

Short abstract
The fibroblast growth factor receptor (FGFR)2 gene has been shown to be amplified in 5-10% of breast cancer patients. A breast cancer cell line developed in our laboratory, SUM-52PE, was shown to have a 12-fold amplification of the FGFR2 gene, and FGFR2 message was found to be overexpressed 40-fold in SUM-52PE cells as compared with normal human mammary epithelial (HME) cells. Both human breast cancer (HBC) cell lines and HME cells expressed two FGFR2 isoforms, whereas SUM-52PE cells overexpressed those two isoforms, as well as several unique FGFR2 polypeptides. SUM-52PE cells expressed exclusively FGFR2-IIIb isoforms, which are high-affinity receptors for fibroblast growth factor (FGF)-1 and FGF-7. Differences were identified in the expression of the extracellular Ig-like domains, acid box and carboxyl termini, and several variants not previously reported were isolated from these cells.
Keywords: alternative splicing, breast cancer, fibroblast growth factor receptor, receptor tyrosine kinase, SUM-52PE
Abstract
Introduction:
The FGFR family of receptor tyrosine kinases includes four members, all of which are highly alternatively spliced and glycosylated. For FGFR2, alternative splicing of the second half of the third Ig-like domain, involving exons IIIb and IIIc, is a mutually exclusive choice that affects ligand binding specificity and affinity [1,2,3]. It appears that the second half of the third Ig-like domain can dictate high affinity for FGF-2 or keratinocyte growth factor (KGF), whereas affinity for FGF-1 appears to remain the same [3]. Alternative splicing of the carboxyl terminus has been shown to involve at least two different exons that can produce at least three different variants. The C1-type and C2-type carboxyl termini are encoded by the same exon, and have two different splice acceptor sites, whereas the C3-type carboxyl terminus is encoded by a separate exon [4]. The biologic significance of the C1 carboxyl terminus, as compared with the shorter C3 variant found primarily in tumorigenic samples, has been studied in NIH3T3 transfection assays, in which C3 variants were able to produce three times more transformed foci in soft agar than C1 variants (both IIIb), whereas full length FGFR2 and FGFR1 (both IIIc variants) showed no transforming activity [4].
Previous studies [5,6] have found amplification and overexpression of FGFR2 in 5-10% of primary breast cancer specimens. A recent study [7] done using a tissue array consisting of 372 primary breast cancer specimens found a 5% incidence of FGFR2 amplification. To our knowledge, none of the HBC cell lines studied thus far have an FGFR2 gene amplification, although overexpression of FGFR2 message and protein has been documented for some breast cancer cell lines [6,8,9].
SUM-52PE is a breast cancer cell line previously isolated in our laboratory that grows under serum-free and epidermal growth factor-free conditions, has high levels of tyrosine-phosphorylated membrane proteins, and has the capacity to invade and grow under anchorage-independent conditions [10,11,12]. This cell line exhibits all of the important hallmarks of transformed, highly malignant cells. Therefore, SUM-52PE was used as a model to study the diversity of FGFR2 expression in a breast cancer cell line that has true amplification and overexpression of FGFR2.
Objectives:
This study was conducted to examine the degree of FGFR2 amplification and overexpression in the breast cancer cell line SUM-52PE. Subsequent sequencing and characterization of individual FGFR2 variants cloned from the SUM-52PE cell line was completed to determine the complexity of FGFR2 alternative splicing in the context of a highly metastatic breast cancer cell line.
Methods:
Southern, Northern and Western blot analyses were done in order to determine the degree of FGFR2 amplification and overexpression in the breast cancer cell line SUM-52PE. Individual FGFR2 variants were cloned out of SUM-52PE using FGFR2-specific primers in a reverse transcription (RT) polymerase chain reaction (PCR). FGFR2 cDNAs were characterized by restriction fragment analysis, sequencing and transient transfection into 293 cells to examine the protein expression of each FGFR2 clone.
Results:
The results of the Southern blot showed that there was a 12-fold amplification of FGFR2 in the SUM-52PE cell line. Northern blot analysis of SUM-52PE showed FGFR2 transcripts to be highly overexpressed compared with other breast cancer cell lines and normal HME cells. Several overexpressed bands of approximately 6.3, 5.0, 4.0, and 2.8kb were observed in SUM-52PE cells. The most prominent band, at 2.8kb, was so abundant that it was difficult to discern other individual bands clearly. Western blot analysis showed that both normal HME and HBC cells expressed two FGFR2 variants of 95 and 135kDa. The SUM-52PE cell line greatly overexpressed not only these two polypeptides, as compared with HME and HBC cells, but also overexpressed two unique variants of FGFR2 - 85 and 109kDa polypeptides - as well as several smaller polypeptides in the 46-53kDa range. The antibody used in Western blot analysis only recognizes FGFR2 isoforms that express the C1 carboxyl termini, therefore greatly underestimating the actual number of different FGFR2 variants that are overexpressed in this cell line.
PCR was performed to determine the proportion of C1/C2 variants as compared with C3 variants in the SUM-52PE cell line. Results of this analysis indicated the presence of all three types of variants in this cell line, although the C1/C2 variants were predominant as compared with the C3 variants in SUM-52PE.
Four different FGFR2-C1 clones were isolated and sequenced from SUM-52PE cells, which differed in their signal sequence, first Ig-like loop, and acid box. Two FGFR2-C2 clones were isolated from the SUM-52PE cell line, which were identical to each other except for the variable expression of the number of Ig-like domains (two or three). Three C3 clones were isolated and sequenced, two of which have not previously been described in the literature. Clone C3-#3 contained two Ig-like domains, but no acid box. C3-#5 was missing the first two Ig-like domains and the acid box, but did contain the third Ig-like domain.
Discussion:
There is an extensive amount of evidence implicating erbB-2, a gene that is overexpressed in approximately 30% of breast cancer cases, as a breast cancer gene [13]. The identification of other breast oncogenes that function in the remaining 70% of cases is an ongoing challenge, as is establishing a causal role for such oncogenes in HME cell transformation.
FGFR1 and FGFR2, previously established oncogenes, were found to be amplified within large amplicons on 8p11 and 10q26, respectively, in the breast cancer cell line SUM-52PE [14]. Previous studies have shown that the FGFR2 gene is amplified in about 5-10% of breast cancer cases.
Our results showed that SUM-52PE cells overexpressed many alternatively spliced isoforms of FGFR2 at both the transcript and protein level as compared with normal HME cells. The variability in FGFR2 isoform expression is complex and involves exon IIIb/c, which encodes the second half of the third Ig-like loop; variations in the carboxyl terminal end of the receptor, involving the C1/C2 or C3 domains; and variable expression of the Ig-like loops and acid box in the extracellular portion of the receptor. The characterization of three unique FGFR2 isoforms that were cloned from SUM-52PE may build on the findings of others concerning the transforming potential of FGFR2 variants [4]. In particular, because it has been demonstrated that expression of C3-IIIb variants may have more transforming activity than C1-IIIb variants, differences between the three C3 clones we have isolated may provide information regarding the influence of particular structural domains on transforming potential.
Ongoing studies are aimed at characterizing the transforming ability of FGFR2 isoforms obtained from SUM-52PE cells by transducing these genes into normal HME cells. By overexpressing FGFR2 isoforms in a physiologically relevant system, we hope to determine the isoform(s) that acts in a dominant way in the process of cell transformation, and to determine whether different regions present in individual clones drive specific phenotypes associated with transformation.
Introduction
The FGFs are a family of polypeptides consisting of 18 different growth factors that bind with varying specificity and affinity to four different FGFRs. FGFs stimulate proliferation of a wide variety of cells of mesenchymal, neuronal, and epithelial origins [15,16], and have been found [17] to induce, inhibit, and maintain cell differentiation in different experimental systems. In addition, FGFs have been shown to be involved in neuronal survival [18], and stimulation of angiogenesis [19] and embryogenesis [20].
The FGFRs are a family of receptors that are characterized by the presence of two or three Ig-like domains and an acid box in the extracellular domain, a transmembrane region, and a split kinase domain in the cytoplasmic domain of the molecule [17]. Binding of FGF to heparin, or cell-surface heparin sulfate proteoglycans, results in high-affinity binding of this complex to FGFRs [219]. FGFRs subsequently undergo dimerization, followed by transphosphorylation on cytoplasmic tyrosine residues [22,23].
FGFRs can be alternatively spliced into a variety of isoforms that may have different functions. For example, at the amino terminus, which encodes the ectodomain of the receptor, alternative splicing of exon III results in the synthesis of either the IIIb or IIIc versions of FGFR2, which differ in their ligand specificities. FGFR2-IIIb, also referred to as KGF receptor, has been shown to be a high-affinity receptor for KGF, whereas FGFR2-IIIc receptors have a high affinity for FGF-2. At the carboxyl terminus, FGFR2 can be alternatively spliced to produce three different variants from two different exons [4]. C1/C2 variants are produced from the same exon with two different splice acceptor sites. C3 variants are produced from a separate exon with a different 3' - noncoding region from that of C1/C2 variants, resulting in a shorter form of FGFR2, which has been found [4] to be the predominant variant in stomach cancer cell lines as compared with normal stomach tissue.
Overexpression and amplification of growth factor receptors are common alterations observed in HBC cells. The FGFR1 and FGFR2 genes are expressed in both normal and breast cancer tissues [24], and alterations, including amplification and overexpression of FGFR1 and FGFR2, have previously been reported in 5-10% of primary breast cancer specimens [5,6]. A recent study [7] that was done using a tissue array consisting of 372 primary breast specimens found a 5% incidence of FGFR2 amplification.
There are at least four studies [6,8,9,24] that have analyzed the presence of FGFR2 message in breast cancer cell lines. These studies show great discrepancy as to the presence or absence of FGFR2 message and level of expression, possibly due to the method of detection (RT-PCR, Northern blot, or ribonuclease protection assay). To our knowledge, none of the HBC cell lines studied thus far have FGFR2 gene amplification, although overexpression of FGFR2 message and protein has been documented for some breast cancer cell lines [6,8,9].
SUM-52PE is a breast cancer cell line previously isolated in this laboratory that grows under serum-free and epidermal growth factor-free conditions, has high levels of tyrosine phosphorylated membrane proteins, and has the capacity to invade basement membranes and grow under anchorage-independent conditions [10,11,12]. Thus, SUM-52PE cells, which have amplifications of both FGFR1 and FGFR2 genes, exhibit all of the important hallmarks of highly malignant cells. In order to understand better the important molecular alterations that lead to this transformed phenotype, we examined the contribution of FGFR2 overexpression to these characteristics. Isolation and characterization of individual FGFR2 isoforms is an important first step in identifying the contribution of this receptor to cell transformation. Therefore, SUM-52PE was used as a model to study the diversity of FGFR2 expression in a breast cancer cell line that has true amplification and overexpression of the FGFR2 gene.
Materials and methods
Southern blot analysis
Genomic DNA (10 μ g) was digested with Hind III, and restriction fragments were separated on an agarose gel and transferred by standard methods to a nylon membrane. The Southern blot was probed with either FGFR1 or FGFR2, generated by RT-PCR of MCF-10A cell RNA. The specific primers used in the RT-PCR reaction were as follows: 5' -CAAACCCAACCGTGTGACCAAAGTG-3' and 5' -CGTGCGAGGCCAAAGTCTGCTATCT-3' for FGFR1; and 5' -GGTCGTTTCATCTGCCTGGT-3' and 5' -CCTTCCCGTTTTTCAGCCAC-3' for FGFR2. The RT-PCR reaction was carried out as described below.
Northern blot analysis
RNA (10 μ g) from a normal HME cell line (MCF-10A) and a panel of HBC cell lines (SUM-52, -149, -185 and -206) was run on a 1% agarose gel containing formaldehyde, and then transferred to a Nytran membrane (Schleicher & Schuell, Keene, NH, USA). The membrane was probed with full-length FGFR2 C1-#1 isolated from the SUM-52PE HBC cell line. Probe (50ng) was labeled using a random priming kit (Gibco BRL, Grand Island, NY, USA) with 5 μ Cu α -P32-dCTP at 25°C for 2h. Unlabeled α -P32-dCTP was removed by centrifugation of the probe through a Quik-Spin (Roche Molecular Biochemicals, Indianapolis, IN, USA) column at 1100 revolutions/min for 4min. Labeled probe (20×106 cpm) was boiled at 100°C for 5min and then used to probe the Northern blot overnight at 42°C. Prehybridization of the membrane occurred for 2.5h at 42°C.
Fibroblast growth factor receptor-2 Western blots
Cells were rinsed twice with ice-cold Hanks' balanced salt solution (Gibco BRL) and then lysed on ice with a buffer consisting of 50mmol/l Tris-HCl (pH8.5); 150mmol/l NaCl; 1% Nonidet P-40 (ICN Biomedical, Inc, Aurora, OH, USA); 5mmol/l ethylene diamine tetra-acetic acid supplemented with 5mmol/l sodium orthovanadate; 50 μ g/ml phenylmethysulfonyl fluoride; 20 μ g/ml aprotinin; and 10 μ g/ml leupeptin. Lysates were spun at 20 800 g at 4°C for 10min and then analyzed for protein using a modified Lowry's method. Whole-cell lysates were resolved on 7.5% polyacrylamide gels, transferred to PVDF membrane (Millipore Corporation, Bedford, MA, USA), and probed with an anti-FGFR2 antibody (Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA), an anti-FGFR2 antibody pre-conjugated with a competitive peptide (Santa Cruz Biotechnology, Inc) at room temperature for 2h, or an anti-Flag antibody (M2 antibody; Sigma, St Louis, MO, USA).
Reverse transcription polymerase chain reaction analysis of exon IIIb/c expression
FGFR2 variants were amplified using exon III specific primers: 5' -CCCGGGTCTAGATTTATAGTGATGCCCAGCCC-3' for FGF-FB; and 5' -CCCGGGGAATTCACCACCATGCAGGCGATTAA-3' for FGF-RB [25]. RT-PCR amplification was carried out by use of the SuperScript One Step RT-PCR system (Gibco BRL) with the addition of 1 μ l Expand High Fidelity enzyme (Roche Molecular Biochemicals). RT was carried out at 50°C for 30min, followed by 94°C for 3min. The PCR reaction was for 25 cycles of 94°C for 30s, 65°C for 30s, and 72°C for 1min, and final extension at 72°C for 7min. Singly, Ava I and Hinc II digestion of the RT-PCR product was performed overnight at 37°C, and then run on 3% NuSieve 3:1 agarose gel (FMC Bioproducts, Rockland, ME, USA).
Isolation of fibroblast growth factor receptor 2 cDNA clones
RNA was isolated from the SUM-52PE cell line using the Trizol isolation technique (Gibco BRL). RT-PCR amplification of FGFR2 was carried out by use of the SuperScript One Step RT-PCR system (Gibco BRL) with the addition of 1 μ l Expand High Fidelity enzyme (Roche Molecular Biochemicals). RT was carried out at 50°C for 30min, followed by 94°C for 3min. Amplification of FGFR2 was carried out using gene-specific primers: 5' -ATGCCCGTAGAGGAAGTGTG-3' for FGFR2 upstream; 5' -AACGCACGTCCACCTTGAGTCCT-3' for C1/C2-specific downstream; and 5' -CTATTACTTGTCATCGTCGTCCTT-GTAGTCGATCTCATTGGTTGTGAG-3' for C3-specific downstream. This was done for 40 cycles of denaturation at 94°C for 15s, annealing at 55°C for 30s, and elongation at 72°C for 3min, and final extension at 72°C for 7min. FGFR2 cDNA was then digested with Sal I/Xba I and ligated into the pZ vector at 14°C overnight using T4 DNA ligase. The ligation reaction was transformed into SURE Escherichia coli cells (Stratagene, La Jolla, CA, USA) as recommended.
Transfection of 293 cells
One-hundred-millimeter dishes of 293 cells at 50% sub-confluence were transfected with 20 μ g plasmid containing individual cDNA clones isolated from the SUM-52PE cell line using the calcium phosphate method. Briefly, 293 cells were incubated in 10% Dulbecco's modified eagle medium with 10mmol/l chloroquine for 5-10min. A reaction mix of 20 μ g cDNA, 1×Hepes-buffered saline, and CaCl2 (JT Baker, Phillipsburg, NJ, USA) was put into 293 dishes. Media was changed 6-8h after transfection and whole-cell lysates were prepared 48h after transfection, as described above. Whole-cell lysates (100 μ g) were resolved on 7.5% polyacrylamide gels, transferred to a polyvinylidene fluoride membrane, and probed with the anti-FGFR2 antibody (Santa Cruz Biotechnology, Inc) or an anti-Flag antibody (M2 antibody, Sigma).
Sequencing of fibroblast growth factor receptor 2 variants
Plasmid cDNA for individually isolated FGFR2 isoforms was prepared and submitted to the University of Michigan DNA Sequencing core with a series of FGFR2-specific primers, as well as vector-specific primers. The sequencing of all FGFR2 variants was performed in both a 5' and a 3' direction, with the exclusion of the transmembrane domain, which was shown to be highly conserved between isoforms.
Results
Amplification of fibroblast growth factor receptor 2 in SUM-52PE cells
In previous experiments, we found by comparative genome hybridization analysis that SUM-52PE cells have large amplifications in the genomic regions of chromosomes 8 and 10, containing the FGFR1 and FGFR2 genes, respectively [14]. In order to characterize the amplification of these candidate breast cancer oncogenes, Southern blot analysis was performed on the HBC cell line SUM-52PE and other breast cancer cell lines [14]. The results of the Southern blot showed fivefold amplification of the FGFR1 gene and a 12-fold amplification of the FGFR2 gene in SUM-52PE cells, as compared with other breast cancer cell lines that do not have genomic amplifications in these regions (Fig. 1).
Figure 1.
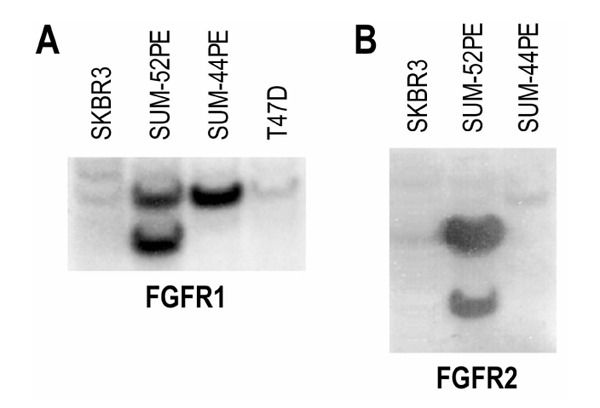
Amplification of FGFR1 and FGFR2 in SUM-52PE cells. (a) Genomic DNA from four breast cancer cells lines (SKBR3, SUM-52PE, SUM-44PE, and T47D) were compared by Southern blot for FGFR1 expression. (b) Genomic DNA from three breast cancer cell lines (SKBR3, SUM-52PE, and SUM-44PE) were compared by Southern blot for FGFR2 expression.
Overexpression of fibroblast growth factor receptor 2 transcript in SUM-52PE cells
Because gene amplification often involves large genomic regions that contain many genes, Northern blot analysis was performed to determine whether the observed amplifications of FGFR1 and FGFR2 correlated with transcript overexpression. Northern blot analysis of SUM-52PE showed FGFR2 transcripts to be highly overexpressed compared with other breast cancer cell lines (Fig. 2a, lanes 2-5) and normal HME cells (Fig. 2a, lane 1). Several overexpressed bands of approximately 6.3, 5.0, 4.0, and 2.8kb were observed in SUM-52PE cells. The most prominent band, at 2.8kb, was so abundant that it was difficult to discern other individual bands clearly. FGFR1 transcript overexpression, on the other hand, was not detected in SUM-52PE (data not shown).
Figure 2.
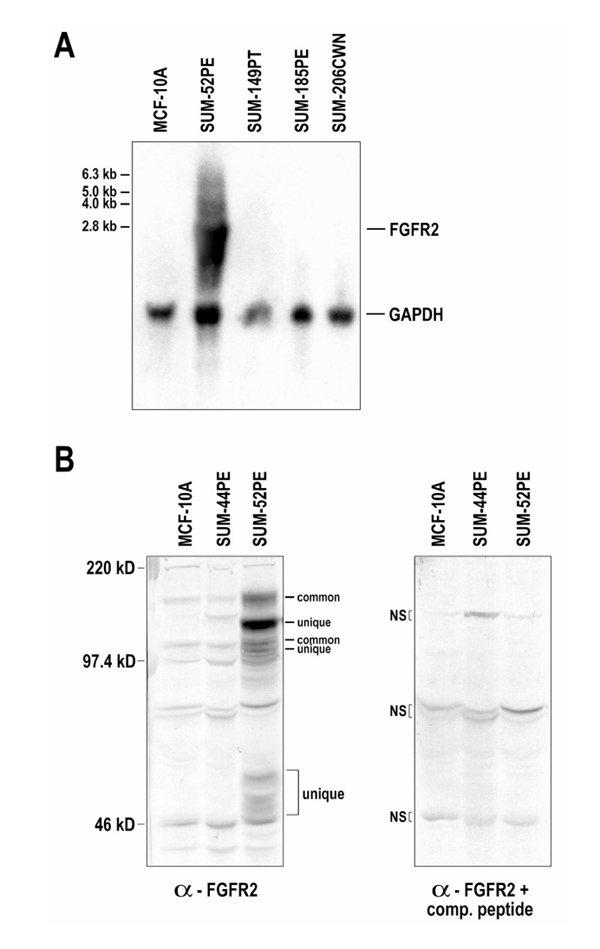
Analysis of FGFR2 expression in HBC and HME cells. (a) A Northern blot probed with full-length FGFR2 is shown. A glyceraldehyde 3' -phosphate dehydrogenase (GAPDH) probe was used to normalize RNA levels. Cell lines shown are as follows: normal HME cell line (MCF-10A, lane 1) and HBC cell lines (lanes 2-5). Size of visualized bands are marked according to kilobase. (b) Whole-cell lysates were immunoblotted with an anti-FGFR2 antibody in the absence (left panel) or presence (right panel) of a competitive peptide. Breast cancer cell lines are in lanes 2 and 3, and a normal HME sample (MCF-10A) is in lane 1. All cell lines expressed two 'common' FGFR2 variants (of 95 and 135 kDa), whereas SUM-52PE expressed at least three 'unique' variants (of 46-53, 85, and 109kDa), as determined by comparison with nonspecific bands (NS; right panel).
Overexpression of fibroblast growth factor receptor 2 variants in SUM-52PE cells
FGFR2 has been reported to be alternatively spliced, resulting in translation of multiple FGFR2 isoforms. Because Northern blot experiments demonstrated a number of different FGFR2 isoforms, Western blot analysis was performed to examine the number and level of expression of FGFR2 protein(s). The results showed that both normal HME and HBC cells (SUM-44PE, SUM-52PE, and others not shown) expressed two isoforms of FGFR2 of 135 and 95kDa (Fig. 2b, left panel, lanes 1-3, polypeptides labeled common). Interestingly, as compared with HME and other HBC cells, the SUM-52PE cell line greatly overexpressed not only these two polypeptides, but also overexpressed two unique isoforms of FGFR2 - 85 and 109kDa polypeptides - as well as several smaller polypeptides in the 46-53kDa range (Fig. 2b, left panel, lane 3, labeled unique). Use of a competitive peptide preconjugated with the anti-FGFR2 antibody confirmed that these bands were specific FGFR2 polypeptides (Fig. 2b, right panel). Use of an antibody isotype control also confirmed the same nonspecific bands as those that were identified by use of the competitive peptide (data not shown).
It is important to note that the polyclonal anti-FGFR2 antibody used in these experiments was created against a peptide derived from the C1 carboxyl terminus of FGFR2. Thus, this antibody only recognizes FGFR2 isoforms that express this carboxyl terminus. Because antibodies that recognize FGFR2 isoforms that express either the C2 or C3 carboxyl termini are not currently available, the results of the Western blot analysis of SUM-52PE cells under-represents the actual number of different FGFR2 proteins expressed in this cell line.
Preliminary characterization of fibroblast growth factor receptor 2 expression in SUM-52PE cells
To begin to characterize the FGFR2 transcripts that were present in SUM-52PE cells, PCR-based assays were performed to estimate the relative proportions of transcripts containing exon IIIb versus IIIc, and the proportion of C1/C2 variants as compared with C3 variants.
RT-PCR analysis using exon III-specific primers was performed on SUM-52PE mRNA, followed by restriction fragment analysis to determine the presence of exon IIIb/c-expressing variants [25]. Exon IIIb contains one unique Ava I site, whereas exon IIIc contains two Hinc III sites. Therefore, the proportion of Ava I digest fragments to Hinc III digest fragments allows for the determination of the proportion of IIIb to IIIc variants present. Using this method, SUM-52PE cells were found to express only IIIb isoforms, because the PCR product obtained was completely digested by Ava I, whereas Hinc III failed to cut any of the amplified product (Fig. 3a).
Figure 3.
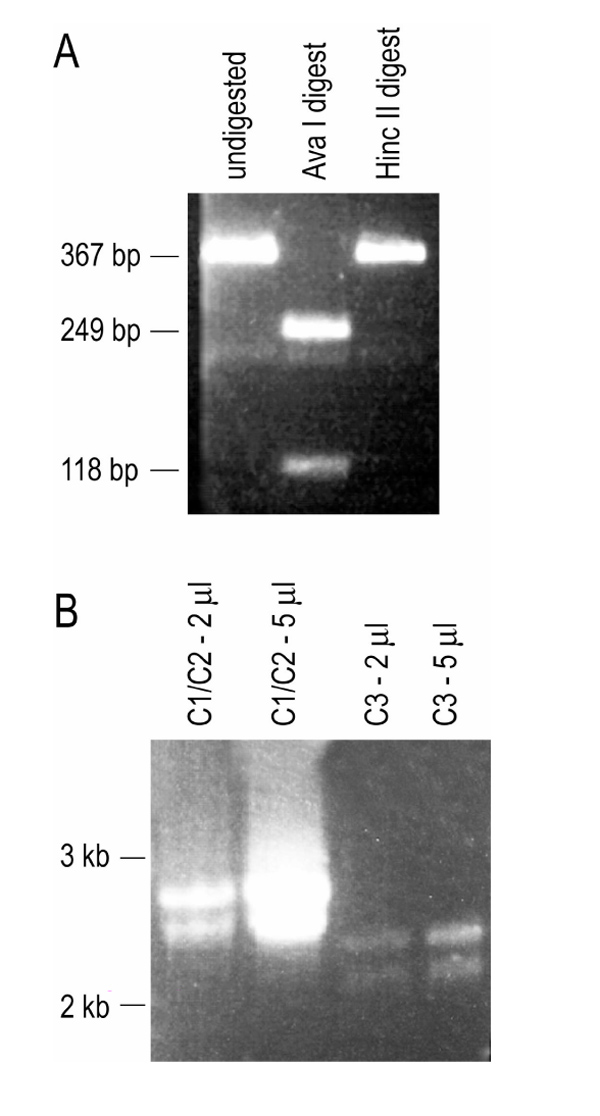
Preliminary characterization of FGFR2 expression in SUM-52PE cells. (a) Exon III-specific primers were used in RT-PCR of SUM-52PE RNA. RT-PCR product was then digested with Ava I or Hinc II at 37°C overnight and then resolved on a 3% NuSieve gel. Exon IIIb contains one unique Ava I site, whereas exon IIIc contains two Hinc II sites, and therefore the proportion of Ava I digest fragments to Hinc II digest fragments determines the proportion of IIIb to IIIc variants present. The presence of 269 and 188 bp fragments generated by Ava I digestion (lane 2) and lack of Hinc II digested products (lane 3) confirms the exclusive presence of exon IIIb in FGFR2 variants in the SUM-52PE cell line. (b) SUM-52PE mRNA was reverse transcribed using an oligo dT primer, and then amplified using a 5' -FGFR2-specific primer and a 3' -specific primer for C1/C2 or C3. Equimolar amounts of primer were used in the PCR reaction, and then 2 or 5 μ l of PCR product were compared on a 0.8% agarose gel. Lane 1, 2 μ l C1/C2 product; lane 2,5 μ l C1/C2 product; lane 3, 2 μ l C3 product; lane 4, 5 μ l C3 product.
C1/C2 variants were amplified using a primer specific to the 3' -noncoding region, whereas C3 variants were amplified using a primer specific for the C3 3' -noncoding region. Results of this analysis indicated the presence of all three types of variants in this cell line, although the C1/C2 variants (Fig. 3b; lanes 1 and 2) appeared to be more abundant than the C3 variants in SUM-52PE (Fig. 3b; lanes 3 and 4).
Isolation and analysis of alternatively spliced fibroblast growth factor receptor 2 variants
The PCR-based approach just described suggested that SUM-52PE cells express exclusively IIIb type receptors, which can contain any of the three carboxyl termini. The data also suggested that C1 and C2 variants are more common than receptors with the C3 terminus. In order to characterize definitively the range of FGFR2 variants expressed by SUM-52PE cells, individual isoforms were cloned and sequenced from SUM-52PE RNA. To isolate specific FGFR2 cDNAs, C1/C2- or C3-specific primers were used in combination with a primer for the 5' end of the gene. RT-PCR amplified FGFR2 products were purified and ligated into the bicistronic vector pZ. Individual clones containing insert were characterized by restriction digest analysis, sequencing, and transient transfection into 293 cells.
FGFR2 cDNA clones isolated from RNA derived from SUM-52PE cells were highly variable and differed with respect to number of Ig-like loops expressed, the presence or absence of the acid box, and the expression of C1, C2, or C3 carboxyl termini. As predicted from the PCR experiments, all FGFR2 variants isolated from SUM-52PE expressed the IIIb exon.
Four different FGFR2-C1 clones were isolated and sequenced (Fig. 4). The largest clone, C1-#38, was a full-length FGFR2-IIIb isoform, which was previously identified in the literature as KGF receptor [3,26]. C1-#38 contained three Ig-like domains, the characteristic acid box, and the C1 exon in the carboxyl terminus. The other three C1 isoforms contained two Ig-like domains, as well as exon IIIb, but differed with respect to the rest of their extracellular domains. C1-#8 contained two Ig-like domains and the acid box in the extracellular region. C1-#1 expressed two Ig-like domains, but was lacking part of the extracellular region corresponding to the acid box. This type of deletion has previously been identified as an FGFR2-IIIc variant, Bek103 [27]. C1-#10 was a unique variant that expressed two Ig-domains and the acid box, but contained a unique deletion of the 5' region of the receptor corresponding to the signal sequence. The functional significance of this type of deletion has yet to be determined.
Figure 4.
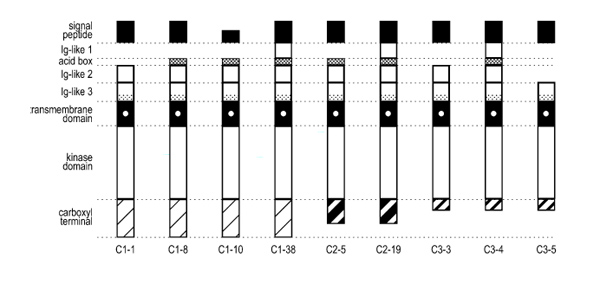
Isolated and sequenced FGFR2 isoforms from SUM-52PE. Isolated FGFR2 variants had variable expression of two to three Ig-like domains, as well as the characteristic acid box in the extracellular portion of the molecule. Variability between isolated clones also existed in the intracellular portion of the molecule, where alternative splicing of exons C1/C2 or C3 created either a full-length carboxyl termini (C1), or truncated versions (C2 or C3).
Two FGFR2-C2 clones were isolated from the SUM-52PE cell line (Fig. 4). Clone C2-#19 contained three Ig-like domains, the acid box, and the C2 exon in the carboxyl termini. Clone C2-#5 contained two Ig-like domains, but was missing part of the extracellular domain, which is similar to, but smaller than the deletion characterized for clone C1-#1. A clone identical to C2-#5 has previously been identified in the literature as human K-sam C2, a variant isolated from the KATO-III human stomach cancer-derived cell line [4]. Both K-sam and clone C2-#5 are missing the same 89 amino acid sequence in the 5' region of FGFR2 corresponding to the first Ig-like domain region.
Three C3 clones were isolated and sequenced, two of which have not previously been described in the literature (Fig. 4). C3-#4, a previously described FGFR2 variant, contained three Ig-like domains and the acid box, and was considered a full-length C3 clone [4]. Clone C3-#3 contained two Ig-like domains, but the lack of the acid box makes this a unique C3 variant. C3-#5 was missing the first two Ig-like domains and the acid box, but did contain the third Ig-like domain. This FGFR2 isoform also has not previously been reported.
Because FGFR2 transcripts from HME cells were not detectable by Northern blot, and because HME cells express FGFR2 protein that is detectable by Western blot, experiments were performed to isolate FGFR2 clones from RNA derived from normal HME cells. After two cycles of RT-PCR and subsequent cloning into pZ, three different FGFR2 isoforms were obtained. As with the SUM-52PE cells, all three isoforms cloned from HME cells contained exon IIIb. Two clones with C1 carboxyl termini were isolated, one of which was full length (the same as clone C1-#38 from SUM-52PE) and the second was missing the first Ig loop (the same as clone C1-#8). One clone containing the C2 terminus was isolated that was otherwise a full-length isoform (the same as clone C2-#19).
Fibroblast growth factor receptor 2 protein expression in transfected 293 whole cell lysates
To assess the ability of the various FGFR2 isoforms isolated from SUM-52PE cell RNA to synthesize protein, and to determine which FGFR2 clones direct the synthesis of protein isoforms detected in Western blots, 293 cells were transiently transfected with individual FGFR2 iso-forms using the bicistronic pZ expression vector and analyzed by Western blot. As can be seen in the left panel of Figure 5a, all four C1-containing isoforms expressed protein in 293 cells and each clone gave rise to an FGFR2 isoform with distinct molecular size. Similarly, all three C3-containing clones expressed protein in 293 cells (Fig. 5b). C3-containing isoforms were visualized using a flag antibody that detects an epitope tag incorporated into the design of the C3-specific primer. As can be seen from the right panel of Figure 5b, the molecular sizes of the C3 variants correspond to what would be predicted on the basis of the size of the individual clones. C3-#4 is full length at the amino terminus, C3-#3 is missing the first Ig loop and the acid box, and C3-#5 is missing the first Ig loop, the acid box and the second Ig loop.
Figure 5.
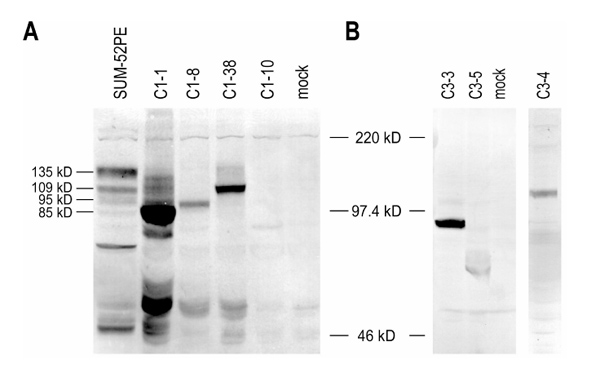
Transient expression of FGFR2 clones isolated from SUM-52PE. (a) Whole-cell lysates from 293 cells that had been transiently transfected with individual FGFR2 clones were immunoblotted with an anti-FGFR2 antibody (lanes 2-5). Whole-cell lysate from the SUM-52PE breast cancer cell line was loaded in lane 1 as a positive control, whereas whole-cell lysates from a mock transfection of 293 cells was loaded in lane six as a negative control. (b) Whole-cell lysates from 293 cells that were transiently transfected with individual FGFR2-C3 clones containing a Flag sequence were immunoblotted with an anti-Flag antibody (lanes 1, 2, and 4). Whole-cell lysates from a mock transfection of 293 cells were loaded in lane 3 as a negative control.
Unfortunately, we cannot assess protein synthesis from any of the C2-containing clones at this time. Because C1 and C2 are derived from the same exon, and because we used a single primer to clone both C1- and C2-containing variants, which did not contain an epitope tag, we are currently unable to detect FGFR2-C2 proteins.
As shown in Figure 2b, two FGFR2-C1 proteins (of 95 and 135kDa) are commonly detected in SUM-52PE cells, HME cells, and other breast cancer cell lines. This is in accord with the observation that both C1 variants cloned from HME cells were also cloned from SUM-52PE cells. Thus, the data suggest that the common band that migrates at 135kDa represents the glycosylated version of the full-length FGFR2-C1 (C1-#38: Fig. 5a; lanes 1 and 4). Similarly, the second common band that migrates at 95kDa is likely derived from clone C1-#8, which is missing the first Ig loop (Fig. 5a; lanes 1 and 3). Clone C1-#1 was highly expressed in 293 cells and resulted in the detection of multiple immunoreactive proteins with a range of sizes (Fig. 5a; lane 2). It is possible that this isoform, which is missing both the first Ig loop and the acid box, is responsible for some of the high- and low-molecular-weight bands detected in SUM-52PE Western blots, although more work will be required to definitively demonstrate this. The contribution of clone C1-#10, which was only weakly expressed in 293 cells, to FGFR2 protein expression in SUM-52PE cells cannot be determined at this time (Fig. 5a; lane 5).
As described above, clone C1-#38 encodes the full-length FGFR2-IIIb/C1 form of the receptor and is expressed at the message level in normal cells and all breast cancer cells examined, including SUM-52PE. However, SUM-52PE cells also have an overexpressed band at 109kDa, which was also detected in 293 cells transfected with clone C1-#38 (Fig. 5a; lane 4) and in HME cells transduced with this clone (not shown). This band was not detected in HME cells or any other breast cancer cells (Fig. 2b; left panel). The molecular size of this band corresponds to the size of the mature, non-processed, full-length protein, suggesting that the 135kDa band corresponds to the fully glycosylated form of the protein. The ability to detect this 109kDa non-processed form of full-length FGFR2-IIIb/C1 only in SUM-52PE cells and cells transduced with clone C1-#38 suggests that the protein is rapidly glycosylated in cells that express physiologic levels of FGFR2. By contrast, in cells that overexpress FGFR2, the 109-kDa nonglycosylated protein accumulates in the cell and is detectable by Western blot as one of the unique bands shown in Figure 2b. Thus, the detection of unique bands in Western blots derived from SUM-52PE cells reflects not only differences in isoform synthesis at the message level, but also differences in protein processing that occurs when the protein is highly overexpressed. At present, the functional consequences of the accumulation of nonglycosylated FGFR2 are not known.
Discussion
The progression of cells from the normal to neoplastic state is a multistep process that involves alterations in multiple signaling pathways. Both epidermal growth factor receptor and erbB-2 have been identified as signaling molecules that play a dominant role in breast cell transformation [28,29]. There is an extensive amount of evidence for erbB-2 as a breast cancer gene that is overexpressed in approximately 30% of breast cancer cases [13]. The identification of other breast oncogenes that function in the remaining 70% of cases is an ongoing challenge, as is establishing a causal role for such oncogenes in HME cell transformation.
Large regions of gene amplification in cancer cells can be detected by techniques such as comparative genomic hybridization and high-density arrays, which helps to localize areas that may contain functional oncogenes. FGFR1 and FGFR2, which were previously established as candidate breast cancer oncogenes, were found to be amplified within large amplicons on 8p11 and 10q26, respectively, in the breast cancer cell line SUM-52PE [14]. Previous studies [5,6,7] have shown that the FGFR2 gene is amplified in about 5-10% of cases. Because genes can be amplified without being overexpressed [14], we chose to examine whether FGFR2 may be an important oncogene in this breast cancer cell line by examining its expression at the mRNA and protein levels. Our results showed that SUM-52PE cells overexpressed many alternatively spliced forms of FGFR2 at both the transcript and protein level, as compared with normal mammary epithelial cells. By contrast, FGFR1 is not expressed in SUM-52 cells.
In contrast to SUM-52PE cells, FGFR2 expression at the message level is very low in HME cells. Indeed, even prolonged exposure of Northern blots to film did not allow the visualization of FGFR2 message in normal cells. However, Western blots did indicate the presence of FGFR2 protein in HME cells. To resolve this apparent paradox, two rounds of RT-PCR were performed using HME cell-derived RNA, which resulted in the isolation of three alternatively spliced forms of FGFR2 message, each of which expressed the IIIb exon. The predicted protein products of these clones correspond to that which was observed in Western blots.
The variability in FGFR2 isoform expression is complex and involves exon IIIb/c, which encodes the second half of the third Ig-like loop, variations in the carboxyl terminal end of the receptor involving the C1/C2 or C3 domains, and variable expression of the Ig-like loops and acid box in the extracellular portion of the receptor.
Alternative splicing of the FGFR2 mRNA that encodes the carboxyl terminus has been shown to involve at least two different exons, which can produce at least three different variants. The C1- and C2-type carboxyl termini are encoded by the same exon, having two different splice acceptor sites, whereas the C3-type carboxyl terminus is encoded by a separate exon [4]. The biologic significance of the full-length carboxyl terminus (C1), as compared with the truncated variant found primarily in tumorigenic samples (C3), has been studied in NIH3T3 transfection assays. The IIIb variants KGF receptor (C1) and K-sam C3 were both able to produce transformed foci, growth in soft agar and tumorigenicity in nude mice as compared with full-length IIIc variants of FGFR2 and FGFR1, which were not transforming [4]. The question of whether C3 variants are more transforming than C1 variants remains to be determined conclusively, because the number of transformed foci obtained using K-sam C3 was only threefold greater than that obtained using KGF receptor (C1) variants. A significant difference between the C3 and C1 termini is that the former does not contain the binding site for phospholipase Cγ. Thus, the ability of the variants of FGFR2 containing the C3 terminus to transform 3T3 cells suggests that signaling through this pathway is not necessary for FGFR2 IIIb to act as an oncogene.
The panel of FGFR2 isoforms isolated from SUM-52PE includes several unique and previously unreported isoforms. The first of these unique variants, C1-#10, contains a large deletion of the 5' region that includes the first Ig-like domain as well as the signal sequence, which could have interesting cellular localization and cell signaling properties due to the absence of part of the signal sequence. Clones C3-#3 and C3-#5 have not previously been reported and are missing the first Ig-like domain and acid box. C3-#5 is also missing the second Ig-like domain. The characterization of these three unique isoforms may build upon the findings of others concerning the transforming potential of FGFR2 variants [4]. In particular, because it has been demonstrated that C3-IIIb variants may have more transforming activity than C1-IIIb variants, differences between the three C3 clones we have isolated may provide information on the influence of particular structural domains on transforming potential.
Previous studies that examined FGFR2 expression in prostate cancer have suggested that a change in the expression from the exon IIIb to IIIc isoform correlates with a progression from an androgen-sensitive to an androgen-insensitive state. RT-PCR analysis on the SUM-52PE breast cancer cell line showed that this cell line exclusively expressed the IIIb FGFR2 isoform (Fig. 3). Exon IIIb expression was also exclusively found in normal luminal HME cells (data not shown). This suggests that exon IIIb to IIIc switching is not necessary for FGFR2 to act as an oncogene when the gene is amplified. Rather, overexpression of one of the common IIIb isoforms or one of the novel variants may be important in driving transformation of HME cells. Ongoing studies are aimed at characterizing the transforming ability of individual FGFR2 isoforms obtained from SUM-52PE cells. These studies will directly test the hypothesis that specific FGFR2 isoforms have transforming activity towards HME cells and will compare variants with the different carboxyl termini. Overexpression of the C1-#38 and C3-#5 FGFR2 clones has been successfully accomplished in both the MCF-10A and H16N2 HME cell lines, and these cells have acquired phenotypes that distinguish them from parental cells (to be described in detail in a separate paper that is in preparation). Thus, by overexpressing FGFR2 isoforms in a physiologically relevant system, we hope to determine the isoform(s) that acts in a dominant way in the process of cell transformation, as well as to determine whether different regions present in individual clones drive specific phenotypes associated with transformation.
Acknowledgments
Acknowledgement
The pZ vector was kindly provided by Genetics Institute, Boston, MA, USA.
References
- Johnson D, Lu J, Chen H, Werner S, Williams L. The human fibroblast growth factor receptor genes: a common structural arrangement underlies the mechanisms for generating receptor forms that differ in their third immunoglobulin domain. Mol Cell Biol. 1991;11:4627–4634. doi: 10.1128/mcb.11.9.4627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champion-Arnaud P, Ronsin C, Gilbert E, Gesnel MC, Houssaint E, Breathnach R. Multiple mRNAs code for proteins related to the BEK fibroblast growth factor receptor. Oncogene. 1991;6:979–987. [PubMed] [Google Scholar]
- Miki T, Bottaro D, Fleming T, et al. Determination of ligand-binding specificity by alternative splicing: two distinct growth factor receptors encoded by a single gene. Proc Natl Acad Sci USA . 1992;89:246–250. doi: 10.1073/pnas.89.1.246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Itoh K, Hattori Y, Sakamoto H. Preferential alternative splicing in cancer generates a K-sam messenger RNA with higher transforming activity. . Cancer Res. 1994;54:3237–3241. [PubMed] [Google Scholar]
- Adnane J, Gaudray P, Dionne CA. BEK and FLG, two receptors to members of the FGF family, are amplified in subsets of human breast cancers. . Oncogene. 1991;6:659–663. [PubMed] [Google Scholar]
- Penault-Llorca F, Bertucci F, Adelaide J. Expression of FGF and FGF receptor genes in human breast cancer. Int J Cancer. 1991;61:170–176. doi: 10.1002/ijc.2910610205. [DOI] [PubMed] [Google Scholar]
- Kononen J, Heisdanen M, Barlund M. Biochip technologies for rapid analysis of gene amplifications in cancer [abstract]. Proc Am Assoc Cancer Res. 1999;40:319. [Google Scholar]
- McLeskey SW, Ding I, Lippman ME, Kern FG. MDA-MB-134 breast carcinoma cells overexpress fibroblast growth factor receptors and are growth-inhibited by FGF ligands. Cancer Res. 1994;54:523–530. [PubMed] [Google Scholar]
- Lorenzi MV, Castagnino P, Chen Q, Chedid M, Miki T. Ligand-independent activation of fibroblast growth factor receptor-2 by carboxyl terminal alterations. Oncogene. 1997;15:817–826. doi: 10.1038/sj.onc.1201242. [DOI] [PubMed] [Google Scholar]
- Ethier SP, Kokeny K, Ridings J, Dilts C. erbB family receptor expression and growth regulation in a newly isolated human breast cancer cell line. Cancer Res. 1996;56:899–907. [PubMed] [Google Scholar]
- Ignatoski KMW, Ethier SP. Constitutive activation of pp125fak in newly isolated human breast cancer cell lines. Breast Cancer Res Treat. 1999;54:173–182. doi: 10.1023/a:1006135331912. [DOI] [PubMed] [Google Scholar]
- Ignatoski KMW, Maehama T, Markwart SM, Dixon JE, Livant DL, Ethier SP. ERBB-2 overexpression confers PI 3 ' -kinase-dependent invasion capacity on human mammary epithelial cells. Br J Cancer. 2000;82:666–674. doi: 10.1054/bjoc.1999.0979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maquire H, Greene M. The new (c-erbB-2) oncogene. Semin Oncol. 1989;16:148–155. [PubMed] [Google Scholar]
- Forozan F, Veldman R, Ammerman CA, et al. Molecular cytogenetic analysis of 11 new breast cancer cell lines. Br J Cancer. 1999;81:1328–1334. doi: 10.1038/sj.bjc.6695007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gospodarowicz D, Neufeld G, Schweiger L. Molecular and biological characterization of FGF, an angiogenic factor which also controls proliferation and differentiation of mesoderm and ectoderm derived cells. . Cell Differ. 1986;19:1–17. doi: 10.1016/0045-6039(86)90021-7. [DOI] [PubMed] [Google Scholar]
- Burgess W, Maciag T. The heparin-binding (fibroblast) growth factor family of proteins. Annu Rev Biochem. 1989;58:575–606. doi: 10.1146/annurev.bi.58.070189.003043. [DOI] [PubMed] [Google Scholar]
- Johnson DE, Williams LT. Structural and functional diversity in the FGF receptor multigene family. Adv Cancer Res. 1992;60:1–41. doi: 10.1016/s0065-230x(08)60821-0. [DOI] [PubMed] [Google Scholar]
- Morrison R, Sharma A, DeVellis J, Bradshaw R. Basic FGF supports the survival of cerebral cortical neurons in primary culture. Proc Natl Acad Sci USA. 1986;83:7537–7541. doi: 10.1073/pnas.83.19.7537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folkman J, Klagsburn M. Angiogenic factors. Science . 1987;235:442–447. doi: 10.1126/science.2432664. [DOI] [PubMed] [Google Scholar]
- Yamaguchi T, Rassant J. Fibroblast growth factors in mammalian development. Curr Opin Genet Dev. 1995;5:485–491. doi: 10.1016/0959-437x(95)90053-j. [DOI] [PubMed] [Google Scholar]
- Vlodavsky I, Miao H-Q, Medalion B, Danagher P, Ron D. Involvement of heparan sulfate and related molecules in sequestration and growth promoting activity of fibroblast growth factor. . Cancer Met Rev. 1996;15:177–186. doi: 10.1007/BF00437470. [DOI] [PubMed] [Google Scholar]
- Shi E, Kan M, Xu J, Wang F, Hou J, McKeehan W. Control of fibroblast growth factor receptor kinase signal transduction by heterodimerization of combinatorial splice variants. Mol Cell Biol. 1993;13:3907–3918. doi: 10.1128/mcb.13.7.3907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellot F, Crumley G, Kaplow JM, Schlessinger J, Jaye M, Dionne CA. Ligand-induced transphosphorylation between different FGF receptors. . EMBO J. 1991;10:2849–2854. doi: 10.1002/j.1460-2075.1991.tb07834.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luqmani YA, Graham M, Coombes RC. Expression of basic fibroblast growth factor, FGFR1 and FGFR2 in normal and malignant human breast and comparison with other normal tissues. Br J Cancer. 1992;66:273–280. doi: 10.1038/bjc.1992.256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carstens RP, Eaton JV, Krigman HR, Walther PJ, Garcia-Blanco MA. Alternative splicing of fibroblast growth factor receptor 2 (FGF-R2) in human prostate cancer. Oncogene. 1997;15:3059–3065. doi: 10.1038/sj.onc.1201498. [DOI] [PubMed] [Google Scholar]
- Dell KR, Williams LT. A novel form of fibroblast growth factor receptor 2. J Biol Chem. 1992;267:21225–21229. [PubMed] [Google Scholar]
- Crumley G, Bellot F, Kaplow JM, Schlessinger J, Jaye M, Dionne CA. High-affinity binding and activation of a truncated FGF receptor by both aFGF and bFGF. Oncogene. 1991;6:2255–2262. [PubMed] [Google Scholar]
- Pierce JH, Arnstein P, DiMarco E, et al. Oncogenic potential of erbB-2 in human mammary epithelial cells. Oncogene . 1991;6:1189–1194. [PubMed] [Google Scholar]
- Zeillinger R, Kury F, Czerwenka K. HER-2 amplification, steroid receptors and epidermal growth factor receptor in primary breast cancer. . Oncogene. 1989;4:109–114. [PubMed] [Google Scholar]