

Abstract
Using a carcinogen-initiated rat model of mammary tumorigenesis, we tested the hypothesis that transforming growth factor (TGF)-βs are useful biomarkers of chemopreventive efficacy in the breast. The chemopreventive agents tested were tamoxifen and the retinoids 9-cis-retinoic acid (9cRA) and N-(4-hydroxyphenyl)retinamide (4-HPR), because both antiestrogens and retinoids have previously been shown to upregulate TGF-βs in vitro. Despite demonstrable chemopreventive efficacy in this model, none of these agents, alone or in combination, had any significant impact on the expression of TGF-βs in the mammary ductal epithelium or periductal stroma as determined by immunohistochemistry. These data suggest that TGF-βs are not likely to be useful biomarkers of chemopreventive efficacy in a clinical setting.
Keywords: antiestrogens, breast cancer, chemoprevention, retinoid, transforming growth factor-βs
Synopsis
Introduction:
Chemoprevention has been defined as the use of noncytotoxic nutrients or pharmacologic agents to enhance intrinsic physiologic mechanisms that protect the organism against the development of mutant clones and their progression to malignant cancer. In a recent landmark trial, tamoxifen, a hormonally active selective estrogen receptor modulator (SERM), was shown to decrease the risk of invasive breast cancer by 49% in asymptomatic, but at-risk women [1]. The search is now on for agents with improved risk-benefit profiles, and for agents that will prevent the subclass of estrogen receptor-negative tumors, the incidence of which was unaffected by the SERMS. Retinoids have already shown potential in this regard [2,3,4,5,6,7]. Because it will not be possible to test many agents in large randomized clinical trials, efforts are underway to develop useful tissue-based surrogate end-point biomarkers that can be used to select only the most promising agents (and doses) for large-scale trials.
Provocative mechanistic connections have been made between the steroid hormone superfamily, including the SERMS and retinoids, and the TGF-β family of multifunctional growth factors [8]. The TGF-β system has tumor suppressor activity, and loss of TGF-β response is associated with advanced disease in many human tumor types, including the breast [9,10]. Conversely, experimental overexpression of TGF-β in the mammary gland protects against tumorigenesis [11]. This strongly suggests that interventions that enhance TGF-β function early in tumorigenesis could delay or prevent the course of the disease. SERMs such as tamoxifen can upregulate TGF-β production and activation by many cell types, including human breast cancer cell lines [12,13,14,15]. Similarly, retinoids can upregulate TGF-β production and activation, both in cell culture and in rats in vivo [16,17]. It is plausible, therefore, that upregulation of endogenous TGF-β could contribute to the chemopreventive efficacy of SERMs and retinoids.
In the present study we used a carcinogen-induced rat mammary carcinogenesis model to test the hypothesis that chemoprevention by tamoxifen and retinoids is associated with local upregulation of TGF-βs in the mammary gland, and that TGF-βs might therefore be useful as potential surrogate end-point biomarkers of chemopreventive efficacy in clinical trials.
Materials and methods:
A standard protocol for induction of breast cancer in female Sprague-Dawley rats using a single dose of N-nitroso-N-methylurea (NMU) at 8 weeks of age was used [4,18]. Chemopreventive agents were incorporated into powdered lab chow [18] and fed ad libitum, beginning 1 week after injection with NMU. The rats were fed 9cRA (Kuraray Company, Osaka, Japan) at 120 mg/kg of diet, tamoxifen (Sigma Chemical Co, St Louis, MO, USA) at 1.0 mg/kg of diet, and 4-HPR (RW Johnson Pharmaceutical Research Unit, Spring House, PA, USA) at 782 mg/kg of diet.
Rats were weighed and palpated for the presence of mammary tumors weekly, and six rats in each experimental group were sacrificed after 6 and 12 weeks of treatment with chemopreventive agent. For experiments to determine the effect of high doses of tamoxifen administered over shorter periods of time, rats were given 10 mg tamoxifen/kg body weight per day intragastrically, or 1 mg tamoxifen/kg in the diet, and were sacrificed after 1 day or 3 weeks of treatment. All palpated tumors were confirmed at necropsy, and mammary glands were fixed in neutral buffered formalin and embedded in paraffin. The number 2 (first thoracic) mammary gland was sectioned for histology and immunohistochemistry.
Immunohistochemical staining was done using rabbit polyclonal antibodies raised against synthetic peptides that correspond to regions in the mature forms of TGF-β1, TGF-β2 and TGF-β3: anti-TGF-β1-LC and anti-TGF-β1-CC [19], anti-TGF-β2 (sc-90; Santa Cruz Biotechnologies Inc, Santa Cruz, CA, USA), and anti-50-60-β3-LC [20], respectively. Anti-latent TGF-β-binding protein (LTBP; Ab39) was raised against the purified full-length platelet LTBP [21]. The antibodies were affinity purified against the immunizing peptide (anti-TGF-β3) or against protein A sepharose (anti-TGF-β1-LC, anti-TGF-β1-CC and anti-TGF-β2). Immunohistochemical staining was performed using an indirect immunoperoxidase detection protocol (Vectastain Elite kit, Vector Laboratories, Burlingame, CA, USA). Staining intensity was scored on a scale of 0-4+, using the mouse embryo control section as a reference standard for each run. Ducts and periductal stroma were scored independently. Staining was scored in a blinded manner by two independent observers, and discrepancies were rescored by consensus. Staining intensity was plotted as the mean ± standard deviation for each experimental group.
Results:
Palpable mammary tumors were first detected after approximately 35 days following initiation of NMU, and by 70 days incidence had reached 100% in rats not treated with chemopreventive agents (Fig. 1a). Tamoxifen, alone or in combination with retinoids, decreased tumor incidence by more than 70% by the end of the study, whereas 9cRA alone decreased it by 50%. 4-HPR alone had a relatively modest effect on tumor incidence in the present study. However, it significantly decreased tumor multiplicity (Fig. 1b), indicating that the dose used was efficacious. There was minimal toxicity associated with the chemopreventive intervention, except in the tamoxifen + 4-HPR group, in which mild toxicity was observed, as judged by the weights of the experimental animals (Fig. 1c).
Figure 1.
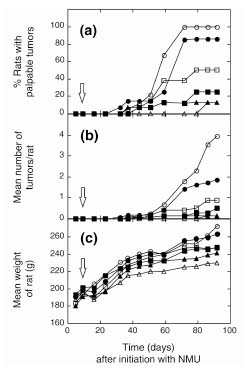
Effect of chemopreventive agents on mammary tumor formation in the NMU rat model of mammary tumorigenesis. Rats were administered NMU at 8 weeks of age (time t = 0), and were randomized into six groups which were given the following chemopreventive agents continuously in their diet, starting 1 week after initiation: no additions (unfilled circle), 4-HPR (filled circle), 9cRA (unfilled square), tamoxifen (filled square), 4-HPR + tamoxifen (unfilled triangle), or 9cRA + tamoxifen (filled triangle). Rats were weighed and palpated for tumors each week. (a) Percentage of rats with palpable tumors. (b) Mean number of tumors per rat. (c) Mean weight of rat. The open arrow indicates the start of the chemopreventive intervention.
All three TGF-β isoforms and the LTBP (part of the naturally occurring latent TGF-β complex) showed broadly similar immunostaining patterns in the mammary glands of untreated rats at 15 weeks of age (Fig. 2). They were present both in the ductal epithelium and in the periductal stroma, suggesting that the TGF-βs are synthesized by the epithelial cells, and possibly stromal cells, and are sequestered in the extracellular matrix. This staining pattern is consistent with a role for the TGF-βs in the maintenance of normal mammary homeostasis.
Figure 2.
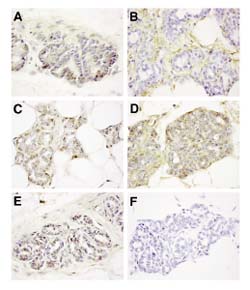
Immunohistochemical analysis of TGF-β expression in the rat mammary gland. The first thoracic (number 2) mammary gland was harvested from 15-week-old rats that had been treated with NMU at age 8 weeks. Sections were immunostained (see Materials and method), with the following antisera: (a) anti-TGF-β1-LC (stains predominantly intracellular TGF-β1); (b) anti-TGF-β1-CC (stains extracellular TGF-β1); (c) anti-TGF-β2; (d) anti-TGF-β3; (e) anti-LTBP; and (f) normal rabbit immunoglobulin. The brown stain indicates a positive immunoperoxidase reaction. Images were shot at 630× original magnification.
None of the chemopreventive agents used, alone or in combination, were found to affect expression of any of the TGF-β isoforms or the LTBP in either ductal epithelium or periductal stroma after 6 weeks of chemopreventive intervention (Fig. 3). The 6-week time point was chosen as representative of the period of preneoplasia, as the majority of the animals had no palpable tumors at this time (Fig. 1). In the study set, eight out of 36 (22%) of the slides showed histologic evidence of hyperplasia, one out of 36 had a ductal carcinoma in situ (mammary intraepithelial neoplasia [22]), and one out of 36 had a carcinoma. We further investigated the effect of tamoxifen at higher doses and earlier time points. In rats that received tamoxifen at 10 mg/kg per day intragastrically (equivalent to 600 mg/day for a human) or 1 mg/kg per day intragastrically (equivalent to 60 mg/day for a human) for either 1 day or 3 weeks, again no consistent changes were seen in TGF-β expression, using either the TGF-β1-CC or the TGF-β2 antibodies (data not shown).
Figure 3.
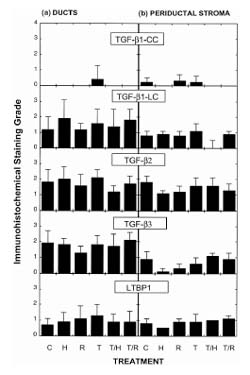
Lack of effect of chemopreventive agents on expression of TGF-βs in the NMU-initiated rat mammary gland. Immunohistochemical staining intensity for TGF-βs and LTBP was determined after 6 weeks of treatment with the following chemopreventive agents: None (C); 4-HPR (H); 9cRA (R); tamoxifen (T); 4-HPR + tamoxifen (T/H); 9cRA + tamoxifen (T/R). Staining intensity was determined independently for the ductal epithelial cells and the periductal stroma (see Materials and method). Staining intensity ranged from 0 to 4 (max). Results are given as the mean ± SD for five or six rats/group. TGF-β1-LC is an antibody that predominantly recognizes intracellular TGF-β1, whereas TGF-β1-CC recognizes extracellular TGF-β1.
After 6 weeks of treatment, we noticed that mammary glands from tamoxifen-treated rats were less developed than those of untreated control animals, having fewer tertiary ducts and terminal end buds, and they could consistently be identified from a blind data set (Fig. 4). By 12 weeks of treatment, all three chemopreventive agents had a significant effect on glandular histology, with tamoxifen and 9cRA showing the greatest suppression of ductal development and lobule formation, and 4-HPR showing a relatively mild effect.
Figure 4.
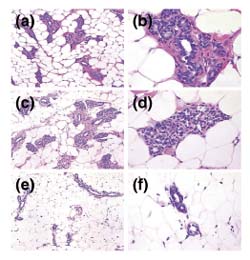
Treatment with tamoxifen affects the histology of the rat mammary gland. Representative hematoxylin and eosin stained sections of the first thoracic gland of 15-week-old rats that had undergone the following treatments: (a, b) No treatment; moderate numbers of mammary gland lobules are present containing primary, secondary and tertiary ductules, as well as developing alveoli. (c, d) Initiation with NMU at 8 weeks of age; no significant histologic differences are noted in mammary gland development from that in untreated control animals. (e, f) Initiation with NMU at 8 weeks, followed by treatment with tamoxifen from 9 to 15 weeks of age; scant numbers of atrophic primary and secondary mammary gland ductules are noted, with no alveolar bud development evident. (a, c, e) Shot at 100×; and (b, d, f) shot at 400× original magnification.
Discussion:
One major goal in the field of prevention is the identification of surrogate biomarkers that might rapidly predict the effect of a given agent on the primary end-point of cancer incidence. The most informative markers are those with modulation that is likely to be directly related to the preventive effect, and a compelling argument can be made that TGF-βs may fall into this category. However, the present data in a well-established preclinical model of breast cancer, employing a variety of highly effective chemopreventive regimens, suggest that this is not the case.
Most of the previous studies on the regulation of TGF-βs by tamoxifen and retinoids have been done in tissue culture [12,13,14,17]. The lack of effect on TGF-β expression in the present in vivo study may reflect the dependence of the response on contextual cues that are only present in the artificial in vitro environment. In an in vivo study [16], all-trans-retinoic acid was shown to cause an upregulation of TGF-β isoforms in rats, with kinetics and isoform selectivity that varied with the target tissue. However, the rats were vitamin A-deficient, and it is not known whether the same effects would be seen in vitamin A-replete animals such as were used in the present study. In a small study in humans [23] tamoxifen treatment was shown to cause a consistent induction in extracellular TGF-β in breast cancer biopsies, when compared with pretreatment biopsies from the same patients, and complex effects of tamoxifen on induction of TGF-β2 in the plasma of patients with metastatic breast cancer have been described [24]. It is possible that tamoxifen is only effective in inducing TGF-β in the context of a tumor, and not in the normal or initiated tissue that was the subject of the present study. However, an optimal surrogate end-point biomarker in a prevention setting needs to be modulated in normal or premalignant tissues. Although we cannot eliminate the possibility of more subtle effects of chemopreventive agents on TGF-β bioavailability or cellular responsiveness, in our preliminary analyses we have seen no effects on the expression of type I and type II TGF-β receptors (data not shown).
There is considerable evidence to suggest that, at late stages in tumorigenesis, TGF-βs can actually promote the tumorigenic process, particularly if the epithelial cells have lost responsiveness to the growth inhibitory effects of TGF-β by this time [9]. While the present work was in progress, a study was reported [25] that showed that loss of the type II TGF-β receptor can already be seen in a significant fraction of hyperplasias without atypia in the human breast. Furthermore, loss of the receptor correlated with increased risk of subsequent development of invasive breast cancer. Thus, loss of TGF-β response may be a very early event in the development of human breast cancer. Because locally elevated TGF-β levels could select for TGF-β-resistant cells, and because TGF-βs can have oncogenic effects on the stroma, it may actually be important for the safety profile of chemopreventive agents to demonstrate that they do not increase TGF-β levels in the at-risk breast. In this regard, this demonstration that the expression of TGF-βs in the preclinical rat model is unaffected by tamoxifen, 9cRA, or 4-HPR may actually have positive implications, because all three agents are already in clinical use.
The NMU-induced rat model of mammary tumorigenesis is widely used for chemoprevention studies, and yields rapid development of hormonally responsive mammary tumors with 100% incidence [2,4,18]. To do this, the initiating agent is given at 8 weeks of age and the chemopreventive agent is started a week later, during the period of active development of the mammary gland. We observed that the histology of the tamoxifen-treated mammary glands differed significantly from control glands when examined after 6 weeks of tamoxifen treatment, showing fewer terminal end-buds and less tertiary branching. Part of the chemopreventive efficacy of antiestrogens and retinoids in this model may therefore be due to a generalized decrease in ductal development. Since chemopreventive agents are unlikely to be given to humans during the pubertal period, this form of preclinical model may not accurately reflect the degree of chemopreventive benefit that could be achieved in humans. Although the accelerated time course and high penetrance of disease reduces the costs of this model, it may be advisable to confirm efficacy of promising agents in a model that delays application of the chemopreventive agent until the mammary gland is fully developed.
In conclusion, we have shown that treatment of rats with tamoxifen or retinoids results in effective chemoprevention of mammary tumorigenesis, without any detectable effect on local expression of TGF-βs. Although we cannot rule out more subtle effects on TGF-β activity, such as the activation of latent forms, the data suggest that TGF-βs are not involved in the underlying molecular mechanism of chemoprevention induced by these agents. This agrees with in vitro work [26] that showed that blockade of TGF-β signaling did not abrogate the growth inhibitory effect of tamoxifen on breast cancer cells. Given the very limited breast tissue available in clinical trials, we do not recommend testing for TGF-βs as a surrogate end-point biomarkers at this time.
Full article
Introduction
Chemoprevention has been defined as the use of noncytotoxic nutrients or pharmacologic agents to enhance intrinsic physiologic mechanisms that protect the organism against the development of mutant clones and their progression to malignant cancer [27]. Members of the nuclear receptor superfamily are considered to be particularly promising targets for chemoprevention, because of their pivotal role in the regulation of metabolic, developmental, and differentiation pathways [28]. In a recent landmark trial [1], tamoxifen, a hormonally active SERM, was shown to decrease the risk of invasive breast cancer by 49% in asymptomatic, but at-risk women. Another SERM, raloxifene, also shows promise [29]. These studies validate the concept of using pharmacologic agents for prevention of human breast cancer in apparently healthy individuals.
The search is now on for agents with improved risk-benefit profiles, and for agents that will prevent the subclass of estrogen receptor-negative tumors, the incidence of which was unaffected by the SERMS. Retinoids, a family of compounds structurally related to vitamin A, have already shown potential in this regard [2,3,4,5,6,7]. Since it will not be possible to test many agents in large randomized clinical trials, efforts are underway to develop useful tissue-based surrogate end-point biomarkers that can be used to select only the most promising agents (and doses) for large-scale trials.
Provocative mechanistic connections have been made between the steroid hormone superfamily, including the SERMS and retinoids, and the TGF-β family of multifunctional growth factors. TGF-βs are potent inhibitors of the growth of many epithelial cell types [8]. Recent work has implicated the TGF-β system as an important tumor suppressor pathway, and loss of TGF-β response is associated with advanced disease in many human tumor types, including the breast [9,10,30,31]. In mouse models, over-expression of TGF-β1 in the mammary gland protects against tumorigenesis [11], whereas local inactivation of the type II TGF-β receptor enhances tumorigenesis [32]. This strongly suggests that interventions that enhance TGF-β function early in tumorigenesis could delay or prevent the course of the disease.
Antiestrogens such as tamoxifen have been shown to upregulate TGF-β production and activation by many cell types, including human breast cancer cell lines [12,13,14,15]. Similarly, retinoids can upregulate TGF-β production and activation, both in cell culture and in rats in vivo [16,17]. Therefore, it is reasonable to propose that some of the chemopreventive efficacy of these agents against breast cancer in vivo could be mediated via a local upregulation of TGF-βs, with concomitant enhancement of tumor suppressor activity.
In the present study, we used a carcinogen-induced rat model of mammary carcinogenesis to test whether chemoprevention by tamoxifen and by two different retinoids (4-HPR, also known as fenretinide; and 9-cRA) is associated with local upregulation of TGF-βs in the initiated mammary gland. If this were the case, TGF-βs might be useful as potential surrogate end-point biomarkers in clinical trials. However, the results show that TGF-β levels, as detected immunohistochemically, are not affected by tamoxifen or retinoids in this preclinical model of early-stage breast cancer.
Materials and method
Mammary carcinogenesis studies
A standard protocol for induction of breast cancer in female Sprague-Dawley rats was used [4,18], with initiation induced by a single intravenous dose of NMU (50 mg/kg body weight) at 8 weeks of age. Retinoids and tamoxifen were incorporated into powdered laboratory chow as described previously [18] and fed ad libitum, beginning 1 week after injection with NMU. Rats were fed 9cRA (Kuraray Company, Osaka, Japan) at 120 mg/kg of diet, tamoxifen (Sigma Chemical Co, St Louis, MO, USA) at 1.0 mg/kg of diet, and 4-HPR (RW Johnson Pharmaceutical Research Institute) at 782 mg/kg of diet.
Rats were weighed weekly and palpated for the presence of mammary tumors. Six rats in each experimental group were sacrificed after 6 and 12 weeks of treatment with chemopreventive agent. The 6-week sacrifice time was chosen for the immunohistochemical studies to represent the period of premalignancy, because the incidence of palpable tumors is less than 20% for all experimental groups at that time. By 12 weeks all rats that have not received a chemopreventive agent have tumors, so the primary purpose of the 12-week sacrifice time was to allow an accurate determination of chemopreventive efficacy for the particular experiment.
For experiments to determine the effect of high doses of tamoxifen administered over shorter periods of time, rats were given 10 mg tamoxifen/kg body weight per day intragastrically or 1 mg tamoxifen/kg in the diet, and were sacrificed after 1 day or 3 weeks of treatment.
All palpated tumors were confirmed at necropsy, and mammary glands were fixed in neutral buffered formalin and embedded in paraffin. The number 2 (first thoracic) mammary gland was sectioned for histology and immunohistochemistry.
Immunohistochemistry of TGF-βs
Immunohistochemical staining was done using rabbit polyclonal antibodies raised against synthetic peptides that correspond to regions in the mature forms of TGF-β1, TGF-β2, and TGF-β3. Antibodies to TGF-β1 were raised against a synthetic peptide corresponding to residues 1-30 of the mature protein in either the Laboratory of Chemoprevention (anti-TGF-β1-LC; NIH, Bethesda, MD, USA) or the Collagen Corporation (anti-TGF-β1-CC; Palo Alto, CA, USA). These antibodies were raised against different preparations of the 1-30 peptide, and they recognize different epitopes of this peptide [19]. The LC antibody usually stains intracellular TGF-β1, whereas the CC antibody stains extracellular TGF-β1. Anti-TGF-β2 (sc-90; Santa Cruz Biotechnologies Inc) was raised to a peptide corresponding to residues 72-99 of the mature TGF-β2. Anti-TGF-β3 (anti-50-60-β3-LC) was raised against residues 50-60 of mature TGF-β3 [20]. Anti-LTBP (Ab39) was raised against the purified full-length platelet LTBP [21].
The antibodies were affinity purified against the immunizing peptide (anti-TGF-β3) or against protein A sepharose (anti-TGF-β1-LC, anti-TGF-β1-CC, and anti-TGF-β2), and have been assayed for specificity by Western blot analysis [19,20,33]. All antibodies reacted with the appropriate TGF-β isoform except anti-TGF-β1-CC, which showed some cross-reactivity with TGF-β3 on Western blots.
Immunohistochemical staining was performed using an indirect immunoperoxidase detection protocol (Vectastain Elite kit; Vector Laboratories) following treatment of sections with hyaluronidase to improve antibody penetration. Optimal antibody concentrations were determined by titration on select samples before analysis of the full experimental set. Staining was shown to be specific in control experiments in which either the primary antibody was preincubated with a 50-fold molar excess of immunizing peptide before being applied to the section (anti-TGF-β2, anti-TGF-β3), or the section was stained with an equivalent concentration of nonimmune rabbit immunoglobulin (anti-TGF-β1-LC, anti-TGF-β1-CC, and anti-LTBP). In analysis of the full experimental set, for any given antibody all sections were stained at the same time so as to be directly comparable, and a normal mouse embryo section was included as a positive control. A normal rabbit immunoglobulin control was also run for the whole set.
Quantitation of immunostaining
Two different systems were used to grade immunostaining. For all samples, staining of the ducts and periductal stroma were scored independently. For samples after 6 weeks of chemopreventive treatment, staining intensity was scored on a scale of 0–4+, using the mouse embryo control section as a reference standard for each run. Staining was scored in a blinded manner by two independent observers, and scores never differed by more than one point. Discrepancies were rescored by consensus. Staining intensity was plotted as the mean ± standard deviation for each experimental group.
Results
Chemopreventive efficacy
Palpable mammary tumors were first detected after approximately 35 days following initation with NMU, and by 70 days incidence had reached 100% in rats not treated with chemopreventive agents (Fig. 1a). Tamoxifen, alone or in combination with retinoids, decreased tumor incidence by more than 70% by the end of the study, whereas 9cRA alone decreased it by 50%. 4-HPR alone had a relatively modest effect on tumor incidence in the present study. However, it significantly decreased tumor multiplicity (Fig. 1b) indicating that the dose used was efficacious. These incidence and multiplicity data are consistent with previously published studies using this model [2,4,34]. The mean weights of the rats in the various experimental groups differed by less than 10% from those of rats not treated with a chemopreventive agent, except for the rats treated with a combination of tamoxifen and 4-HPR, in which the mean weight was approximately 15% lower than the that in untreated group by the end of the study (Fig. 1c). This suggests that there was minimal toxicity associated with the chemopreventive intervention, except in the tamoxifen + 4-HPR group, in which mild toxicity was observed.
Effect of chemopreventive agents on TGF-β expression in initiated mammary gland
Figure 2 shows the typical immunohistochemical staining pattern for the TGF-βs and the LTBP (part of the naturally occurring latent TGF-β complex) in initiated mammary glands of 15-week-old rats that had not been treated with chemopreventive agents. All three TGF-β isoforms and the LTBP showed broadly similar staining patterns. They were present both in the ductal epithelium and in the periductal stroma, suggesting that the TGF-βs are synthesized by the epithelial cells, and possibly stromal cells, and are sequestered in the extracellular matrix. This staining pattern is consistent with a role for the TGF-βs in the maintenance of normal mammary homeostasis.
None of the chemopreventive agents used, alone or in combination, were found to affect expression of any of the TGF-β isoforms or the LTBP in either ductal epithelium or periductal stroma after 6 weeks of chemopreventive intervention (Fig. 3). The 6-week time point was chosen as representative of the period of preneoplasia, because the majority (>80%) of the rats had no palpable tumors at this stage (Fig. 1). Human clinical material for biomarker analysis in primary chemoprevention studies is also likely to comprise normal and initiated, at-risk epithelium with some early preneoplastic changes, but without evidence of major neoplastic change.
In the 6-week study set, eight out of 36 (22%) of the glands sampled showed histologic evidence of hyperplasia. In addition, one out of 36 glands sampled had a lesion with the appearance of a ductal carcinoma in situ (focal necrosis, marked atypia and abundant mitoses, placing it in the category of 'mammary intraepithelial neoplasia' by the Annapolis naming convention [22]), and an additional one had an invasive carcinoma. Both the in situ and the invasive carcinomas were in the control group. However, both samples also had histologically normal ducts on the same slide, which were scored for the analysis. There was no difference in staining between the ducts that were proximal to the tumor and those that were more distal, and neither were there any differences in staining observed between histologically normal and hyperplastic ducts in any of the samples analyzed (data not shown). Since the focus of the present study was on TGF-β expression changes in the preneoplastic gland, staining of the tumors was not scored for the analysis, but in the two cases present, the staining did not differ significantly from that of the surrounding normal-appearing ducts (not shown).
Assuming that a rat weighs approximately 250 g and eats 10 g of chow each day, the equivalent human doses to those used in this study would be approximately 2.5 mg/day tamoxifen, 2 g/day 4-HPR, and 290 mg/day 9cRA. The human doses that are currently being used in clinical trials are 20 mg/day for tamoxifen, 0.2-0.4 g/day for 4-HPR, and 100-250 mg/day for 9cRA. The different doses of tamoxifen and 4-HPR used in humans and rats probably reflect interspecies differences in pharmacokinetics. However, because the efficacious dose for tamoxifen was 10 times lower in the rat than the dose that has previously been shown to upregulate TGF-β1 expression in human breast tumor tissue [23], we looked at the effect of a higher dose of tamoxifen in the rats. Furthermore, because one study [35] showed that TGF-β may be transiently upregulated early after administration of tamoxifen, we also looked at earlier time points. In rats receiving tamoxifen at 10 mg/kg per day intragastrically (equivalent to 600 mg/day for a human) or 1 mg/kg per day (equivalent to 60 mg/day for a human) for either 1 day or 3 weeks, again no consistent changes were seen in TGF-β expression, using either the TGF-β1-CC or the TGF-β2 antibodies (data not shown).
Effect of chemopreventive agents on the histology of the mammary gland
While scoring the immunohistochemical slides, we noticed that the mammary histology appeared to be altered in rats treated with chemopreventive agents. Hematoxylin and eosin stained sections from the samples after 6 weeks of treatment were analyzed by a veterinary pathologist (MAE). The mammary glands from tamoxifen-treated rats were less developed than those of untreated control animals, having fewer tertiary ducts and terminal end-buds, and could consistently be identified from a blind data set (Fig. 4). By 12 weeks of treatment, all three chemopreventive agents had a significant effect on glandular histology, with tamoxifen and 9cRA showing the greatest suppression of ductal development and lobule formation, and 4-HPR showing a relatively mild effect.
Discussion
TGF-βs as candidate biomarkers
One major goal in the field of prevention is the identification of surrogate biomarkers that might rapidly predict the effect of a given agent on the primary end-point of cancer incidence. There is an extensive literature showing that steroid hormone superfamily members, such as antiestrogens and retinoids, can upregulate TGF-β activity in a variety of systems [12,13,14,15,16,17,23,24,35]. This suggested that the chemopreventive action of these agents against breast cancer could be mediated in part through enhancing the tumor suppressor activity of the endogenous TGF-β system, and thus that changes in TGF-β expression might serve as useful surrogate end-point biomarkers of chemopreventive efficacy. However, here we used the NMU-induced rat model of mammary carcinogenesis to show that the chemopreventive effect of tamoxifen and two retinoids is not associated with any consistent changes in TGF-β levels, at least as determined immunohistochemically.
Comparison with earlier studies
Most of the previous studies on the regulation of TGF-βs by tamoxifen and retinoids [12,13,14,17] have been done in tissue culture. Thus, the lack of effect on TGF-β expression in the present in vivo study might reflect the dependence of the response on contextual cues that are only present in the artificial in vitro environment. In one previous in vivo study [16] all-trans-retinoic acid upregulated TGF-β expression in rats, with kinetics and isoform selectivity that varied with the target tissue. However, the rats were vitamin A-deficient, and it is not known whether the same effects would be seen in vitamin A-replete animals, such as were used in the present study, or whether the response would vary with the specific retinoid used.
There are some data that support an effect of tamoxifen on upregulation of TGF-βs in vivo in humans. Three months of tamoxifen treatment was shown to cause a consistent induction in extracellular TGF-β in breast cancer biopsies, when compared with pretreatment biopsies from the same patients [23]. Furthermore, complex effects of tamoxifen on induction of TGF-β2 in the plasma of patients with metastatic breast cancer have been described [24]. It is possible that tamoxifen is only effective in inducing TGF-β in the context of a tumor, and not in normal or initiated tissue, which was the subject of the present study. This issue could be reassessed in preclinical models using the same agents to treat established proliferative intraepithelial neoplasia. However, for ease of tissue acquisition, an optimal surrogate end-point biomarker in a prevention setting needs to be modulated in normal or premalignant tissues.
Alternative levels of regulation of the bioefficacy of TGF-βs
Interestingly, in cell culture, both tamoxifen and all-trans-retinoic acid have been shown [12,13,14,17] to increase the fraction of TGF-β in its biologically active, as opposed to its latent form. The current method for discriminating between active and latent TGF-β in tissue samples requires the use of frozen sections and immunofluorescence techniques, which are not practical for routine clinical use [36]. As simpler assays become available, however, the issue of possible changes in TGF-β activation status should be readdressed. Retinoids can also affect cellular responsiveness to TGF-βs at the level of receptor expression and downstream events [37,38]. To date, expression of TGF-β receptors and downstream signaling components such as the Smads have not been well-characterized in this rat model, but in our preliminary analyses we saw no effect of retinoids on type I and type II TGF-β receptor expression in the mammary gland (data not shown). At this time, however, we certainly cannot rule out the possibility that tamoxifen and retinoids may be having subtle effects on the TGF-β system at levels other than the regulation of TGF-β expression.
Lack of effect of chemopreventive agents on TGF-β expression may have positive implications
There is considerable evidence to suggest that, at late stages in tumorigenesis, TGF-βs can actually promote the tumorigenic process, particularly if the epithelial cells have lost responsiveness to the growth regulatory effects of TGF-β by this time [9,39,40,41]. Thus, advanced human tumors show increased levels of TGF-β expression [42,43,44], and TGF-βs are known to suppress the immunosurveillance system, to enhance angiogenesis, invasion and metastasis, and to increase drug resistance [45,46,47,48].
In the colon, loss of the type II TGF-β receptor occurs at the late adenoma to carcinoma transition [49], suggesting that early premalignant lesions retain TGF-β responsiveness and would be amenable to interventions that enhance TGF-β activity. However, while the present work was in progress, a study was reported [25] showing that loss of the type II TGF-β receptor can already be seen in a significant fraction of hyperplasias without atypia in the human breast. Furthermore, loss of the receptor correlated with increased risk of subsequently developing invasive breast cancer. Thus, unlike in the colon, loss of TGF-β response may be a very early event in the development of human breast cancer.
Since locally elevated TGF-β levels may select for TGF-β-resistant cells, and because TGF-βs can have oncogenic effects on the stroma, it may actually be important for the safety profile of chemopreventive agents to demonstrate that they do not increase TGF-β levels in the at-risk breast. For example, tamoxifen resistance in a xenograft model of advanced human breast cancer, was recently shown [50] to be associated with an increase in TGF-βs and concomitant immunosuppressive effects on natural killer cells. In this regard, our demonstration that the expression of TGF-βs in the preclinical rat model is unaffected by tamoxifen, 9cRA, and 4-HPR may actually have positive implications, because these agents are already in clinical use.
Limitations of the NMU-induced rat model of mammary carcinogenesis
The NMU-induced rat model of mammary tumorigenesis is widely used for chemoprevention studies and yields rapid development of hormonally responsive mammary tumors with 100% incidence [2,4,18]. To do this, the initiating agent is given at 8 weeks of age, during early puberty, and the chemopreventive agent is typically given continuously, starting 1 week later. Since sexual maturity is achieved at approximately 11 weeks of age in rats, this means that the chemopreventive agent is given during a period of active development of the mammary gland.
In the present study we observed that the histology of the tamoxifen-treated mammary gland differed significantly from control glands when examined after 6 weeks of tamoxifen treatment. Specifically, there were fewer terminal end-buds and less tertiary branching, which are indicative of a delay or arrest in normal mammary development. This is consistent with the known requirement for estrogen for proper mammary development [51]. We saw a lesser effect with 4-HPR, although this type of phenomenon has also been described in the literature for retinoids [4]. Thus, part of the chemopreventive efficacy of antiestrogens and retinoids in this model may be due to a generalized decrease in ductal development. Because chemopreventive agents are unlikely to be given to humans during the pubertal period, this form of preclinical model may not accurately reflect the degree of chemopreventive benefit that could be achieved in humans. Although the accelerated time course and high penetrance of disease reduces the costs of this model, it may be advisable to confirm efficacy of promising agents in a model that delays application of the chemopreventive agent until the mammary gland is fully developed.
Conclusion
We have shown that treatment of rats with tamoxifen or retinoids results in effective chemoprevention of mammary tumorigenesis, without any detectable effect on local expression of TGF-βs. Although we cannot rule out more subtle effects on TGF-β activity, such as the activation of latent forms, the data suggest that the underlying molecular mechanism of chemoprevention by these agents does not involve increases in TGF-β expression. This agrees with in vitro work showing that blockade of TGF-β signalling did not abrogate the growth inhibitory effect of tamoxifen on breast cancer cells [26]. Given the very limited breast tissue available in clinical chemoprevention trials, we do not recommend testing for TGF-βs as surrogate end-point biomarkers at this time.
Acknowledgments
Acknowledgements
We thank Drs Calle Heldin and Kohei Miyazono for the generous gift of Ab39, and Dr Clint Grubbs for the rat tissues treated with high levels of tamoxifen for short durations. We are grateful to Dr Donald Gardner for veterinary pathology expertise, to Dr Michael Sporn for many inspiring discussions on the concept and practice of chemoprevention, and to Dr Anita Roberts for continued support and guidance. This study was funded in part by an American Society of Clinical Oncology Young Investigator Award to Dr Zujewski.
References
- Fisher B, Costantino JP, Wickerham DL, Redmond CK, Kavanah M, Cronin WM, Vogel V, Robidoux A, Dimitrov N, Atkins J, Daly M, Wieand S, Tan-Chiu E, Ford L, Wolmark N. Tamoxifen for prevention of breast cancer: report of the National Surgical Adjuvant Breast and Bowel Project P-1 Study. J Natl Cancer Inst. 1998;90:1371–1388. doi: 10.1093/jnci/90.18.1371. [DOI] [PubMed] [Google Scholar]
- Anzano MA, Byers SW, Smith JM, Peer CW, Mullen LT, Brown CC, Roberts AB, Sporn MB. Prevention of breast cancer in the rat with 9-cis-retinoic acid as a single agent and in combination with tamoxifen. Cancer Res. 1994;54:4614–4617. [PubMed] [Google Scholar]
- De Palo G, Camerini T, Marubini E, Costa A, Formelli F, Del Vecchio M, Mariani L, Miceli R, Mascotti G, Magni A, Campa T, Di Mauro MG, Attili A, Maltoni C, Del Turco MR, Decensi A, D'Aiuto G, Veronesi U. Chemoprevention trial of contralateral breast cancer with fenretinide. Rationale, design, methodology, organization, data management, statistics and accrual. Tumori. 1997;83:884–894. doi: 10.1177/030089169708300603. [DOI] [PubMed] [Google Scholar]
- Moon RC, Thompson HJ, Becci PJ, Grubbs CJ, Gander RJ, Newton DL, Smith JM, Phillips SL, Henderson WR, Mullen LT, Brown CC, Sporn MB. N-(4-Hydroxyphenyl)retinamide, a new retinoid for prevention of breast cancer in the rat. Cancer Res. 1979;39:1339–1346. [PubMed] [Google Scholar]
- Anzano MA, Peer CW, Smith JM, Mullen LT, Shrader MW, Logsdon DL, Driver CL, Brown CC, Roberts AB, Sporn MB. Chemoprevention of mammary carcinogenesis in the rat: combined use of raloxifene and 9-cis-retinoic acid. J Natl Cancer Inst. 1996;88:123–125. doi: 10.1093/jnci/88.2.123. [DOI] [PubMed] [Google Scholar]
- Gottardis MM, Bischoff ED, Shirley MA, Wagoner MA, Lamph WW, Heyman RA. Chemoprevention of mammary carcinoma by LGD1069 (Targretin): an RXR-selective ligand. Cancer Res. 1996;56:5566–5570. [PubMed] [Google Scholar]
- Veronesi U, De Palo G, Marubini E, Costa A, Formelli F, Mariani L, Decensi A, Camerini T, Del Turco MR, Di Mauro MG, Muraca MG, Del Vecchio M, Pinto C, D'Aiuto G, Boni C, Campa T, Magni A, Miceli R, Perloff M, Malone WF, Sporn MB. Randomized trial of fenretinide to prevent second breast malignancy in women with early breast cancer. J Natl Cancer Inst. 1999;91:1847–1856. doi: 10.1093/jnci/91.21.1847. [DOI] [PubMed] [Google Scholar]
- Roberts AB, Sporn MB. The transforming growth factors-β. Handbook of Experimental Pharmacology. Peptide Growth Factors and Their Receptors. Edited by Sporn MB, Roberts AB. Berlin: Springer-Verlag; 1990;95(1):419–472. [Google Scholar]
- Reiss M. Transforming growth factor-β and cancer: a love-hate relationship? Oncol Res. 1997;9:447–457. [PubMed] [Google Scholar]
- Gobbi H, Arteaga CL, Jensen RA, Simpson JF, DuPont WD, Olson SJ, Schuyler PA, Plummer WDJ, Page DL. Loss of expression of transforming growth factor β type II receptor correlates with high tumour grade in human breast in-situ and invasive carcinomas. Histopathology. 2000;36:168–177. doi: 10.1046/j.1365-2559.2000.00841.x. [DOI] [PubMed] [Google Scholar]
- Pierce DF, Jr, Gorska AE, Chytil A, Meise KS, Page DL, Coffey RJ Jr, Moses HL. Mammary tumor suppression by transforming growth factor β 1 transgene expression. Proc Natl Acad Sci USA. 1995;92:4254–4258. doi: 10.1073/pnas.92.10.4254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colletta AA, Wakefield LM, Howell FV, van Roozendaal KE, Danielpour D, Ebbs SR, Sporn MB, Baum M. Anti-oestrogens induce the secretion of active transforming growth factor β from human fetal fibroblasts. Br J Cancer. 1990;62:405–409. doi: 10.1038/bjc.1990.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colletta AA, Wakefield LM, Howell FV, Danielpour D, Baum M, Sporn MB. The growth inhibition of human breast cancer cells by a novel synthetic progestin involves the induction of transforming growth factor β. J Clin Invest. 1991;87:277–283. doi: 10.1172/JCI114983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knabbe C, Lippman ME, Wakefield LM, Flanders KC, Kasid A, Derynck R, Dickson RB. Evidence that transforming growth factor-β is a hormonally regulated negative growth factor in human breast cancer cells. Cell. 1987;48:417–428. doi: 10.1016/0092-8674(87)90193-0. [DOI] [PubMed] [Google Scholar]
- Koli KM, Ramsey TT, Ko Y, Dugger TC, Brattain MG, Arteaga CL. Blockade of transforming growth factor-β signaling does not abrogate antiestrogen-induced growth inhibition of human breast carcinoma cells. J Biol Chem. 1997;272:8296–8302. doi: 10.1074/jbc.272.13.8296. [DOI] [PubMed] [Google Scholar]
- Glick AB, Abdulkarem N, Flanders KC, Lumadue JA, Smith JM, Sporn MB. Complex regulation of TGFβ expression by retinoic acid in the vitamin A-deficient rat. Development. 1991;111:1081–1086. doi: 10.1242/dev.111.4.1081. [DOI] [PubMed] [Google Scholar]
- Glick AB, Flanders KC, Danielpour D, Yuspa SH, Sporn MB. Retinoic acid induces transforming growth factor-β2 in cultured keratinocytes and mouse epidermis. Cell Regul. 1989;1:87–97. doi: 10.1091/mbc.1.1.87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anzano MA, Smith JM, Uskokovic MR, Peer CW, Mullen LT, Letterio JJ, Welsh MC, Shrader MW, Logsdon DL, Driver CL. 1 alpha,25-Dihydroxy-16-ene-23-yne-26,27-hexafluorocholecal-ciferol (Ro24/-5531), a new deltanoid (vitamin D analogue) for prevention of breast cancer in the rat. Cancer Res. 1994;54:1653–1656. [PubMed] [Google Scholar]
- Flanders KC, Thompson NL, Cissel DS, Van Obberghen-Schilling E, Baker CC, Kass ME, Ellingsworth LR, Roberts AB, Sporn MB. Transforming growth factor-β1: histochemical localization with antibodies to different epitopes. J Cell Biol. 1989;108:653–660. doi: 10.1083/jcb.108.2.653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flanders KC, Ludecke G, Engels S, Cissel DS, Roberts AB, Kondaiah P, Lafyatis R, Sporn MB, Unsicker K. Localization and actions of transforming growth factor-βs in the embryonic nervous system. Development. 1991;113:183–191. doi: 10.1242/dev.113.1.183. [DOI] [PubMed] [Google Scholar]
- Kanzaki T, Olofsson A, Moren A, Wernstedt C, Hellman U, Miyazono K, Claesson-Welsh L, Heldin CH. TGF-β1 binding protein: a component of the large latent complex of TGF-β1 with multiple repeat sequences. Cell. 1990;61:1051–1061. doi: 10.1016/0092-8674(90)90069-q. [DOI] [PubMed] [Google Scholar]
- Cardiff RD, Anver MR, Gusterson BA, Hennighausen L, Jensen RA, Merino MJ, Rehm S, Russo J, Tavassoli FA, Wakefield LM, Ward JM, Green JE. The mammary pathology of genetically engineered mice: the consensus report and recommendations from the Annapolis meeting. Oncogene. 2000;19:968–988. doi: 10.1038/sj.onc.1203277. [DOI] [PubMed] [Google Scholar]
- Butta A, MacLennan K, Flanders KC, Sacks NP, Smith I, McKinna A, Dowsett M, Wakefield LM, Sporn MB, Baum M, Colletta AA. Induction of transforming growth factor β1 in human breast cancer in vivo following tamoxifen treatment. Cancer Res. 1992;52:4261–4264. [PubMed] [Google Scholar]
- Kopp A, Jonat W, Schmahl M, Knabbe C. Transforming growth factor β2 (TGF- β2) levels in plasma of patients with metastatic breast cancer treated with tamoxifen. Cancer Res. 1995;55:4512–4515. [PubMed] [Google Scholar]
- Gobbi H, DuPont WD, Simpson JF, Plummer WDJ, Schuyler PA, Olson SJ, Arteaga CL, Page DL. Transforming growth factor-β and breast cancer risk in women with mammary epithelial hyperplasia. J Natl Cancer Inst. 1999;91:2096–2101. doi: 10.1093/jnci/91.24.2096. [DOI] [PubMed] [Google Scholar]
- Koli KM, Ramsey TT, Ko Y, Dugger TC, Brattain MG, Arteaga CL. Blockade of transforming growth factor-β signaling does not abrogate antiestrogen-induced growth inhibition of human breast carcinoma cells. J Biol Chem. 1997;272:8296–8302. doi: 10.1074/jbc.272.13.8296. [DOI] [PubMed] [Google Scholar]
- Sporn MB. Chemoprevention of cancer. Lancet. 1993;342:1211–1213. doi: 10.1016/0140-6736(93)92189-z. [DOI] [PubMed] [Google Scholar]
- Prevention of cancer in the next millennium: report of the Chemoprevention Working Group to the American Association for Cancer Research. Cancer Res. 1999;59:4743–4758. [PubMed] [Google Scholar]
- Cummings SR, Eckert S, Krueger KA, Grady D, Powles TJ, Cauley JA, Norton L, Nickelsen T, Bjarnason NH, Morrow M, Lippman ME, Black D, Glusman JE, Costa A, Jordan VC. The effect of raloxifene on risk of breast cancer in postmenopausal women: results from the MORE randomized trial. Multiple Outcomes of Raloxifene Evaluation. JAMA. 1999;281:2189–2197. doi: 10.1001/jama.281.23.2189. [DOI] [PubMed] [Google Scholar]
- Markowitz SD, Roberts AB. Tumor suppressor activity of the TGF-β pathway in human cancers. Cytokine Growth Factor Rev. 1996;7:93–102. doi: 10.1016/1359-6101(96)00001-9. [DOI] [PubMed] [Google Scholar]
- Hata A, Shi Y, Massague J. TGF-β signaling and cancer: structural and functional consequences of mutations in smads. Mol Med Today. 1998;4:257–262. doi: 10.1016/s1357-4310(98)01247-7. [DOI] [PubMed] [Google Scholar]
- Bottinger EP, Jakubczak JL, Haines DC, Bagnall K, Wakefield LM. Transgenic mice overexpressing a dominant-negative mutant type II transforming growth factor β receptor show enhanced tumorigenesis in the mammary gland and lung in response to the carcinogen 7,12-dimethylbenz-[a]-anthracene. Cancer Res. 1997;57:5564–5570. [PubMed] [Google Scholar]
- Flanders KC, Roberts AB, Ling N, Fleurdelys BE, Sporn MB. Antibodies to peptide determinants in transforming growth factor β and their applications. Biochemistry. 1988;27:739–746. doi: 10.1021/bi00402a037. [DOI] [PubMed] [Google Scholar]
- McCormick DL, Moon RC. Retinoid-tamoxifen interaction in mammary cancer chemoprevention. Carcinogenesis. 1986;7:193–196. doi: 10.1093/carcin/7.2.193. [DOI] [PubMed] [Google Scholar]
- Grainger DJ, Witchell CM, Metcalfe JC. Tamoxifen elevates transforming growth factor-β and suppresses diet-induced formation of lipid lesions in mouse aorta. Nature Med. 1995;1:1067–1073. doi: 10.1038/nm1095-1067. [DOI] [PubMed] [Google Scholar]
- Barcellos-Hoff MH, Ehrhart EJ, Kalia M, Jirtle R, Flanders K, Tsang ML. Immunohistochemical detection of active transforming growth factor-β in situ using engineered tissue. Am J Pathol. 1995;147:1228–1237. [PMC free article] [PubMed] [Google Scholar]
- Yoshizawa M, Miyazaki H, Kojima S. Retinoids potentiate transforming growth factor-β activity in bovine endothelial cells through up-regulating the expression of transforming growth factor-β receptors. J Cell Physiol. 1998;176:565–573. doi: 10.1002/(SICI)1097-4652(199809)176:3<565::AID-JCP13>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- Newman MJ. Inhibition of carcinoma and melanoma cell growth by type 1 transforming growth factor β is dependent on the presence of polyunsaturated fatty acids. Proc Natl Acad Sci USA. 1990;87:5543–5547. doi: 10.1073/pnas.87.14.5543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumont N, Arteaga CL. Transforming growth factor-β and breast cancer: tumor promoting effects of transforming growth factor-β. Breast Cancer Res. 2000;2:125–132. doi: 10.1186/bcr44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akhurst RJ, Balmain A. Genetic events and the role of TGF β in epithelial tumour progression. J Pathol. 1999;187:82–90. doi: 10.1002/(SICI)1096-9896(199901)187:1<82::AID-PATH248>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- Barrack ER. TGFβ in prostate cancer: a growth inhibitor that can enhance tumorigenicity. Prostate. 1997;31:61–70. doi: 10.1002/(sici)1097-0045(19970401)31:1<61::aid-pros10>3.0.co;2-m. [DOI] [PubMed] [Google Scholar]
- Gorsch SM, Memoli VA, Stukel TA, Gold LI, Arrick BA. Immunohistochemical staining for transforming growth factor β1 associates with disease progression in human breast cancer. Cancer Res. 1992;52:6949–6952. [PubMed] [Google Scholar]
- Walker RA, Dearing SJ. Transforming growth factor β1 in ductal carcinoma in situ and invasive carcinomas of the breast. Eur J Cancer. 1992;28:641–644. doi: 10.1016/s0959-8049(05)80116-9. [DOI] [PubMed] [Google Scholar]
- Dalal BI, Keown PA, Greenberg AH. Immunocytochemical localization of secreted transforming growth factor-β1 to the advancing edges of primary tumors and to lymph node metastases of human mammary carcinoma. Am J Pathol. 1993;143:381–389. [PMC free article] [PubMed] [Google Scholar]
- Arteaga CL, Hurd SD, Winnier AR, Johnson MD, Fendly BM, Forbes JT. Anti-transforming growth factor (TGF)-β antibodies inhibit breast cancer cell tumorigenicity and increase mouse spleen natural killer cell activity. Implications for a possible role of tumor cell/host TGF-β interactions in human breast cancer progression. J Clin Invest. 1993;92:2569–2576. doi: 10.1172/JCI116871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohmori T, Yang JL, Price JO, Arteaga CL. Blockade of tumor cell transforming growth factor-βs enhances cell cycle progression and sensitizes human breast carcinoma cells to cytotoxic chemotherapy. Exp Cell Res. 1998;245:350–359. doi: 10.1006/excr.1998.4261. [DOI] [PubMed] [Google Scholar]
- Fajardo LF, Prionas SD, Kwan HH, Kowalski J, Allison AC. Transforming growth factor β1 induces angiogenesis in vivo with a threshold pattern. Lab Invest. 1996;74:600–608. [PubMed] [Google Scholar]
- Oft M, Heider KH, Beug H. TGFβ signaling is necessary for carcinoma cell invasiveness and metastasis. Curr Biol. 1998;8:1243–1252. doi: 10.1016/s0960-9822(07)00533-7. [DOI] [PubMed] [Google Scholar]
- Grady WM, Rajput A, Myeroff L, Liu DF, Kwon K, Willis J, Markowitz S. Mutation of the type II transforming growth factor-β receptor is coincident with the transformation of human colon adenomas to malignant carcinomas. Cancer Res. 1998;58:3101–3104. [PubMed] [Google Scholar]
- Arteaga CL, Koli KM, Dugger TC, Clarke R. Reversal of tamoxifen resistance of human breast carcinomas in vivo by neutralizing antibodies to transforming growth factor-β. J Natl Cancer Inst. 1999;91:46–53. doi: 10.1093/jnci/91.1.46. [DOI] [PubMed] [Google Scholar]
- Korach KS, Couse JF, Curtis SW, Washburn TF, Lindzey J, Kimbro KS, Eddy EM, Migliaccio S, Snedeker SM, Lubahn DB, Schomberg DW, Smith EP. Estrogen receptor gene disruption: molecular characterization and experimental and clinical phenotypes. Recent Prog Horm Res. 1996;51:159–186. [PubMed] [Google Scholar]