

Abstract
We tested the signal value of song overlapping in banded wrens (Thryothorus pleurostictus), using interactive playback to either overlap or alternate with their songs. Males shortened song duration and decreased variability in song length when their songs were overlapped by playback, suggesting that they were attempting to avoid being overlapped and perhaps being less aggressive. A novel finding was an effect of long-term prior experience: song lengths remained relatively short in alternating trials that followed two or more days after overlapping trials. Approach responses to the two treatments did not differ overall, but there was a parallel effect of prior experience: males tended to stay further from the speaker during alternating treatments if they had previously been overlapped by playback. Some females paired to the male subjects sang in response to playback and were also influenced by prior experience, singing more during alternating trials that had not been preceded by an overlapping trial. Male overlappers may signal dominance over a rival to other male or female receivers in a communication network, but it is currently unclear whether overlapping indicates motivation to escalate an aggressive interaction, or whether this singing strategy is related to male quality. Banded wrens are long-lived and maintain year-round territories, so modifying responses to rivals based on prior experience is likely to be important for success.
Keywords: song overlap, interactive signals, communication networks, male quality, female song
Introduction
Overlapping signals are a feature of many communication systems, and may mask or enhance one another, depending on factors like signal modality and the importance of individual distinctiveness. In many insects and anurans, signal synchrony or alternation is a consequence of males attempting to produce the leading signals preferred by females (Gerhardt and Huber, 2002; Greenfield et al., 1997). In some systems, the temporal overlap of signals may benefit interactants. For example, groups of lionesses (Panthera leo) increase the effectiveness of their aggressive display by roaring in chorus (McComb et al., 1994), and synchronous vibrational signaling by aggregations of treehopper (Umbonia crassicornis) nymphs elicits maternal antipredator behavior that individual vibrational signals do not (Cocroft, 1996; Cocroft, 1999). However, interactions involving signal overlap are often competitive. The spatial overlap of chemical signals that occurs when rodents overmark one another appears to be competitive (reviewed in Johnston, 2003). Likewise, the temporal overlap of auditory signals in interacting territorial songbirds is agonistic (Todt and Naguib, 2000).
Song overlapping is a relatively infrequently used strategy in many avian systems, but the contexts in which it occurs and the response it elicits from both receivers and eavesdroppers suggest that it is a directed aggressive signal (Tables 1-3). Birds generally avoid singing when conspecifics are singing (Gochfeld, 1978; Wasserman, 1977), but increased numbers of neighbors can increase the chance of overlapping, and some individuals may be more likely to overlap neighbor songs than others (Hultsch and Todt, 1982; Popp, 1989). Furthermore, song playback simulating intrusion can elicit overlapping (Balsby and Dabelsteen, 2001; Brindley, 1991; Langemann et al., 2000; Naguib, 1999). Responses of birds to being overlapped vary. Some respond aggressively with increased song rates when their songs are experimentally overlapped (Dabelsteen et al., 1997; Naguib, 1999). Many increase variability in the timing of their songs, and some reduce approach and song duration (Dabelsteen et al., 1996; Mennill and Ratcliffe, 2004b; Osiejuk et al., 2004). There is evidence that prior experience of being overlapped influences responses (Naguib, 1999).
Table 1.
Studies on song overlapping: sender perspective
| Species | Context | Source |
|---|---|---|
| white-throated sparrow | avoid overlapping conspecific song | (Wasserman, 1977) |
| Zonotrichia albicollis | ||
| lesser skylark | avoid overlapping conspecific song | (Gochfeld, 1978) |
| Alauda gulgula | ||
| ovenbird | > with increasing number of neighbors | (Popp, 1989) |
| Seiurus aurocapillus | ||
| nightingale | some males overlap, others follow | (Hultsch and Todt, 1982) |
| Luscinia megarhynchos | overlap songs of simulated intruders | (Naguib, 1999) |
| whitethroat | overlap songs of simulated intruders | (Balsby and Dabelsteen, 2001) |
| Sylvia communis | ||
| great tit | overlap intruder unless overlapped | (Langemann et al., 2000) |
| Parus major | ||
| European robin | strangers > neighbors; | (Brindley, 1991) |
| Erithacus rubecula | playback in territory > outside |
Table 3.
Studies on song overlapping: eavesdropper perspective (responses to overlapper vs. overlapped in simulated vocal interaction)
| Species | Approach | Amount of song | Source |
|---|---|---|---|
| nightingale great tit | > time near overlapper | >, > time singing > | (Naguib and Todt, 1997) (Peake et al., 2001) |
| black-capped chickadee | approach overlapper | (Mennill and Ratcliffe, 2004a) |
Individual males may differ in their preferred singing patterns, but few studies have investigated male characteristics associated with these differences. Males may vary consistently in how they respond to simulated intrusion (Nowicki et al., 2002), and a few studies have shown that such individual differences are related to male characteristics such as age or dominance rank (Hyman et al., 2004; Mennill and Ratcliffe, 2004a; Mennill and Ratcliffe, 2004b). Though studies focusing on song overlapping have found individual differences in the use of overlapping (Hultsch and Todt, 1982) and in responsiveness to being overlapped (Naguib, 1999), characteristics of these males were not compared and further work is needed to determine whether song overlapping is related to male quality.
We used an interactive playback experiment to test the communication function of overlapping songs in banded wrens. Socially monogamous pairs are territorial and do not migrate. Males are considerably more vocal than females, and have repertoires of around 20 song types, many of which are shared among neighbors (Molles and Vehrencamp, 1999). Matching song types of rivals in vocal interactions is an aggressive signal eliciting rapid approach (Molles and Vehrencamp, 2001). Males also sometimes overlap the songs of rivals during countersinging interactions but overlapping is correlated with song rate, suggesting that some overlapping may be accidental (Burt and Vehrencamp, 2005). To determine whether song overlapping has signal value in banded wrens, we tested whether males responded differently to a simulated intruder that responded to their songs with alternating or overlapping songs. We also monitored the response of females during playback. We replicated the paired experimental design of Naguib (1999), presenting both treatments to each subject in either ‘escalating’ or ‘de-escalating’ sequences and testing for effects of prior experience. However, we increased the time between presentation of alternating and overlapping playback from just a few minutes to several days to determine whether such effects might persist over the longer term.
Methods
We conducted playback experiments from 16 May to 26 June 2003 in Santa Rosa National Park in the Guanacaste Conservation Area, Costa Rica. For details on the habitat and study population see Molles and Vehrencamp (1999). Subjects were 20 individually color-banded males resident on territories in the study area. Breeding stage of subjects varied and was not always known, so the experiment was designed to minimize variance due to breeding stage by testing the treatment effect within subjects in trials separated by just a few days.
Experimental design
We used a paired design, exposing each subject to the same playback songs in two interactive trials, an overlapping treatment and an alternating treatment. Trials on the same male were conducted 2 to 5 days apart, and successive males received treatments in the opposite sequence so that half the subjects received alternating, and the other half overlapping, treatments first. We employed a two-speaker design to standardize the state and location of subjects at the start of the experiment. We used a lure speaker to attract the subject to the area and stimulate singing, and then played experimental stimuli from the second speaker. To minimize interference from neighbors, we placed the experimental speaker well within the territory boundaries of the subject, approximately 1.5 to 2.0 m high. We marked a radius of 15m around the speaker with flagging tape in four directions for quantifying approach. We placed the lure speaker approximately 20m from the experimental speaker, at a similar height, and also well within the subject's territory. The two speakers, RadioShack Amplified Speaker System AMX 18, were connected to a Compaq Evo N800c laptop computer with volumes adjusted to approximate natural wren singing levels (70 dB at 10m, measured with a RadioShack Realistic sound pressure level meter 33-2050). Song files were played from within the program Syrinx-PC (www.syrinxpc.com - developed by John Burt, University of Washington, Seattle) with a single keystroke.
We played a 45-second lure and waited for the subject to approach the lure speaker and sing a song. If the subject did not respond within 5 minutes, we played the lure a second time, and abandoned the trial for that day if the subject again failed to respond. Seven males that did not respond were tested again later, and all except one responded and were included in the final analysis. When the bird sang its second song after the end of the lure, we started the experiment by broadcasting a song from the experimental speaker, so that the song started immediately after the start of the subject's song (overlapping treatment), or after the end of the subject's song (alternating treatment). We played songs in response to each of the subject's songs, ending playback after 10 songs had been broadcast. We continued to monitor the subject for three minutes after the last playback song.
Playback stimuli
We selected high quality recordings of several different song types from focal recordings of dawn and early morning song bouts of 13 males to use as playback stimuli. Using songs from 13 males in playback to 20 subjects raises the issue of pseudoreplication. However, this low level of replication was used to reduce the influence of source males on treatment sequence: songs from the seven source males that were played to two subjects were balanced with respect to treatment sequence, so one subject received the alternating treatment first and the other received the overlapping treatment first. Each subject heard several songs from one non-neighboring source male (2 to 5 territories away). We used one song type to create the 45-second lure containing four repeats of that song, and two different song types for experimental playback, switching between types halfway through. Exactly the same lure and experimental playback songs were used for the two trials on each subject. We used the cursor-delimited filter in the program Syrinx-PC to filter out noise and amplified all recorded songs to the same peak amplitude.
The interactive playback method results in differences between paired trials caused by variation in bird and experimenter behavior that may include factors other than the one of interest. We chose to standardize the amount of playback between trials by playing the same song types the same number of times. Because playback songs were broadcast in response to subject songs, the duration of playback period depended on the rate at which the subject sang. Thus the experimental period comprised a playback period of variable duration (mean = 4.5 min, range 1 to 19.5 min) and a 3-minute post-playback period. There was no difference in the duration of the playback period in alternating and overlapping trials (Wilcoxon signed rank Z = −0.5, n = 20, p = 0.6). In two trials, males moved away from the speakers during the playback period and may have countersung with neighbors in the distance. Because this happened late in the playback period (after the eighth and ninth playback songs respectively), we included these experiments, but scored song rate and song type diversity as missing values. In one of these trials, one of the playback songs overlapped a female song by mistake. In six trials, males sang one or two songs during the playback period that were not responded to with playback songs. In one trial only 9 playback songs were broadcast, and in another 11 songs were broadcast.
We confirmed that the two treatments did in fact differ in the factor of interest by quantifying the extent of overlapping. The proportion of all subject songs during the playback period that were overlapped was significantly higher during overlapping (median = 0.9, range 0.6-1.0) than alternating treatments (median = 0, range 0-0.2; Wilcoxon signed rank Z = −3.8, p < 0.001, n = 18). For those subject songs responded to with playback, the time from the end of the subject song to the start of the playback song differed significantly between treatments (paired t = 15.6, p <0.001, df = 19). In alternating treatments, playback songs started about 1 second after the end of the subject songs (1.03 ± 0.07 sec), while in overlapping treatments, playback started about 1 second before the end of the subject songs (−1.27 ± 0.16 sec). This translated into a significant difference in the proportion of the length of subject songs that was overlapped during alternating (median = 0.0, range 0-0.12) and overlapping trials (median = 0.47, range 0.26-0.68; Wilcoxon signed rank Z = −3.9, p < 0.001, n = 20).
Response to playback
We used the playback trial logging feature in Syrinx-PC to record the times all sound files were played and to create an audio record of the entire trial, recording vocal responses of the subjects using a Sennheiser ME67 directional microphone connected to the laptop computer. We extracted the following measures of male vocal response from the audio record: 1) Song length - mean length of subject songs responded to with playback; 2) Song length CV - coefficient of variation of song lengths, 100*SD/M; 3) Inter-song interval CV - coefficient of variation of intervals between the starts of consecutive subject songs responded to with playback; 4) Song rate - songs/minute over the entire experimental period; and 5) Diversity - song types/song over the entire experimental period. In addition, we observed approach to the experimental speaker and extracted the following measures: 1) Closest approach and 2) Prop. time close – proportion of time spent within 15m of the experimental speaker over the entire experimental period, including the post-playback period. We also noted whether males overlapped playback songs. We recorded female songs and computed female song rate for the experimental period.
Analysis
Log transformation normalized vocal measures of response, and square root transformation normalized closest approach data. The proportion of time spent close to the speaker was bimodal and no transformation normalized it. Associations between different types of response were explored using Pearson correlations or Spearman correlations (Table 4). We used a Principal Components Analysis of the three correlated vocal responses to generate a combined score (PC1) for each male. This vocal response PC1 was normally distributed, had an eigenvalue of 1.99 and explained 66% of the variance. Component weightings were: Song rate = −0.89, Diversity = 0.75, Inter-song interval CV = 0.80. The two correlated approach variables were analyzed separately to take advantage of parametric methods for analyzing closest approach.
Table 4.
Relationships between measures of response (Pearson correlations or Spearman's rho).
| Length | Length CV | Closest approach | Prop. time close | Song rate | Diversity | I.S.I. CV | |
|---|---|---|---|---|---|---|---|
| Song length CV | 0.05 | ||||||
| Closest approach | −0.06 | 0.01 | |||||
| Prop. time close | −0.09 | 0.13 | −0.81*** | ||||
| Song rate | −0.01 | −0.02 | 0.07 | −0.03 | |||
| Diversity | 0.07 | 0.21 | −0.00 | 0.12 | −0.54*** | ||
| Inter-song interval CV | −0.07 | 0.22 | −0.26 | 0.18 | −0.63*** | 0.35* | |
| Female song rate | −0.09 | 0.47** | −0.25 | 0.16 | −0.09 | 0.05 | 0.11 |
p < 0.05
p < 0.01
p < 0.001
For normally distributed measures of response we used repeated measures general linear models to test treatment (alternate, overlap) as a within-subject effect, and sequence (alternate first, overlap first) as a between-subject effect, and we also tested the interaction between them to assess the effect of prior experience. For proportion of time spent close, we tested for a treatment effect using Wilcoxon's signed ranks tests to compare alternating and overlapping trials within males. To assess the effect of prior experience on proportion of time spent close, we analyzed overlapping and alternating trials separately, using Mann-Whitney tests to compare males that received the overlapping treatment first with those that received it second, and likewise for alternating treatments. Female song rate was also not normalized by transformation, and was analyzed with the same method.
We examined the structure of male songs sung during the playback period to determine the mechanism underlying observed variation in song length. Songs consist of a series of phrases with a terminal trill, and different song types vary in length depending on the number and type of phrases they contain, as well as the number of elements within phrases (Fig. 1). Song length might vary because males select song types from their repertoire that are short (Fig. 1a, b) or long (Fig. 1c, d), or because they interrupt a song (omit the terminal trill, Fig. 1e), or combine two songs to form a compound song (Fig. 1f). The most striking difference in length was between alternating and overlapping trials of males that received the alternating treatment first (see Results and Fig. 2a), so we compared songs of these ten subjects. We counted the number of songs responded to with playback that were interrupted, compound, and short in overlapping and alternating trials. To categorize song types as ‘short’, we measured the length of exemplars of each song type in the subject's repertoire using recordings of dawn and early morning song, calculated the mean song length for each song type, and then designated the shortest third of the repertoire as ‘short’ song types. We used a logistic regression to determine the contribution of short types, interrupted songs, and compound songs to the difference in song length between alternating and overlapping trials.
Figure 1.
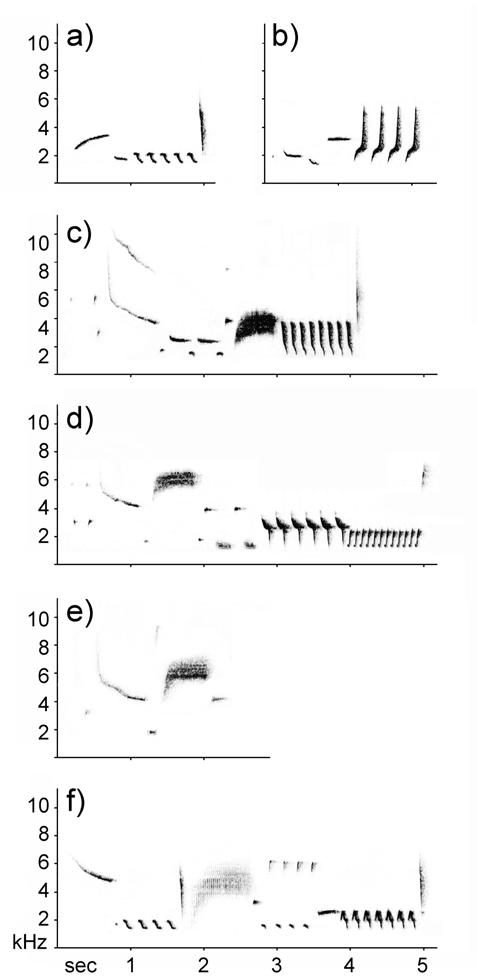
Sonagrams illustrating two of the song types categorized as ‘short’ (a, b), two of the song types categorized as ‘long’ (c, d), one interrupted song (e), and one compound song (f). The number and type of phrases constituting songs varies between song types, but songs tend to start with softer elements and end in a loud trill.
Figure 2.
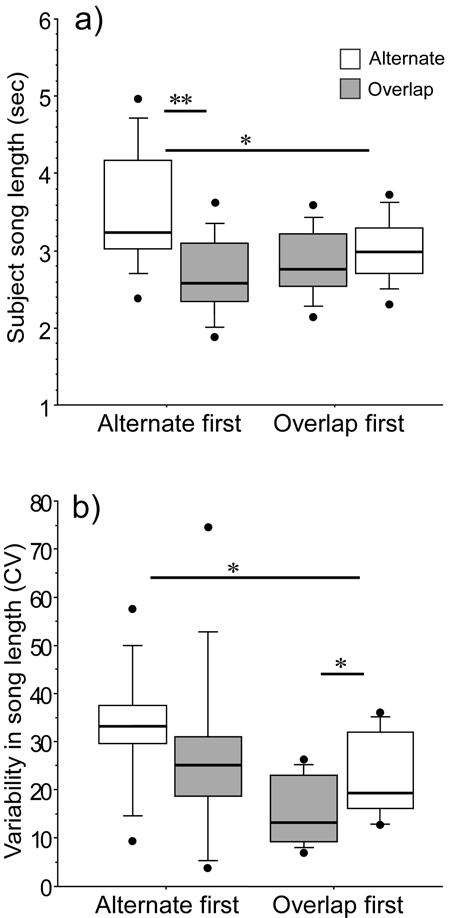
Length (a) and variability in the length (b) of male songs responded to with alternating or overlapping playback. Responses of males that received the alternating trial first are presented separately from those that received the overlapping trial first. Asterisks indicate significant post-hoc differences (** p < 0.01) and trends (* 0.05<p<0.1). Box plots enclose the interquartile range within the box – the central line shows the median, and cases in the 10th percentile at each extreme are shown as outlying points.
Following Naguib (1999), we tested for individual differences in male response by conducting separate one-way ANOVA's for each subject, with subject song length as the response variable and treatment as the explanatory variable. We tested for a relationship between male quality and their responsiveness to overlapping by comparing male weight, nesting success, their minimum age (number of years they were known to have been resident on the territory), and whether or not they survived until the following year.
We used SPSS 12.0 for Windows for computation. Means are presented with standard errors. Data presented graphically are shown untransformed for ease of interpretation.
Results
Overall response to playback
Banded wrens approached the speaker and sang in response to playback (Table 4). Males that approached the speaker closest also spent a greater proportion of time within 15 m of the speaker. Males that sang at high rates also sang with lower song type diversity and less variable song intervals. However, approach and vocal responses were uncorrelated. Males sang with more variable song lengths when their females sang at high rates, but all other measures of male response were not correlated with female song rate (Table 4). Individual males showed consistency between the two trials in their closest approach (Pearson correlation = 0.45, n = 20, p = 0.05), but not in vocal behavior (all p > 0.3).
Males sometimes responded to simulated intrusion by overlapping playback songs (15% of 40 trials). Though males only overlapped playback in 6 trials, there was a trend for less close approach in these trials than in those where males did not overlap playback (closest approach in trials with overlapping = 16.2 ± 3.1 m, with no overlapping = 9.9 ± 1.2 m; t = −1.95, df = 38, p = 0.06).
Female banded wrens seldom sing and can be cryptic at times (pers obs, Molles and Vehrencamp, 1999). However, females were clearly present in at least 24 trials, and sang during the experimental period in 21 trials (52.5%) with low song rates overall (median 0.30 songs/min, range 0 – 3.5).
Male vocal response to overlapping and alternating playback
Male songs were significantly shorter during overlapping than alternating treatments, and this effect was influenced by the sequence in which treatments were presented (Table 5). Post-hoc comparisons showed that songs were significantly shorter in overlapping than alternating trials for males that received the alternating treatment first, but not for those that received the overlapping treatment first (Fig. 2a). There was a trend for songs to be shorter in alternating trials given after overlapping trials than in alternating trials given first, but no difference in song length during overlapping trials associated with order of presentation (Fig. 2a). Males singing shorter songs had a smaller proportion of their song lengths overlapped by playback during overlapping trials (Pearson correlation between song length and % overlap = 0.71, p < 0.001, n = 20). Song length was less variable in overlapping than alternating trials, and it was also less variable among males that received the overlapping trial first than among those that received the alternating trial first (Table 5, Fig. 2b).
Table 5.
Results of repeated measures GLMs assessing effects on male responses.
| Treatment | Sequence | Treatment*Sequence | |
|---|---|---|---|
| (F1,18) | (F1,18) | (F1,18) | |
| Song length | 8.7*** | 0.2 | 5.6** |
| Song length CV | 5.2** | 4.4** | 0.04 |
| Closest approach | 0.4 | 2.9* | 1.6 |
| Vocal response PC1 | 0.3 | 1.5 | 0.2 |
0.1 =< p < 0.05
p =< 0.05
p =< 0.01
The reduction in song length in overlapping trials presented after alternating trials was attributable primarily to males singing more short song types and interrupting more songs during overlapping than alternating trials (short types: 4.8 ± 0.82 in overlap and 2.4 ± 0.87 in alternate, log likelihood χ21 = 5.9, p = 0.02; interrupted songs: 1.2 ± 0.44 in overlap and 0.4 ± 0.24 in alternate, log likelihood χ21 = 4.1, p = 0.04). The number of compound songs males sang did not contribute significantly to the difference between alternating and overlapping trials (1.3 ± 0.51 in overlap and 1.9 ± 0.70 in alternate, log likelihood ch χ21 = 0.3, p = 0.57).
Song rate, song type diversity, and variability of inter-song intervals did not differ significantly between alternating and overlapping treatments. Vocal response (PC1) did not vary with treatment, sequence, or their interaction (Table 5, Fig. 3).
Figure 3.
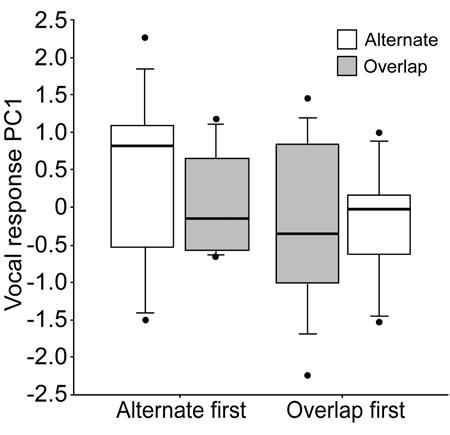
Vocal response to alternating and overlapping playback. Males with high PC1 scores sang with higher song type diversity, more variable inter-song intervals, and at a lower song rate. There were no post-hoc contrasts with p < 0.1.
Males overlapped playback in two overlapping trials and four alternating trials. Three of the four males that overlapped playback during alternating trials had previously been exposed to overlapping treatments, suggesting that prior experience with being overlapped by playback may have increased the likelihood that males would overlap playback songs, though the sample size was too small for inference.
Male approach to overlapping and alternating playback
Males exposed to alternating playback first showed a non-significant trend to approach the speaker more closely than males exposed to overlapping playback first (sequence effect), but there were no overall differences associated with treatment, or the interaction of treatment with sequence (Table 5). Examination of post-hoc contrasts showed that during alternating treatments, those males that had previously been overlapped tended not to come as close to the speaker as males that had not previously been exposed to playback (Fig. 4a). Closest approach during overlapping trials was unaffected by the order of presentation (Fig. 4a).
Figure 4.
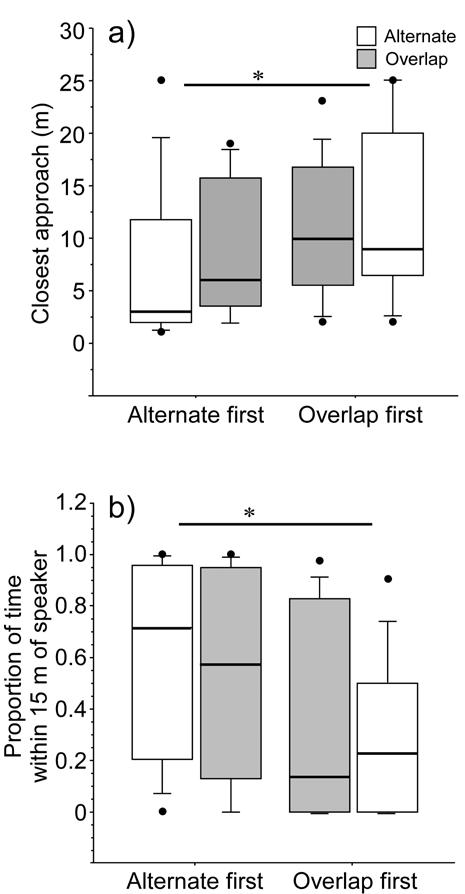
Distance of the closest approach to the experimental speaker (a) and the proportion of the experimental period that males spend within 15m of the experimental speaker (b) during alternating and overlapping trials. Responses of males that received the alternating trial first are presented separately from those that received the overlapping trial first. Asterisks indicate post-hoc trends (* 0.05<p<0.1).
The proportion of time males spent close to the speaker was similar in alternating and overlapping trials overall (Fig. 4b; Wilcoxon signed rank Z = −0.2, n = 19, p = 0.83). However, during alternating treatments, those males that had previously been overlapped tended to spend less time close to the speaker than males that had not previously been exposed to playback (Mann-Whitney U = 22.0, p = 0.06). This was not the case in overlapping trials: regardless of whether the overlapping trial was first or second, males spent similar amounts of time close to the speaker (Mann-Whitney U = 39.0, p = 0.40).
Male characteristics
Individual males varied in the extent to which they responded differently to the two treatments. Five males sang significantly shorter songs during overlapping than alternating trials (one-way ANOVAs F1,18 range = 9.0 to 27.2, p < 0.01; three of these males had received the alternating treatment first). Three males showed a trend in the same direction (F1,18 range = 3.0 to 3.4, 0.05 < p < 0.10; one of these males had received the alternating treatment first). The remaining 12 individuals did not vary song length in response to the two different treatments (F1,18 range = 0.08 to 2.7, 0.12 < p < 0.79). We compared several male quality characteristics of these ‘non-responders’ with males that showed some difference in song length between the two treatments. Males that responded by tending to sing shorter songs when their songs were overlapped did not differ from non-responding males in weight, their probability of fledging chicks, age, or how likely they were to survive until the following year (Table 6).
Table 6.
Male characteristics (Means ± SD or probability): males that responded to being overlapped (n = 8) versus non-responders (n = 12).
| Responder | Non-responder | Test | N(df) | P | |
|---|---|---|---|---|---|
| Weight (season adj g) | 20.5 ± 1.1 | 20.4 ± 0.9 | MWU = 38.0 | 19 (17) | 0.65 |
| Nest success (prob 1+fledgl) | 0.33 | 0.67 | Fisher exact | 20 (1) | 0.31 |
| Minimum age (yrs) | 1.9 ± 2.0 | 2.2 ± 1.6 | MWU = 41.0 | 20 (18) | 0.58 |
| Survival (prob survive) | 0.63 | 0.67 | Fisher exact | 20 (1) | 0.99 |
Few males overlapped playback, and they did not differ detectably from males that did not overlap playback in weight, age, or how likely they were to survive until the following year (Table 7; nesting success was unknown for 2 of 5 overlappers and therefore not presented).
Table 7.
Male characteristics (means ± SD or probability): males that overlapped playback songs (n = 5) versus males that did not overlap playback (n = 15).
| Overlapper | Non-overlapper | Test | N(df) | P | |
|---|---|---|---|---|---|
| Weight (season adj g) | 20.5 ± 1.4 | 20.3 ± 0.9 | MWU = 28.0 | 19(17) | 0.84 |
| Minimum age (yrs) | 2.0 ± 1.4 | 3.4 ± 3.1 | MWU = 20.0 | 20(18) | 0.12 |
| Survival (prob survive) | 0.6 | 0.67 | Fisher exact | 20(1) | 0.79 |
Males exposed to the alternating treatment first did not differ significantly from males that received the overlapping treatment first in any of the male quality characteristics assessed (Table 8).
Table 8.
Male characteristics (means ± SD or probability): males receiving overlapping treatment first versus alternating treatment first.
| Ovr first | Alt first | Test | N(df) | P | |
|---|---|---|---|---|---|
| Weight (season adj g) | 20.2 ± 0.9 | 20.4 ± 0.9 | MWU = 37.5 | 19 (17) | 0.57 |
| Nest success (prob 1+fledgl) | 0.67 | 0.33 | Fisher exact | 20 (1) | 0.61 |
| Minimum age (yrs) | 3.4 ± 3.8 | 2.7 ± 2.5 | MWU = 40.0 | 20 (18) | 0.44 |
| Survival (prob survive) | 0.5 | 0.8 | Fisher exact | 20 (1) | 0.35 |
Female response to overlapping and alternating playback
Females did not sing at different rates overall in trials where playback alternated with versus overlapped their partners (Wilcoxon signed rank Z = −1.6, n = 19, p = 0.12), but like males, their response was influenced by the sequence in which treatments were presented (Fig. 5). Females sang significantly more during alternating trials if their partner had not previously been overlapped (Mann-Whitney U = 15.5, p < 0.01). However, females sang at similar low rates during overlapping trials regardless of whether the overlapping trial was first or second (Mann-Whitney U = 42.0, p = 0.84).
Figure 5.
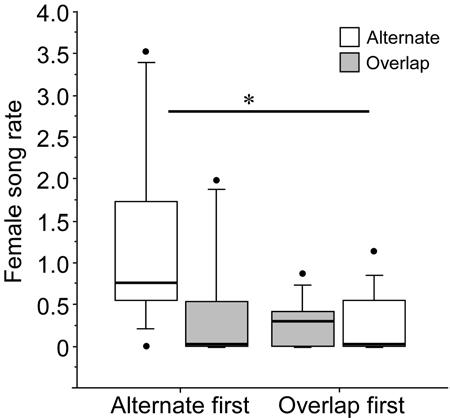
Female song rate (/min) during trials when playback alternated with or overlapped the songs of their partner. Responses of females paired with males that received the alternating trial first are presented separately from those that received the overlapping trial first. Asterisks indicate significant differences (** p < 0.01).
Discussion
Overlapping elicited an aversive response from banded wrens. Males shortened their songs, apparently to avoid being overlapped. This effect was maintained in alternating trials conducted several days after overlapping trials, indicating that males modify their behavior based on prior experience with a rival. Males also tended to stay further from the speaker if they had previously been overlapped. Shorter songs and reduced approach suggest a less aggressive response, indicating that overlapping may be so threatening that it inhibits aggression. Female response showed a similar effect of prior experience, with females singing at lower rates when playback overlapped the songs of their partners, and during alternating trials that had been preceded by an overlapping trial. There were individual differences among males in their responsiveness to the treatments, but we found no difference in the quality of ‘responding’ versus ‘non-responding’ males.
Male banded wrens sang songs that were shorter and had less variable lengths during overlapping trials than during alternating trials. Experimental studies on song overlapping have consistently shown that birds modify the timing of their songs to avoid being overlapped (Tables 1-3). Black-capped chickadees (Poecile atricapillus) also sang shorter songs when overlapped by playback, with a similar trend in corn buntings (Miliaria calandra). In contrast to banded wrens, variability in song length increased in response to overlapping in black-capped chickadees and great tits (Parus major; Tables 1-3). Nightingales (Luscinia megarhynchos) and black-capped chickadees increased the variability of intervals between songs when overlapped (Tables 1-3), but inter-song intervals were unaffected by overlapping in banded wrens.
Like nightingales and black-capped chickadees, male banded wrens reduced song length by interrupting more songs in overlapping than alternating trials (Mennill and Ratcliffe, 2004b; Naguib, 1999). In addition, banded wrens selectively used more short song types from their repertoire. Repertoires of song types increase signal diversity, but different song types are sometimes considered functionally equivalent (Krebs, 1976). However, there is increasing evidence that song types (Vehrencamp, 2001) and some structural features (Podos, 1997; Staicer, 1996) may differ functionally. Banded wrens selectively use song types of longer duration and broader bandwidth during intense dawn chorus singing versus later in the morning, and when counter-singing at the territory edge versus solo singing in the territory centre (Trillo and Vehrencamp, 2005). This use of longer songs during more aggressive contexts is consistent with our interpretation of reduced aggression during overlapping treatments in this study.
Despite paired trials being separated by two to five days, the responses of male banded wrens were influenced by prior experience with an overlapping ‘intruder’. The reduction in song length associated with being overlapped was significantly influenced by the sequence in which treatments were presented. The interaction between treatment and sequence was not significant for any other measure of response (Table 5), but the reversal in the direction of the difference in median responses to the two treatments associated with sequence of presentation evident for song length was also evident for all other measures of response (Fig. 2, 3, and 4). The trend for males that had previously been overlapped to sing shorter songs during alternating playback (Fig. 2a) was paralleled by trends for males previously overlapped by playback to sing songs with less variable lengths (Fig. 2b), approach the speaker less closely, and spend less time close to the speaker (Fig. 4). This was not merely a consequence of general previous experience with playback, as responses to overlapping trials did not differ depending on whether they followed an alternating trial or not.
Female banded wrens responded to playback of male song by singing and were also influenced by prior experience of a male ‘intruder’ overlapping the songs of their partner, reducing the intensity of their response during alternating trials that had been preceded by an overlapping trial several days before. This is the first study to document effects of song overlapping between males on the behavior of females during playback. Other studies on signal overlap have shown that female assessment of relative male quality and subsequent choices about pairing and mating are influenced by the relative timing of signals in vocal interactions between males, with females preferring males that do the overlapping to those that are overlapped (Mennill et al., 2002; Miyazaki and Waas, 2002; Otter et al., 1999). In banded wrens, females may have been inciting competition between males to facilitate assessment of male quality, or they may have been engaging in inter-sexual territorial defense. Banded wrens do not duet (Molles and Vehrencamp, 1999), but most of their congeners do (Brown and Lemon, 1979; Farabaugh, 1982) and partners defend territories together (Logue and Gammon, 2004). In banded wrens, male and female song rates were not correlated and their approaches did not appear to be closely coordinated (males also occasionally chased their partners during playback), but further work is needed to determine whether or not males and females coordinate their response to simulated intrusion.
This is the first field study to demonstrate an effect of being overlapped maintained over several days in both male and female responses to playback. Further testing is required to determine whether this was a generalized response to all intruders, or specific to the individual (or song types or location) associated with the prior interaction. In a study on two captive blackbirds that avoided singing from perches where their song was consistently overlapped, this conditioned aversion diminished only slightly over four days without playback (Todt, 1981). A field study on nocturnally singing nightingales where the two trials were presented a few minutes apart also showed an effect of prior experience being overlapped (Naguib, 1999). Male nightingales sang at higher rates during overlapping than alternating trials, and at higher rates in alternating trials that followed overlapping trials than in those presented first. The latter may have been a consequence of high arousal during overlapping causing high song rates in the alternating trial following soon after (Naguib, 1999). The reduced song length of banded wrens in alternating trials following overlapping trials was clearly not an immediate effect of being overlapped, and is consistent rather with a functional modification of response due to prior experience with a rival (Naguib, 1999).
Individual males showed some consistency between trials in the closeness of their approach to playback, and individuals also differed in their responsiveness to the treatments. However, we found no significant differences in the characteristics of males who responded to overlapping by reducing song length. We also found no differences in the characteristics of males that overlapped playback versus those that did not. However few males overlapped playback in this study, and a larger sample size is necessary to determine whether males that overlap differ in quality from males that do not overlap. A possible alternative explanation for response differences between males receiving different treatment sequences might be that there were some underlying differences in male ‘quality’ in the two groups of males. However, we found no differences in any of the measures tested.
Song overlapping is viewed as a directed agonistic signal (Todt and Naguib, 2000), yet overlapping playback seldom elicits aggressive responses from receivers. Though banded wrens use longer song types in more aggressive contexts (Trillo and Vehrencamp, 2005), they were less likely to use long song types or approach closely if they had previously been overlapped. This contrasts with the rapid approach elicited by song type matching, another singing strategy used in this species (Molles and Vehrencamp, 2001). Though European robins (Erithacus rubecula) and nightingales sang at higher song rates when their songs were overlapped by playback, and robins switched more quickly to an aggressive mode of singing, ‘twittering,’ no studies have found that overlapping playback elicits closer approaches or longer songs from receivers than alternating playback (Table 2). Rather, black-capped chickadees sang shorter songs when overlapped, with a similar trend in corn buntings. Corn buntings also spent less time close to the speaker during overlapping trials, while yellowhammers (Emberiza citrinella) were slower to approach the speaker (Table 2). The slower approach by yellowhammers may have been a proximate effect of overlapping playback limiting the ability of subjects to detect and locate their ‘rival’ (Osiejuk et al., 2004). However, this was clearly not the case in banded wrens, as the tendency to spend less time close to the speaker occurred when playback songs followed subject songs, in alternating trials presented after overlapping trials. The tendency for banded wrens to reduce approach and song length if they had previously been overlapped suggests rather that overlapping is so threatening that it is intimidating, and reduces the likelihood of aggressive close approaches and long song types. Intrusions into the territory center are uncommon in our population, and playback simulating an intruder singing well within territory boundaries and overlapping 10 songs in succession would have been extremely threatening.
Table 2.
Studies on song overlapping: receiver perspective (responses to overlapping vs. alternating playback)
| Species | Approach | Amount of song | Song timing | Source |
|---|---|---|---|---|
| European robin | = | >, twitter sooner | NA | (Dabelsteen et al., 1997) |
| nightingale | NA | > | > interrupted, > variable intervals | (Naguib, 1999) |
| great tit | = | = | > variable length | (Dabelsteen et al., 1996) |
| yellowhammer Emberiza citrinella | slower | = | NA | (Osiejuk et al., 2004) |
| corn bunting Miliaria calandra | < | = | < length (trend) | (Osiejuk, Ratynska, Cygan, in prep) |
| black-capped chickadee Poecile atricapillus | = | = | > interrupted, < length, > variable length, > variable intervals | (Mennill and Ratcliffe, 2004b) |
Examining overlapping from the perspective of the signaler, rather than the receiver, may shed light on why overlapping elicits an aversive response. Some signals are aversive because they are given at close range and associated with willingness to escalate an interaction or a high probability of attack. For example, screeching in red squirrels (Tamiasciurus hudsonicus) is closely associated with attack by the signaler and retreat by the receiver (Lair, 1990). Likewise, the A-songs of willow warblers (Phylloscopus trochilus) are associated with overt attack behavior and tend to cause receivers to withdraw (Jarvi et al., 1980). The honesty of such signals is likely to be maintained by vulnerability to retaliation by the receiver (Vehrencamp, 2000). Robins were more likely to overlap songs of strangers than those of neighbors, and approached songs of strangers more closely than those of neighbors (though the correlation between overlap and approach was not explicitly tested), and playback from just inside the territory boundary was more likely to be overlapped than playback from further away outside the territory (Brindley, 1991). However, great tits overlapped playback from a distance and, when playback escalated, they approached more closely but overlapped playback less (Langemann et al., 2000). Likewise, the closest approach of banded wrens that overlapped playback tended not to be as close as that of males that did not overlap playback. Further work from the signaler's perspective to identify behaviors associated with overlapping is necessary to determine whether song overlapping in birds indicates willingness to escalate an interaction.
Individuals also attend to the timing of vocal interactions between others, discriminating between overlapper and overlapped in these interactions. Studies on the response of eavesdroppers suggest that signal overlap is aggressive because it indicates dominance over a rival to other potential receivers by masking the signal of a rival, without necessarily indicating willingness to attack. For example, eavesdropping males respond more aggressively to leaders of dyadic interactions (Naguib et al., 1999), but if the second signal overlaps the first, then the overlapper elicits a more aggressive response than the overlapped (leading) male (Mennill and Ratcliffe, 2004a; Naguib and Todt, 1997). Females birds may prefer the male that does the overlapping over males who are overlapped (Mennill et al., 2002; Miyazaki and Waas, 2002; Otter et al., 1999). In rodents too, females may devalue males whose scent has been overmarked, or may show no preference for them (reviewed in Johnston, 2003). Females prefer leading males in many insects and anurans, and this tends to lead to call alternation among neighboring males when song rates are low, and call synchrony when song rates are high (Greenfield, 1994; Greenfield et al., 1997). A notable exception to synchrony as an epiphenomenon among anurans is the running frog (Kassina fusca), where females prefer leaders of overlapping interactions if the percentage overlap is high, and followers if it is lower (Grafe, 1999). Males in this species time their signals competitively, overlapping rival males at the low percentage overlap that results in a female preference for followers (Grafe, 1999).
In summary, receivers generally respond to overlappers by temporal modification of their own signals to avoid being overlapped. Being overlapped influences the behavior of male banded wrens and their partners for a number of days following the interaction. Being overlapped may be costly if overlapping indicates dominance to other same- or opposite-sex individuals in the communication network. Though song overlapping in birds is considered an aggressive signal, it rarely elicits overt aggression in response, and further work is needed to determine whether signalers accompany song overlapping with other aggressive behaviors to escalate an interaction. Likewise, further investigation of the individual characteristics of males who overlap other males, or who are responsive to being overlapped, is necessary to determine the causes and consequences of these individual differences.
Acknowledgements
We would like to thank Stephanie Lessard-Pilon for her assistance in the field with conducting the experiment. The Area Conservacion de Guanacaste gave permission to work in Santa Rosa National Park, and Roger Blanco and Maria Marta Chavarria provided support and facilitated our research there. Thorsten Balsby and Carlos Botero discussed ideas about vocal interactions, and Tomasz Osiejuk, Renata Sousa-Lima and three anonymous reviewers provided comments on the manuscript. This research was funded by the National Institute of Mental Health (grant R01-MH60461).
References
- Balsby TJS, Dabelsteen T. The meaning of song repertoire size and song length to male whitethroats Sylvia communis. Behav Process. 2001;56:75–84. doi: 10.1016/s0376-6357(01)00189-9. [DOI] [PubMed] [Google Scholar]
- Brindley EL. Response of European robins to playback of song: neighbour recognition and overlapping. Anim Behav. 1991;41:503–512. [Google Scholar]
- Brown RN, Lemon RE. Structure and evolution of song form in the wrens Thryothorus sinaloa and T. felix. Behav Ecol Sociobiol. 1979;5:111–131. [Google Scholar]
- Burt JM, Vehrencamp SL. Dawn chorus as an interactive communication network. In: McGregor PK, editor. Animal communication networks. Cambridge University Press; Cambridge: 2005. pp. 320–343. [Google Scholar]
- Cocroft RB. Insect vibrational defence signals. Nature. 1996;382:679–680. [Google Scholar]
- Cocroft RB. Offspring-parent communication in a subsocial treehopper (Hemiptera: Membracidae: Umbonia crassicornis) Behaviour. 1999;136:1–21. [Google Scholar]
- Dabelsteen T, McGregor PK, Holland J, Tobias JA, Pedersen SB. The signal function of overlapping singing in male robins. Anim Behav. 1997;53:249–256. [Google Scholar]
- Dabelsteen T, McGregor PK, Shepherd M, Whittaker X, Pedersen SB. Is the signal value of overlapping different from that of alternating during matched singing in great tits. J Avian Biol. 1996;27:189–194. [Google Scholar]
- Farabaugh SM. The ecological and social significance of duetting. In: Kroodsma DE, Miller EH, editors. Acoustic communication in birds. Academic Press; New York: 1982. pp. 85–124. [Google Scholar]
- Gerhardt HC, Huber F. Acoustic communication in insects and anurans. The University of Chicago Press; Chicago: 2002. [Google Scholar]
- Gochfeld M. Intraspecific social stimulation and temporal displacement of songs of the lesser skylark, Alauda gulgula. Z Tierpsychol. 1978;48:337–344. [Google Scholar]
- Grafe TU. A function of synchronous chorusing and a novel female preference shift in an anuran. Proc R Soc Lond B. 1999;266:2331–2336. [Google Scholar]
- Greenfield MD. Cooperation and conflict in the evolution of signal interactions. Ann Rev Ecol Syst. 1994;25:97–126. [Google Scholar]
- Greenfield MD, Tourtellot MK, Snedden WA. Precedence effects and the evolution of chorusing. Proc R Soc Lond B. 1997;264:1355–1361. [Google Scholar]
- Hultsch H, Todt D. Temporal performance roles during vocal interactions in nightingales (Luscinia megarhynchos B) Behav Ecol Sociobiol. 1982;11:253–260. [Google Scholar]
- Hyman J, Hughes M, Searcy WA, Nowicki S. Individual variation in the strength of territory defense in male song sparrows: Correlates of age, territory tenure, and neighbor aggressiveness. Behaviour. 2004;141:15–27. [Google Scholar]
- Jarvi T, Radesater T, Jakobsson S. The song of the willow warbler Phylloscopus trochilus with special reference to singing behavior in agonistic situations. Orn Scand. 1980;11:236–242. [Google Scholar]
- Johnston RE. Chemical communication in rodents: From pheromones to individual recognition. J Mammal. 2003;84:1141–1162. [Google Scholar]
- Krebs JR. Habituation and song repertoires in the great tit. Behav Ecol Sociobiol. 1976;1:215–227. [Google Scholar]
- Lair H. The calls of the red squirrel - a contextual analysis of function. Behaviour. 1990;115:254–282. [Google Scholar]
- Langemann U, Tavares JP, Peake TM, McGregor PK. Response of great tits to escalating patterns of playback. Behaviour. 2000;137:451–471. [Google Scholar]
- Logue DM, Gammon DE. Duet song and sex roles during territory defence in a tropical bird, the black-bellied wren, Thryothorus fasciatoventris. Anim Behav. 2004;68:721–731. [Google Scholar]
- McComb K, Packer C, Pusey A. Roaring and numerical assessment in contests between groups of female lions, Panthera leo. Anim Behav. 1994;47:379–387. [Google Scholar]
- Mennill DJ, Ratcliffe LM. Do male black-capped chickadees eavesdrop on song contests? A multi-speaker playback experiment. Behaviour. 2004a;141:125–139. [Google Scholar]
- Mennill DJ, Ratcliffe LM. Overlapping and matching in the song contests of black-capped chickadees. Anim Behav. 2004b;67:441–450. [Google Scholar]
- Mennill DJ, Ratcliffe LM, Boag PT. Female eavesdropping on male song contests in songbirds. Science. 2002;296:873–873. doi: 10.1126/science.296.5569.873. [DOI] [PubMed] [Google Scholar]
- Miyazaki M, Waas JR. ‘Last word’ effects of male advertising calls on female preference in little blue penguins. Behaviour. 2002;139:1413–1423. [Google Scholar]
- Molles LE, Vehrencamp SL. Repertoire size, repertoire overlap, and singing modes in the banded wren (Thryothorus pleurostictus) Auk. 1999;116:677–689. [Google Scholar]
- Molles LE, Vehrencamp SL. Songbird cheaters pay a retaliation cost: evidence for auditory conventional signals. Proc R Soc Lond B. 2001;268:2013–2019. doi: 10.1098/rspb.2001.1757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naguib M. Effects of song overlapping and alternating on nocturnally singing nightingales. Anim Behav. 1999;58:1061–1067. doi: 10.1006/anbe.1999.1223. [DOI] [PubMed] [Google Scholar]
- Naguib M, Fichtel C, Todt D. Nightingales respond more strongly to vocal leaders of simulated dyadic interactions. Proc R Soc Lond B. 1999;266:537–542. [Google Scholar]
- Naguib M, Todt D. Effects of dyadic vocal interactions on other conspecific receivers in nightingales. Anim Behav. 1997;54:1535–1543. doi: 10.1006/anbe.1997.9997. [DOI] [PubMed] [Google Scholar]
- Nowicki S, Searcy WA, Krueger T, Hughes M. Individual variation in response to simulated territorial challenge among territory-holding song sparrows. J Avian Biol. 2002;33:253–259. [Google Scholar]
- Osiejuk TS, Ratynska K, Cygan JP. Signal value of alternating and overlapping singing in the Yellowhammer Emberiza citrinella. J Ethol. 2004;22:55–61. [Google Scholar]
- Otter K, McGregor PK, Terry AMR, Burford FRL, Peake TM, Dabelsteen T. Do female great tits (Parus major) assess males by eavesdropping? A field study using interactive song playback. Proc R Soc Lond B. 1999;266:1305–1309. [Google Scholar]
- Peake TM, Terry AMR, McGregor PK, Dabelsteen T. Male great tits eavesdrop on simulated male-to-male vocal interactions. Proc R Soc Lond B. 2001;268:1183–1187. doi: 10.1098/rspb.2001.1648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podos J. A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes:Emberizidae) Evolution. 1997;51:537–551. doi: 10.1111/j.1558-5646.1997.tb02441.x. [DOI] [PubMed] [Google Scholar]
- Popp JW. Temporal aspects of singing interactions among territorial ovenbirds (Seiurus aurocapillus) Ethology. 1989;82:127–133. [Google Scholar]
- Staicer CA. Acoustical features of song categories of the Adelaide's warbler (Dendroica adelaidae) Auk. 1996;113:771–783. [Google Scholar]
- Todt D. On functions of vocal matching - effect of counter-replies on song post choice and singing. Z Tierpsychol. 1981;57:73–93. [Google Scholar]
- Todt D, Naguib M. Vocal interactions in birds: The use of song as a model in communication. Adv Study Behav. 2000;29:247–296. [Google Scholar]
- Trillo PA, Vehrencamp SL. Song types and their structural features are associated with specific contexts in the banded wren. Anim Behav. 2005;70:921–935. doi: 10.1016/j.anbehav.2005.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vehrencamp SL. Handicap, index, and conventional signal elements of bird song. In: Espmark YO, Amundsen T, Rosenqvist G, editors. Animal signals: signalling and signal design in animal communication. Tapir Academic Press; Trondheim: 2000. pp. 277–300. [Google Scholar]
- Vehrencamp SL. Is song-type matching a conventional signal of aggressive intentions? Proc R Soc Lond B. 2001;268:1637–1642. doi: 10.1098/rspb.2001.1714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wasserman FE. Intraspecific acoustical interference in the white-throated sparrow (Zonotrichia albicollis) Anim Behav. 1977;25:949–952. [Google Scholar]