

Short abstract
An emerging family of structurally distinct dual-specificity serine, threonine and tyrosine phosphatases that act on MAP kinases consists of ten members in mammals, and members have been found in animals, plants and yeast. Three subgroups have been identified that differ in exon structure, sequence and substrate specificity.
Abstract
Mitogen-activated protein MAP kinases are key signal-transducing enzymes that are activated by a wide range of extracellular stimuli. They are responsible for the induction of a number of cellular responses, such as changes in gene expression, proliferation, differentiation, cell cycle arrest and apoptosis. Although regulation of MAP kinases by a phosphorylation cascade has long been recognized as significant, their inactivation through the action of specific phosphatases has been less studied. An emerging family of structurally distinct dual-specificity serine, threonine and tyrosine phosphatases that act on MAP kinases consists of ten members in mammals, and members have been found in animals, plants and yeast. Three subgroups have been identified that differ in exon structure, sequence and substrate specificity.
The mitogen-activated protein (MAP) kinases are evolutionary conserved enzymes that play an important role in orchestrating a variety of cellular processes, including proliferation, differentiation and apoptosis [1,2]. To date, four major discrete groups of MAP kinases have been identified in mammalian cells, known as the extracellular signal-regulated kinases (ERK1 and ERK2), the c-Jun amino-terminal kinases (JNK1, JNK2 and JNK3), the p38 kinases (p38α, p38β, p38γ and p38δ) and ERK5/Big MAP kinase 1 (BMK1). MAP kinases are part of a three-tiered cascade consisting of a MAP kinase, a MAP kinase kinase (MAPKK, MKK or MEK) and a MAP kinase kinase kinase (MAPKKK or MEKK). Phosphorylation of the threonine and tyrosine residues in the T-loop of the MAP kinase, by its upstream kinase, results in activation. The activation of MAP kinases is not a simple switch, however, as both the duration and magnitude of activation is crucial in determining the physiological outcome in cells [3]. Thus, it seems likely that dephosphorylation of the MAP kinases is vital for their control. This is achieved by removal of phosphate groups from either the threonine residue or the tyrosine residue, or both. Both protein serine/threonine phosphatases and protein tyrosine phosphatases have been reported to dephosphorylate MAP kinases. In the past decade, however, some dual-specificity phosphatases (DSPs) have been recognized as key players for inactivating different MAP kinase isoforms; this class of phosphatases has been designated MAP kinase phosphatases (MKPs).
Although the MKPs form a structurally and functionally distinct subclass among the large number of protein phosphatases present in eukaryotic genomes, there is a great deal of confusion regarding their classification. In this article we discuss the family of 'classical' MKPs that have been definitively shown to play a role in the de-activation of MAP kinases. Where relevant, we also mention the closely related low-molecular-weight phosphatases, whose role in MAP kinase regulation is controversial. Other more distantly related DSP families, including members of the FYVE family, which have high sequence similarity to myotubularin, and of the Cdc25 family, which dephosphorylate cyclin-dependent kinases, are beyond the scope of this review.
Gene organization and evolutionary history
Gene organization
To date, 10 genes encoding members of the classical MKP family have been isolated and characterized from mammalian genomes (see Table 1). They all share some common features, including an extended active-site motif with significant sequence similarity to the corresponding region of the VH-1 protein tyrosine phosphatase that was isolated from vaccinia virus [4]. In addition, their amino termini contain two short regions that are homologous to sequences that flank the Cdc25 phosphatase catalytic site [5] and a cluster of basic amino-acid residues that play an important role in binding to the MAP kinases [6]. The MKPs can be further subdivided into groups depending on their substrate specificity for the MAP kinases, subcellular localization (nuclear, cytoplasmic or both) and the structural organization of their genes.
Table 1.
Classification and chromosomal and subcellular localization of DUSP genes
| DUSP | Subgroup | Human protein | Species ortholog | Chromosomal localization | Subcellular localization | References |
| DUSP1 | I | hVH1/CL100 | MKP-1/3CH134 (m) | 5q35 | Nuclear | [8,60,61,62] |
| DUSP2 | I | hPAC-1 | --- | 2p11.2-2q11 | Nuclear | [10,42,62] |
| DUSP4 | I | hVH2/TYP1 | MKP-2 (r) | 8p12-p11 | Nuclear | [42,63,64,65] |
| DUSP5 | I | hVH3/B23 | Cpg21 (r) | 10q25 | Nuclear | [48,62,66] |
| DUSP6 | II | PYST1 | MKP-3/rVH6 (r) | 12q22-q23 | Cytosolic | [49,50,51,65] |
| DUSP7 | II | PYST2/B59 | MKP-X (r) | 3p21 | Cytosolic | [49,51,65,67] |
| DUSP8 | III | hVH5 | M3/6 (m) | 11p15.5 | Nuclear /cytosolic | [13,43,44] |
| DUSP9 | II | MKP-4 | --- | Xq28 | Nuclear /cytosolic | [45] |
| DUSP10 | II(?) | MKP-5 | --- | 1q32 | Nuclear /cytosolic | [46,52] |
| DUSP16 | III | MKP-7 | MKP-M (m) | 12p12 | Cytosolic | [14,15,36] |
Compilation of DUSP genes identified in the human and other mammalian genomes. The DUSPs are classified into subgroups I, II or III (see text for details). The various alternative names for each gene in human (h) or rat (r) or mouse (m) are given. The chromosomal location of each gene and the subcellular localization of the protein product of each gene are also indicated. Multiple names are used in the literature for each member of the MAP kinase phosphatase gene family. To avoid confusion in this article, we have used the gene names recommended by the human gene nomenclature committee, which uses the root DUSP (for dual specificity phosphatase). Note that the designation of DUSP16 is provisional.
A gene closely related to the MKP family is STYX. The protein encoded by STYX contains many of the hallmarks of the MKPs but lacks a critically important catalytically active cysteine residue, having a glycine instead [7]. Interestingly, alteration of this glycine residue to cysteine renders the STYX protein catalytically active, suggesting that other critical residues have been retained. STYX appears to function as a phosphotyrosine/phosphoserine/phosphothreonine-binding protein analogous to the SH2 and PTB domains. It is unclear whether it plays any role in signaling through the MAP kinase pathways, however.
Evolutionary history
On the basis of structures predicted from genomic sequence, the MKPs can readily be divided into the following subgroups: subgroup I, DUSP1, DUSP2, DUSP4 and DUSP5; subgroup II, DUSP6, DUSP7, DUSP9 and DUSP10; and subgroup III, DUSP8 and DUSP16 (Figure 1, Table 1). Division into these three subgroups is also supported by phylogenetic analysis (Figure 2), and to a considerable extent by substrate preference.
Figure 1.
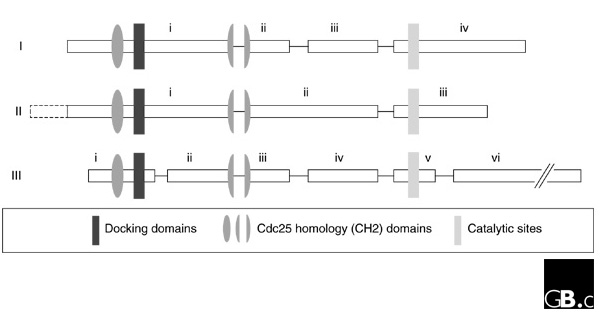
Gene structure of the DUSPs. Three distinct gene structures have been described for the DUSPs, allowing their classification into subgroups I, II and III. Structural motifs (CH2 domains, docking domain and catalytic domain) in the encoded proteins are indicated by shaded shapes and exons by boxes and roman numerals. The dashed box in subgroup II is an alternatively spliced exon.
Figure 2.
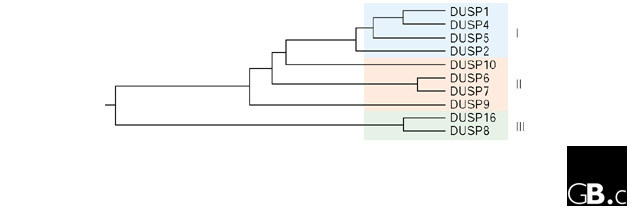
Phylogenetic analysis of DUSP sequences. Human DUSP amino-acid sequences were aligned and a phylogenetic tree derived using Clustal W [68] on DNASTAR. The length of the branches is proportional to sequence divergence between proteins. Subgroups I, II, and III are shaded (see text for further details).
The genes of subgroup I consist of four exons [8,9,10,11], and the exact positions of the introns are highly conserved. The first exon encodes the amino terminus of each of the three proteins and ends within the second of the Cdc25 homology (CH2) domains. The active-site motif of all four proteins is encoded within exon 4, and the length of exon 3 is identical for all these DUSPs, suggesting that they arose from a common ancestral gene.
Subgroup II genes (DUSP6,7 and 9, and provisionally 10) consist of three exons [12]. As with subgroup I, exon 1 encodes the amino terminus and the 3'-most exon encodes the active site. Unlike subgroup I, however, the central portion of the protein is encoded by a single exon. DUSP10 has an intron/exon structure consistent with a placement within subgroup II, but it shares some features with members of subgroup III, such as a similar MAP kinase docking-site motif and similar substrate specificity. In addition, it has an extended amino-terminal region that is not present in any of the other DUSPs. Assignment of DUSP10 to subgroup II or III or to its own subgroup may need re-evaluation in the future.
The genes of subgroup III (DUSP8 and DUSP16) consist of six exons [13,14,15]. The lengths of exons 2, 3, 4 and 5 are identical in the two genes. Unlike the two previous subgroups, however, exon 1 encodes only the first of the CH2 domains. Exon 2 ends within the same region of the second CH2 domain as does exon 1 of subgroups I and II, however. The central portion of the protein is encoded by two exons, as in subgroup I, rather than the single exon of subgroup II. This suggests that subgroup II genes may have diverged earlier and that subgroup I and III genes may be more closely related to each other. Exon 5 encodes the catalytic site of these DUSPs but not the carboxyl terminus; the sixth exon encodes the large extended carboxyl terminus, with the intron between the catalytic domain and the carboxyl terminus of the protein being diagnostic of this subgroup.
Overall, the high conservation of some intron/exon boundaries within all three subgroups of DUSPs indicates a common ancestral gene for each subgroup. The similarities of substrate specificity among subgroup members suggest that these ancestral genes are themselves likely to have shown some substrate preference.
Dual specificity phosphatases have been found in a wide variety of phylogenetically distinct eukaryotes, including yeasts, worms, flies and plants, as well as mammals. Although budding yeast does not encode any predicted protein tyrosine kinases, it has 16 dual specificity and low-molecular-weight phosphatases and five phosphatase-like STYX genes. A direct role has been demonstrated for the budding yeast dual specificity phosphatase gene MSG5 in the regulation of the MAP kinase Fus3 [16]. Six MKPs have been noted in the Drosophila melanogaster genome sequence [17]. Of these, two have been characterized: puckered has been shown to encode a MKP that inactivates basket, the Drosophila equivalent of mammalian JNK [18]; in addition, a Drosophila DUSP6-like gene has recently been identified [19]. The sequence of the Caenorhabditis elegans genome includes 26 predicted DSPs, including some MKPs [20]. One of these, LIP-1, has been suggested to inactivate MAP kinase, mediating inhibition by the ligand Notch of a signaling pathway involving the GTPase Ras, during C. elegans vulval development [21]. Moreover, at least two MKPs have been identified in Arabidopsis thaliana, one of which is required for relief from genotoxic stress [22].
Characteristic structural features
Catalytic domain
All the DUSPs share strong amino-acid sequence identity over their catalytic domains (37%-50%), with members of certain subgroups sharing up to 75% identity (for example, DUSP6 with DUSP7 and DUSPQ or DUSP8 with DUSP16). The catalytic domain invariably contains the highly conserved consensus sequence DX26(V/L)X(V/I)HCXAG(I/V)SRSXT(I/V)XXAY(L/I)M, in the single-letter amino-acid code where X is any amino acid. The three amino acids indicated in bold have been shown to be absolutely essential for catalysis. The cysteine is required for the nucleophilic attack of the phosphorus of the substrate and the formation of the thiol-phosphate intermediate; the conserved arginine binds the phosphate group of the phosphotyrosine or phospho-threonine, enabling transition-state stabilization; whereas the aspartate enhances catalysis by protonating the leaving group oxygen [23].
Determination of the crystal structure of the catalytic domain of DUSP6 [24] has revealed that this domain adopts a shallow cleft conformation, similar to that of VH1-related phosphatase (VHR) [25]. Within this cleft, the phosphatase is able to accommodate both phosphotyrosine and phosphothreonine side chains. In the absence of substrate, DUSP6 exists in a low-activity state, with key residues (Arg299 and Asp262 in DUSP6) disengaged from interaction. Upon binding of the substrate ERK2, however, the active-site residues are rearranged, adopting a catalytically active conformation [24] and enabling the dephosphorylation of both Thr183 and Tyr185 in ERK2 (Figure 3b). In particular, Asp262 (the conserved aspartate in the consensus sequence) is located in a loop, 5.5 Å away from the nucleophilic cysteine and arginine in the active site. This suggests that the loop must undergo conformational rearrangement for activation. Upon binding of the substrate ERK2, closure of the Asp262 loop over the active site occurs, positioning the aspartate residue for catalysis. Thus DUSP6 adopts a catalytically active conformation [24].
Figure 3.
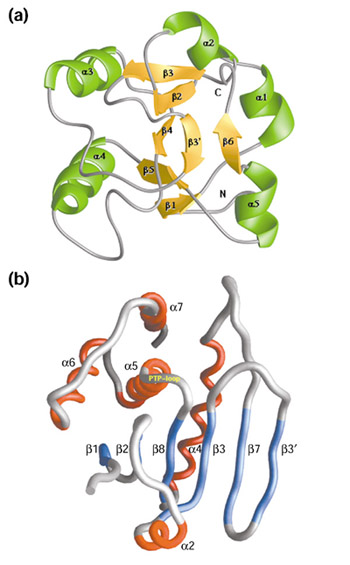
Three-dimensional structure of DUSP6. (a) The amino-terminal Erk-binding (EB) domain, reproduced with permission from [38]. (b) The catalytic domain, reproduced with permission from [24].
CH2 domains
The amino terminus is much less conserved in the DUSPs than the catalytic domain. Nevertheless, all DUSPs contain at their amino terminus two conserved regions that show similarity to the Ccd25 phosphatase, designated CH2 domains. In Cdc25, these domains flank the catalytic site of the enzyme, whereas in the DUSPs they are found upstream of the active site [5]. They have been suggested to play a role in substrate binding, but this has yet to be formally demonstrated.
Docking sites
All DUSPs have, near to their amino termini, a MAP kinase docking site, which consists of a cluster of positively charged amino acids [6]. It has been proposed that the number of consecutive positively charged residues in this docking site of MKPs may play a role in determining binding specificity and therefore catalytic activity (Figure 4) [14]. The corresponding docking site on the MAP kinases has been shown to consist of negatively charged residues (known as the common docking domain, CD), suggesting that electrostatic interactions are critical for the binding of MAP kinases and MKPs. Consistent with this suggestion, an altered form of ERK2 (D319N) corresponding to the Drosophila sevenmaker mutation is highly compromised in its ability to bind to, and activate, DUSP6 [26,27]. The low-molecular-weight DSPs do not have the MAP kinase docking site present in MKPs, but VHR (a low-molecular-weight DSP) has been shown to be capable of inactivating ERK in cells [28,29], suggesting that it may have a distinct mechanism of interaction.
Figure 4.
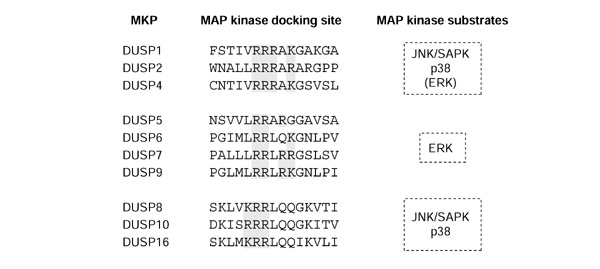
Classification of DUSPs on the basis of MAP kinase docking site. The sequence thought to be responsible for MAP kinase interaction in DUSPs is shown, as is substrate preference. This results in a DUSP subclassification similar to that obtained by analysis of gene structure (Figure 1) or amino-acid sequence similarity (Figure 2). Adapted from [14].
Further regions that are potentially important for docking to and activation by MAPKs have now been defined in certain subsets of MKPs. The FXFP motif, and variants of this sequence (present in DUSP1, DUSP4, DUSP6, DUSP7 and DUSP9) has been shown to mediate ERK binding [30,31,32]. Another such region is the D-domain (or D-box), which comprises a cluster of basic residues amino-terminal to an L/I-X-L/I motif [30,32,33]. The consensus for this site was derived from alignment of the sequence of the delta domain of c-Jun, the docking site for JNK, with similar motifs found in other transcription factors that are known targets of the MAP kinases. This domain is present in all MKPs identified to date [34] and is thought to mediate binding to both JNK and ERK MAP kinases [30,32]. Given the diversity of docking sites, the prevailing view is that the affinity of the MKPs for their substrates may be governed by the number, type and accessibility of docking sites.
PEST sequences
The subgroup III DUSPs, DUSP8 and DUSP16, have an extended carboxyl terminus containing PEST sequences (abundant in proline, glutamate, serine and threonine residues) that are frequently found in rapidly degraded proteins [35]. Removal of the PEST sequences from these proteins can result in their stabilization ([36] and our unpublished observations), consistent with a role for the domain in rapid turnover.
Regulation of MKPs
In 1998, Arkinstall's group [27] demonstrated that binding of DUSP6 to purified ERK2 enhances the catalytic activity of the phosphatase. Since then, several papers have attempted to elucidate the mechanism of catalytic activation of DUSP6 [37,38] and to define the regions of interaction between DUSP6 and ERK2. The derivation of the solution structure of the ERK2-binding (EB) domain of DUSP6 [38] has provided considerable insight into this issue (Figure 3a). The EB domain of DUSP6 contains a positively charged cluster of arginine residues that binds to a highly negatively charged region of ERK2. This binding causes a conformational change in DUSP6, affecting the association of its EB domain with its catalytic domain, which results in the re-positioning of the conserved aspartate residue (Asp262 in DUSP6). These conformational effects, in conjunction with the catalytic domain binding to phosphorylated ERK2, allosterically trigger the rearrangement of DUSP6 active-site residues, resulting in a high-activity state of the phosphatase (Figure 5). This mechanism of activation of DUSP6 by its substrate may be generally applicable to the DUSPs; recent data support a similar model for both DUSP4 and DUSP9 [27,39], and binding of DUSP1 to p38 results in the catalytic activation of the phosphatase [40].
Figure 5.
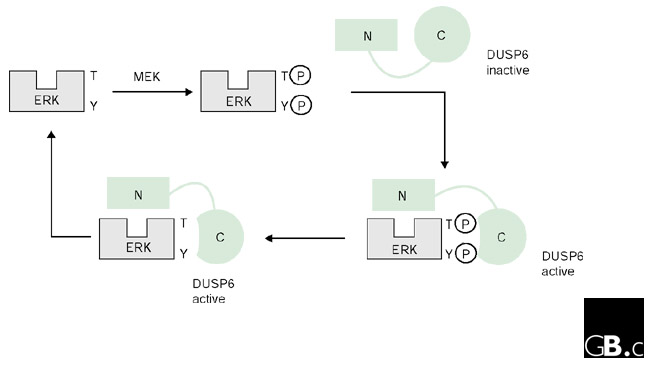
Activation of DUSP6 phosphatase by interaction with substrate ERK. This is a simplified schematic representation of ERK binding to DUSP6, resulting in catalytic activation. The carboxy-terminal catalytic domain is represented by a circle and the amino terminus containing the ERK binding (EB) domain by a rectangle. DUSP6 appears to exist in a low-activity state until binding to ERK through the EB domain results in a conformational change, triggering activation of the phosphatase. This results in ERK dephosphorylation and subsequent dissociation of the complex.
An alternative regulatory mechanism is exemplified by the binding of DUSP1 to ERK1, which results in phosphorylation of the phosphatase [41]. This phosphorylation does not directly affect the phosphatase activity of DUSP1, but instead leads to its stabilization, by reducing the rate of proteosome-mediated degradation of this rather labile protein [41].
Localization and function
Most DUSPs display wide tissue distribution but some show a rather restricted expression pattern. These include: DUSP2, which is predominantly expressed in hematopoietic tissues [42]; DUSP8, which is expressed mainly in brain, as well as in heart and lung [43,44]; DUSP9, which is detected in placenta, kidney and fetal liver [45]; and DUSP10, which appears to be restricted to liver and skeletal muscle [46]. Detailed information on expression patterns during development is not available for most DUSPs.
The DUSPs vary in their subcellular locations and can be subdivided into three major groups on the basis of location. One group consists of members expressed exclusively in the nucleus, including DUSP1, DUSP2, DUSP4 and DUSP5 [42,47,48] whereas another group includes DUSP6, DUSP7 and DUSP16, which are predominantly expressed in the cytoplasm [12,14,15,49,50,51]. Some DUSPs (DUSP8, DUSP9 and DUSP10) show both cytoplasmic and nuclear localizations [44,45,46,52]. It is of interest that DUSP9 shows punctate nuclear staining, which is reminiscent of the pattern displayed by promyelocytic leukemia (PML) protein [45]. Moreover, DUSP9 and PML co-localize in a significant proportion of cells, suggesting a potential role for DUSP9 as a regulator of kinases that phosphorylate PML. The recently identified DUSP16 has been shown to act as a shuttle protein [15]; it has both an active nuclear localization signal (NLS) and a nuclear export signal (NES), and it has been suggested to function by translocating into the nucleus, where it interacts with and dephosphorylates its substrates (JNK and/or p38) and then subsequently transports them to the cytoplasm. The sequence of the NES region of DUSP8 is very similar to that of DUSP16, and it therefore seems possible that DUSP8 also shuttles between cytoplasm and nucleus.
Despite the wealth of data supporting the role of MKPs in the regulation of MAP kinases in cell-culture systems, there is as yet no direct in vivo evidence confirming this in mammals. Disruption of the DUSP1 gene in mice causes no obvious phenotype [53]. Moreover, DUSP1-deficient embryonic fibroblasts and stem cells display no abnormalities in the regulation of MAP kinases. Genetic studies in yeasts, Drosophila and Arabidopsis have provided evidence for a role for MKPs in the regulation of MAP kinases in vivo, however. In Drosophila, mutations in puckered (encoding an MKP) cause severe developmental defects, resulting in embryonic lethality due to hyperactivation of Drosophila JNK and failure of dorsal closure [18]. Moreover, a mutant has been identified in Arabidopsis that develops normally but is hypersensitive to genotoxic stress as a result of disruption of a gene that encodes a MKP, AtMKP1. AtMKP1 is required for maintaining the MAP kinase activity at levels that contribute to genotoxic stress resistance rather than those that trigger cell death [22].
Substrate specificity
Assessment of substrate specificity for the DUSPs has proved problematic because many of the assays used do not reflect the situation in vivo. From data published so far, however, it seems that many of the DUSPs have preferred substrates. For example, after transfection into mammalian cells, DUSP6 completely inactivates ERK1 and ERK2, even at low concentrations, but not JNK or p38 kinases [49,54]; similarly, DUSP9 also displays some specificity towards ERK [45]. In contrast, DUSP8 and DUSP16 appear highly specific for the inactivation of JNK and p38 kinases but fail to inhibit ERK activation [14,15,54]; similarly, DUSP10 displays specificity for p38 and JNK kinases [46,52]. These 'specificities' should be treated with caution, as they may be specific to a certain cell type or physiological status [26,55,56]. Some of the specificity may reside in the docking sites present on the various MKPs, but it must be emphasized that access to substrate may be an equally important factor in regulating specificity in vivo.
Frontiers
MAP kinase activation has been implicated in oncogenic transformation, tumor formation and metastasis [57,58,59]. This suggests that the MKPs could have a role in tumorigenesis, perhaps as tumor suppressors. Several MKPs have been mapped to regions of the human genome altered in human cancer, but direct evidence for such a role is at present lacking. It will be interesting to see whether this family of genes plays a part in the development of cancer or other diseases.
The lack of relevant mammalian models makes it difficult to assign a physiological role to these phosphatases. Given the overlapping substrate specificity displayed by members of the various subgroups, in addition to the similar tissue distributions, it seems likely that there will be a certain degree of redundancy. This issue could be addressed by combined gene targeting of the members of each subgroup. Such an approach, in conjunction with further genetic biochemical and cell biological analyses of other model organisms should result in the elucidation of the precise biological roles of these enzymes in controlling key signaling pathways. Further insight into the regulation of these phosphatases by their substrates will be gained by determining the three-dimensional structures of representative members of each subgroup, both free and in complex with their targets.
Finally, the adoption by the community of a common nomenclature (such as the DUSP system used here and recommended by the Human Gene Nomenclature committee) will considerably facilitate progress in this important area.
Acknowledgments
Acknowledgements
We thank Den Moore for figure preparation. A.T. is a Marie Curie Fellow. We thank Cancer Research UK and Breakthrough Breast Cancer for financial support.
References
- Schaeffer HJ, Weber MJ. Mitogen-activated protein kinases: specific messages from ubiquitous messengers. Mol Cell Biol. 1999;19:2435–3444. doi: 10.1128/mcb.19.4.2435. A review discussing how the regulation and localization of the MAP kinases, in conjunction with the participation of scaffold proteins and adapters, may determine different biological outcomes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang L, Karin M. Mammalian MAP kinase signalling cascades. Nature. 2001;410:37–40. doi: 10.1038/35065000. A comprehensive review of mammalian MAP kinase signaling, focusing on the functions of the MAPK cascades. [DOI] [PubMed] [Google Scholar]
- Marshall CJ. Specificity of receptor tyrosine kinase signalling: transient versus sustained extracellular signal regulated kinase activation. Cell. 1995;80:179–185. doi: 10.1016/0092-8674(95)90401-8. An excellent review on the role of the ERK MAP kinase pathway in differentiation and cell-fate determination: Marshall proposes that the duration of ERK activation is crucial in determining the biological outcome. [DOI] [PubMed] [Google Scholar]
- Guan K, Broyles SS, Dixon JE. A Tyr/Ser protein phosphatase encoded by vaccinia virus. Nature. 1991;350:359–362. doi: 10.1038/350359a0. Cloning of VH1, the first member of the dual specificity phosphatase family. Demonstration that recombinant VH1 hydrolyzes substrates containing both phosphotyrosine and phosphoserine. [DOI] [PubMed] [Google Scholar]
- Keyse SM, Ginsburg M. Amino acid sequence similarity between CL100, a dual-specificity MAP kinase phosphatase, and cdc25. Trends Biochem Sci. 1993;18:377–378. doi: 10.1016/0968-0004(93)90092-2. Identification of two regions of amino-acid sequence similarity between the two phosphatases CL100 (DUSP1) and Cdc25. The authors propose a possible role for these regions in substrate interaction. [DOI] [PubMed] [Google Scholar]
- Tanoue T, Adachi M, Moriguchi T, Nishida E. A conserved docking motif in MAP kinases common to substrates, activators and regulators. Nat Cell Biol. 2000;2:110–116. doi: 10.1038/35000065. Identification of a docking site in ERK2, designated common docking domain, which is used in common for recognition and binding of its activators, substrates and regulators. A corresponding domain is also found in p38 and JNK/SAPK kinases. [DOI] [PubMed] [Google Scholar]
- Wishart MJ, Dixon JE. Gathering STYX: phosphatase-like form predicts functions for unique protein-interaction domains. Trends Biochem Sci. 1998;23:301–306. doi: 10.1016/s0968-0004(98)01241-9. A comprehensive review of STYX domains. The authors present examples of several phosphatase-like proteins containing STYX domains and discuss their potential roles in mediating intracellular signaling. [DOI] [PubMed] [Google Scholar]
- Noguchi T, Metz R, Chen L, Mattei M-G, Carrasco D, Bravo R. Structure, mapping, and expression of erp, a growth factor-inducible gene encoding a nontransmembrane protein tyrosine phosphatase, and effect of ERP on cell growth. Mol Cell Biol. 1993;13:5195–5205. doi: 10.1128/mcb.13.9.5195. Structure and chromosomal localization of the murine erp gene (DUSP1). Stable expression of ERP in transfected NIH 3T3 fibroblasts resulted in growth inhibition and altered cell morphology. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwak S, Hakes D, Martell K, Dixon JE. Isolation and characterization of a human dual specificity protein-tyrosine phosphatase gene. J Biol Chem. 1994;269:3596–3604. Isolation of the human CL100 (DUSP1) gene, the characterization of its promoter and determination of the intron/exon boundaries. CL100 expression in discrete neuronal populations is also presented. [PubMed] [Google Scholar]
- Yi H, Morton CC, Weremowicz S, McBride OW, Kelly K. Genomic organization and chromosomal localization of the DUSP2 gene, encoding a MAP kinase phosphatase, to human 2p11.2-q11. Genomics. 1995;28:92–96. doi: 10.1006/geno.1995.1110. Structure and chromosomal localization of the human DUSP2 gene. [DOI] [PubMed] [Google Scholar]
- Zhang T, Choy M, Jo M, Roberson MS. Structural organization of the rat mitogen-activated protein kinase phosphatase 2 gene. Gene. 2001;273:71–79. doi: 10.1016/S0378-1119(01)00574-1. Genomic organization of the rat MKP-2 (DUSP4) gene, and characterization of its 5' untranslated region. Comparison of the structure of human MKP-1 (DUSP1) and rat MKP-2 genes. [DOI] [PubMed] [Google Scholar]
- Dowd S, Sneddon AA, Keyse SM. Isolation of the human genes encoding the Pyst1 and Pyst2 phosphatases: characterization of Pyst2 as a cytosolic dual-specificity MAP kinase phosphatase and its catalytic activation by both MAP and SAP kinases. J Cell Sci. 1998;111:3389–3399. doi: 10.1242/jcs.111.22.3389. Genomic organization of the human genes encoding the Pyst1 (DUSP6) and Pyst2 (DUSP7) phosphatases. The subcellular localization of Pyst2 protein is also determined. [DOI] [PubMed] [Google Scholar]
- Nesbit MA, Hodges MD, Campbell L, de Meulemeester TM, Alders M, Rodrigues NR, Talbot K, Theodosiou AM, Mannens MA, Nakamura Y, et al. Genomic organization and chromosomal localization of a member of the MAP kinase phosphatase gene family to human chromosome 11p15.5 and a pseudogene to 10q11.2. Genomics. 1997;42:284–294. doi: 10.1006/geno.1997.4737. Structure of human DUSP8 and its localization to chromosome 11p15.5. The authors also report the identification of a pseudogene on chromosome 10q11.2. [DOI] [PubMed] [Google Scholar]
- Tanoue T, Yamamoto T, Maeda R, Nishida E. A novel MAPK phosphatase MKP-7 acts preferentially on JNK/SAPK and p38α and β MAPKs. J Biol Chem. 2001;276:26629–26639. doi: 10.1074/jbc.M101981200. Identification and characterization of human MKP-7 (DUSP16), which shows substrate preference for JNK and p38. The authors also propose a tentative classification of MKPs based on the sequence characteristics of their MAP kinase-docking sites. [DOI] [PubMed] [Google Scholar]
- Masuda K, Shima H, Watanabe M, Kikuchi K. MKP-7, a novel mitogen-activated protein kinase phosphatase, functions as a shuttle protein. J Biol Chem. 2001;276:39002–39011. doi: 10.1074/jbc.M104600200. The authors present the cloning of human and mouse MKP-7 (DUSP16). They propose that this phosphatase also functions as a shuttle protein as it has both a nuclear export and a nuclear localization signal in its sequence. [DOI] [PubMed] [Google Scholar]
- Doi K, Gartner A, Ammerer G, Errede B, Shinkawa H, Sugimoto K, Matsumoto K. MSG5, a novel protein phosphatase promotes adaptation to pheromone. EMBO J. 1994;13:61–70. doi: 10.1002/j.1460-2075.1994.tb06235.x. Isolation of the phosphatase MSG5 as a suppressor of pheromone-stimulated G1 arrest in yeast cells. Loss of Msg5 function leads to increased activation of Fus3. These findings coupled with the in vitro dephosphorylation of Fus3 by Msg5 suggested a role for Msg5 in negative feedback control of MAP kinase signaling. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morrison DK, Murakami MS, Clephon V. Protein kinases and phosphatases in the Drosophila genome. J Cell Biol. 2000;150:F57–F62. doi: 10.1083/jcb.150.2.f57. A review of the kinases and phosphatases in the Drosophila genome. Provides an excellent link to a detailed table of all the genes identified in Drosophila. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin-Blanco E, Gampel A, Ring J, Virdee K, Kirov N, Tolkovsky AM, Martinez-Arias A. puckered encodes a phosphatase that mediates a feedback loop regulating JNK activity during dorsal closure in Drosophila. Genes Dev. 1998;12:557–570. doi: 10.1101/gad.12.4.557. The identification of puckered, encoding a Drosophila MKP homolog. Mutations in this gene led to cytoskeletal defects resulting in failure of dorsal closure. Martin-Blanco et al. demonstrate that this phosphatase mediates a feedback loop regulating JNK activity during Drosophila development. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S-H, Kwon H-B, Kim Y-S, Ryu J-H, Kim K-S, Ahn Y, Lee W-J, Choi K-Y. Isolation and characterization of a Drosophila homologue of mitogen-activated protein kinase phosphatase-3 which has a high substrate specificity towards extracellular-signal-regulated kinase. Biochem J. 2002;361:143–151. doi: 10.1042/0264-6021:3610143. Characterization of Drosophila MKP-3 (DUSP6). When expressed in cultured Schneider cells, DMKP-3 inhibits DERK activity and binds with DERK via its amino-terminal domain. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plowman GD, Sudarsanam S, Bingham J, Whyte D, Hunter T. The protein kinases of Caenorhabditis elegans : a model for signal transduction in multicellular organisms. Proc Natl Acad Sci USA. 1999;96:13603–13610. doi: 10.1073/pnas.96.24.13603. A comprehensive review of the kinase and phosphatase families identified in C. elegans. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berset T, Hoier EF, Battu G, Canevascini S, Hajnal A. Notch inhibition of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval development. Science. 2001;291:1055–1058. doi: 10.1126/science.1055642. Genetic dissection of the role of LIP-1 phosphatase during cell fate determination in vulval development in the worm. [DOI] [PubMed] [Google Scholar]
- Ulm R, Revenkova E, di Sansebastiano G-P, Bechtold N, Paszkowski J. Mitogen-activated protein kinase phosphatase is required for genotoxic stress relief in Arabidopsis. Genes Dev. 2001;15:699–709. doi: 10.1101/gad.192601. The identification of an Arabidopsis mutant that is hypersensitive to genotoxic stress treatments (UV-C and methyl methanesulphonate). This sensitivity is due to disruption of the gene AtMKP1, encoding an Arabidopsis MKP homolog, which is a key regulator of MAP kinase activity in vivo. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denu JM, Dixon JE. Protein tyrosine phosphatases: mechanisms of catalysis and regulation. Curr Opin Chem Biol. 1998;2:633–641. doi: 10.1016/s1367-5931(98)80095-1. This review outlines the common catalytic mechanism employed by protein tyrosine phosphatases and ways of regulating phosphatase activity. [DOI] [PubMed] [Google Scholar]
- Stewart AE, Dowd S, Keyse SM, McDonald NQ. Crystal structure of the MAPK phosphatase Pyst1 catalytic domain and implications for regulated activation. Nat Struct Biol. 1999;6:174–181. doi: 10.1038/5861. The crystal structure of the catalytic domain of Pyst1 (DUSP6). This reveals that in the absence of substrate the active site is distorted, suggesting that it undergoes conformational change upon ERK2 binding. Using site-directed mutagenesis Stewart et al. identify an aspartate that is essential for catalysis. [DOI] [PubMed] [Google Scholar]
- Yuvaniyama J, Denu JM, Dixon JE, Saper MA. Crystal structure of the dual-specificity protein phosphatase VHR. Science. 1996;272:1328–1331. doi: 10.1126/science.272.5266.1328. The authors determine the crystal structure of the human dual specificity phosphatase VHR, and show that it forms a shallow active-site cleft that allows for the hydrolysis of phosphoserine, phosphothreonine or phosphotyrosine residues. [DOI] [PubMed] [Google Scholar]
- Chu Y, Solski PA, Khosravi-Far R, Der CJ, Kelly K. The mitogen-activated protein kinase phosphatases PAC1, MKP-1 and MKP-2 have unique substrate specificities and reduced activity in vivo toward the ERK2 sevenmaker mutation. J Biol Chem. 1996;271:6497–6501. doi: 10.1074/jbc.271.11.6497. The authors demonstrate that the phosphatases PAC-1 (DUSP2), MKP-1 (DUSP1) and MKP-2 (DUSP4) show unique substrate specificities. PAC-1 is specific for ERK and p38, MKP-2 for JNK, and ERK and MKP-1 for p38 and JNK. All three phosphatases show reduced activity toward the ERK2 sevenmaker mutant. [DOI] [PubMed] [Google Scholar]
- Camps M, Nichols A, Gillieron C, Antonsson B, Muda M, Chabert C, Boschert U, Arkinstall S. Catalytic activation of the phosphatase MKP-3 by ERK2 mitogen-activated protein kinase. Science. 1998;280:1262–1265. doi: 10.1126/science.280.5367.1262. This paper shows that MKP-3 (DUSP6) is activated by direct binding to purified ERK2 and that this activation is specific to ERK2. Similarly, MKP-4 (DUSP9) is also activated by ERK2 binding. The authors suggest that this mechanism may underlie the substrate specificity of this group of enzymes. [DOI] [PubMed] [Google Scholar]
- Todd JL, Tanner KG, Denu JM. Extracellular regulated kinases (ERK) 1 and ERK2 are authentic substrates for the dual-specificity protein-tyrosine phosphatase VHR. J Biol Chem. 1999;274:13271–13280. doi: 10.1074/jbc.274.19.13271. Demonstration that immunodepletion of endogenous VHR eliminates the dephosphorylation of cellular ERK, suggesting that VHR specifically inactivates ERK in vivo. [DOI] [PubMed] [Google Scholar]
- Alonso A, Saxena S, Williams S, Mustelin T. Inhibitory role for dual specificity phosphatase VHR in T cell antigen receptor and CD28-induced Erk and Jnk activation. J Biol Chem. 2001;276:4766–4771. doi: 10.1074/jbc.M006497200. Evidence that VHR, when expressed exogenously in Jurkat T cells, counteracts the Erk and Jnk MAP kinases and all reporter genes depending on them. The authors also show that catalytically inactive VHR behaves as a 'dominant negative' mutant. [DOI] [PubMed] [Google Scholar]
- Jacobs D, Glossip D, Xing H, Muslin AJ, Kornfeld K. Multiple docking sites on substrate proteins form a modular system that mediates recognition by ERK MAP kinase. Genes Dev. 1999;13:163–175. Suggestion that the FXFP motif is a docking site that mediates ERK binding to substrates in multiple protein families. The authors also identify another motif, designated D domain/box, that mediates binding to both ERK and JNK kinases. Their findings suggest that the overlapping substrate specificities of these kinases may result from recognition of shared or unique docking sites. [PMC free article] [PubMed] [Google Scholar]
- Zhou B, Wu L, Shen K, Zhang J, Lawrence DS, Zhang Z-Y. Multiple regions of MAP kinase phosphatase 3 are involved in its recognition and activation by ERK2. J Biol Chem. 2001;276:6506–6515. doi: 10.1074/jbc.M009753200. Using site-directed mutagenesis, three regions within the MKP-3 (DUSP6) sequence that are important for its binding to ERK2 are determined. In addition, the authors demonstrate that the FTAP motif in the carboxyl terminus of the phosphatase is absolutely essential for ERK2-induced MKP-3 activation. [DOI] [PubMed] [Google Scholar]
- Fantz DA, Jacobs D, Glossip D, Kornfeld K. Docking sites on substrate proteins direct extracellular signal-regulated kinase to phosphorylate specific residues. J Biol Chem. 2001;276:27256–27265. doi: 10.1074/jbc.M102512200. The paper focuses on the functions of the FXFP and D-domain docking sites. In addition to mediating ERK binding, these sites can direct phosphorylation on specific serine/threonine residues adjacent to proline residues. [DOI] [PubMed] [Google Scholar]
- Yang S-H, Whitmarsh AJ, Davis RJ, Sharrocks AD. Differential targeting of MAP kinases to the ETS-domain transcription factor Elk-1. EMBO J. 1998;17:1740–1749. doi: 10.1093/emboj/17.6.1740. Evidence that, in addition to the ERK MAP kinases, the JNKs are also targeted to Elk-1 by the D domain, and that this targeting is essential for phosphorylation and activation of Elk-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson TR, Biggs JR, Winbourn SE, Kraft AS. Regulation of dual-specificity phosphatases M3/6 and hVH5 by phorbol esters. J Biol Chem. 2000;275:31755–31762. doi: 10.1074/jbc.M004182200. An examination of the regulation of M3/6/hVH-5 (DUSP8) in response to PMA treatment. Activation of JNK stimulates M3/6 phosphorylation and a delta-domain like motif in M3/6, the deletion of which inhibits the phosphatase activity of the enzyme, is identified. [DOI] [PubMed] [Google Scholar]
- Rechsteiner M, Rogers SW. PEST sequences and regulation by proteolysis. Trends Biochem Sci. 1996;21:267–271. This review focuses on the experimental evidence supporting the hypothesis that PEST sequences target proteins for rapid degradation, and on the mechanisms responsible for this degradation. [PubMed] [Google Scholar]
- Matsuguchi T, Musikacharoen T, Johnson TR, Kraft AS, Yoshikai Y. A novel mitogen-activated protein kinase phosphatase is an important negative regulator of lipopolysaccharide-mediated c-Jun N-terminal kinase activation in mouse macrophage cell lines. Mol Cell Biol. 2001;21:6999–7009. doi: 10.1128/MCB.21.20.6999-7009.2001. Cloning of mouse MKP-M (mouse ortholog of DUSP16), a cytosolic MKP, which shows substrate specificity for JNK and p38. MKP-M is constitutively expressed in mouse macrophage cell lines with increased expression upon LPS stimulation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fjeld CC, Rice AE, Kim Y, Gee KR, Denu JM. Mechanistic basis for catalytic activation of mitogen-activated protein kinase phosphatase-3 by extracellular signal-regulated kinase. J Biol Chem. 2000;275:6749–6757. doi: 10.1074/jbc.275.10.6749. This paper provides biochemical evidence that ERK activates MKP-3 (DUSP6) through the stabilization of the active phosphatase conformation. [DOI] [PubMed] [Google Scholar]
- Farooq A, Chaturvedi G, Mujtaba S, Plotnikova O, Zeng L, Dhalluin C, Ashton R, Zhou M-M. Solution structure of ERK2 binding domain of MAPK phosphatase MKP-3: structural insights into MKP-3 activation by ERK2. Mol Cell. 2001;7:387–399. doi: 10.1016/s1097-2765(01)00186-1. The solution structure of the ERK2 binding domain of MKP-3 (DUSP6). The authors also use biochemical analysis and define regions within the amino terminus of MKP-3 that are used for binding ERK2 but also interact with the carboxy-terminal catalytic domain, and propose a mechanism for the catalytic activation of MKP-3. [DOI] [PubMed] [Google Scholar]
- Chen P, Hutter D, Yang X, Gorospe M, Davis RJ, Liu Y. Discordance between the binding affinity of mitogen-activated protein kinase subfamily members for MAP kinase phosphatase-2 and their ability to activate the phosphatase catalytically. J Biol Chem. 2001;276:29440–29449. doi: 10.1074/jbc.M103463200. The catalytic activity of recombinant MKP-2 (DUSP4) is enhanced by ERK1 and JNK1. The authors also show that MKP-2 binds preferentially to ERK and p38, and define the region of interaction within the amino-mutant. terminus of the phosphatase. [DOI] [PubMed] [Google Scholar]
- Hutter D, Chen P, Barnes J, Liu Y. Catalytic activation of mitogen-activated protein (MAP) kinase phosphatase-1 by binding to p38 MAP kinase: critical role of the p38 C-terminal domain in its negative regulation. Biochem J. 2000;352:155–163. doi: 10.1042/0264-6021:3520155. MKP-1 (DUSP1) binds directly to p38 MAP kinase both in vitro and in vivo and this interaction results in the catalytic activation of MKP-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brondello J-M, Pouyssegur J, McKenzie FR. Reduced MAP kinase phosphatase-1 degradation after p42/p44MAPK-dependent phosphorylation. Science. 1999;286:2514–2517. doi: 10.1126/science.286.5449.2514. MKP-1 (DUSP1) is a labile protein and is targeted for proteosomal degradation via ubiquitination but phosphorylation of MKP-1 by ERK leads to the stabilization of the phosphatase. [DOI] [PubMed] [Google Scholar]
- Rohan PJ, Davis P, Moskaluk CA, Kearns M, Krutzsch H, Siebenlist U, Kelly K. PAC-1: a mitogen-induced protein nuclear tyrosine phosphatase. Science. 1993;259:1763–1766. doi: 10.1126/science.7681221. The PAC-1 (DUSP2) phosphatase, cloned from human T cells. PAC-1 is predominantly expressed in hematopoietic tissues and is localized to the nucleus. [DOI] [PubMed] [Google Scholar]
- Martell KJ, Seasholtz AF, Kwak SP, Clemens KK, Dixon JE. hVH-5: a protein tyrosine phosphatase abundant in brain that inactivates mitogen-activated protein kinase. J Neurochem. 1995;65:1823–1833. doi: 10.1046/j.1471-4159.1995.65041823.x. Isolation and characterization of human hVH-5 (DUSP8), which is abundant in brain. The expression pattern of hVH-5 in mouse embryos reveals abundant and wide distribution in the central and peripheral nervous system. [DOI] [PubMed] [Google Scholar]
- Theodosiou AM, Rodrigues NR, Nesbit MA, Ambrose HJ, Paterson H, McLellan-Arnold E, Boyd Y, Leversha MA, Owen N, Blake DJ, et al. A member of the MAP kinase phosphatase gene family in mouse containing a complex trinucleotide repeat in the coding region. Hum Mol Genet. 1996;5:675–684. doi: 10.1093/hmg/5.5.675. This paper describes the cloning and characterization of the mouse M3/6 (DUSP8) gene, which maps to distal mouse chromosome 7 and is predominantly expressed in brain. The subcellular localization of M3/6 is cell-type-dependent. [DOI] [PubMed] [Google Scholar]
- Muda M, Boschert U, Smith A, Antonsson B, Gillieron C, Chabert C, Camps M, Martinou I, Ashworth A, Arkinstall S. Molecular cloning and functional characterization of a novel mitogen activated protein kinase phosphatase, MKP-4. J Biol Chem. 1997;272:5141–5151. doi: 10.1074/jbc.272.8.5141. Human MKP-4 (DUSP9), a phosphatase that is highly selective for ERK, maps to human Xq28 and displays distinct subcellular localization. [DOI] [PubMed] [Google Scholar]
- Theodosiou A, Smith A, Gillieron C, Arkinstall S, Ashworth A. MKP5, a new member of the MAP kinase phosphatase family, which selectively dephosphorylates stress-activated kinases. Oncogene. 1999;18:6981–6988. doi: 10.1038/sj/onc/1203185. Cloning of human MKP5 (DUSP10) and mapping to human chromosome 1q32. Demonstration that MKP5 preferentially binds to and inactivates the p38 and JNK kinases. [DOI] [PubMed] [Google Scholar]
- Guan K-L, Butch E. Isolation and characterization of a novel dual specific phosphatase, HVH2, which selectively dephosphorylates the mitogen-activated protein kinase. J Biol Chem. 1995;270:7197–7203. doi: 10.1074/jbc.270.13.7197. Isolation and characterization of human HVH2 (DUSP4), a nuclear MKP that shows substrate selectivity for ERK1 and ERK2. [DOI] [PubMed] [Google Scholar]
- Kwak SP, Dixon JE. Multiple dual specificity protein tyrosine phosphatases are expressed and regulated differentially in liver cell lines. J Biol Chem. 1995;270:1156–1160. doi: 10.1074/jbc.270.3.1156. Identification of human hVH-3 (DUSP5) and comparison of its levels of expression and tissue distribution to other known members of the family of MKPs. [DOI] [PubMed] [Google Scholar]
- Groom LA, Sneddon AA, Alessi DR, Dowd S, Keyse SM. Differential regulation of the MAP, SAP and RK/p38 kinases by Pyst1, a novel cytosolic dual-specificity phosphatase. EMBO J. 1996;15:3621–3632. Identification of human Pyst1 (DUSP6) and Pyst2 (DUSP7). Pyst 1 is the first member of the MKPs that is localized exclusively in the cytoplasm,is not encoded by an immediate early gene and shows substrate specificity for ERK. [PMC free article] [PubMed] [Google Scholar]
- Mourey RJ, Vega QC, Campbell JS, Wenderoth MP, Hauschka SD, Krebs EG, Dixon JE. A novel cytoplasmic dual specificity protein tyrosine phosphatase implicated in muscle and neuronal differentiation. J Biol Chem. 1996;271:3795–3802. doi: 10.1074/jbc.271.7.3795. The cloning and characterization of rat rVH6 (DUSP6), a cytoplasmic MKP. The authors also show that in PC12 cells the expression of rVH6 is induced during nerve growth factor-mediated differentiation, whereas in muscle cells it is highly expressed during proliferation. [DOI] [PubMed] [Google Scholar]
- Muda M, Boschert U, Dickinson R, Martinou J-C, Martinou I, Camps M, Schlegel W, Arkinstall S. MKP-3, a novel cytosolic protein-tyrosine phosphatase that exemplifies a new class of mitogen-activated protein kinase phosphatase. J Biol Chem. 1996;271:4319–4326. doi: 10.1074/jbc.271.8.4319. This paper describes the cloning and characterization of rat MKP-3 (DUSP6) and MKP-X (DUSP7), and shows that MKP-3 is a cytosolic MKP that specifically inactivates ERK. Expression studies of MKP-3 reveal that it is enriched within the CA1 layer of the hippocampus. [DOI] [PubMed] [Google Scholar]
- Tanoue T, Moriguchi T, Nishida E. Molecular cloning and characterization of a novel dual specificity phosphatase, MKP-5. J Biol Chem. 1999;274:19949–19956. doi: 10.1074/jbc.274.28.19949. Human MKP-5 (DUSP10), which shows substrate specificity for p38 and JNK, is localized in both the cytoplasm and the nucleus and its expression is induced by stress stimuli. [DOI] [PubMed] [Google Scholar]
- Dorfman K, Carrasco D, Gruda M, Ryan C, Lira SA, Bravo R. Disruption of the erp/mkp-1 gene does not affect mouse development: normal MAP kinase activity in ERP/MKP-1-deficient fibroblasts. Oncogene. 1996;13:925–931. MKP-1 (DUSP1) knockout mice show no phenotypic or histological abnormalities. [PubMed] [Google Scholar]
- Muda M, Theodosiou A, Rodrigues N, Boschert U, Camps M, Gillieron C, Davies K, Ashworth A, Arkinstall S. The dual specificity phosphatases M3/6 and MKP-3 are highly selective for inactivation of distinct mitogen-activated protein kinases. J Biol Chem. 1996;271:27205–27208. doi: 10.1074/jbc.271.44.27205. Demonstration that M3/6 (DUSP8) displays high substrate specificity for JNK and p38 whereas MKP-3 (DUSP6) is highly selective for ERK MAP kinases. [DOI] [PubMed] [Google Scholar]
- Hirsch DD, Stork PJS. Mitogen-activated protein kinase phosphatases inactivate stress-activated protein kinase pathways in vivo. J Biol Chem. 1997;272:4568–4575. doi: 10.1074/jbc.272.7.4568. Examination of the ability of MKP-1 (DUSP1) and MKP-2 (DUSP4) to dephosphorylate JNK1 and JNK2. Both phosphatases are able to block the activation of c-Jun, and in PC12 cells they are both transcriptionally induced upon nerve growth factor (NGF) stimulation. [DOI] [PubMed] [Google Scholar]
- Reffas S, Schlegel W. Compartment-specific regulation of extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK) mitogen-activated protein kinases (MAPKs) by ERK-dependent and non-ERK-dependent inductions of MAPK phosphatase (MKP)-3 and MKP-1 in differentiating P19 cells. Biochem J. 2000;352:701–708. doi: 10.1042/0264-6021:3520701. Regulation of MKP-1 (DUSP1) and MKP-3 (DUSP6) during induction of neuronal differentiation in mouse P19 cells. The authors demonstrate that the levels of the two phosphatases increase with different kinetics, in different cell compartments and through distinct pathways. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansour SJ, Matten WT, Hermann AS, Candia JM, Rong S, Fukasawa K, Vande Woude GF, Ahn NG. Transformation of mammalian cells by constitutively active MAP kinase kinase. Science. 1994;265:966–970. doi: 10.1126/science.8052857. Expression of constitutively active MAPKK mutants in cells results in activation of AP-1-regulated transcription and formation of transformed foci and tumors in nude mice. [DOI] [PubMed] [Google Scholar]
- Cowley S, Paterson H, Kemp P, Marshall CJ. Activation of MAP kinase kinase is necessary and sufficient for PC12 differentiation and for transformation of NIH 3T3 cells. Cell. 1994;77:841–852. doi: 10.1016/0092-8674(94)90133-3. Constitutively active and interfering mutants of MAPKK1 are generated and their effects in PC12 and NIH 3T3 cells are assessed. In these cell lines, activation of MAPKK is necessary and sufficient for cell differentiation and proliferation, respectively. [DOI] [PubMed] [Google Scholar]
- Webb CP, Van Aelst L, Wigler MH, Vande Woude GF. Signaling pathways in Ras mediated tumorigenicity and metastasis. Proc Natl Acad Sci USA. 1998;95:8773–8778. doi: 10.1073/pnas.95.15.8773. The tumorigenic and metastatic potential of effector domain mutants of oncogenic Ras is assessed. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keyse SM, Emslie EA. Oxidative stress and heat shock induce a human gene encoding a protein-tyrosine phosphatase. Nature. 1992;359:644–647. doi: 10.1038/359644a0. Cloning of the first member of the MKPs, human CL100 (DUSP1). They demonstrate that it has phosphatase activity and show that it is induced by oxidative stress and heat shock. [DOI] [PubMed] [Google Scholar]
- Charles CH, Abler AS, Lau LF. cDNA sequence of a growth factor-inducible immediate early gene and characterization of its encoded protein. Oncogene. 1992;7:187–190. The paper describes the isolation and characterization of the mouse 3CH134 (DUSP1) gene. [PubMed] [Google Scholar]
- Martell KJ, Kwak S, Hakes DJ, Dixon JE, Trent JM. Chromosomal localization of four human VH1-like protein-tyrosine phosphatases. Genomics. 1994;22:462–464. doi: 10.1006/geno.1994.1411. The chromosomal localization of DUSP1, DUSP2 and DUSP5 genes. [DOI] [PubMed] [Google Scholar]
- King AG, Ozanne BW, Smythe C, Ashworth A. Isolation and characterization of a uniquely regulated threonine, tyrosine phosphatase (TYP 1) which inactivates ERK2 and p54jnk. Oncogene. 1995;11:2553–2563. Isolation of TYP1 (DUSP4), a nuclear MKP that specifically inactivates ERK2 and JNK. [PubMed] [Google Scholar]
- Misra-Press A, Rim CS, Roberson MS, Stork PJS. A novel mitogen-activated protein kinase phosphatase. J Biol Chem. 1995;270:14587–14596. doi: 10.1074/jbc.270.24.14587. Isolation and characterization of rat MKP-2 (DUSP4), mapping of its expression in the rat brain and demonstration that both NGF and epidermal growth factor (EGF) induce a rapid increase in MKP-2 mRNA levels. [DOI] [PubMed] [Google Scholar]
- Smith A, Price C, Cullen M, Muda M, King A, Ozanne B, Arkinstall S, Ashworth A. Chromosomal localization of three human dual specificity phosphatase genes (DUSP4, DUSP6, and DUSP7). Genomics. 1997;42:524–527. doi: 10.1006/geno.1997.4756. Mapping the human phosphatases DUSP4, 6 and 7 to chromosomes 8p12-p11, 12q22-23 and 3p21, respectively. [DOI] [PubMed] [Google Scholar]
- Ishibashi T, Bottaro DP, Michieli P, Kelley CA, Aaronson SA. A novel dual specificity phosphatase induced by serum stimulation and heat shock. J Biol Chem. 1994;269:29897–29902. Report of the isolation and characterization of human B23 (DUSP5). Demonstration that B23 displays substrate selectivity for ERK and that it is induced by serum stimulation and heat shock. [PubMed] [Google Scholar]
- Shin D-Y, Ishibashi T, Choi TS, Chung E, Chung IY, Aaronson SA, Bottaro DP. A novel human ERK phosphatase regulates H-ras and v-raf signal transduction. Oncogene. 1997;14:2633–2639. doi: 10.1038/sj.onc.1201106. The paper describes the cloning of the human B59 (DUSP7) MKP. It also shows that cotransfection of NIH3T3 cells with B59 inhibits morphological transformation caused by H-ras and v-raf oncogenes. [DOI] [PubMed] [Google Scholar]
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. A widely used method for the alignment of multiple protein sequences. [DOI] [PMC free article] [PubMed] [Google Scholar]