

Abstract
Objective
To identify portal segmentation and a portal fissure in the caudate lobe of the human liver in relation to the hepatic venous system and the external notch at the caudal edge of the caudate lobe.
Summary Background Data
Although the anatomy of the caudate lobe has been studied, the detailed anatomy has not yet been clarified; this is necessary to develop safe procedures for caudate lobe resection.
Methods
A total of 88 formalin-fixed human livers were dissected to visualize the portal vein and hepatic vein systems of the caudate lobe in relation to the external notch.
Results
The patterns of portal branching were classified into two types. In 58 livers (67.4%), the territories of the first-order portal branches were clearly divided into two areas (the Spiegel lobe and the paracaval portion). In the remaining 28 livers (32.6%), the territories of the second-order portal branches were clearly divided into two areas. These two areas were distinctly separated by an internal plane, which was coincident with the external notch. The caudate lobe had a systematized hepatic venous system that consisted of one (87.5%) or two (11.4%) proper hepatic veins and plural accessory hepatic veins. The proper hepatic veins laid along the internal plane between these two portal areas.
Conclusions
The caudate lobe exhibited distinct portal segmentation with a portal fissure that was indicated internally by the proper hepatic vein and externally by the notch at the caudal edge of the caudate lobe.
The caudate lobe has been studied from various aspects due to its unique anatomical location in the liver and its characteristic configuration, which resembles a joining pin of a fan. 1–3 Recently, isolated resection of the caudate lobe 4–6 and caudate lobe resection combined with major hepatectomy procedures to treat hepatocellular carcinomas and hepatic hilar bile duct cancer have increased in number, 7–10 expanding our knowledge of the caudate lobe. 11–14
The caudate lobe was regarded as an independent segment and was classified by Couinaud 3 as segment 1 of the liver, which consisted of eight segments. Kumon 15 first divided the caudate lobe into three parts: the Spiegel lobe, which was equivalent to Couinaud’s segment 1 and the conventional caudate lobe, the paracaval portion, and the caudate processus. Couinaud 16 as well as Healey and Schory 1 later subdivided the caudate lobe into two subsegments, named the left (segment 1l) and right dorsal segments (segment 1r). Couinaud’s segment 1r is equal to Kumon’s paracaval portion, and Couinaud 17 called it segment IX, which encircled the side of the inferior vena cava. In Kumon’s and Couinaud’s divisions of the caudate lobe, no detailed description of the boundaries among them was provided.
The ligamentum venosum lies to the left of the caudate lobe and enters the inferior vena cava. It has been used as a conventional index of the median border between the Spiegel lobe (segment 1) and the paracaval portion (segment IX). 4,5 However, it was identified as a vestigial organ of the ductus venosus in the embryo, and it is unclear whether it is correlated with the anatomical boundary of the caudate lobe. 14,16
While performing hepatectomies, we noticed a notch at the caudal edge of the caudate lobe in approximately half of the patients. This external notch may be a vestige of the portal segmentation of the caudate lobe, as demonstrated in animal livers. The livers of animals such as rats and mice are clearly lobed, and the caudate lobe of these animals has two distinctly independent lobes (anterior and posterior) and a paracaval portion. 18 If the structure of the liver is embryologically similar between these animals and humans, the caudate lobe of the human liver may demonstrate portal segmentation with a portal fissure, and the external notch may represent the portal fissure.
In previous studies, the anatomy of the caudate lobe has been studied mainly in terms of the distribution of the portal branches, 19 although the hepatic venous system is considered an essential component of the anatomical structure. Therefore, this study was performed to clarify the interrelations among portal segmentation, the hepatic venous system, and the external notch of the caudate lobe.
MATERIALS AND METHODS
Eighty-eight livers from adult cadavers obtained from the Department of Anatomy, Gunma University School of Medicine, were dissected. These livers were formalin-fixed (10%) and preserved in the same solution at 5%. The specimens that underwent fine dissection were assessed for shape and the existence of the notch, identified as a small sagittal fissure at the caudal edge of the caudate lobe. The portal venous system was then investigated. The portal branches of the caudate lobe were defined as the branches from the fissure of the ligamentum venosum to the right extremity of the caudate processus. As indicated by Couinaud 3 and Kumon, 15 the branches that paralleled the right side of the inferior vena cava were not included.
The following features of the portal branches of the caudate lobe were observed by dissecting the caudate lobe: the number, the origin (from the main portal trunk, the bifurcation of the portal vein, the right and left portal veins), the territories (the Spiegel lobe, the paracaval portion, and the caudate processus), and the classification of size. The comparatively stronger branches of larger size (≥2 mm) were defined as the primary portal branches, and the smaller ones (<2 mm) as the accessory portal branches of the caudate lobe.
The hepatic venous system of the caudate lobe was then investigated after the posterior wall of the inferior vena cava was cut in a longitudinal direction. From the inside of the lumen, we investigated the following factors concerning the hepatic venous system of the caudate lobe: the number, the territories of drainage, and the classification of size. The large hepatic veins (≥3 mm) that had a common channel with the other small veins were categorized as the proper hepatic veins, and the small veins (<3 mm) were classified as the accessory hepatic veins. The accessory hepatic veins were further divided into two groups, middle-sized (1–3 mm) and thread-sized (<1 mm). The interrelations among the portal and hepatic venous systems and the external notch at the caudal edge of the caudate lobe were then investigated.
In this study, Kumon’s segmentation 15 was applied to the anatomical description of the caudate lobe and Couinaud’s segmentation 3 was used for the nomenclature of the other parts of the liver.
RESULTS
External Notch of the Caudate Lobe
The shape and size of the caudate lobe were variable in each case. A typical caudate lobe with the most common size and shape is shown in Figure 1, in which an external notch is clearly present at the caudal edge. The location of the notch varied according to the relative development of the paracaval portion and the Spiegel lobe. The external notch was observed in 44/82 (53.7%) livers, whereas in the remaining 38 no notch could be identified. In six livers, the notch could not be determined due to a former dissection.
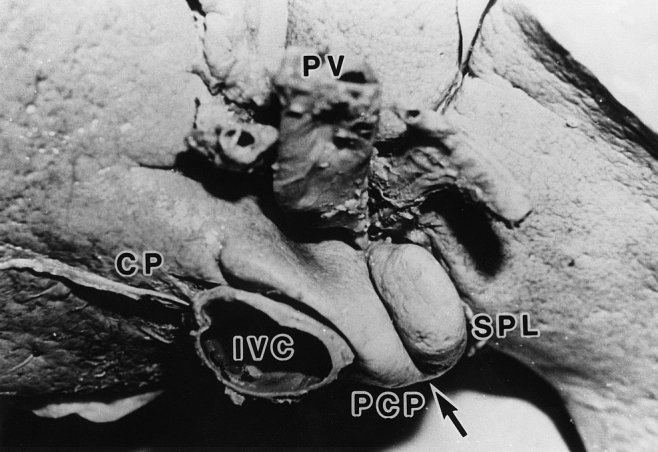
Figure 1. Outer view of the caudate lobe from the caudal side. An external notch (arrow) divides the Spiegel lobe (SPL) from the paracaval portion (PCP). CP, caudate processus; IVC, inferior vena cava; PV, portal vein.
Portal Venous System of the Caudate Lobe
Number, Origin, and Classification of the Portal Branches
The number of portal branches of the caudate lobe varied from 1 to 6, totaling 253 vessels, with an average of 2.94 vessels per caudate lobe. The number of livers that had one to six portal branches were 7 (8.1%), 28 (32.5%), 26 (30.2%), 17 (19.8%), 4 (4.7%) and 4 (4.7%), respectively. Livers with two to four portal branches were dominant.
The caudate branches ramified from the left portal vein in 123/253 (48.6%) branches, from the right portal vein in 58/253 (22.9%), from the bifurcation of the portal vein in 43/253 (17.0%), and from the main portal trunk in 29/253 (11.5%). The most common origin of the portal branches was the left portal vein (Table 1).
Table 1. RELATION BETWEEN THE ORIGIN OF PORTAL BRANCHES AND THE THREE AREAS OF THE CAUDATE LOBE
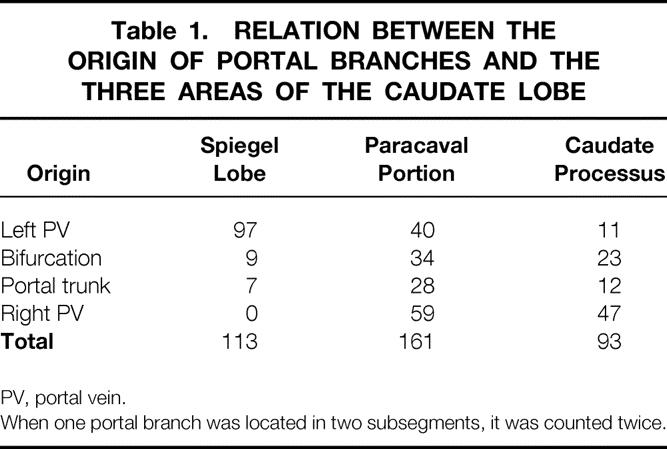
PV, portal vein.
When one portal branch was located in two subsegments, it was counted twice.
Primary portal branches were found in 137/253 (54.1%) branches, among which 81 branches (59.1%) originated from the left portal vein. The remaining smaller ones, named the accessory portal branches, were observed in 116/253 (45.9%) portal branches. The portal branches of 2 of the 88 livers could not be determined due to a former dissection.
Territory of Portal Branches and Portal Segmentation
In terms of the relations among three areas of the caudate lobe and the number of portal branches, 113/367 (30.8%) branches in the Spiegel lobe, 161/367 (43.9%) branches in the paracaval portion, and 93/367 (25.3%) branches in the caudate processus were determined (Table 1). In this calculation, when one portal branch was located in two areas, it was counted twice. Therefore, the total number of counted portal branches was 367, which was larger than the absolute number (253) of portal branches.
In terms of the origin of the portal branches, the Spiegel lobe was principally supplied by the branches originating from the left portal vein (97/113, 85.8%), whereas the paracaval portion was supplied with combinations of branches stemming from different points, especially from the trunk or the bifurcation of the portal vein. In the caudate processus, the portal branches that originated from the right portal vein were dominant (47/93, 50.5%), and most of them for the caudate processus stemmed from the branches that supplied other subsegments (Table 1).
Although many combinations of portal branches were found in the caudate lobe, the patterns of portal branching were classified into two types (Fig. 2). In one type (n = 58, 67.4%), the territories of the first-order portal branches were clearly divided into two areas, and these two areas were distinctly separated by the intersegmental plane (Fig. 2A). In the remaining 28 livers (32.6%), one of the first-order portal branches simultaneously supplied two areas as if it had no specific territory. However, the territories of the second-order portal branches were clearly divided into two areas by the intersegmental plane (Fig. 2B). These results indicated that a boundary existed between the Spiegel lobe and the paracaval portion. No clear boundary was found between the caudate processus and the paracaval portion.
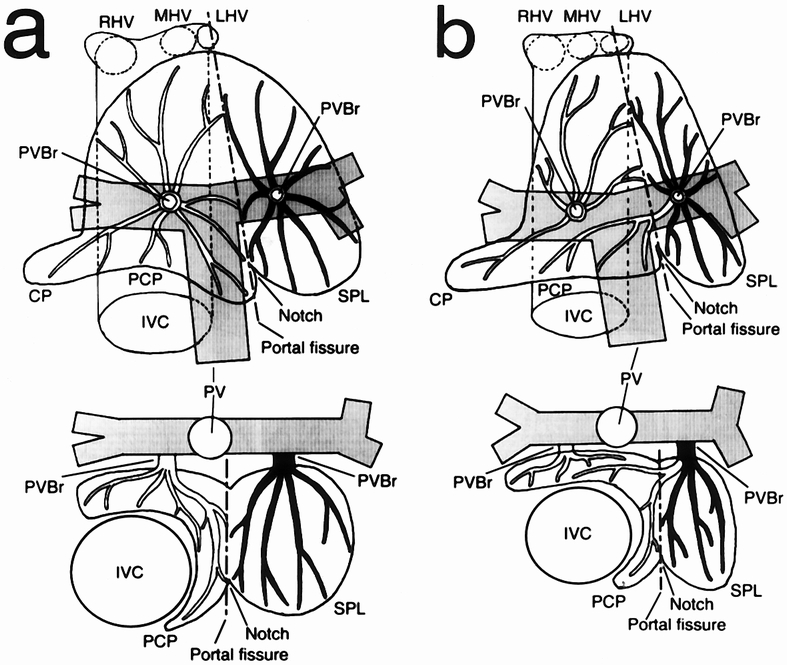
Figure 2. Two types of portal venous system in the caudate lobe. (A) The territories of the portal branches are clearly divided into two areas along the line between the external notch and the point at which the ductus venosus attaches to the inferior vena cava. (B) Although a portal branch originating from the left portal vein simultaneously supplied the two areas, the territories of the second-order portal branches were clearly divided along the line indicated by the external notch. Upper figure shows a top view; lower figure shows a cross-sectional view at the level of the porta hepatis. IVC, inferior vena cava; LHV, left hepatic vein; MHV, middle hepatic vein; PV, portal vein; PVBr, portal branch (black = portal branch of the Spiegel lobe [SPL]; white = portal branch of the paracaval portion [PCP] and the caudate processus [CP]); RHV, right hepatic vein. A dotted line indicates the portal fissure.
Hepatic Venous System of the Caudate Lobe
Fundamental Structure
The hepatic venous system of the caudate lobe consisted of one (77/88, 87.5%) or two (10/88, 11.4%) proper hepatic veins and the plural small accessory hepatic veins (Fig. 3). Most proper hepatic veins entered the inferior vena cava from the caudate lobe via a large common channel at the left anterior side of the inferior vena cava. It initially ramified small branches into the paracaval portion from its common channel and then went through the intersegmental plane between the Spiegel lobe and the paracaval portion and finally reached the external notch, dividing small branches into these two areas. The number and size of the accessory hepatic veins varied in each case, and these hepatic veins were distributed in all areas of the caudate lobe.
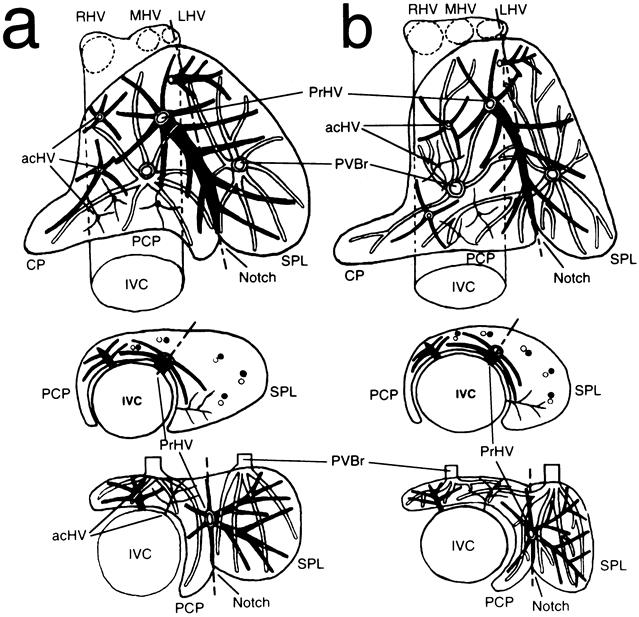
Figure 3. Hepatic venous system (black) of the caudate lobe with the correlation between the portal venous system (white) and the external notch. Hepatic venous system consists of proper (PrHV) and plural accessory (acHV) hepatic veins. The PrHV lies along the plate between two portal segments that depend on the territories of the first-order branches (A) and those of the second-order branches (B). The portal fissure is indicated internally by the PrHV and externally by the notch at the lower edge of the caudate lobe. Upper figure shows a top view; middle figure shows a cross-sectional view at the level of the root of the PrHV; lower figure shows a cross-sectional view at the level of the porta hepatis. CP, caudate processus; IVC, inferior vena cava; LHV, left hepatic vein; MHV, middle hepatic vein; PCP, paracaval portion; PVBr, portal branch; RHV, right hepatic vein; SPL, Spiegel lobe. A dotted line indicates the portal fissure.
Area of Drainage
The drainage of the Spiegel lobe depended mainly on the proper hepatic vein and partly on the middle-sized accessory hepatic veins. The combination of the proper and accessory hepatic veins to drain the Spiegel lobe is shown in Table 2. In one liver, no proper hepatic vein was observed but five middle-sized accessory hepatic veins drained the Spiegel lobe.
Table 2. DRAINAGE OF SPIEGEL LOBE WITH COMBINATIONS OF PROPER AND ACCESSORY HEPATIC VEINS
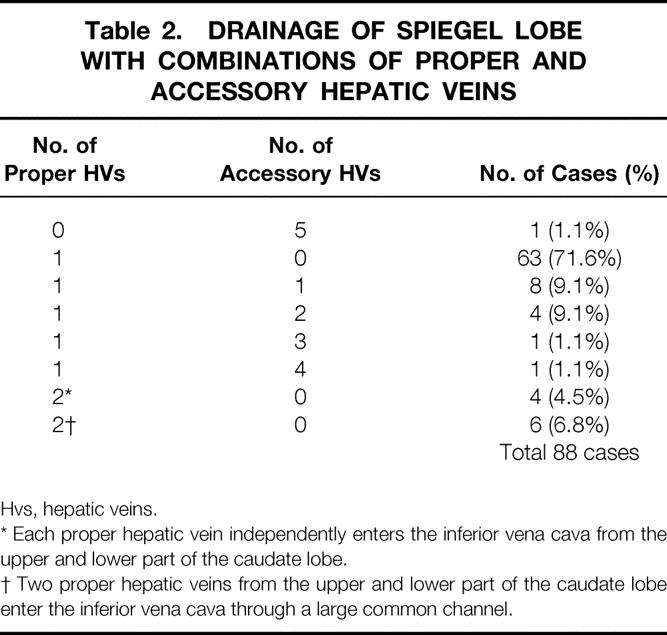
Hvs, hepatic veins.
* Each proper hepatic vein independently enters the inferior vena cava from the upper and lower part of the caudate lobe.
† Two proper hepatic veins from the upper and lower part of the caudate lobe enter the inferior vena cava through a large common channel.
The hepatic venous system of the paracaval portion was supported by three groups of hepatic veins. The first group consisted of small branches ramified radially from the common channel of the proper hepatic vein. These veins covered the drainage of a considerable area of the paracaval portion. The second group was composed of middle-sized accessory hepatic veins, which mainly drained the right part of the paracaval portion. One to four middle-sized accessory hepatic veins were observed in 38.4% of the livers in the paracaval portion. The third group included thread-sized accessory hepatic veins. The caudal portion of the paracaval portion was partly drained by these veins.
The drainage of the caudate processus was complex. It was occasionally drained by several thread-sized accessory hepatic veins or by a branch from the common channel of the proper hepatic vein. Moreover, the thread-sized accessory hepatic veins of the paracaval portion sometimes aided drainage of the caudate processus.
Correlation Among Portal Segmentation, Proper Hepatic Vein, and External Notch of Caudate Lobe
The trunk of the proper hepatic vein was set deep in the caudate lobe, directed anterocaudally from the inferior vena cava, and its ramifying pattern exhibited marked regularity. When the caudate lobe was divided into two areas along the line of the external notch, the trunk of the proper hepatic vein was found to lie on the intraparenchymal boundary between these two areas, which were coincident with the Spiegel lobe and the paracaval portion, respectively. This observation indicated that the caudate lobe had distinct portal segmentation with a portal fissure that was located internally by the proper hepatic vein and externally by the notch at the caudal edge (Fig. 3).
DISCUSSION
In the embryonic stage, the portal and hepatic venous systems of the liver develop first, followed by the hepatic arterial and biliary systems. Investigations into the structures of the portal and the hepatic veins have been conducted to clarify the true anatomical structures of the liver. 20,21 However, it was considered difficult to identify a pattern of portal branches in the caudate lobe because of the large number of individual variations in these vessels. 8,22 Nevertheless, a basic feature of the portal vein system of the caudate lobe was presented by Kumon 15 based on a cast study of the human liver. His study first showed the existence of portal segmentation in the caudate lobe, especially the existence of the special territory, the paracaval portion encircling the inferior vena cava in a semicircumferential manner, but he did not refer to the internal boundary (portal fissure) separating the Spiegel lobe from the paracaval portion. Couinaud 16 reported that in 34 of 96 casts, the hepatic vein lay in the fissural plane, which was variably oriented from a frontal to sagittal position between the afferent left and right portal branches. These observations suggest the existence of portal segmentation and a portal fissure of the caudate lobe. However, in the remaining 62 cases he did not observe distinct portal segmentation or a portal fissure in the caudate lobe because the hepatic vein was intermingled with the afferent portal branches, so that he did not amplify it on the boundary between segment 1 (Spiegel lobe) and segment IX (paracaval portion).
In our study, two types of portal segmentation were observed. In 67.4% of the livers, the distribution of the first-order portal branches was clearly separated along the fissural plane between the Spiegel lobe and the paracaval portion. In the other 32.6% of the livers, portal segmentation could not be determined according to the distribution of the first-order portal branches because these branches simultaneously supplied two areas. Our former cases may correspond to Couinaud’s casts that had hepatic veins in a fissural plane, but the latter cases may be related to Couinaud’s 62 casts in which the hepatic vein was intermingled with the portal branches. However, in the latter cases, although portal segmentation was intermingled with the hepatic vein, a fissural plane was clearly observed when portal segmentation was defined according to the distribution of the second-order portal branches. The results proved that the caudate lobe has distinct portal segmentation and a portal fissure that is located internally by the proper hepatic vein.
The clear boundary that separated the caudate processus from the paracaval portion could not be determined by dissecting the portal branches and the hepatic veins, although the caudate processus was observed as a small pyramidal shape at the right caudal portion of the caudate lobe, and all the studies described above accepted its existence. This result may be attributed to an insufficient volume to reveal exclusive portal branches of the caudate processus.
It is difficult to determine the boundary between the paracaval portion and the right liver when resection of the caudate lobe is performed. 23 Takayama et al 6 showed that the margins of the caudate processus and the right border of the paracaval portion could be recognized by indentations that corresponded to the right posterior branches of the Glisson sheath using a counterstaining technique. However, no other superficial structures that indicated the border between the paracaval portion and the right liver could be found by dissecting our formalin-fixed livers.
Studies have been performed on the hepatic venous system of the caudate lobe. Our study was the first to indicate that the hepatic veins of the caudate lobe had clear systematized structures in relation to portal segmentation. Nakamura and Tsuzuki 24 classified the hepatic veins of the caudate lobe into four types according to number and location. Foucou et al 2 reported two groups of hepatic veins (the large dorsal vein and the small dorsal vein) in the caudate lobe, whereas Minh et al 13 described three kinds of hepatic veins (the middle hepatic vein, the inferior hepatic vein, and the superior hepatic vein). The hepatic veins identified by Nakamura and Tsuzuki, the large dorsal vein by Foucou et al, and the middle hepatic vein by Minh et al corresponded to our proper hepatic vein. In addition, the small hepatic veins detected by Foucou et al and the superior or inferior hepatic veins by Minh et al corresponded to our accessory hepatic veins of the caudate lobe. These studies recognized that the caudate lobe had both large (proper) and small (accessory) hepatic veins. However, no study except that by Couinaud 16 considered these hepatic veins in relation to the portal segmentation of the caudate lobe. In all of our livers, the proper hepatic veins lay along the fissural plane between the Spiegel lobe and the paracaval portion as if the middle hepatic vein lay along the main portal fissure between the right and left liver. Our results showed that the proper hepatic vein is internally located by the portal fissure between the two portal areas, the Spiegel lobe and the paracaval portion.
Few studies have examined the external index that separates the Spiegel lobe from the paracaval portion in the caudate lobe. In more than half of the livers, the caudate lobe showed an external notch at the caudal edge. This external notch has not been closely analyzed because most anatomical studies are performed by cast studies. However, in cast studies the external notch disappeared with hepatic parenchyma as a result of the caustic. It may be a vestige of the portal segmentation of the caudate lobe, as has been demonstrated in animal livers. Our results confirmed that the external notch can be used as an index to separate the Spiegel lobe from the paracaval portion.
Our results provide new information regarding the anatomy of the caudate lobe and are important for safe procedures to manage hepatic malignant neoplasms originating in the caudate lobe and extended left hepatectomy with the caudate lobe in donors for living related liver transplantation. 25
Acknowledgments
The authors thank Prof. Harunori Ishikawa, technician Fujio Takahashi, and Mitsuaki Shikata, Second Department of Anatomy, Gunma University School of Medicine, for their suggestions, advice, and cooperation.
Footnotes
Correspondence: Kimitaka Kogure, MD, PhD, First Department of Surgery, Gunma University School of Medicine, 3-39-15 Showamachi, Maebashi 371-8511, Japan.
Supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science and Culture of Japan (08557076).
Accepted for publication March 12, 1999.
References
- 1.Healey JE, Schory PC. Anatomy of the biliary ducts within the human liver. Arch Surg 1953; 66:599–616. [DOI] [PubMed] [Google Scholar]
- 2.Foucou B, Said HO, Joyeux A, et al. Le segment I du foie ou lobe de Spigel: Etude anatomique et interet chirurgical. J Chir (Paris) 1983; 120:179–186. [PubMed] [Google Scholar]
- 3.Couinaud C. Portal segmentation. In: Couinaud C, ed. Controlled hepatectomies and exposure of the intrahepatic bile ducts. Paris: C. Couinaud; 1981: 9–27.
- 4.Yanaga K, Matsumata T, Hayashi H, et al. Isolated hepatic caudate lobectomy. Surgery 1993; 115:757–761. [PubMed] [Google Scholar]
- 5.Kosuge T, Yamamoto J, Takayama T, et al. An isolated, complete resection of the caudate lobe, including the paracaval portion, for hepatocellular carcinoma. Arch Surg 1994; 129:280–284. [DOI] [PubMed] [Google Scholar]
- 6.Takayama T, Tanaka T, Higaki T, et al. High dorsal resection of the liver. J Am Coll Surg 1994; 179:72–75. [PubMed] [Google Scholar]
- 7.Nimura Y, Hayakawa N, Kamiya J, et al. Hepatic segmentectomy with caudate lobe resection for bile duct carcinoma of the hepatic hilus. World J Surg 1990; 14:535–544. [DOI] [PubMed] [Google Scholar]
- 8.Elias D, Lasser PH, Desruennes E, et al. Surgical approach to segment I for malignant tumors of the liver. Surg Gynecol Obstet 1992; 175:17–24. [PubMed] [Google Scholar]
- 9.Bartlett D, Fong Y, Blumgart LH. Complete resection of the caudate lobe of the liver: technique and results. Br J Surg 1996; 83:1076–1081. [DOI] [PubMed] [Google Scholar]
- 10.Takayama T, Makuuchi M, Kosuge T, et al. A hepatoblastoma originating in the caudate lobe radically resected with the inferior vena cava. Surgery 1991; 109:208–213. [PubMed] [Google Scholar]
- 11.Mizumoto R, Suzuki H. Surgical anatomy of the hepatic hilum with special reference to the caudate lobe. World J Surg 1988; 12:2–10. [DOI] [PubMed] [Google Scholar]
- 12.Heloury Y, Leborgne J, Rogez JM, et al. Review of clinical anatomy: the caudate lobe of the liver. Surg Radiol Anat 1988; 10:83–91. [DOI] [PubMed] [Google Scholar]
- 13.Minh TV, Galizia G, Lieto E. L’anatomie du lobe de Spigel du foie. Aspects nouveaux et applications chirurgicales. Ann Chir (France) 1992; 46:309–319. [PubMed] [Google Scholar]
- 14.Hasegawa H, De Cervens T, Yamasaki S, et al. Strategie chirurgicale pour les carcinomes hepatocellulaires du lobe caude. J Chir (Paris) 1991; 128:533–540. [PubMed] [Google Scholar]
- 15.Kumon M. Anatomy of the caudate lobe with special reference to portal vein and bile duct. Acta Hepatol Jpn 1985; 26:1193–1199. [Google Scholar]
- 16.Couinaud C. Posterior or dorsal liver. In: Couinaud C, ed. Surgical anatomy of the liver revisited. Paris: C. Couinaud; 1989: 123–134.
- 17.Couinaud C. The paracaval segments of the liver. J Hepato Bil Panc Surg 1994; 2:145–151. [Google Scholar]
- 18.Kogure K, Ishizaki M, Nemoto M, et al. The naming of the hepatic lobes of the rats. Acta Hepatol Jpn 1986; 27:1155–1160. [Google Scholar]
- 19.Franceschini LJ, Ortale JR. Ramification of the portal vein at the porta hepatis in humans. Surg Radiol Anat 1995; 17:35–39. [DOI] [PubMed] [Google Scholar]
- 20.Lassau JP. L’organogenese du foie humain. Contribution a l’etude de l’angioarchitecture hepatique. Quoted in: Couinaud C, ed. Surgical anatomy of the liver revisited. Paris: C. Couinaud; 1989:17.
- 21.Nawar NNY, Mikhail Y, Bahi el Din IA. Fetal hepatic vessels and subsegmentation with evidence of further subdivision. Acta Anat 1980; 108:389–393. [DOI] [PubMed] [Google Scholar]
- 22.Laux G, Rapp PE. Le dispositif veineux du lobe de Spigel. CR Assoc Anat Bordeaux 1953; 78:264–271. [Google Scholar]
- 23.Yamane T, Mori K, Sakamoto K, et al. Intrahepatic ramification of the portal vein in the right and caudate lobes of the liver. Acta Anat 1988; 133:162–172. [DOI] [PubMed] [Google Scholar]
- 24.Nakamura S, Tsuzuki T. Surgical anatomy of the hepatic veins and the inferior vena cava. Surg Gynecol Obstet 1981; 152:43–51. [PubMed] [Google Scholar]
- 25.Miyagawa S, Hashikura Y, Miwa S, et al. Concomitant caudate lobe resection as an option for donor hepatectomy in adult living related liver transplantation. Transplantation 1998; 66:661–663. [DOI] [PubMed] [Google Scholar]