status of the Blood, 4th edCopenhagen: Munksgaard, 1974.
- Grogono AW, Byles PH, Hawke W. An in vivo representation of acid–base balance. Lancet. 1976;i:499–500. doi: 10.1016/s0140-6736(76)90792-3. [DOI] [PubMed] [Google Scholar]
- Severinghaus JW. Acid–base balance nomogram: a Boston-Copenhagen détente. Anesthesiology. 1976;45:539–541. [PubMed] [Google Scholar]
- Brackett NC, Cohen JJ, Schwartz WB. Carbon dioxide titration curve of normal man. N Engl J Med. 1965;272:6–12. doi: 10.1056/NEJM196501072720102. [DOI] [PubMed] [Google Scholar]
- Prys-Roberts C, Kelman GR, Nunn JF. Determinants of the in vivo carbon dioxide titration curve in anesthetized man. Br J Anesth. 1966;38:500–509. doi: 10.1093/bja/38.7.500. [DOI] [PubMed] [Google Scholar]
- Schlichtig R. Acid–base balance (quantitation). Textbook of Critical Care. Edited by Grenvik A, Shoemaker PK, Ayers S, Holbrook PR.Philadelphia, PA: WB Saunders Co, 1999. pp. 828–839.
- Magder S. Pathophysiology of metabolic acid–base disturbances in patients with critical illness. Critical Care Nephrology. Edited by Ronco C, Bellomo R. Dordrecht, The Netherlands: Kluwer Academic Publishers, 1997. pp. 279–296.
- Gattinoni L, Lissoni A. Respiratory acid–base disturbances in patients with critical illness. Critical Care Nephrology. Edited by Ronco C, Bellomo R. Dordrecht, The Netherlands: Kluwer Academic Publishers, 1998. pp. 297–312.
- Figge J, Mydosh T, Fencl V. Serum proteins and acid–base equilibria: a follow-up. J Lab Clin Med. 1992;120:713–719. [PubMed] [Google Scholar]
- Kellum JA, Kramer DJ, Pinsky MR. Strong ion gap: a methodology for exploring unexplained anions. J Crit Care. 1995;10:51–55. doi: 10.1016/0883-9441(95)90016-0. [DOI] [PubMed] [Google Scholar]
- Gilfix BM, Bique M, Magder S. A physical chemical approach to the analysis of acid–base balance in the clinical setting. J Crit Care. 1993;8:187–197. doi: 10.1016/0883-9441(93)90001-2. [DOI] [PubMed] [Google Scholar]
- Mecher C, Rackow EC, Astiz ME, Weil MH. Unaccounted for anion in metabolic acidosis during severe sepsis in humans. Crit Care Med . 1991;19:705–711. doi: 10.1097/00003246-199105000-00018. [DOI] [PubMed] [Google Scholar]
- Kirschbaum B. Increased anion gap after liver transplantation. . Am J Med Sci. 1997;313:107–110. doi: 10.1097/00000441-199702000-00007. [DOI] [PubMed] [Google Scholar]
- Kellum JA, Bellomo R, Kramer DJ, Pinksy MR. Hepatic anion flux during acute endotoxemia. J Appl Physiol. 1995;78:2212–2217. doi: 10.1152/jappl.1995.78.6.2212. [DOI] [PubMed] [Google Scholar]
- Kellum JA, Kramer DJ, Pinsky MR. Closing the GAP: a simple method of improving the accuracy of the anion gap [abstract].Chest. Chest. 1996;110(suppl 4):18S. [Google Scholar]
- Bourke E, Haussinger D. pH homeostasis: the conceptual change. The Kidney Today. Selected Topics in Renal Science. Edited by Berlin GM. Contrib Nephrol. Basel: Karger, 1992. pp. 58–88. [DOI] [PubMed]
- Oliver J, Bourke E. Adaptations in urea and ammonium excretion in metabolic acidosis in the rat: a reinterpretation. Clin Sci Mol Med. 1975;48:515–520. doi: 10.1042/cs0480515. [DOI] [PubMed] [Google Scholar]
- Atkinson DE, Bourke E. pH Homeostasis in terrestrial vertebrates; ammonium ion as a proton source. Comparative and Environmental Physiology. Mechanisms of Systemic Regulation, acid–base Regulation, Ion Transfer and Metabolism. Edited by Heisler N. Springer: Berlin, 1995. pp. 1–26.
- Moore EW. The alkaline tide. Gastroenterology. 1967;52:1052–1054. [PubMed] [Google Scholar]
- Kellum JA, Bellomo R, Kramer DJ, Pinsky MR. Splanchnic buffering of metabolic acid during early endotoxemia. J Crit Care. 1997;12:7–12. doi: 10.1016/s0883-9441(97)90020-4. [DOI] [PubMed] [Google Scholar]
- Cushing H. Concerning the poisonous effect of pure sodium chloride solutions upon the nerve muscle preparation. Am J Physiol. 1902;6:77ff. [Google Scholar]
- Shires GT, Tolman J. Dilutional acidosis. Ann Intern Med. 1948;28:557–559. doi: 10.7326/0003-4819-28-3-557. [DOI] [PubMed] [Google Scholar]
- Kellum JA, Bellomo R, Kramer DJ, Pinsky MR. Etiology of metabolic acidosis during saline resuscitation in endotoxemia. . Shock. 1998;9:364–468. doi: 10.1097/00024382-199805000-00009. [DOI] [PubMed] [Google Scholar]
- Scheingraber S, Rehm M, Sehmisch C, Finsterer U. Rapid saline infusion produces hyperchloremic acidosis in patients undergoing gynecologic surgery. Anesthesiology. 1999;90:1265–1270. doi: 10.1097/00000542-199905000-00007. [DOI] [PubMed] [Google Scholar]
- Waters JH, Miller LR, Clack S, Kim JV. Cause of metabolic acidosis in prolonged surgery. Crit Care Med. 1999;27:2142–2146. doi: 10.1097/00003246-199910000-00011. [DOI] [PubMed] [Google Scholar]
- Fencl V, Leith DE. Stewart's quantative acid–base chemistry: applications in biology and medicine. Respir Physiol. 1993;91:1–16. doi: 10.1016/0034-5687(93)90085-o. [DOI] [PubMed] [Google Scholar]
- Jabor A, Kazda A. Modeling of acid–base equilibria. Acta Anaesth Scand. 1995;39(suppl 107):119–122. doi: 10.1111/j.1399-6576.1995.tb04345.x. [DOI] [PubMed] [Google Scholar]
- Kellum JA. Recent advances in acid–base physiology applied to critical care. Yearbook of Intensive Care and Emergency Medicine. Edited by Vincent JL. Heidelberg: Springer-Verlag, 1998. pp. 579–587.
- Wilkes P. Hypoproteinemia, SID, and acid–base status in critically ill patients. J Appl Physiol. 1998;84:1740–1748. doi: 10.1152/jappl.1998.84.5.1740. [DOI] [PubMed] [Google Scholar]