

Short abstract
Iodinated oestradiol-labeled oestrogen receptor (ER) isoforms devoid of amino-terminal ABC domains represent about two-thirds of the whole receptor population detected in cytosol samples from human breast cancers. This high frequency could not be ascribed to the expression of truncated mRNAs, or to the proteolysis of the native ER peptide at the time of homogenization or assay, suggesting an intracellular proteolysis. Free amino-terminal and ligand-binding domains maintained together within oligomeric structure(s); increase of ionic strength separated them. The amino-terminal region was consistently detected in the cell nucleus by specific immunohistochemistry leading to the concept of a potential intranuclear association between ER cleavage products and/or other regulatory proteins.
Keywords: estrogen receptor domains, hydroxylapatite, immunohistochemistry, primary breast cancer
Abstract
Introduction:
We previously reported that about two-thirds of [125I]oestradiol-labelled cytosolic ERs from breast cancer samples eluted as low-molecular-weight isoforms (≤ 37 kDa, size-exclusion fast pressure liquid chromatography [FPLC]). These isoforms failed to adsorb strongly to hydroxylapatite at high ionic strength, a property that was ascribed to receptors devoid of amino-terminal ABC domains. In view of recent data concerning intracellular proteolysis of several transcriptional regulators, the possibility of such behaviour for ER was assessed.
The clinical significance of ER measurement in breast cancer cytosols is well established; approximately 50% of ER-positive cases respond to endocrine therapy. Whether such a poor correlation is related to a high proportion of cleaved ER is a question of prime importance. Failure of routine ER assays to discriminate between full-length and cleaved receptors led us to develop an oestradiol-binding assay based on hydroxylapatite adsorption.
The aims of the present study were to demonstrate that hydroxylapatite adsorption assay easily identifies cleaved cytosolic ER forms and to assess the origin of such ER forms.
Method:
Breast cancer cytosols classified as ER-positive according to [3H]oestradiol-binding assay (dextran-coated charcoal [DCC]) were subjected to hydroxylapatite adsorption. ER isoforms covalently labeled with [125I]tamoxifen aziridine (TAZ) released from this matrix with 0.5 mol/l KCl were subsequently immunoprecipitated with a panel of monoclonal antibodies raised against various domains of ER (H222 [E], H226 [C] or ER1D5 [AB]) before being subjected to SDS-gel electrophoresis.
Three approaches were used to identify the origins of the cleaved ER forms: potential truncated ER-α messenger RNAs that may encode ER isoforms of low molecular weights (Northern blot assay) were sought by using ER-α full-length probe; heat treatment of tumour cytosols in the absence or presence of a cocktail of protease inhibitors was performed; and the molecular weight of intracellular ER molecules was determined by in situ [125I]TAZ-labelling, which minimizes ER proteolysis.
Breast cancer samples classified as ER-positive according to both biochemical (cytosolic DCC assay) and histochemical (ER1D5 monoclonal antibody) criteria were labelled with [3H]oestradiol and were subsequently subjected to hydroxylapatite adsorption. Hydroxylapatite extraction index (EI) is defined as a ratio of the specifically bound [3H]oestradiol released from the hydroxylapatite matrix with KCl to the total amount of the specifically bound [3H]oestradiol extracted successively with KCl and ethanol: EI= ([3H]oestradiol) [KCl] × 100/([3H]oestradiol) [KCl] + ([3H]oestradiol) [EtOH]. The EI was calculated for each cytosol in order to evaluate the amount of cleaved ER forms present. Persistence of adsorption ER to hydroxylapatite in the presence of KCl (low EI) and ER1D5 positivity established by immunohistochemistry are two independent criteria for the presence of amino-terminal ABC domains. We therefore assessed whether hydroxylapatite determinations performed on cytosols are related to immuno-histochemistry data.
Results:
Cytosol pools labelled with [125I]TAZ gave different electrophoretic patterns depending on the nature of the anti-ER monoclonal antibody used in the immunoprecipitation step preceding electrophoresis. The carboxyl-terminal-specific antibody H222 precipitated all ER isoforms (full-length 67 kDa ER, and cleavage products of 50 and 37-28 kDa), whereas the amino-terminal-specific antibodies H226 and ER1D5 precipitated only the full-length and a partially truncated isoform. Adsorption of this labelled cytosol pool onto hydroxylapatite with subsequent KCl extraction yielded ER isoforms with molecular weights between 37 and 28 kDa when immunoprecipitation of the elutes was carried out using H222. The absence of these isoforms after exposure of the elutes to H226 or ER1D5 demonstrated truncation of these isoforms at a site(s) downstream of ABC domains.
Total RNA from 46 tumours was exposed to ER-α full-length probe (Northern blot). All tumours expressed a full-length 6.6-kb ER mRNA; small-sized isoforms were not recorded. A good correlation resulted when amounts of 6.6-kb ER mRNA estimated by densitometry were compared with corresponding [3H]oestradiol-binding capacities (DCC assay), thereby rejecting the concept that low-molecular-weight isoforms were encoded by truncated ER mRNA.
We next investigated whether such isoforms might be generated by proteolysis. Cytosol samples of a series of breast tumours were labelled with [125I]TAZ in the presence of a cocktail of protease inhibitors. These inhibitors failed to maintain the full-length 67 kDa ER by SDS-PAGE. In situ [125I]TAZ-labelling of receptors associated with a protein extraction procedure minimizing their proteolysis displayed multi-bands electrophoretic patterns, almost identical to those found under conventional methods. Hence, ER molecular heterogeneity appears to result from an intracellular proteolysis. ER1D5 immunostaining scores (ISs) of a series of 15 tumours were significantly correlated with ER levels, as measured by hydroxylapatite assay of corresponding cytosols (total number of binding sites). Sequential extraction of bound [3H]oestradiol from hydroxylapatite with KCl and ethanol revealed an EI of over 30% in the large majority of these cytosols, indicating a high frequency of cleaved ER isoforms. Of note, no significant correlation between IS and EI data was recorded, suggesting that ABC and E domains are separated at high ionic strength, but are apparently held together within the cell nucleus in oligomeric structures.
Discussion:
Endogenous proteolysis is a regulatory mechanism in many cellular processes, such as cell cycle progression and transcriptional regulation. The present data extend this concept to ER. Indeed, proteolysis-generated ER fragments appear to be held together within the cell in oligomeric structures. Because ER proteolysis is probably relevant to several oestrogen target tissues, we suggest that the protein environment, which differs among tissues, may be a factor of major importance in the formation of distinct oligomeric structures, which elicit specific biological responses. The possibility of heterogeneous association between cleaved ER and regulatory proteins might perhaps result in a spectrum of such transcriptional activities. In this context, we propose that a complementary hydroxylapatite extraction assay (EI assessment) should be added to the usual tests to identify ER-positive tumours. Such a complementary test would provide an estimate of the level of cleaved ER forms, which may have biological and/or clinical relevance.
Introduction
Assessment of the ER status in breast cancer samples is currently used to select patients for endocrine (tamoxifen) therapy [1,2,3]. Biochemical determinations of receptor concentration are based on the measurement of the tritiated oestradiol-binding capacity of cytosol samples; immunoenzymatic measurements (Abbott's ER enzyme immunoassay) usually give similar data because they use monoclonal antibodies against epitopes that are localized at both edges of the hormone-binding domain (E domain) [4]. Histochemical data established with various anti-ER monoclonal antibodies are in agreement with these biochemical assays, at least on a qualitative basis [5,6,7]. Of note, histochemical determinations identify ER essentially in the cell nucleus, leading to the concept of a possible dissociation of nuclear oligomers with concomitant release of the receptor into the cytosol at the time of homogenization.
Approximately 50% of ER-positive breast cancers respond to endocrine therapy [8]. A defect in the synthesis and/or turnover of ER that leads to the emergence of altered receptors has been suggested as an explanation for this relatively poor correlation between treatment response and ER status [9,10]. In agreement with this, we found that about two-thirds of [125I]oestradiol-labelled cytosolic ERs elute as low-molecular-weight isoforms (≤ 37 kDa, by size exclusion FPLC) that fail to adsorb strongly to hydroxylapatite at high ionic strength [11], a property ascribed to receptors that are devoid of amino-terminal ABC domains [12] required for transcription activity. ER proteolysis may lead to such a situation. Thus, a differential susceptibility of the hinge region (D domain) that separates such ABC domains from the hormone-binding domain to endoproteolytic cleavage has long been recognized [13]; whole hormone-binding domain itself also contains a few specific motifs that are accessible to proteolytic attacks [14,15]. Of note, an oestradiol-inductible protease activity, which is detected in mouse reproductive tissues, has been reported to produce a cleaved ER that is devoid of its amino-terminal region, but which has a nuclear localization capacity [16]. Hence, cleaved ER isoforms may maintain a regulatory activity at the genomic level.
The relevance of the molecular heterogeneity of ER in breast cancer is still a matter of controversy [17,18]. Our observation that a given ER cleavage pattern is relevant to the whole tumour mass (ie various samples taken from a given patient usually display identical radiolabelled SDS-PAGE profiles when their receptor isoforms were labelled with [3H]TAZ [19]) accredit the analysis of this question. Whether ER degradation occurs within the cell or is a product of homogenization or assay is a question of prime interest, in view of recent data regarding the biological significance of intracellular proteolysis of transcriptional regulators, such as the nuclear factor-κB, p53, c-JUN, sterol-regulated element-binding proteins and MATα2 [20,21]. Of note, activation of nuclear factor-κB requires proteasome-dependent proteolysis, leading to the processing of a precursor (nuclear factor-κB1) into the mature form [22]. Association of nuclear factor-κB with a specific inhibitory protein (IκB), which masks its nuclear localization signal, constitutes a functional system in which DNA binding/activation and regulatory/inhibitory functions are performed by two separate proteins that can interact [23]. Similar association of distinct functional domains may be applicable to cleaved ER isoforms, because the native form is also subject to proteasomal degradation [24,25,26].
In the present paper we demonstrate that ER isoforms of low molecular weight could not be ascribed to the expression of truncated mRNAs, or to the proteolysis of the native peptide at the time of assay, suggesting a degradative mechanism at the intracellular level. Moreover, we establish systematic immunohistochemical identification of ER amino-terminal domains within the cell nucleus with an anti-ER monoclonal antibody (ER1D5) that is raised against this part of the receptor molecule. All of these findings support the concept of a potential intranuclear association between cleaved ER and other regulatory proteins.
Material and methods
Reagents
[3H]oestradiol (approximately 100 Ci/mmol) was obtained from Amersham (Buckinghamshire, UK); [125I]TAZ (approximately 2000 Ci/mmol) was obtained from Trigone Ligands/Biocode (Liège, Belgium); and unlabelled oestradiol was obtained from Sigma (St Louis, MO, USA). Reagents for SDS-PAGE as well as Bio-Gel HTP hydroxylapatite were from Bio-Rad (Richmond, CA, USA); and H222 and H226 anti-ER monoclonal antibodies were a gift from Dr D Cotter (Abbott Laboratories, North Chicago, IL, USA). ER1D5 anti-ER monoclonal antibody was from Immunotech (Marseille-Luminy, France). Protease inhibitors (AEBSF, 4- [2-aminoethyl]-benzenesulphonyl fluoride; PMSF, phenylmethylsulphonylfluoride; antipain; chymostatin; leupeptin; and calpastatin] were from Euro Biochem (Bierges, Belgium).
Oestrogen receptor preparations
Primary breast cancer samples (for steroid hormone receptor measurements) were obtained from our surgery department. For ER measurement, they were homogenized in 10 mmol/l phosphate (pH 7.4) containing 1 mmol/l EDTA and 1 mmol/l monothioglycerol using a whole glass homogenizer. Homogenates were subsequently centrifuged for 1 h at 100 000 g. Supernatants were stored in liquid nitrogen until they were labelled, which was within the following week.
Human full-length ER produced in yeast [27] was provided by Dr P Sjöholm (Karo Bio, Huddinge, Sweden). This preparation was at a concentration of approximately 1 pmol/ml phosphate buffer (pH 7.5), and was stored at -70°C. For experiments, aliquots were diluted with 10 mmol/l Tris-HCl buffer (pH 8), containing bovine serum albumin (fraction V; Sigma) to reach a final protein concentration of approximately 1 mg/ml.
Cytosols from MCF-7 cells growing in monolayer culture were prepared as described previously [28].
Oestrogen receptor assays
Dextran-coated charcoal assay
ER preparations were labelled with increasing concentrations of [3H]oestradiol (range 0.25-5 nmol/l) in the absence and presence of a 200-fold excess of unlabelled oestradiol (overnight incubation at 0°C) and unbound ligands were removed using the DCC adsorption method (0.5% charcoal, 0.05% dextran) [29]. Specific binding was calculated from the difference between radioactivity levels measured in the absence and presence of unlabelled oestradiol. Binding capacities were expressed per milligram of protein, the latter being measured using the Bio-Rad assay. Parameters of the binding reaction (n, number of binding sites; Kd, dissociation constant) were estimated using the LIGAND v4.5 program (Dr PJ Munson, National Institutes of Health, Bethesda, MD, USA), and the data were plotted according to the method of Scatchard [30]. The same procedure was applied for [125I]TAZ, except that a 5 nmol/l saturating concentration was used.
Hydroxylapatite assay and extraction index
ER preparations adsorbed on hydroxylapatite pellets (prepared in 10 mmol/l Tris-HCl buffer, pH 8) were labelled with increasing concentrations of [3H]oestradiol (range 0.25-5 nmol/l) in the presence and absence of a 200-fold excess of unlabelled oestradiol (overnight incubation at 0°C) [12]. Bound [3H]oestradiol complexes were then successively extracted with 0.5 mol/l KCl or absolute ethanol. Specific binding was calculated from the difference between radioactivity levels measured in the absence and presence of unlabelled oestradiol and the data were analyzed according to the method of Scatchard [30]. EI was defined as a ratio of the specifically bound [3H]oestradiol released with KCl to the total amount of the specifically bound [3H]oestradiol extracted successively with KCl and ethanol: EI= ([3H]oestradiol) [KCl]×100/([3H]oestradiol) [KCl]+ ([3H]oestradiol) [EtOH]. High values of EI were considered to indicate the absence of ABC ER domains [11].
The same procedure was applied for [125I]TAZ, except that a 5 nmol/l saturating concentration was used. Because ethanol was unable to elute the covalently bound [125I]TAZ from the ER in the final step of the conventional hydroxylapatite assay, extraction of labelled receptor was carried out with 0.5 mol/l phosphate. Bound [3H]oestradiol was measured with Ecoscint H scintillation fluid (National Diagnostics, Atlanta, GA, USA) in a Wallac 1409 liquid scintillation counter (Wallac Oy, Turku, Finland) with 45-60% counting efficiency; bound [125I]TAZ was measured in a γ-detector (Crystal 5400 Series, United Technologies Packard, IL, USA).
Heat treatment of breast tumour cytosols
Tumour cytosols were heated at 37°C for 2 min in the presence or absence of a cocktail of protease inhibitors (final concentrations: 1 mmol/l for AEBSF, antipain and chymostatin; 5 mmol/l for PMSF; 0.1 mmol/l for leupeptin; and 0.1 mg/ml for calpastatin). After treatment, samples were labelled with 1 nmol/l [125I]TAZ (1 h at 0°C) in the presence or absence of a 200-fold excess of radioinert oestradiol, immunoprecipitated with H222 anti-ER monoclonal antibody, and then analyzed using SDS-PAGE.
SDS-PAGE analysis for cytosolic oestrogen receptors
ER preparations (cytosol/KCl extract from hydroxylapatite) were covalently labelled with 1 nmol/l [125I]TAZ (1 h incubation at 0°C) in the absence or presence of a 200-fold excess of unlabelled oestradiol; unbound ligand was then removed with DCC treatment. Labelled ERs were subsequently immunoprecipitated with H222, H226, or ER1D5 anti-ER monoclonal antibodies, and then antirat IgG agarose was added. Resulting suspensions were centrifuged and the pellets washed before being solubilized in lysis buffer (4% SDS, 20% glycerol, 10% mercaptoethanol, 0.05% bromophenol blue in 500 mmol/l Tris-HCl; pH 6.8). Lysates were subjected to electrophoresis on a 10% polyacrylamide gel in buffer (25 mmol/l Tris-HCl, 0.1% SDS, 192 mmol/l glycine; pH 8.2). Gels were stained, dried and finally exposed to Kodak X-OMAT films (Eastman Kodak Cy, Rochester, NY, USA) for 1 day for autoradiographical visualization of radioactive bands [19]. The molecular weights of the labelled peptides were estimated according to the migration of 14-94 kDa protein standards (Amersham Pharmacia Biotech, Uppsala, Sweden).
SDS-PAGE analysis for tissue oestrogen receptors
Sliced breast cancer tissues (thickness varied among samples, but was always less than 0.5 mm) were incubated for 1 h at 37°C with [125I]TAZ (final concentration 1 nmol/l) in Krebs-Ringer phosphate buffer (pH 7.4), with 2% bovine albumin and 0.25% glucose [31,32]. Then, all tissues were washed with Krebs-Ringer phosphate buffer and mixed with the same solution, but containing 1% SDS, 1.6 mmol/l EDTA and 2% β-mercaptoethanol.Tissue slices were briefly and delicately homogenized, and heated to 100°C for 2 min. Protein extraction was carried out by phenol and precipitation by acetone containing 0.1 mol/l acetic acid [32]. After 2 h at -20°C, the extracts were centrifuged. After washing, the pellets were solubilized in lysis buffer and subjected to electrophoresis as described above.
Oestrogen receptor mRNA measurement
Total RNA was extracted with TRIzol reagent (Boehringer-Mannheim, Mannheim, Germany), dissolved in RNase-free water and quantified by spectrophotometry at 260-280 nm. Aliquots of 30 μg RNA/15 μl were electrophoresed through a 1% agarose formaldehyde gel, capillary transferred to a Hybond-N membrane (Amersham), and treated according to the manufacturer's instructions. An EcoRI fragment (1300 pb) of pOR3 used as an ER-α mRNA probe was from the American Type Culture Collection (Rockville, MD, USA). Blots were hybridized sequentially with [32P]-labelled ER cDNA probe (109 cpm/mg cDNA, produced by random priming [Boehringer Mannheim]). Prehybridization (4 h) and hybridization (18 h) were performed as described previously [33]. The membranes were then washed with sodium citrate solutions of increasing stringency, the last wash being performed in 0.3 × sodium citrate solutions containing 0.1% SDS. Blots were visualized by exposure of the membranes for 1 day to Kodak XAR-5 film in an autoradiography cassette with an intensifying screen.
Immunohistochemistry
Breast cancer sections submitted to formalin fixation and paraffin embedding were immunostained with ER1D5 monoclonal antibody (Immunotech, Marseille-Luminy, France) using microwave pretreatment for antigen retrieval. Briefly, the sections were soaked in buffer (sodium citrate 10 mmol/l; pH 6) and heated for three cycles, each for 5 min at 600 W. After microwave treatment, sections were allowed to cool in room temperature and incubated with ER1D5 (used at 1:150 dilution) for 1 h. Sections were processed using a multilink supersensitive streptavidin-biotin detection system (Biogenex, San Ramon, CA, USA), employing diaminobenzidine as chromogen and haematoxylin counterstaining.
The nuclear staining was classified in intense, moderate, weak or nonstained categories. The immunostaining score (IS) was based on the staining intensity among these categories and on the percentage of stained cells. The IS was defined as a weighted sum of the percentages in the following manner: IS= 4a + 2b + c, where a is the percentage of intensely stained cells, b is the percentage of moderately stained cells and c is the percentage of weakly stained cells [34]. This weighted distribution of positive cells is based on the previously demonstrated linear relationship between the subjectively determined optical density and the concentration of histochemical concentration product measured by microspectrophotometry [35].
Results
Adsorption of oestrogen isoforms onto hydroxylapatite at low ionic strength
Previous studies from our laboratory [12,36] revealed that ER preparations (human ER expressed in yeast, cytosols from MCF-7 breast cancer cells and uterus) adsorbed onto hydroxylapaatite display a lower [3H]oestradiol-binding capacity than those provided by the conventional DCC assay; interference of the phosphocalcic matrix of the hydroxylapatite with ligand binding was advocated to explain this discrepancy. Assessment of cytosols from human primary breast tumours revealed that this property also holds for their receptors.
In 97 cases out of a series of 102 cytosols, assessment of the binding parameters of [3H]oestradiol (multipoint Scatchard plot analysis) gave Kd values of the same order of magnitude for both assays (median Kd values of 0.30 for DCC and 0.40 nmol/l for hydroxylapatite; Table 1). Binding capacities established by these two methods were also significantly correlated (r = 0.79; P < 0.001), with systematically higher values for the DCC assay (slope of the regression line= 1.47; Fig 1). The remaining five outlayers bound [3H]oestradiol with a relatively lower affinity (Kd about ten times higher; Table 1). These outliers were also characterized by a weak [3H]oestradiol-binding capacity by the DCC assay and a wide range of values by the hydroxylapatite procedure (Fig. 1; inset). They did not differ from the others with regard to their protein contents, thus refuting the hypothesis of false-negative or false-positive cases associated with a low or high protein level [37]. The nature of these low affinity-binding sites remains unknown.
Table 1.
Comparison of Kd values estimated by DCC and hydroxylapatite (HAP) assays in the whole series of human breast cancer cytosols(102 cases) as well as in the five cases not included in the correlation
| 102 Cases | 97 Cases | 5 Cases | ||||
| Range | Median | Range | Median | Range | Median | |
| Dissociation constants (nmol/l) | ||||||
| DCC | 0.001-02.74 | 0.31 | 0.001-2.74 | 0.30 | 0.010-02.70 | 0.45 |
| HAP | 0.001-17.90 | 0.42 | 0.001-2.92 | 0.40 | 0.080-17.90 | 4.26 |
| Protein (mg/ml) | ||||||
| 0.80-11.00 | 4.52 | 0.80-11.00 | 4.41 | 0.80-10.11 | 4.01 | |
Figure 1.
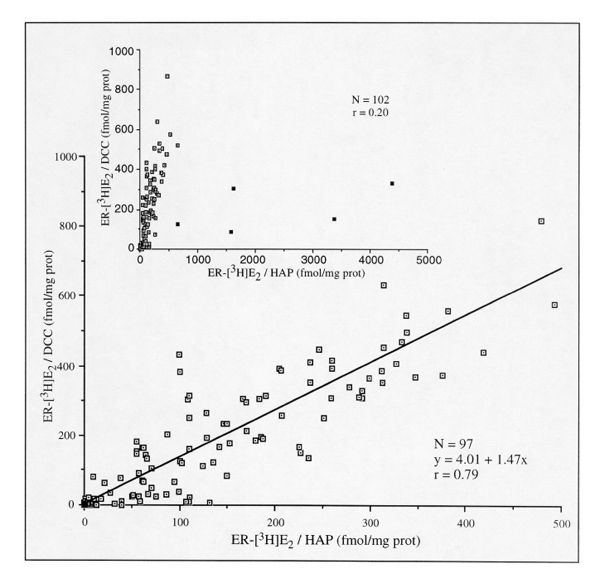
Comparison of [3H]oestradiol (E2)-binding capacities of a series of human breast cancer cytosols simultaneously measured by DCC and hydroxylapatite (HAP) assays (ethanolic extraction). The ordinate corresponds to values established by DCC, and the abscissa to those by hydroxylapatite (data established by Scatchard plot analysis). A significant correlation was obtained between the two assays in 97 out of 102 (95%) samples; five outlayers are represented by closed symbols in the insert.
Using [125I]TAZ instead of [3H]oestradiol as the labelling agent in another series of 36 cytosols, we established a similar correlation (r = 0.83; P < 0.001; Fig. 2). The slope of the regression line was similar to that established with [3H]oestradiol (1.40), indicating a similar interference of the phosphocalcic matrix of the hydroxylapatite in the binding of [125I]TAZ. Hence, at low ionic strength, ligand-binding capacities of cytosolic ER from breast tumours are significantly correlated when assessed by DCC and hydroxylapatite assays.
Figure 2.
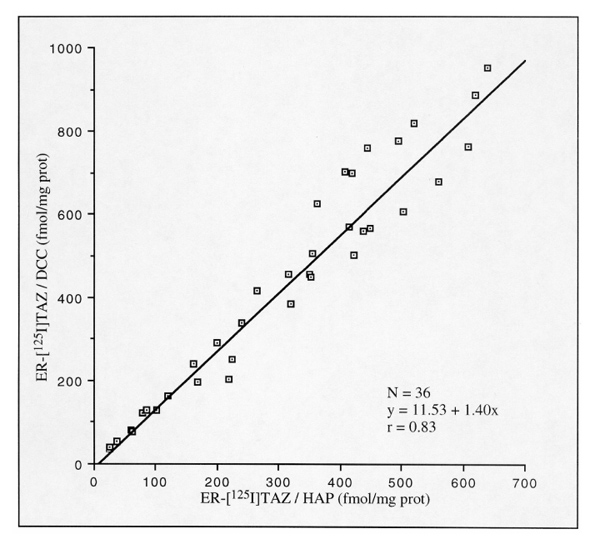
Comparison of [125I]TAZ-binding capacities (5 nmol/l) of a series of human breast cancer cytosols simultaneously measured by DCC and hydroxylapatite (HAP) assays (phosphate extraction).
Release of cleaved oestrogen receptor isoforms from hydroxylapatite with KCl
Data reported above suggest that the ER domains that are required for hydroxylapatite adsorption (ABC domains) and ligand-binding (E domain) are present in most tumour cytosols. In view of the high frequency of cleaved ER isoforms in breast cancer [11], such domains should be dissociated at high ionic strength, as demonstrated here using [125I]TAZ as a labelling agent.
ER isoforms labelled with radiolabelled TAZ were easily detected by successive immunoprecipitation and SDS-PAGE (full-length 67 kDa and cleavage products of 50 and 37-28 kDa) [19,38,39]. Using this approach we observed that a pool of breast cancer cytosols labelled with [125I]TAZ gave different electrophoretic patterns, depending on the nature of the anti-ER monoclonal antibody used in the immunoprecipitation step preceding electrophoresis (H222, H226, ER1D5; Fig. 3a, left). The carboxyl-terminal-specific antibody H222 precipitated all ER isoforms, whereas the amino-terminal-specific antibodies H226 and ER1D5 precipitated only the full-length and a partially truncated 50 kDa isoform, indicating a lack of corresponding antigenic sites in the 37-28 kDa isoforms. Of note, the 50 kDa band was more intensely labelled with H226. The [125I]TAZ-labelling intensity of all of these bands was suppressed with a 200-fold excess of unlabelled oestradiol, establishing their specificity.
Figure 3.
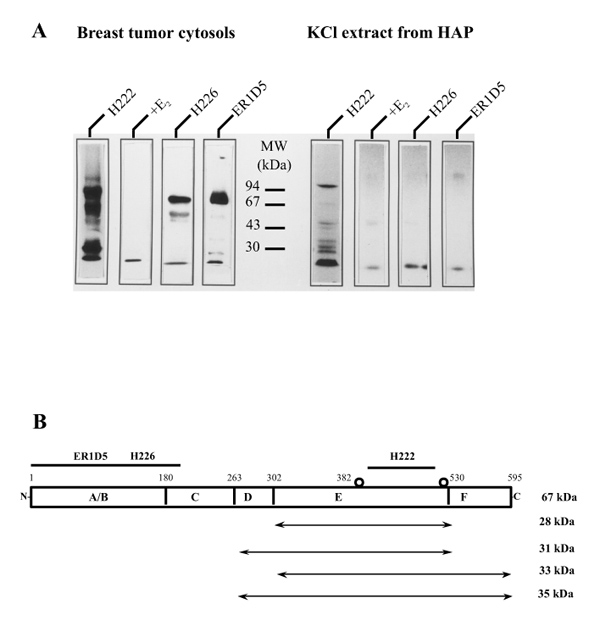
(A) Molecular weight and monoclonals anti-ER recognition of [125I]TAZ-labelled ER isoforms extracted from hydroxylapatite (HAP) with KCl. (left) Part of human breast cancer cytosol pool, after labelling with 1 nmol/l [125I]TAZ for 1 h at 0°C in the presence or absence of a 200-fold excess of radioinert oestradiol, was immunoprecipitated with H222, H226 or ER1D5 anti-ER monoclonal antibodies, and then analyzed by SDS-PAGE and autoradiography. (right) Another part of this cytosol pool was adsorbed onto HAP, labelled with 1 nmol/l [125I]TAZ, extracted with 0.5 mol/l KCl, and immunoprecipitated before being subjected to electrophoresis. (B) Presumed structure of ER isoforms extracted from HAP with KCl. Potential sites of covalent attachment of TAZ [40,41] are indicated by open circles; antigenic sites for anti-ER monoclonal antibodies are shown above ER structure. The predicted ER isoforms extracted from HAP as well as their sizes determined by SDS-PAGE are shown below.
Part of this [125I]TAZ-labelled cytosol pool was adsorbed onto hydroxylapatite and subsequently subjected to KCl extraction (Fig. 3a; right). Elutes were then immunoprecipitated before being subjected to SDS-PAGE. Absence of 67 and 50 kDa isoforms under all immunoprecipitation conditions confirmed their adherence onto hydroxylapatite due to their strong interaction (ABC domains) with the matrix. On the contrary, and as expected, ER isoforms with a molecular weight of between 37 and 28 kDa were detected in the elutes when the immunoprecipitation was carried out by H222; their absence after exposure to H226 or ER1D5 confirmed the cleavage of these isoforms at a site(s) downstream of ABC domains. Hence, hydroxylapatite extraction assay easily identifies ER isoforms that lack amino-termini. Figure 3b illustrates the presumed structure of these various ER isoforms, as well as their sizes as determined by SDS-PAGE.
Assessment of oestrogen receptor mRNA size in breast cancer
Total RMA from 46 breast tumours was qualitatively and quantitatively analyzed by hybridization with an ER-α full-length probe (Northern blotting). All tumours expressed a full-length 6.6-kb ER mRNA (small-sized species were not recorded). Moreover, a good correlation was obtained when the amount of 6.6-kb ER mRNA estimated by densitometry was compared with corresponding [3H]oestradiol-binding capacities (DCC values; Fig. 4). Hence, ER isoforms of low molecular weight did not appear to be encoded by truncated ER mRNAs, suggesting that they were generated by proteolysis. Whether such a phenomenon is an intracellular process is analyzed below.
Figure 4.
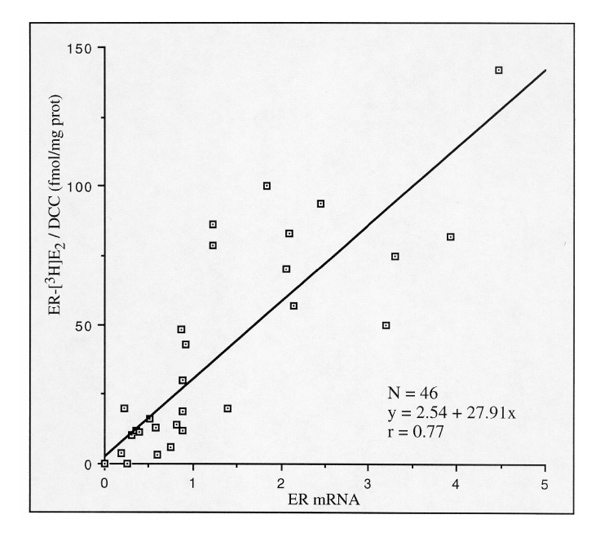
Correlation between ER mRNA and ER (by DCC assay). Breast tumours mRNA abundance (intensity of the 6.6-kb band) was expressed relatively to mRNA levels in MCF-7 cells (MCF-7 = 1).
Origin of cleaved oestrogen receptor forms
Cytosol samples of a series of breast tumours were labelled with [125I]TAZ in the presence of a cocktail of compounds that are known to inhibit the action of a wide range of proteolytic activities (final concentrations: 1 mmol/l for AEBSF, antipain and chymostatin; 5 mmol/l for PMSF; 0.1 mmol/l for leupeptin; and 0.1 mg/ml for calpastatin); this cocktail was added before or shortly after the homogenization of the samples. These inhibitors failed to maintain the native 67 kDa nature of the receptor as demonstrated by SDS-PAGE and autoradiography (Fig. 5); the ER electrophoretic pattern was not significantly modified, still showing bands of low molecular weight.
Figure 5.
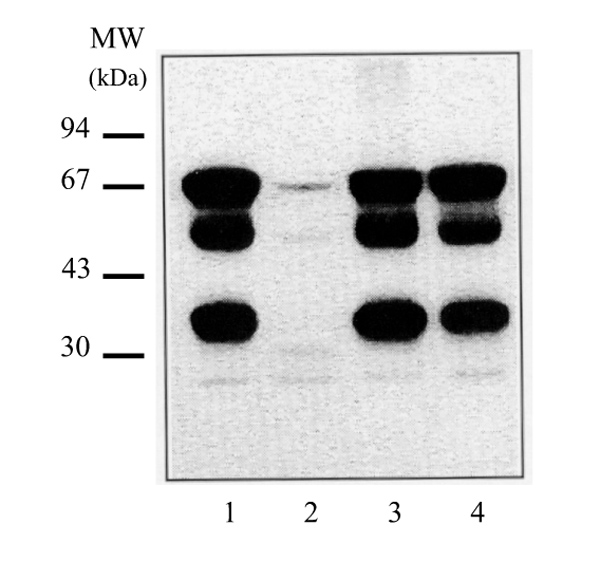
Effect of heat treatment on the relative expression of ER isoforms. Human breast cancer cytosols were heated at 37°C for 2 min in the presence or absence of protease inhibitors. They were then labelled with [125I]TAZ in the presence or absence of a 200-fold excess of radioinert oestradiol, immunoprecipitated with H-222 anti-ER monoclonal antibodies, and then subjected to SDS-PAGE. Lane 1, unheated control; lane 2, plus an excess of oestradiol; lane 3, 2 min heating in the absence of protease inhibitors; lane 4, 2 min heating in the presence of protease inhibitors.
In order to determine the molecular weight of intracellular ER molecules, we applied an in situ labelling approach along with an extraction procedure to minimize ER proteolysis [31,32]; tumour slices were incubated with [125I]TAZ at 37°C before homogenization and lysis at 100°C in the presence of SDS. Labelled proteins extracted with phenolic solution were precipitated by acetone and finally identified by SDS-PAGE. ER electrophoretic patterns remained almost identical to those usually found with cytosols, still showing bands of truncated receptors (mainly 50 and 37-28 kDa; Fig. 6). Hence, the high proportion of the low-molecular-weight isoforms appeared already to be present within tumour samples.
Figure 6.
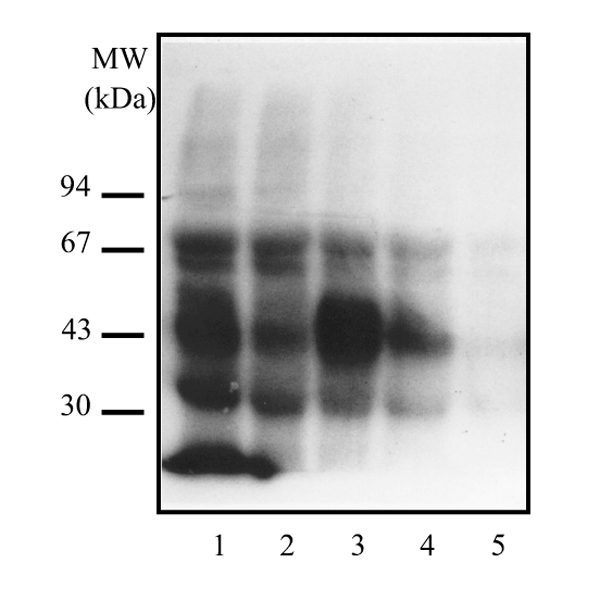
In situ labelling of ER with [125I]TAZ. Breast tissues slices (samples 1-4, ER positive; sample 5, ER-negative) were incubated with 1 nmol/l [125I]TAZ for 1 h at 0°C and the unbound ligand was removed. Then, all tissues were mixed with Krebs-Ringer phosphate buffer containing 1% SDS, 1.6 mmol/l EDTA and 2% β-mercaptoethanol, and briefly homogenized. After lysis at 100°C, proteins were extracted with phenol, precipitated by acetone and were finally analyzed by SDS-PAGE. The figure shows the electrophoretic patterns of these tissue ERs.
Finally, breast cancer cytosols were heated at 37°C for 2 min in the absence or presence of a cocktail of protease inhibitors in order to determine whether they possess proteolytic activities that are able to cleave native ER. Samples were subsequently adsorbed onto hydroxylapatite and successively subjected to KCl and ethanol extraction. No significant increase in EI was recorded (Table 2); inhibitors also failed to induce any drastic change in EI values, refuting the hypothesis that isoforms devoid of ABC domains may emerge during assay procedures. Of note, treatment at 37°C of control preparations of native ER (human recombinant ER, MCF-7 cells) resulted in similar behaviour. Hence, major proteolysis did not occur at the time of manipulation. Taken together, these data suggest that dominant ER proteolytic cleavage is an intracellular process.
Table 2.
Comparison of EIs of adsorbed ER onto hydroxylapatite matrix in the absence and presence of protease inhibitors
| EI (%) | ||||
| Without protease inhibitors | With protease inhibitors | |||
| ER | 0°C | 37°C | 0°C | 37°C |
| Cytosol from tumors | 69 ± 9 | 82 ± 16 | 61 ± 7 | 59 ± 13 |
| Recombinant hER | <10 | <10 | <10 | 12 ± 5 |
| MCF-7 cells | <10 | 11 ± 2 | 13 ± 02 | 16 ± 4 |
hER, human full-length ER.
Hydroxylapatite extraction index and heterogeneity of oestrogen receptors
Data reported here clearly show that low-molecular-weight ER isoforms extracted from hydroxylapatite matrix with KCl were not recognized by the ER1D5 monoclonal antibody. Because this antibody is often used in immunohistochemical assessment of ERs, we assessed whether immunohistochemical data are related to hydroxylapatite ER adsorption characteristics measured in cytosolic preparations from the corresponding tumours. For this purpose, cytosols from a set of 15 ER-positive tumours (by DCC assay), for which nuclear ERs had been detected by immunohistochemistry (IS cutoff ≥ 5), were labelled with [3H]oestradiol and were then subjected to hydroxylapatite assay (Table 3). A significant correlation between the two sets of measurement was recorded (IS versus total number of binding sites assayed by hydroxylapatite, r = 0.71; P < 0.001; Table 3). Sequential extraction of bound [3H]oestradiol from hydroxylapatite with KCl and ethanol revealed an EI of over 30% in the large majority of these cytosols (11/15), indicating a high frequency of cleaved ER. Of note, no significant correlation between IS and EI data (r = 0.2; P > 0.05) was detected, clearly establishing that identification of ABC domains within the cell (indicated by IS) does not imply the presence of (native) full-length ER in the corresponding cytosol.
Table 3.
Comparative analysis of ER by immunohistochemistry (ER1D5), DCC ([3H]oestradiol-binding) and hydroxylapatite ([3H]oestradiol-binding and EI)
| Tissular ER | Cytosolic ER | |||
| IHC | DCC | Hydroxylapatite | ||
| Case no. | IS | n (fmol/mg protein) | n (fmol/mg protein)* | EI (%)† |
| 1 | 10 | 39 | 27 (20 + 7) | 74.5 |
| 2 | 20 | 69 | 62 (54 + 8) | 86.8 |
| 3 | 20 | 32 | 67 (21 + 46) | 31.5 |
| 4 | 40 | 38 | 35 (12 + 23) | 34.4 |
| 5 | 130 | 23 | 130 (91 + 39) | 70.0 |
| 6 | 130 | 83 | 151 (62 + 89) | 41.1 |
| 7 | 160 | 163 | 166 (30 + 136) | 18.0 |
| 8 | 160-200 | 257 | 209 (58 + 151) | 27.5 |
| 9 | 260 | 23 | 103 (64 + 39) | 62.1 |
| 10 | 360 | 265 | 230 (114 + 116) | 49.6 |
| 11 | 360-400 | 400 | 360 (190 + 170) | 52.6 |
| 12 | 360-400 | 354 | 337 (82 + 255) | 24.5 |
| 13 | 360-400 | 178 | 193 (60 + 133) | 31.0 |
| 14 | 360-400 | 92 | 174 (36 + 138) | 20.8 |
| 15 | 360-400 | 312 | 390 (156 + 234) | 40.0 |
*Hydoxylapatite corresponds specifically to [3H]oestradiol extracted successively from hydroxylapatite with KCl and ethanol (ie 27 = 20 + 7). †EI = ([3H] oestradiol) [KCl] × 100/([3H]oestradiol) [KCl] + ([3H]oestradiol) [EtOH]. IHC, immunohistochemistry; IS, immunostaining score.
Discussion
Cytosolic ER contains several sites that are sensitive to proteolytic cleavage. On limited digestion, ER fragments of 50 and 37 kDa and lower molecular weight were recorded in uterine cytosol preparations [42]. The presence of such isoforms has generally been attributed to proteolysis of the native ER [43,44]. Regarding ER prepared from breast cancer samples, we previously suggested [11] that proteolytic events that operate at the time of tissue processing might explain the observed extreme ER molecular heterogeneity. Data presented here clearly refute this hypothesis. A high proportion of these isoforms is derived from an intracellular proteolytic activity, as shown by electrophoresis of in situ [125I]TAZ-labelled ER and addition of protease inhibitors to cytosols. In this regard, the present results are in agreement with those reported recently by Navarro et al [45] who demonstrated that the inclusion of PMSF, aprotinin and leupeptin to human uterine cytosolic ER did not prevent the 8S to 4S transformation known to be associated with ER cleavage [46]. Hence, truncated ER isoforms are probably produced before contact with protease inhibitors.
On breast tumour slices, staining intensities (ISs) established with the ER1D5 monoclonal antibody used here have often been reported to be correlated with [3H]oestradiol binding capacities of corresponding cytosolic extracts [5,6,47]. Hence, a consensus seems to exist between immunohistochemical detection of the amino-terminal region of ER (ABC domains) and biochemical measurement of its carboxyl-terminal region (E domain). In agreement with this, we reported a significant correlation between immunohistochemical data and [3H]oestradiol-binding capacities measured by hydroxylapatite assay; ABC domains are indeed required for hydroxylapatite adsorption [12]. For both [3H]oestradiol- and [125I]TAZ labelled-ER isoforms, we found an excellent correlation between DCC values and hydroxylapatite adsorption data, suggesting the concomitant presence of free ABC and E domains in most cytosols (proteolytic fragments physically separated from each other at high ionic strength; high EI values). Hence, these domains would be held together within the cell nucleus because of the tendency of the receptor to be included into oligomeric structures. Corroborating these views of intracellular association between ER fragments, Fritsch et al [42] described large oligomeric complexes containing proteolysis-generated ER fragments in rat uterus.
When expressed individually as separate polypeptides in ER-negative breast cancer cells (MDA-MB-231), neither ABCD (AF-1 containing) nor EF (AF-2 containing) domains activated transcription from a hormone-inducible reporter gene (3ERE-pS2-CAT) [48]. Coexpression of the steroid receptor coactivator-1 protein with either ABCD or EF alone was also ineffective [49]. On the contrary, when expressed together, amino- and carboxyl-terminal ER regions interacted in an oestradiol-dependent manner to reconstitute ER transcriptional activity [48]. In this context, it should be emphasized that steroid receptor coactivator-1 has also been suggested to promote the association between separate amino- and carboxyl-terminal regions of ER, allowing full ER activation [49]. In view of these data, one may speculate that the secondary and tertiary structures of the receptor peptide required for its transcriptional ability would not be lost when it has been subjected to endogenous proteolytic activity. Hence, intracellular ER fragmentation should not necessarily imply lack of transcriptional activity.
Endogenous proteolysis is a mechanism of regulation of many cellular processes, such as cell cycle progression and transcriptional regulation [50,51]. Recent data concerning cell-specific regulation of oestrogen target gene expression in various rat tissues [52] and cell lines [53] demonstrated different levels of coactivators and corepressors. Hence, protein environment would be a factor of major importance in the ability of ER to transcribe genes. Although not presently demonstrated in breast cancer, the possibility of heterogeneous association between cleavage products of ER and other regulatory proteins could be proposed, generating a situation that may mimic the behaviour of chimaeric receptors. Thus, domain-swapping experiments in which the ER amino-terminal domain was switched with that of the glucocorticoid receptor yielded a receptor that upregulates transcription of glucocorticoid-responsive target genes when treated with oestradiol [54]. Investigation of homogenous association and/or heterogeneous association between cleaved ER regions and regulatory proteins would be informative with regard to the validity of this hypothesis.
In view of these considerations, it seems that analysis of the biological role and potential clinical relevance of endogenous ER proteolysis is of prime importance. The hydroxylapatite extraction assay described here, which is extremely simple, would provide a criteria for the detection of tumours characterized by a high amount of full-length ER (low EI). Such tumours may perhaps be considered an indication for adjuvant hormone therapy if we refer to earlier sucrose gradient sedimentation data concerning hormone dependency of tumours that express native and/or cleaved ER forms [17]; very high EI may perhaps be an index of poor prognosis. In practice, for clinical correlation studies devoted to analysis of these hypotheses, we propose that a hydroxylapatite extraction assay should be performed as a complement to the usual immunohistochemical test (or Abbott's ER enzyme immunoassay [4]), in view of the small size of most tumours that restrict the cytosol amount. If our speculations concerning the biological/clinical relevance of the detection of cleaved ER isoforms are verified, then the introduction of such a hydroxylapatite assay into routine practice would be helpful to orientate therapy.
Acknowledgments
Acknowledgement
This work was supported by grants from the Fonds Medic and the Fonds J-C Heuson. YM is a recipient of a grant from the Fonds National de la Recherche Scientifique - Télévie. ML is a recipient of a grant from Les Amis de l'Institut Bordet. FJ is a recipient of a grant from the Fonds J-C Heuson. We thank Dr A Buggenhout for reading of the manuscript.
References
- Heuson JC, Leclercq G, Longeval E, Deboel MC, Mattheiem WH, Heimann R (Edited by McGuire WL, Carbone PP, Volmer EP. New York: Raven Press.) Estrogen receptors: pronostic significance in breast cancer. Estrogen Receptors in Human Breast Cancer. 1975. pp. 57–72.
- McGuire WL. Hormone receptors: their role in predicting prognosis and response to endocrine therapy. Semin Oncol. 1978;5:428–433. [PubMed] [Google Scholar]
- DeSombre ER, King WJ, Green GL, Jensen EV (Edited by Jordan VC. Wisconsin: University of Wisconsin Press.) Estrogen receptor studies. Laboratory investigations and clinical applications. . Estrogen/Antiestrogen Action and Breast Cancer Therapy. 1986. pp. 303–323.
- Leclercq G, Bojar H, Goussard J, Nicholson RI, Pichon MF, Piffanelli A, Pousette A, Thorpe S, Lonsdorfer M. Abbott monoclonal enzyme immunoassay measurement of estrogen receptors in human breast cancer: a european multicenter study. Cancer Res. 1986;46 (Suppl):S4233–S4236. [PubMed] [Google Scholar]
- al Saati T, Clamens S, Cohen-Knafo E, Faye JC, Prats H, Coindre JM, Wafflart J, Caveriviere P, Bayard F, Delsol G. Production of monoclonal antibodies to human estrogen-receptor protein (ER) using recombinant ER (RER). Int J Cancer. 1993;55:651–654. doi: 10.1002/ijc.2910550423. [DOI] [PubMed] [Google Scholar]
- Kell DI, Kamel OW, Rouse RV. Immunohistochemical analysis of breast carcinoma estrogen and progesterone receptors in paraffin-embedded tissues: correlation of clones ER1D5 and 1A6 with a cytosol-based hormone receptor assay. Appl Immunohistochem. 1993;1:275–281. [Google Scholar]
- Wong SY, Purdie A, Sewell HF, Wilkinson L, Angus B, Westley B, Horne CH. Immunohistochemical assessment of ER-P31, a mouse anti-oestrogen receptor protein monoclonal antibody in human breast cancers: comparison with ER-ICA (Abbott) and radioligand assays. Tumor Biol. 1991;12:16–23. doi: 10.1159/000217683. [DOI] [PubMed] [Google Scholar]
- Clark GM, McGuire WL. Steroid receptors and other pronostic factors in primary breast cancer. Semin Oncol. 1988;15:20–25. [PubMed] [Google Scholar]
- Wittliff JL. Steroid-hormone receptors in breast cancer. . Cancer. 1984;53:630–643. doi: 10.1002/1097-0142(19840201)53:3+<630::aid-cncr2820531308>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- Sluyser M, Wittliff JL. Influence of estrogen receptor variants in mammary carcinomas on the pronostic reliability of the receptor assay. . Mol Cell Endocrinol. 1992;85:83–88. doi: 10.1016/0303-7207(92)90127-r. [DOI] [PubMed] [Google Scholar]
- Maaroufi Y, Trivedi S, Leclercq G. Major molecular weight heterogeneity of estrogen receptor from breast cancer is not related to neoplasia. Cancer Biochem Biophys. 1995;15:67–78. [PubMed] [Google Scholar]
- Maaroufi Y, Leclercq G. Importance of A/B and C domains of the estrogen receptor for its adsorption to hydroxylapatite. J Ster Bioch Mol Biol. 1994;48:155–163. doi: 10.1016/0960-0760(94)90262-3. [DOI] [PubMed] [Google Scholar]
- Murayama A, Fukai F, Murachi T. Action of calpain on the basic estrogen receptor molecule of porcine uterus. J Biochem. 1984;95:1697–1704. doi: 10.1093/oxfordjournals.jbchem.a134783. [DOI] [PubMed] [Google Scholar]
- Thole HH, Maschler I, Jungblut PW. Surface mapping of the ligand-filled C-terminal half of the porcine estradiol receptor by restricted proteolysis. Eur J Biochem. 1995;231:510–516. doi: 10.1111/j.1432-1033.1995.tb20726.x. [DOI] [PubMed] [Google Scholar]
- Seielstad DA, Carlson KE, Kushner PJ, Greene GL, Katzenellenbogen JA. Analysis of the structural core of the human estrogen receptor ligand binding domain by selective proteolysis/mass spectrometry analysis. . Biochemistry. 1995;34:12605–12615. doi: 10.1021/bi00039a016. [DOI] [PubMed] [Google Scholar]
- Horigome TF, Ogata TS, Golding F, Korach KS. Estradiol-stimulated proteolytic cleavage of the estrogen receptor in mouse uterus. Endocrinology. 1988;123:2540–2548. doi: 10.1210/endo-123-5-2540. [DOI] [PubMed] [Google Scholar]
- Wittliff JL, Savlov ED (Edited by McGuire WL, Carbone PP, Volmer EP. New York: Raven Press.) Estrogen binding capacity of cytoplasmic forms of estrogen receptors in human breast cancer. Estrogen Receptors in Human Breast Cancer. 1975. pp. 73–86.
- Freedman B, Hawkins RA. Sedimentation profiles of the estrogen receptor from breast tumors of man and rat using vertical-tube rotor centrifugation. J Endocrinol. 1980;86:431–442. doi: 10.1677/joe.0.0860431. [DOI] [PubMed] [Google Scholar]
- Piccart MJ, Muquardt C, Bosman C, Pirotte P, Veenstra S, Grillo F, Leclercq G. Comparison between tritiated estradiol and tamoxifen aziridine for measuring estrogen receptors in human breast cancer cytosols. . J Natl Cancer Inst. 1991;83:1553–1559. doi: 10.1093/jnci/83.21.1553. [DOI] [PubMed] [Google Scholar]
- Pahl HL, Bäuerle PA. Control of gene expression by proteolysis. . Curr Opin Cell Biol. 1996;8:340–347. doi: 10.1016/s0955-0674(96)80007-x. [DOI] [PubMed] [Google Scholar]
- Spataro V, Norbury C, Harris AL. The ubiquitin-proteasome pathway in cancer. Br J Cancer. 1998;77:448–455. doi: 10.1038/bjc.1998.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plombella VJ, Rando OJ, Goldberg AL, Maniatis T. The ubiquitin-proteasome pathway is required for processing the NF-kappa B1 precursor protein and the activation of NF-kappa B. Cell. 1994;78:773–785. doi: 10.1016/s0092-8674(94)90482-0. [DOI] [PubMed] [Google Scholar]
- Bäuerle PA, Baltimore D. NF-κB: ten years after. Cell. 1996;87:13–20. doi: 10.1016/s0092-8674(00)81318-5. [DOI] [PubMed] [Google Scholar]
- El Khissiin A, Leclercq G. Implication of proteasome in estrogen receptor degradation. FEBS Lett. 1999;448:160–166. doi: 10.1016/s0014-5793(99)00343-9. [DOI] [PubMed] [Google Scholar]
- Nawaz Z, Lonard DM, Dennis AP, Smith CL, O'Malley BW. Proteasome-dependent degradation of the human estrogen receptor. . Proc Natl Acad Sci USA. 1999;96:1858–1862. doi: 10.1073/pnas.96.5.1858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alarid ET, Bakopoulos N, Solodin N. Proteasome-mediated proteolysis of estrogen receptor: a novel component in autologous down-regulation. Mol Endocrinol. 1999;13:1522–1534. doi: 10.1210/mend.13.9.0337. [DOI] [PubMed] [Google Scholar]
- Wooge CH, Nilsson GM, Heierson A, McDonnell DP, Katzenellenbogen BS. Structural requirements for high affinity ligand binding by estrogen receptors: a comparative analysis of truncated and full length estrogen receptors expressed in bacteria, yeast, and mammalian cells. Mol Endocrinol. 1992;6:861–869. doi: 10.1210/mend.6.6.1495491. [DOI] [PubMed] [Google Scholar]
- Leclercq G, Legros N, Piccart MJ. Accumulation of a non-binding form of estrogen receptor in MCF-7 cells under hydroxytamoxifen treatment. . J Steroid Biochem Mol Biol. 1992;41:545–552. doi: 10.1016/0960-0760(92)90381-r. [DOI] [PubMed] [Google Scholar]
- EORTC: Breast Cancer Cooperative Group. Revision of the standards for the assessment of hormone receptors in human breast cancer. . Eur J Cancer. 1980;16:1513–1515. doi: 10.1016/0014-2964(80)90064-x. [DOI] [PubMed] [Google Scholar]
- Scatchard G. The attraction of proteins for small molecules and ions. Ann NY Acad Sci. 1949;51:660–672. [Google Scholar]
- Terenius L. Selective retention of estrogen isomers in estrogen-dependent breast tumors of rats demonstrated by in vitro methods. Cancer Res. 1968;28:328–337. [PubMed] [Google Scholar]
- Toulas C, Guilbaud N, Delassus F, Bayard F, Faye JC. Modulation of the estrogen receptor structure, evidence of a heterogeneity. J Receptor Res. 1990;10:1–9. doi: 10.3109/10799899009064654. [DOI] [PubMed] [Google Scholar]
- Borràs M, Lacroix M, Legros N, Leclercq G. Estrogen receptor-negative/progesterone receptor-positive Evsa-T mammary tumor cells: a model for assessing the biological property of this peculiar phenotype of breast cancers. Cancer Lett. 1997;120:23–30. doi: 10.1016/s0304-3835(97)00285-1. [DOI] [PubMed] [Google Scholar]
- Deugnier Y, Margules S, Brissot P, Deugnier MA, Bisconte JC, Bourel M, Ferrand B. Comparative study between biochemical and histological methods and image analysis in liver iron overload. J Clin Pathol . 1982;35:45–51. doi: 10.1136/jcp.35.1.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weintraub J, Weintraub D, Redard M, Vassilakos P. Evaluation of estrogen receptors by immunocytochemistry on fine-needle aspiration biopsy specimens from breast tumors. Cancer. 1987;60:1163–1172. doi: 10.1002/1097-0142(19870915)60:6<1163::aid-cncr2820600602>3.0.co;2-d. [DOI] [PubMed] [Google Scholar]
- Maaroufi Y, Ben Hardouze A, Leclercq G. Decrease of estrogen receptor binding capacity by calcium. J Receptor Signal Transduction Res. 1997;17:833–853. doi: 10.3109/10799899709039159. [DOI] [PubMed] [Google Scholar]
- Powel BL, de la Graza M, Clark GM, McGuire WL. Estrogen receptor measurement in low protein breast cancer cytosols. Breast Cancer Res Treat. 1981;1:33–35. doi: 10.1007/BF01807889. [DOI] [PubMed] [Google Scholar]
- Trivedi S, Piccart M, Muquardt C, Gilot N, Hadiy S, Patel D, Leclercq G. Tamoxifen aziridine labeling of the estrogen receptor: potential utility in detecting biologically aggressive breast tumors. Breast Cancer Res Treat. 1996;40:231–241. doi: 10.1007/BF01806811. [DOI] [PubMed] [Google Scholar]
- Maaroufi Y, Quivy J, Trivedi S, Gilot N, Leclercq G. 4-iodotamoxifen aziridine, a new affinity labeling agent for the rapid detection of estrogen receptor isoforms. J Steroid Biochem Mol Biol. 1998;67:95–104. doi: 10.1016/s0960-0760(98)00102-2. [DOI] [PubMed] [Google Scholar]
- Harlow KW, Smith DN, Katzenellenbogen JA, Greene GL, Katzenellenbogen BS. Identification of cysteine 530 as the covalent attachment site of an affinity-labeling estrogen (ketonenestrol aziridine) and antiestrogen (tamoxifen aziridine) in the human estrogen receptor. J Biol Chem . 1989;264:17476–17485. [PubMed] [Google Scholar]
- Reese JC, Wooge CH, Katzenellenbogen BS. Identification of two cysteines closely positioned in the ligand binding pocket of the human estrogen receptor: roles in ligand binding and transcriptional activity. . Mol Endocrinol. 1992;6:2160–2166. doi: 10.1210/mend.6.12.1491695. [DOI] [PubMed] [Google Scholar]
- Fritsch M, Anderson I, Gorski J. Structural characterization of trypsinized estrogen receptor. Biochemistry. 1993;32:14000–14008. doi: 10.1021/bi00213a033. [DOI] [PubMed] [Google Scholar]
- Wilson EA, Yang F, Rees D. Estradiol and progesterone binding in uterine leiomyomata and in normal uterine tissues. Obstet Gynecol . 1980;55:20–24. [PubMed] [Google Scholar]
- Gregory MR, Notides AC. Characterization of two uterine proteases and their actions on the estrogen receptor. Biochemistry . 1982;21:6452–6458. doi: 10.1021/bi00268a021. [DOI] [PubMed] [Google Scholar]
- Navarro D, León L, Chirino R, Fernández L, Pestano J, Díaz-Chico BN. The two native estrogen receptor forms of 8S and 4S present in cytosol from human uterine tissues display opposite reactivities with the antiestrogen tamoxifen aziridine and the estrogen responsive element. . J Steroid Biochem Mol Biol. 1998;64:49–58. doi: 10.1016/s0960-0760(97)00134-9. [DOI] [PubMed] [Google Scholar]
- Schneider SL, Dao TL. Effect of Ca2+ and salt on forms of estradiol cytoplasmic receptor in human neoplasmic breast tissue. . Cancer Res. 1977;37:382–387. [PubMed] [Google Scholar]
- Mauri FA, Veronese S, Frigo B, Girlando S, Losi L, Gambacorta M, Dalla Palma P, Barbareschi M. ER1D5 and H222 (ER-ICA) antibodies to human estrogen receptor protein in breast carcinomas: results of a multicentric comparative study. Appl Immunohistochem. 1994;2:157–163. [Google Scholar]
- Kraus WL, McInerney EM, Katzenellenbogen BS. Ligand-dependent, transcriptionally productive association of the amino- and carboxyl-terminal regions of a steroid hormone nuclear receptor. Proc Natl Acad Sci USA. 1995;92:12314–12318. doi: 10.1073/pnas.92.26.12314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McInerney EM, Tsai MJ, O'Malley BW, Katzenellenbogen BS. Analysis of estrogen receptor transcriptional enhancement by a nuclear hormone receptor coactivator. Proc Natl Acad Sci USA. 1996;93:10069–10073. doi: 10.1073/pnas.93.19.10069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hochstrasser M. Ubiquitin, proteasomes, and the regulation of intracellular protein degradation. Curr Opin Cell Biol. 1995;7:215–223. doi: 10.1016/0955-0674(95)80031-x. [DOI] [PubMed] [Google Scholar]
- King RW. How proteolysis drives the cell cycle. Science . 1996;274:1652–1659. doi: 10.1126/science.274.5293.1652. [DOI] [PubMed] [Google Scholar]
- Misiti S, Schomburg L, Yen PM, Chin WW. Expression and hormonal regulation of coactivator and corepressor genes. Endocrinology . 1998;139:2493–2500. doi: 10.1210/endo.139.5.5971. [DOI] [PubMed] [Google Scholar]
- Folkers GE, van der Burg B, van der Saag PT. Promoter architecture, cofactors, and orphan receptors contribute to cell-specific activation of the retinoid acid receptor beta2 promoter. . J Biol Chem. 1998;273:32200–32212. doi: 10.1074/jbc.273.48.32200. [DOI] [PubMed] [Google Scholar]
- Green S, Chambon P. Oestradiol induction of a glucocorticoid-responsive gene by a chimaeric receptor. Nature . 1987;325:75–78. doi: 10.1038/325075a0. [DOI] [PubMed] [Google Scholar]