

Summary
The PITP family is one of the two families of phosphoinositide transfer proteins that can bind and exchange one molecule of either phosphatidylinositol or phosphatidylcholine and facilitate the transfer of these lipids between different membrane compartments; the other is the structurally unrelated Sec14p family. PITPs have recently been shown to be critical regulators of phosphoinositides in several cellular compartments, where they participate in signal transduction and in membrane traffic. PITPs were originally defined as soluble, 35 kDa proteins that contain a single structural domain. More recently, however, the PITP domain has also been found in the larger rdgB proteins. Soluble PITPs are found in many organisms, including mammals, Caenorhabditis elegans (worms), Drosophila melanogaster (flies), and Dictyostelium discoideum (slime molds), but not in yeasts or plants. Dysfunction of PITPs leads to neurodegeneration; this highlights the need to understand the biochemical and physiological functions of these proteins in cells.
Two families of proteins are able specifically to transfer phosphatidylinositol (PI) and phosphatidylcholine (PC) in eukaryotic cells (Figure 1), namely the PITP and Sec14p families [1]. They share no obvious sequence or structural similarity, but ectopically expressed PITPs can rescue sec14 mutants in yeast. Likewise, in functional studies in mammalian cells, ectopically expressed Sec14p can be used to compensate for the loss of PITPs [2,3]. Whereas Sec14p isoforms are ubiquitous in eukaryotes, members of the PITP family appear to be absent from plants and fungi. It is likely that endogenous Sec14p and PITP family members normally have distinct biological roles, and the two families should not be confused. Accordingly, PITPs define a discrete family, which forms the subject of this review.
Figure 1.
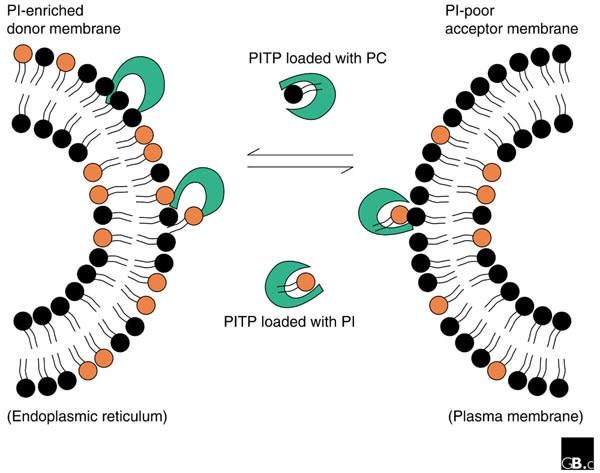
PITPs bind to PI and PC and transfer them between membrane compartments (for example, the endoplasmic reticulum and plasma membrane, as shown). PITPs were first purified by virtue of their ability to transfer PI between two membrane compartments in vitro. PITPs are always occupied by a single molecule of either phosphatidylinositol (PI) or phosphatidylcholine (PC). The affinity for PI is 16-fold higher than for PC, and depending on the relative distribution of PI and PC, PITPs can re-distribute PI at the expense of PC.
Gene organization and evolutionary history
The first mammalian PITP was identified as a 35 kDa protein with 271 amino acids and no sequence similarity to any known protein [4]. Three subfamilies can now be defined (Figure 2) and all isoforms have an amino-terminal PITP-like domain. Non-systematic nomenclature has arisen as a result of the different methods by which each isoform was identified. All three types occur in humans: the first comprises the small PITPα and PITPβ proteins, which were identified by virtue of their transfer activity in vitro; the second comprises the large rdgBα (also called M-RdgB1, Nir2 and PITPnm) and Nir3 (also called M-RdgB2) proteins; and the third type comprises the rdgBβ protein, which is intermediate in size and was identified by homology to rdgBα [5]. The rdgB acronym is derived from a retinal degeneration mutant phenotype (type B) in Drosophila, and the Nir acronym is derived from a reported interaction with the amino-terminal domain of the Pyk2 tyrosine kinase (Pyk2 N-terminal domain-interacting receptor). A third protein, termed Nir1, was also identified, but this lacks a PITP domain [6]. Mammalian rdgBα is 39% identical in sequence to Drosophila rdgB, and mammalian Nir3 is 46% identical to Drosophila rdgB and 56% identical to mammalian rdgBα [7].
Figure 2.
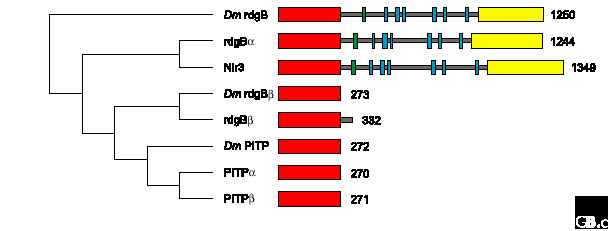
Structural relationships within the PITP family. A ClustalW alignment of human and Drosophila (Dm) PITP sequences was made using MacVector (version 7.0, Oxford Molecular). A bootstrapped dendrogram (1,000 replications) was constructed by rooting with the Drosophila rdgB sequence, using uncorrected, neighbor-joining parameters and ignoring gap sites. The corresponding domain topologies on the right illustrate the number of amino-acid residues in each protein and the position of PITP domains (red), Ca2+-binding domains (green), short hydrophobic regions (blue) and Pyk2-binding domains (yellow).
Intron-exon boundaries are not conserved between different isoforms, and diversity may be enhanced by variation in mRNA splicing [8]. The recent completion of the human genome has revealed that the human PITP chromosomal loci are located at approximately 11q13 (rdgBα), 12q24 (Nir3), 17q13 (PITPα), 17q23-24 (rdgBβ) and 22q12 (PITPβ).
To date, the aforementioned five PITP isoforms have been identified in humans, and one member of each of the three subfamilies can be found in the Drosophila melanogaster genome. Three isoforms can be identified in the genome of C. elegans, including a PITPα or β and a rdgBα or Nir3 homolog; the open reading frame of the third gene is too poorly defined to classify. Two genes have been identified in Dictyostelium discoideum, both of which appear to be PITPα or β homologs [9]. The existence of multiple isoforms within subfamilies appears to be a late event in evolution as, for example, the rat PITPα and PITPβ protein sequences are more similar to each other (77% identity) than to any other PITP. The same is true for murine rdgBα and Nir3 (56% identity) and the Dictyostelium PITP1 and PITP2 sequences (52% identity). Consequently, the different functions of mammalian PITPα and PITPβ will not necessarily parallel the functions of Dictyostelium PITP1 and PITP2; indeed, the subcellular localization of the mammalian and Dictyostelium proteins do not fully correspond [9].
The absence of PITPs from plants, fungi and bacteria is consistent with a role for these proteins in a subset of phosphoinositide signaling pathways unique to animals. It is unlikely, however, that PITPs are functionally redundant in vivo. For example, reduction of PITPα expression in the vibrator mutant mouse leads to neurodegeneration in the presence of normal, ubiquitous PITPβ expression [10]. Furthermore, although the PITP domain of Drosophila rdgBα can rescue the rdgB-null phenotype, rat PITPα is ineffective [11]. Finally, the as-yet undefined functional properties of at least some PITP isoforms appear to be conserved between species as expression of murine rdgBα fully rescues rdgB-null Drosophila [8].
Characteristic structural features
The amino acid sequence of the PITP domain is highly conserved in all isoforms and no characteristic short sequence motifs have been identified. (The sequence can be found under the Pfam IDs PF02121 or IP_trans [12], the PRINTS IDs PR00391 or PITransfer [13] or the INTERPRO ID IPR001666 [14]). The only solved crystal structure in this family is of PITPα bound to PC [15]. On the basis of this structure, PITP domains were suggested to comprise three regions (Figure 3a), of which the amino-terminal lipid-binding region contains the most highly conserved residues. This region contains an eight-stranded, concave, mostly antiparallel β-sheet and two helices (Figure 3b), which encircle the bound lipid and are structurally homologous to the START family of lipid-binding proteins [16]. As there is no apparent sequence similarity between PITPs and START proteins, it appears that these families may have evolved convergently. The carboxy-terminal helical region of the PITP domain shows greatest sequence variation and may play an important role in membrane binding. Finally, the intervening loop region, which contains the Ser166 phosphorylation site, has been suggested to mediate several reported protein-protein interactions [15].
Figure 3.
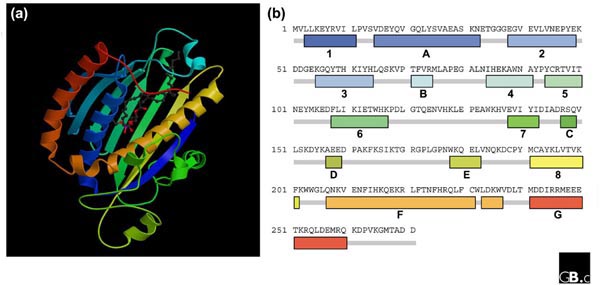
Primary, secondary and tertiary structure of PITPα. The tertiary (a) and primary and secondary (b) structures of rat PITPα, shown bound to sn-1,2-dioleoyl-PC [12], are graded by color between the amino (blue) and the carboxyl (red) termini (see Protein Data Bank (PDB) ID 1FVZ [39]). In (b), the seven α helices and the eight β sheets are labeled A-G and 1-8, respectively.
Assignments to the PITPα/β and rdgBβ subfamilies are made according to the similarity of the whole PITP domain rather than any motif. Although human rdgBβ has a short carboxy-terminal extension, the Drosophila isoform does not. In contrast, the rdgBα/Nir3 subfamily is additionally characterized by the presence of an acidic Ca2+-binding domain (Pfam-B_23582 [12]), six short hydrophobic regions, a putative metal-ion-binding domain (Pfam DDHD [12]), and a carboxy-terminal Pyk2-binding domain [6] (Figure 2). The presence of the Pyk2-binding domain in Drosophila rdgB has been noted to require a frame-shift alteration of the previously published sequence [6]. The hydrophobic regions are probably not transmembrane segments as previously proposed [17], because, although murine rdgBα associates with a particulate brain and retinal fraction on centrifugation, as do other membrane proteins, it can be extracted using buffers containing high salt concentration, high pH or denaturing agents [7]. Sequence features distinguishing individual isoforms in each subfamily remain unclear at present, and may ultimately be better defined by functional information.
Mutation of Thr59, a putative protein kinase C phosphorylation site, in PITPα has been shown to affect PI binding [18], and this residue is completely conserved in all subfamilies. Modeling the position of PI in the PITPα crystal structure suggests that a hydrogen bond may exist between Thr59 and the inositol residue [15]. The only established post-translational modification, however, is phosphorylation of Ser166 in PITPα by protein kinase C activity. Ser166 is conserved in nearly all PITPs and its phosphorylation has been suggested to affect subcellular localization [19].
Localization and function
Although PITPs are defined by their ability to bind one molecule of either PI or PC, the affinity of PITPα for PI is 16-fold greater than for PC. This reflects the lower levels of PI than PC in cells, and typically 40% of the PITPα and β proteins are liganded with PC compared to 60% with PI. In addition to their lipid-binding properties, PITPs can transfer PI or PC from one membrane to another down a concentration gradient without input of energy (Figure 1). Thus, PITPs solubilize lipids from membranes and can facilitate their movement through the aqueous phase.
PITPα and PITPβ are expressed ubiquitously in all tissues. In brain, PITPα is very abundant and can represent 0.1% of brain cytosolic protein [20]. During embryonic and early post-natal stages, expression of the genes for both PITPα and PITPβ is detected widely throughout the entire developing central nervous system. In the adult brain, PITPα is expressed in almost all neurons, whereas the expression of PITPβ is lower than at earlier stages in the entire gray matter regions except for the cerebellar cortex [21]. The cellular concentration of PITPα in brain is estimated at 5-10 μM. Most tissues contain both isoforms, but in neutrophils PITPβ is the major isoform and the cellular concentration of PITPβ in these cells is estimated to be between 5 μM and10 μM. Within cells, PITPs are localized in different compartments: PITPα is present in the cytosol and the nucleus, and PITPβ is localized in the Golgi and cytosol but not the nucleus [9,22].
Genetic data indicate that a decrease in PITPα levels leads to neurodegeneration in the mouse [10]. The vibrator mutation appeared spontaneously in three individuals of a litter of nine DBA/2J mice at the Jackson laboratory in 1961. The homozygous vibrator mutation causes an early-onset progressive action tremor, degeneration of neurons in the brain stem and spinal cord, and juvenile death. The mutation has been identified as an intracisternal-A-particle retroposon insertion in intron 4 of the PITPα gene, causing a fivefold reduction in RNA and protein levels. The vibrator phenotype is suppressed in heterozygous mice and in one intercross. The major suppressor locus, termed Modifier of vibrator-1, maps to proximal chromosome 19 but has not yet been identified.
The biochemical function of PITPα has been extensively examined. Several laboratories have used permeabilized cell preparations from which the cytosolic proteins have been depleted. Measurements of phospholipase-C-mediated signaling [23], regulated exocytosis [24], the formation of secretory granules [25] and acid secretion from gastric glands [26] have been examined, and all these responses were found to be dependent on cytosolic proteins. In all cases, PITPα was purified from brain cytosol as the major reconstituting factor [27]. Several of these functions are summarized in Figure 4. Phospholipase C hydrolyses PI bisphosphate to generate the second messengers (PIP2) diacylglycerol and inositol trisphosphate. Activation of G-protein-coupled receptors or receptor tyrosine kinases is responsible for increasing phospholipase C activity, and PITPα was identified as an essential component in ensuring substrate supply to phospholipase C [28,29]. Analysis of regulated exocytosis was also identified as being dependent on PITPα [18,30,31]. In this case, PITPα functioned together with PI phosphate 5-kinase, indicating that the synthesis of PIP2 was required for the secretory pathway.
Figure 4.
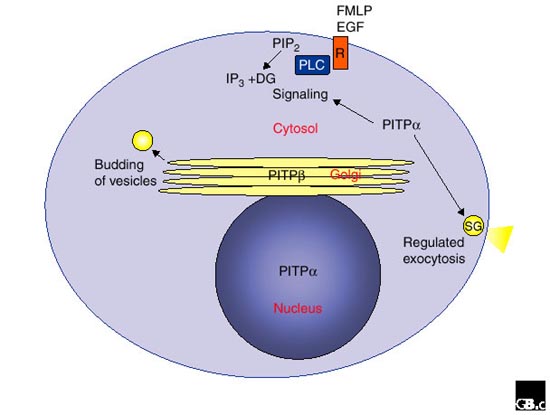
Functions and location of PITPα and PITPβ. PITPα is primarily localized in the cytosol and the nucleus. The major functions of PITP α to be identified are in phospholipase-C-mediated hydrolysis of phosphatidylinositol bisphosphate (PIP2) and in maintaining a pool of PIP2 for exocytosis. PITPβ is primarily localized in the Golgi and the cytosol, and it is involved in the budding of vesicles by making available a pool of phosphoinositides. Abbreviations: DG, diacylglycerol; EGF, epidermal growth factor; FMLP, N-formyl-methionyl-leucyl-phenylalanine; IP3, inositol-1,4,5-trisphosphate; PLC, phospholipase C; R, receptor for EGF and FMLP; SG, secretory granule.
Several reports have demonstrated the ability of PITPα to associate with class I and class III PI 3-kinases [32,33] and type II PI 4-kinase [29] and for rdgBα to associate with type III PI 4-kinase [34]. In addition, rdgBα and Nir3 have been shown to associate with Pyk2, a Ca2+-dependent protein tyrosine kinase [6]. The physiological importance of these different interactions remains unclear, however.
In flies, phototransduction is dependent on a G-protein-coupled phospholipase C signaling pathway and the rdgB protein is essential for phototransduction. The Drosophila rdgB gene was discovered in three independent screens for mutants that exhibited either abnormal photoreceptor physiology or retinal degeneration. The rdgB mutant has a defective light response and its photoreceptors subsequently degenerate, more so in the light. The rdgB gene encodes a 160 kDa protein that is found in the retina, optic lobes, ocelli and in the central brain. In the photoreceptor cell, which consists of an array of densely packed microvilli called the rhabdomere and a non-rhabdomeric cell body, RdgB localizes to the subrhabdomeric cisternae (SRC), which are extensions of the endoplasmic reticulum that act as an intracellular Ca2+ store and also function to deliver protein components and phospholipids such as rhodopsin and PI to the rhabdomere [17]. The SRC and the photoreceptor membrane are in close proximity, separated by a gap of only 10 nm, which may be narrow enough for the transfer of PI between the apposing membranes by the cytoplasmic domain of the rdgB protein [35].
The mouse and human rdgB homologs are strongly expressed in retina, olfactory bulb and brain and moderately expressed in other tissues, including lung, liver, kidney and spleen [36,37]. Mammalian rdgBα can phenotypically rescue Drosophila rdgB mutants. Human rdgBα maps to chromosome 11q13.1, a region known to contain several retinopathy loci, including Best disease and Bardet-Biedl syndrome-1 [8,37]. Furthermore, in the mouse a dramatic increase in rdgBα expression is seen on the day 17 of gestation, when brain development is at its maximum.
Mammalian Nir3 is selectively expressed in neurons, with high levels in the retina and the dentate gyrus of the hippocampus in the mouse. Unlike rdgBα, Nir3 rescues only photoreceptor degeneration without fully restoring the light response, indicating a functional difference between the two RdgB homologs [7].
Frontiers
Proteins with a PITP domain are a relatively small family of PI-binding proteins and belong to a larger superfamily of proteins that bind hydrophobic ligands. Despite a wealth of data indicating that PITP proteins play a central role in membrane traffic and signaling, how they execute their function at the molecular level remains to be understood. PITPs appear to coordinate the levels of phosphoinositides in different membrane compartments during periods when high levels of these lipids are needed for both trafficking and signaling purposes, the way the proteins are harnessed in living cells remains to be understood, however.
At the molecular level, understanding of the mechanism by which a PITP molecule abstracts a lipid from a bilayer and facilitates exchange awaits a detailed analysis of the physical properties of the protein and its ability to interact with membranes. A critical point is that PITPs need a sufficiently high affinity for membranes to allow them to release and exchange their bound lipid, but this affinity should be sufficiently low that the protein can move rapidly away from the membrane. Whether lipid composition or curvature of membranes [38] play roles in modulating this affinity of PITPs for membranes in vivo needs further analysis. Furthermore, how does this biochemical function relate to the requirement for PITPs in membrane budding and fusion? Regulation of these proteins by phosphorylation may provide another important functional determinant. Whether this modification serves as a way of regulating directional transfer of lipids also remains to be studied.
Finally, comprehending the ways in which the regulatory actions of PITPs intertwine with cell signaling and membrane trafficking may provide insights into the neurodegen-eration observed in mouse and fly models that lack PITPs. Here the creation of more refined mouse models with inducible and tissue-specific expression, as well as studies of gene expression during development, will be instrumental in providing insights into the different physiological functions of this versatile protein family.
References
- Wirtz KW. Phospholipid transfer proteins. Annu Rev Biochem. 1991;60:73–99. doi: 10.1146/annurev.bi.60.070191.000445. A comprehensive review of different lipid transfer protein families. [DOI] [PubMed] [Google Scholar]
- Tanaka S, Hosaka K. Cloning of a cDNA encoding a second phosphatidylinositol transfer protein of rat brain by complementation of the yeast sec14 mutation. J Biochem. 1994;115:981–984. doi: 10.1093/oxfordjournals.jbchem.a124448. First cloning of PITPβ from its ability to rescue a temperature-sensitive mutant of the yeast Sec14p PI transfer protein. [DOI] [PubMed] [Google Scholar]
- Cunningham E, Tan SK, Swigart P, Hsuan J, Bankaitis V, Cockcroft S. The yeast and mammalian isoforms of phosphatidylinositol transfer protein can all restore phospholipase C-mediated inositol lipid signalling in cytosol-depleted RBL-2H3 and HL60 cells. Proc Natl Acad Sci USA. 1996;93:6589–6593. doi: 10.1073/pnas.93.13.6589. The similar ability of recombinant Sec14p, PITPa and PITPb to support PLC signaling is demonstrated here, and challenges the notion that PITPs may be enzyme cofactors. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickeson SK, Lim CN, Schuyler GT, Dalton TP, Helmkamp GJ, Yarbrough LR. Isolation and sequence of cDNA clones encoding rat phosphatidylinositol transfer protein. J Biol Chem. 1989;264:16557–16564. First cloning of a PITP, demonstrating that it has a completely novel sequence and that it is widely expressed in rat tissues. [PubMed] [Google Scholar]
- Fullwood Y, dos Santos M, Hsuan JJ. Cloning and characterization of a novel human phosphatidylinositol transfer protein, rdgBβ. J Biol Chem. 1999;274:31553–31558. doi: 10.1074/jbc.274.44.31553. First cloning of a rdgBβ using homology to the Drosophila rdgB sequence. [DOI] [PubMed] [Google Scholar]
- Lev S, Hernandez J, Martinez R, Chen A, Plowman G, Schlessinger J. Identification of a novel family of targets of PYK2 related to Drosophila retinal degeneration B (rdgB) protein. Mol Cell Biol. 1999;19:2278–2288. doi: 10.1128/mcb.19.3.2278. A yeast two-hybrid screen using the amino-terminal domain of Pyk2 led to the identification of three Nir Pyk2-binding proteins with different expression patterns. All bind Ca2+, but the significance of the interaction with Pyk2 remains unknown. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu C, Vihtelic TS, Hyde DR, Li T. A neuronal-specific mammalian homolog of the Drosophila retinal degeneration B gene with expression restricted to the retina and dentate gyrus. J Neurosci. 1999;19:7317–7325. doi: 10.1523/JNEUROSCI.19-17-07317.1999. Cloning of murine m-rdgB-2 (Nir3), demonstration of neuronal-specific expression, and separation of light-response and degeneration phenotypes by differential rescue in Drosophila. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang JT, Milligan S, Li Y, Chew CE, Wiggs J, Copeland NG, Jenkins NA, Campochiaro PA, Hyde DR, Zack DJ. Mammalian homolog of Drosophila retinal degeneration B rescues the mutant fly phenotype. J Neurosci. 1997;17:5881–5890. doi: 10.1523/JNEUROSCI.17-15-05881.1997. Cloning of murine m-rdgB-1 (Nir2), demonstration of neuronal-specific expression, and complete rescue of the rdgB phenotype in Drosophila. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swigart P, Insall R, Wilkins A, Cockcroft S. Purification and cloning of phosphatidylinositol transfer proteins from Dictyostelium discoideum : homologues of both mammalian PITPs and Saccharomyces cerevisiae sec14p are found in the same cell. Biochem J. 2000;347:837–843. doi: 10.1042/0264-6021:3470837. First cloning of PITPs in D. discoideum, showing differential expression during development. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton BA, Smith DJ, Mueller KL, Kerrebrock AW, Bronson RT, van Berkel V, Daly MJ, Kruglyak L, Reeve MP, Nemhauser JL, et al. The vibrator mutation causes neurodegeneration via reduced expression of PITP alpha: positional complementation cloning and extragenic suppression. Neuron. 1997;18:711–722. doi: 10.1016/s0896-6273(00)80312-8. Identification of the first mammalian PITP mutation as a PITPα hypomorph, and mapping of a suppressor locus, Mvb-1, on chromosome 19. The mechanism causing brain-stem and spinal-cord neurodegeneration remains unknown, but an unexpected reduction of neurofilament light chain was detected. [DOI] [PubMed] [Google Scholar]
- Milligan SC, Alb JG, Jr, Elagina RB, Bankaitis VA, Hyde DR. The phosphatidylinositol transfer protein domain of Drosophila retinal degeneration B protein is essential for photoreceptor cell survival and recovery from light stimulation. J Cell Biol. 1997;139:351–363. doi: 10.1083/jcb.139.2.351. A role for Drosophila rdgB in the termination and recovery stages of phototransduction is deduced. Expression of the PITP domain alone, but not of a PC-specific mutant or of rat PITPα, rescued the light and degeneration phenotypes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfam http://www.sanger.ac.uk/Software/Pfam/ A large, searchable collection of multiple sequence alignments and analytical models covering many common protein domains.
- PRINTS http://www.bioinf.man.ac.uk/dbbrowser/PRINTS/ A searchable compendium of protein fingerprints or conserved motifs used to characterize a protein family.
- INTERPRO http://www.ebi.ac.uk/interpro/ InterPro provides an integrated, searchable view of the commonly used signature databases, including Pfam, PRINTS and others.
- Yoder MD, Thomas LM, Tremblay JM, Oliver RL, Yarbrough LR, Helmkamp GM., Jr Structure of a multifunctional protein. Mammalian phosphatidylinositol transfer protein complexed with phosphatidylcholine. J Biol Chem. 2001;276:9246–9252. doi: 10.1074/jbc.M010131200. First three-dimensional definition of PITP structure showing a clear difference from Sec14p. The position of PI remains unknown, but has been modeled. [DOI] [PubMed] [Google Scholar]
- Ponting CP, Aravind L. START: a lipid-binding domain in StAR, HD-ZIP and signalling proteins. Trends Biochem Sci. 1999;24:130–132. doi: 10.1016/s0968-0004(99)01362-6. Identification of the putative, lipid-binding START domain in apparently unrelated proteins by sensitive sequence comparisons. All proteins containing this domain, including the structurally related PITPs, are notably absent from yeast. [DOI] [PubMed] [Google Scholar]
- Vihtelic TS, Goebl M, Milligan S, O'Tousa JE, Hyde DR. Localization of Drosophila retinal degeneration B, a membrane-associated phosphatidylinositol transfer protein. J Cell Biol. 1993;122:1013–1022. doi: 10.1083/jcb.122.5.1013. Drosophila rdgB is found in the endoplasmic reticulum of specific neurons. The PITP and acidic domains are shown to have PI transfer and Ca2+-binding activities, respectively. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alb JJ, Gedvilaite A, Cartee RT, Skinner HB, Bankaitis VA. Mutant rat phosphatidylinositol/phosphatidylcholine transfer proteins specifically defective in phosphatidylinositol transfer: implications for the regulation of phospholipid transfer activity. Proc Natl Acad Sci, USA. 1995;92:8826–8830. doi: 10.1073/pnas.92.19.8826. Random PITPα mutations were screened in a sec14ts yeast background to identify several residues required for PI transfer. A mechanism is proposed by which PI transfer could be regulated in vivo via phosphorylation of PITPα by protein kinase C. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Tiel CM, Westerman J, Paasman M, Wirtz KW, Snoek GT. The protein kinase C-dependent phosphorylation of serine 166 is controlled by the phospholipid species bound to the phosphatidylinositol transfer protein alpha. J Biol Chem. 2000;275:21532–21538. doi: 10.1074/jbc.M002203200. An agonist-dependent protein kinase C phosphorylation site on PITPα is identified at Ser166. A model of PITPα regulation is proposed based on the preferential phosphorylation of the PC-bound form. [DOI] [PubMed] [Google Scholar]
- Venuti SE, Helmkamp GM., Jr Tissue distribution, purification and characterization of rat phosphatidylinositol transfer protein. Biochim Biophys Acta. 1988;946:119–128. doi: 10.1016/0005-2736(88)90464-6. Characterization of physical and lipid transfer properties of PITPα. [DOI] [PubMed] [Google Scholar]
- Utsunomiya A, Owada Y, Yoshimoto T, Kondo H. Localization of gene expression for phosphatidylinositol transfer protein in the brain of developing and mature rats. Brain Res Mol Brain Res. 1997;45:349–352. doi: 10.1016/s0169-328x(97)00030-2. An in situ hybridization study of PITPα and PITPβ expression, which suggests a role for PITPs in neuronal development. [DOI] [PubMed] [Google Scholar]
- De Vries KJ, Westerman J, Bastiaens PI, Jovin TM, Wirtz KW, Snoek GT. Fluorescently labeled phosphatidylinositol transfer protein isoforms (alpha and beta), microinjected into fetal bovine heart endothelial cells, are targeted to distinct intracellular sites. Exp Cell Res. 1996;227:33–39. doi: 10.1006/excr.1996.0246. Ectopic and endogenous PITPα and PITPβ are found predominantly, but not exclusively, in the cytosol and Golgi and the cytosol and nucleus, respectively. [DOI] [PubMed] [Google Scholar]
- Thomas GM, Cunningham E, Fensome A, Ball A, Totty NF, Truong O, Hsuan JJ, Cockcroft S. An essential role for phosphatidylinositol transfer protein in phospholipase C-mediated inositol lipid signaling. Cell. 1993;74:919–928. doi: 10.1016/0092-8674(93)90471-2. First demonstration of an acute requirement for PITPα in phospholipase C-β signaling. [DOI] [PubMed] [Google Scholar]
- Hay JC, Martin TF. Phosphatidylinositol transfer protein required for ATP-dependent priming of Ca2+-activated secretion. Nature. 1993;366:572–575. doi: 10.1038/366572a0. PITP is found to be one of three cytosolic factors required for the ATP-dependent priming of neuroendocrine secretion. A PIP kinase was subsequently found to be another factor (see [30]), but the third factor remains unknown. [DOI] [PubMed] [Google Scholar]
- Ohashi M, Jan de Vries K, Frank R, Snoek G, Bankaitis V, Wirtz K, Huttner WB. A role for phosphatidylinositol transfer protein in secretory vesicle formation. Nature. 1995;377:544–547. doi: 10.1038/377544a0. PITP is found to be one of two cytosolic factors required for the ATP-dependent formation of post-Golgi secretory vesicles in a cell-free preparation. The second factor remains unknown. [DOI] [PubMed] [Google Scholar]
- Akagi K, Nagao T, Urushidani T. Reconstitution of acid secretion in digitonin-permeabilized rabbit gastric glands - identification of cytosolic regulatory factors. J Biol Chem. 2001;276:28171–28178. doi: 10.1074/jbc.M101190200. PITP is found to be one of two cytosolic factors that stimulate gastric secretion from a permeabilised-cell preparation. The second factor remains unknown. [DOI] [PubMed] [Google Scholar]
- Cockcroft S. Phosphatidylinositol transfer proteins: a requirement in signal transduction and vesicle traffic. BioEssays. 1998;20:423–432. doi: 10.1002/(SICI)1521-1878(199805)20:5<423::AID-BIES9>3.3.CO;2-U. A review of PITP, rdgB and Sec14p proteins. [DOI] [PubMed] [Google Scholar]
- Cunningham E, Thomas GM, Ball A, Hiles I, Cockcroft S. Phosphatidylinositol transfer protein dictates the rate of inositol trisphosphate production by promoting the synthesis of PIP2. Curr Biol. 1995;5:775–783. doi: 10.1016/s0960-9822(95)00154-0. A kinetic study of PITPα in PLCβ signaling suggests that PITPα is an enzyme cofactor. [DOI] [PubMed] [Google Scholar]
- Kauffmann ZA, Thomas GM, Ball A, Prosser S, Cunningham E, Cockcroft S, Hsuan JJ. Requirement for phosphatidylinositol transfer protein in epidermal growth factor signaling. Science. 1995;268:1188–1190. doi: 10.1126/science.7761838. PITP is found to be acutely required in phospholipase C-γ signaling and to associate with a signaling complex in response to growth factor stimulation. [DOI] [PubMed] [Google Scholar]
- Hay JC, Fisette PL, Jenkins GH, Fukami K, Takenawa T, Anderson RA, Martin TF. ATP-dependent inositide phosphorylation required for Ca2+-activated secretion. Nature. 1995;374:173–177. doi: 10.1038/374173a0. A PIP kinase is found to be one of three cytosolic factors required for the ATP-dependent priming of neuroendocrine secretion. PITP had previously been found to be another factor (see [24]), but the third factor remains unknown. [DOI] [PubMed] [Google Scholar]
- Fensome A, Cunningham E, Tan SK, Swigart P, Thomas G, Hsuan J, Cockcroft S. ARF and PITP restore GTPγS-stimulated protein secretion from cytosol-depleted HL60 cells by promoting PIP2 synthesis. Curr Biol. 1996;6:730–738. doi: 10.1016/s0960-9822(09)00454-0. PIP 5-kinase activity is found to be stimulated by PITPα and the small GTPase ARF1, the latter via stimulation of phospholipase D. [DOI] [PubMed] [Google Scholar]
- Panaretou C, Domin J, Cockcroft S, Waterfield MD. Characterization of p150, an adaptor protein for the human phosphatidylinositol (PtdIns) 3-kinase. Substrate presentation by phosphatidylinositol transfer protein to the p150-PtdIns 3-kinase complex. J Biol Chem. 1997;272:2477–2485. doi: 10.1074/jbc.272.4.2477. PITPα is found to associate with and stimulate a PI 3-kinase activity found on an intracellular membrane compartment (see also [33]). [DOI] [PubMed] [Google Scholar]
- Jones SM, Alb JG, Jr, Phillips SE, Bankaitis VA, Howell KE. A phosphatidylinositol 3-kinase and phosphatidylinositol transfer protein act synergistically in formation of constitutive transport vesicles from the trans-Golgi network. J Biol Chem. 1998;273:10349–10354. doi: 10.1074/jbc.273.17.10349. PITPα is found to stimulate a GTP-dependent PI 3-kinase activity found in a trans-Golgi network preparation (as the enzyme was PI-specific, it is likely to be the Vps34p homolog studied in [32]). [DOI] [PubMed] [Google Scholar]
- Aikawa Y, Kuraoka A, Kondo H, Kawabuchi M, Watanabe T. Involvement of PITPnm, a mammalian homologue of Drosophila rdgB, in phosphoinositide synthesis on Golgi membranes. J Biol Chem. 1999;274:20569–20577. doi: 10.1074/jbc.274.29.20569. Ectopic, overexpressed Nir2 (rdgBα) was localized to Golgi membranes in COS-7 cells via the hydrophobic regions and the carboxy-terminal domain. The PITP and acidic domains appear to mediate a functional interaction with type III PI 4-kinase to promote the synthesis of PI 4-phosphate. [DOI] [PubMed] [Google Scholar]
- Suzuki E, Hirosawa K. Immunolocalization of a Drosophila phosphatidylinositol transfer protein (rdgB) in normal and rdgA mutant photoreceptor cells with special reference to the subrhabdomeric cisternae. J Electron Microsc. 1994;43:183–189. Immunoelectron microscopy revealed that Drosophila rdgB is localized to the subrhabdomeric cisternae (contiguous with the endoplasmic reticulum) and juxtaposed regions of the rhabdomeric microvilli (part of the plasma membrane of photoreceptor cells). [PubMed] [Google Scholar]
- Aikawa Y, Hara H, Watanabe T. Molecular cloning and characterization of mammalian homologues of the Drosophila retinal degeneration B gene. Biochem Biophys Res Commun. 1997;236:559–564. doi: 10.1006/bbrc.1997.7009. Cloning of human and mouse rdgBα. The murine isoform was mapped close to the Mvb-1 locus in chromosome 19. [DOI] [PubMed] [Google Scholar]
- Guo J, Yu FX. Cloning and characterization of human homologue of Drosophila retinal degeneration B: a candidate gene for degenerative retinal diseases. Dev Genet. 1997;20:235–245. doi: 10.1002/(SICI)1520-6408(1997)20:3<235::AID-DVG6>3.3.CO;2-Z. Cloning of human rdgBα and identification of expression exclusively in rat neuronal cells. The human isoform was mapped on chromosome 11q13, close to several retinopathy loci. [DOI] [PubMed] [Google Scholar]
- Komatsu H, Bouma B, Wirtz KW, Taraschi TF, Janes N. Activity of phosphatidylinositol transfer protein is sensitive to ethanol and membrane curvature. Biochem J. 2000;348:667–673. doi: 10.1042/0264-6021:3480667. Low concentrations of ethanol or chloroform and increasing convex curvature were found to enhance PI transfer by PITPα in vitro. It is suggested that PITPs may preferentially deliver PI to cellular membrane domains that are more convex and/or have a low surface density of lipid. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Protein Data Bank http://www.rcsb.org/pdb/ The single worldwide repository for the processing and distribution of three-dimensional biological macromolecular structure data.