

Abstract
Human fibroblasts immortalized by Simian Virus 40 (SV40) are widely employed for cell and molecular biology model of study. Indeed, SV40 transmission to humans was believed to occur only under exceptional situations. The oncogenic potential of SV40 in laboratory animals is well established, whereas its involvement in human carcinogenesis is still a matter of active investigations. A recent report links SV40 exposure with the development of a brain tumor in a laboratory researcher. In previous studies, episomal viral DNA was detected in SV40 stably transformed and immortalized fibroblast cell lines. In this study, we report molecular and biological characterizations of SV40 DNA in human fibroblast cells. Our results indicate that SV40 is able to establish a persistent infection in long-term immortalized human fibroblasts, resulting in the production of an infectious viral progeny, which is able to infect both monkey and human cells. These data indicate that SV40-immortalized human fibroblasts may represent a source of SV40 infection. To avoid the SV40 infection, careful attention should be given by operators to this SV40-cell model of study.
INTRODUCTION
In a recent report (1), the authors proposed that direct exposure to Simian Virus 40 (SV40) lead to brain tumor development in a laboratory researcher. SV40 is a natural infectious agent for monkeys, whereas more recent investigations indicate that this viral agent is also a human virus (2). It is well known that SV40 was administered to humans through contaminated vaccines, mainly anti-polio vaccines between 1955 and 1963 (3,4). The possibility of a human-to-human transmission of SV40 was taken into consideration only recently (5). Indeed, SV40 DNA sequences were detected in normal and neoplastic tissues of persons too young (1 to 30 y) or too old (60 to 85 y) to have been vaccinated with SV40-contaminated polio vaccines (2). This finding may also explain the lack of difference in cancer incidence between individuals vaccinated with SV40-contaminated and SV40-free polio vaccines (6). SV40 sequences were detected in blood and sperm specimens of normal individuals and blood samples of patients (reviewed in [2]) while SV40 virions were found in urine samples (7), indicating that blood, sperm, and urine may represent routes/vehicles of transmission of SV40 horizontal infection in humans. Support to the diffusion of SV40 in the human population is provided by the presence of SV40 sequences in human brain tumors, other neoplasms, and normal tissues of children and adults (reviewed in 2;8–11); specific SV40-neutralizing antibodies in human sera (12,13); and SV40 large T antigen (Tag) antibodies in sera of mesothelioma patients (14). A scientific panel recently established the importance of assessing the ways of contagion and the mechanisms of SV40 transmission in humans (5).
How SV40 may establish a persistent infection in human cells is poorly understood. It has been reported that some human cells such as human spongioblasts, fetal neural cells, and tumor cell lines are lytically infected by SV40 (15,16). On the other hand, human mesothelial cells do not support the lytic infection efficiently, but rather they are transformed at high rate by SV40 and release SV40 virions soon after infection (17,18). A different behavior was described for SV40-infected human fibroblast cell lines. Indeed, they (1) produce a limited SV40 viral progeny, (2) are transformed, and (3) may become immortalized but at a low rate (19). Immortalized human fibroblasts have been shown to contain SV40 DNA in the integrated form and not to produce a wild-type SV40 progeny (20–22). However, on this experimental model, few studies reported the persistence of SV40 mutants, lacking the expression of the small t antigen (tag) and with a deleted form of the large T antigen (Tag), able to complete a replicative cycle (21,22). Recently, it has been demonstrated that human SV40-immortalized fibroblasts might contain different-sized SV40 genomes, including complete viral genomes (23). SV40-immortalized human fibroblasts have been widely employed as cell and molecular biology model to study DNA replication and repair, cell cycle, immortalization and transformation (8). SV40-immortalized fibroblast cell lines were therefore broadly disseminated in many research laboratories.
Following the report by Arrington and colleagues (1) describing the SV40 detection in a brain tumor of a scientist who worked with a SV40-immortalized fibroblast cell line, we investigated long-term SV40-immortalized human fibroblasts. We show that these cells, which are routinely used in many laboratories since the 1980s, may represent a source of SV40 infection. In a previous study, we reported the presence of episomal SV40 DNA in 3 of 9 SV40-immortalized human fibroblast cell lines, showing that these cells release an infectious viral progeny (24). Here, we describe the molecular characterization of the viral DNA persisting as an episome in these human cells. In addition, we further assess that even though the MRC5-SV2 fibroblasts have been cultured for over 2 decades as immortal cell lines, they support a complete SV40 replicative cycle and release an infectious wild-type viral progeny.
MATERIALS AND METHODS
Cell Lines and Cell Culture
MRC5-SV2 cells are derived from fetal lung fibroblast strain MRC-5 by infection with SV40 strain VA45-54-2 (20,24). CV-1 cells are monkey kidney cells fully permissive to SV40 infection. Cell cultures were maintained in Dulbecco’s modified Eagle’s medium–F12 (DMEM) supplemented with 10% fetal calf serum.
For infections, viral inoculum was diluted in sterile phosphate-buffered saline (PBS). Incubations with mild shaking were carried out for 2 h at room temperature, in 6 wells plates, in a final volume of 0.5 mL. The viral inoculum was then removed, and cells, after rinsing them with phosphate-buffered saline, were cultured in Dulbecco’s modified Eagle’s medium–F12 supplemented with 1% fetal calf serum.
Viral DNA and RNA Extractions
Viral DNA was extracted from SV40-infected cell culture medium. One hundred microliters of infected cell culture medium was incubated 1 h at 37 °C with 5 μg DNAse I and 10 μg of RNAse A to remove contaminant cellular nucleic acids. Sample proteins were lysed by adding 25 μL of 5% sodium dodecyl sulfate (SDS), 50 mM EDTA, pH 7.5, at room temperature for 10 min. Viral DNA was extracted twice with phenol/chloroform mixture and then ethanol precipitated. Pellet DNA was resuspended in 20 μL of Tris 10mM, EDTA 1mM.
Total cytoplasmic RNA was extracted using the commercial kit RNeasy mini (Qiagen, Milano, Italy) following the manufacturer’s instruction.
Polymerase Chain Reaction (PCR) and Reverse-transcriptase Polymerase Chain Reaction (RT-PCR) analysis
PCR amplifications were carried out with 30 ng viral DNA or 100 ng cellular genomic DNA with SV40 specific primer sets (25,26).
For reverse transcription (RT), 5 μg RNA was resuspended in 100 mL of a buffer containing 40 mM Tris-HCl, pH 7.5, 10 mM NaCl, 6 mM MgCl2. Contaminant DNA was removed by 2 repeated treatments with RNase-free DNase (50 U) (Roche Diagnostic, Monza, Italy) at 37 °C for 20 min, followed by phenol extraction and ethanol precipitation. The RT reaction was carried out with the SuperScrip III Reverse Transcriptase from Invitrogen (Invitrogen, Milano, Italy) as indicated by the supplier. The cDNA obtained was then amplified by PCR with primers specific for the VP1 and the Tag sequences (26).
Western Blot Analysis
Proteins from 105 normal or SV40-infected cells were separated by 10% polyacrylamide gel electrophoresis (PAGE) and transferred to a nitrocellulose filter. Five micrograms of 45 kDa albumin was used as molecular weight marker. SV40-VP1 was detected using a goat anti-SV40 serum and anti-goat immunoglobulin G coupled to peroxidase. The membrane was incubated for 1 h with primary antibody, diluted 1:1000 in blocking buffer, and again for 1 h with secondary antibody used at dilution of 1:5000 in blocking buffer. Immunocomplex was detected by the Enhanced Chemiluminescence (ECL) protocol (Amersham, Milano, Italy) following the manufacturer’s instructions.
Antibodies and Immunofluorescence Labeling
Cells were grown in 12 mm coverslip glass. CV-1 cells were infected as described above and stained for immunofluorescence 24 h post infection (p.i.) The preparation of mouse pAb 101 anti-Tag antibody (DBA, Milano, Italy), fluorescein-conjugated anti-mouse, goat anti-SV40 antiserum, and fluorescein-conjugated anti-goat, as well as the condition for immunofluorescence microscopy, have been previously described (27,28). Cells were counterstained with 0.5 μg/mL of 4′, 6-diamino-2-phenylindole (Sigma, Milano, Italy).
RESULTS
Molecular Characterization of SV40 Episome DNA
A detailed molecular characterization was performed on MRC5-SV2 cell line (20,24) for which the sequence of the transforming SV40 strain (VA45-54-2) is available (GenBank nr AF156105).
SV40 DNA isolated from the culture medium of immortalized fibroblasts, hereafter indicated as 45-54-2MRC5, was fully characterized by PCR and DNA sequencing as previously described (25)(24) (data not shown). These analyses were carried out to verify the viral DNA and to assess that laboratory contaminations with other SV40 strains did not occur. The amplification pattern of different SV40 DNA regions present in immortalized fibroblasts is indistinguishable from that of the input SV40 strain, indicating that no deletions or rearrangements occurred during the long-term cultures. To assess the specificity of these data, PCR products of marker DNA regions (Figure 1A), which are known to differ in distinct SV40 strains were sequenced. DNA sequence identity detected in the recovered viral DNA and the parental virus strain further confirmed the lack of SV40 DNA rearrangements, mutations, or deletions. Altogether these experiments established that the episomal viral DNA present in the immortalized cell line MRC5-SV2 belongs to the SV40 VA45-54-2 strain used as input virus.
Figure 1.
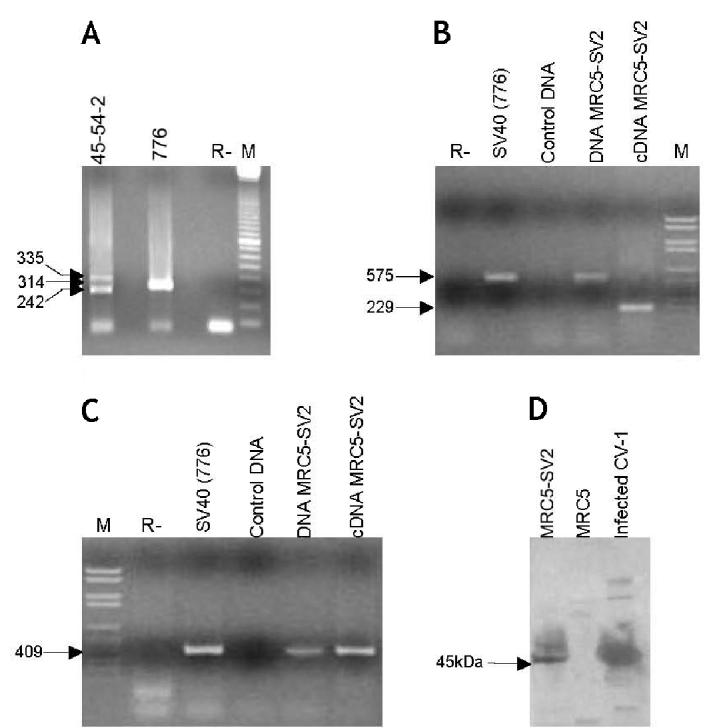
Complete molecular characterization of the SV40 particles present in the MRC5-SV2 immortalized human fibroblast cell line, from the DNA to the protein level. A: PCR amplification with RA1 and RA2 primers (Lednicky and Butel 2001[40]) of the SV40 regulatory region. 45-54-2 viral DNA was extracted from the culture medium of MRC5-SV2 cells as described in Materials and Methods. The DNA of SV40 strain 776, used as control, shows a single PCR product of 314 bp (arrow), as expected. In SV40 strain 45-54-2 the RA1 primer anneals on 2 repeated sequences, with the consequent amplification of 2 fragments, one of 335 bp and the other of 242 bp (arrows). R–, control of the PCR without DNA template; M, 100 base-pair ladder (Amersham, Milano, Italy) as marker. B, C: RT-PCR experiments on cDNA from MRC5-SV2 cells. A single reverse transcription reaction was performed on MRC5-SV2 total cytoplasmic RNA extracted from 106 cells using the Rneasy kit (Qiagen, Milano, Italy). Genomic DNA extracted from MRC5-SV2 cells was amplified for comparison. SV40 DNA, strain 776, was employed as positive control, whereas normal human DNA was used as negative control. R–, control of the PCR without DNA template; M, molecular weight marker VI (Roche Diagnostics, Monza, MI, Italy). B: Amplification carried out using a primer pair located upstream and downstream the Tag intron, which generates an amplimer of 229 bp on the cDNA and one of 575 on the genomic DNA (arrows) (26). C: Amplification of 409 bp within the VP1 sequence obtained with the primer pair VPA-VPB (Martini et al., 2002 (26)). (D) Western blot analysis carried out on total protein extracts from 105 cells separated on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis. A goat anti-SV40 serum was used for primary incubation, followed by an anti-goat IgG coupled to peroxidase for detection. Protein extract form CV-1 cells harvested 24 h after SV40 infection, was used as positive control, whereas protein extract from normal human MRC5 fibroblasts was the negative control.
SV40 DNA from Immortalized Human Fibroblasts Is Biologically Active
Once we assessed the presence of wild-type episomal SV40 DNA in human immortalized fibroblasts, we then verified whether the virus was able to multiply in human cells. Tag and VP1 transcripts, representative of early and late viral genes, were tested by RT-PCR (Figures 1B and 1C). A single RT reaction was performed on total cytoplasmic RNA, DNA-free, from MRC5-SV2 cells. Tag and VP1 specific transcripts were investigated on the same cDNA sample. To confirm the absence of contaminant DNA, a primer pair located upstream and downstream the Tag intron, was used for the PCR amplification of the viral oncoprotein coding sequences, as previously described (26). Indeed, on the c-DNA sample, these primers generated a fragment of only 229 bp, while the viral genomic amplification product of 575 bp was absent (see Figure 1B). These data confirmed that RNA contamination with viral and cellular genomic DNA did not occur.
Expression of the structural protein VP1 was tested on the same cDNA using the primer pair, which amplifies 409 bp of the VP1 coding sequence on both genomic viral DNA and cDNA (25). Because the genomic DNA was not present in the cDNA sample, we may infer the amplification product obtained for the VP1 sequence reflects the presence of mRNA specific of this late viral gene (see Figure 1C).
To verify whether the late mRNAs were also translated, the VP1 protein was analyzed by Western blotting. Using a goat anti-SV40 serum on total protein extracts of MRC5-SV2 and CV-1 cells 24 h post-infection, the 45 kDa band corresponding to the VP1 monomer was identified both in the infected cells and immortalized human fibroblasts, while it was absent in MRC5 normal fibroblasts used as control (see Figure 1D). These results support the evidence that a viral complete replicative cycle occurs in the immortalized fibroblasts, suggesting the release of a SV40 infectious viral progeny in the culture medium.
SV40 Persistent Infection in Immortalized Human Fibroblasts
We previously demonstrated, by transfection and co-culturing experiments with permissive CV-1 cells, that 45-54-2MRC5 was biologically active (24). Herein, the viral titer was determined by endpoint dilution assay in CV-1 permissive cells. Culture medium of human immortalized fibroblasts or CV-1 cells infected with 45-54-2MRC5 were employed. To avoid any possible interference by cellular debris, the experiments were carried out after filtering the culture medium through 45 nm filters. The viral progeny released by the human cells (45-54-2MRC5) gave a titer of 103 plaque-forming units (pfu)/mL, whereas a higher titer, 106 pfu/mL, was obtained from CV-1 cells infected by the same SV40 strain (45-54-2CV-1) (Table 1). It is worth noting that the latency period, before the appearance of the cell lysis, was significantly higher in SV40 infections performed at the same multiplicity of infection (MOI) with 45-54-2MRC5 compared with 45-54-2CV-1. For example, at MOI of 10, SV40 45-54-2MRC5 induces cell lysis in additional 10 d compared with SV40 45-54-2CV-1.The same infection assay was employed in human normal fibroblasts, obtaining a perfect match with the results observed in CV-1 cells. Once assessed that SV40-immortalized fibroblasts release viral progenies, which were infectious for both monkey and human cells, we investigated how MRC5-SV2 cells can be maintained in culture as immortal cell line without lysis. One possible explanation is that the virus lytic cycle takes place only in a fraction of the human cells. To verify this hypothesis, we tested by immunostaining the presence of Tag and VP1 in MRC5-SV2 cells (Figure 2). In this instance, every cell was Tag-positive, but 82.6% of the cells displayed a diffuse cytoplasmic staining (see Figure 2B) instead of the well-established nuclear staining present in permissive cells (see Figure 2A). VP1 protein was detectable only in 77% of human cells, being the specific staining mainly diffuse in the cytoplasm (see Figure 2D). In permissive monkey cells the VP1 was detected, as expected, at nuclear and perinuclear levels (see Figure 2C).
Table 1.
Viral titers showed by SV40 VA45-54-2 strain released by immortalized human fibroblast (45-54-2MRC5) and infected CV-1 cells (45-54-2CV-1)
| Viral stock | Titera |
|---|---|
| Crudeb 45-54-2MRC5 | 103 pfu/mL |
| Filteredc 45-54-2MRC5 | 103 pfu/mL |
| Crudeb 45-54-2CV-1 | 106 pfu/mL |
Viral titers were determined in CV-1 permissive cell monolayers (105 cells/monolayer) by the endpoint dilution technique. Cells were observed for lysis to occur from day 4 p.i. to day 24 p.i. Matching results were obtained infecting normal human fibro-blasts with the same viral stocks employed in CV-1 cells.
Crude indicates the medium of infected cells without any treatment.
Filtered indicates the medium of infected cells processed though 45-nm filters.
Figure 2.
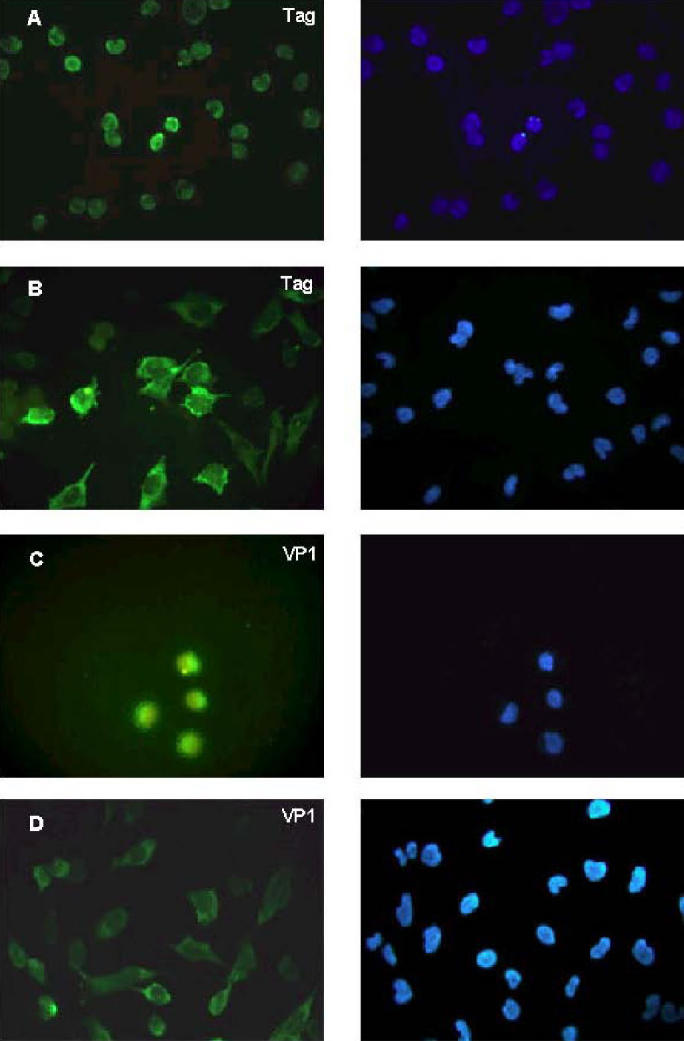
Cellular localization of viral Tag and VP1 proteins in SV40-infected CV-1 cells (A, C) and in human SV40-immortalized fibroblasts MRC5-SV2 (B, D). Fixed cells were incubated with the Tag specific monoclonal antibody pAb101 (DBA Italia, Segrate, Italy) (A, B,) to localize the Tag, or with goat anti-SV40 serum (C, D) to localize the VP1. An adequate secondary antibody conjugated with fluorescein isothiocyanate (FITC) was used to reveal the signals (left column). Nuclei were visualized with 4′, 6-diamino-2-phenylindole (0.5 mg/mL) (Sigma, Milano, Italy) (right column). Left and right panels represent the same field analyzed for the 2 different dyes.
DISCUSSION
It is well known that SV40 was administered to human population through contaminated vaccines (3,29). The possibility of a human-to-human transmission was taken into consideration only recently (2,30). Indeed, if SV40 is able to multiply in human cells, maintaining its original biological activity, the SV40 spreading in humans can be more than a hypothesis. This possibility is suggested by recent studies reporting SV40 footprints in human tumors and normal tissues (2,9,10). Moreover, in a recent report (1), the authors proposed that direct exposure to SV40 lead to development of a brain tumor in a laboratory researcher. Furthermore, complete SV40 genomes have been cloned from a long-term SV40-immortalized human fibroblast cell line (GM00637) (23), which is widely employed in many laboratories.
Our study indicates that SV40 can establish a persistent infection in human fibroblasts lasting after the “crisis” stage. Indeed, we show that the SV40 genome characterized in MRC5-SV2 immortalized cells is wild-type and indistinguishable from the input virus. Furthermore, these cells release an infectious viral progeny able to infect with the same efficiency both monkey and human cells. The appearance of cytopathic effect induced by the viruses recovered from the human immortalized cells is slower than that induced by the same virus released by permissive CV1 cells. At present, it is not clear why SV40 viruses 45-54-2MRC5 and 45-54-2CV-1 behave differently during cell infection. Probably, some properties of the SV40 virions may depend on cellular factors rather than on specific SV40 mutations. In SV40-immortalized human cells Tag and VP1 were detected mainly in the cytoplasm, suggesting specific interactions with host proteins present in this compartment (31–35). On the other hand, it is well established that SV40 virion morphology is strictly dependent on nuclear host proteins (36,37), and it has been shown that 45-54-2MRC5 virions display an altered morphology in comparison to wild type 45-54-2CV-1 (24). Further investigations are needed to elucidate the reciprocal influence of viral and cellular proteins during post-translation process and probably in viral particles assembly. Our data on VP1 coding sequences and Western blot analysis in human fibroblasts indicate that this viral protein is wild-type. Thus, it is reasonable to infer that these cells could influence viral capsid proteins at the post-translation level. Indeed, it has been demonstrated that heat shock cognate protein 70 (hsc70) binds to VP1 immediately after translation (38), and very likely, it plays an active role during virus assembly, inhibiting premature assembly in the cytosol (39).
It is worth noting that MRC5-SV2 cells were reported to be virus producers before the “crisis” stage, but then they lost the ability to shed virus after the immortalization occurred (>100 population doublings in culture) (20). It is therefore possible that before the “crisis” stage SV40 induces in human fibroblasts an “acute” infection, which is somehow controlled by those cells overtaking crisis, in a sort of latency period. During this stage, immortalized human fibroblasts do not produce a viral progeny, but they allow the replication of the episomal SV40 DNA. Subsequently, in a fraction of the cell population, a complete replicative SV40 cycle occurs, thus establishing a persistent infection with virus production.
This scenario suggests to interpret with caution the data on the biological characterization of SV40-immortalized human fibroblasts. Indeed, the possibility of a SV40 persistent infection in human fibroblasts represents a new element to take into consideration in evaluating the biological activity of this viral agent in the human host.
The different aspects of the SV40 behavior indicate that laboratory investigators are exposed to SV40 infection not only during specific procedures involving virus spread, such as sonication, but also during routine laboratory techniques, as cell trypsinization and centrifugations. Altogether these data suggest one should handle SV40 with additional care, indicating in the biohazard laboratory the appropriate containment for this oncogenic virus.
Acknowledgments
This study was granted by Associazione Italiana per la Ricerca sul Cancro and M.I.U.R. Cofin and local projects. Laura Iaccheri was supported by a fellowship from the Fondazione Cassa di Risparmio di Cento.
Footnotes
Online address: http://www.molmed.org>
REFERENCES
- 1.Arrington AS, Moore MS, Butel JS. SV40-positive brain tumor in scientist with risk of laboratory exposure to the virus. Oncogene. 2004;23:2231–5. doi: 10.1038/sj.onc.1207341. [DOI] [PubMed] [Google Scholar]
- 2.Barbanti-Brodano G, et al. Simian virus 40 infection in humans and association with human diseases: results and hypotheses. Virology. 2004;318:1–9. doi: 10.1016/j.virol.2003.09.004. [DOI] [PubMed] [Google Scholar]
- 3.Shah K, Nathanson N. Human exposure to SV40: review and comment. Am J Epidemiol. 1976;103:1–12. doi: 10.1093/oxfordjournals.aje.a112197. [DOI] [PubMed] [Google Scholar]
- 4.Carbone M, et al. Simian virus-40 large-T antigen binds p53 in human mesotheliomas. Nat Med. 1997;3:908–12. doi: 10.1038/nm0897-908. [DOI] [PubMed] [Google Scholar]
- 5.Ferber D. Virology. Monkey virus link to cancer grows stronger. Science. 2002;296:1012–5. doi: 10.1126/science.296.5570.1012. [DOI] [PubMed] [Google Scholar]
- 6.Strickler HD, et al. Contamination of poliovirus vaccines with simian virus 40 (1955–1963) and subsequent cancer rates. JAMA. 1998;279:292–5. doi: 10.1001/jama.279.4.292. [DOI] [PubMed] [Google Scholar]
- 7.Li RM, Branton MH, Tanawattanacharoen S, Falk RA, Jennette JC, Kopp JB. Molecular identification of SV40 infection in human subjects and possible association with kidney disease. J Am Soc Nephrol. 2002;13:2320–30. doi: 10.1097/01.asn.0000028249.06596.cf. [DOI] [PubMed] [Google Scholar]
- 8.Butel JS, Lednicky JA. Cell and molecular biology of simian virus 40: implications for human infections and disease. J Natl Cancer Inst. 1999;91:119–34. doi: 10.1093/jnci/91.2.119. [DOI] [PubMed] [Google Scholar]
- 9.Barbanti-Brodano G, et al. BK and JC human polyomaviruses and simian virus 40: natural history of infection in humans, experimental oncogenicity, and association with human tumors. Adv Virus Res. 1998;50:69–99. doi: 10.1016/s0065-3527(08)60806-4. [DOI] [PubMed] [Google Scholar]
- 10.Jasani B, et al. Association of SV40 with human tumours. Semin Cancer Biol. 2001;11:49–61. doi: 10.1006/scbi.2000.0346. [DOI] [PubMed] [Google Scholar]
- 11.Garcea RL, Imperiale MJ. Simian virus 40 infection of humans. J Virol. 2003;77:5039–45. doi: 10.1128/JVI.77.9.5039-5045.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jafar S, Rodriguez-Barradas M, Graham DY, Butel JS. Serological evidence of SV40 infections in HIV-infected and HIV-negative adults. J Med Virol. 1998;54:276–84. doi: 10.1002/(sici)1096-9071(199804)54:4<276::aid-jmv7>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- 13.Basetse HR, Lecatsas G, Gerber LJ. An investigation of the occurrence of SV40 antibodies in South Africa. S Afr Med J. 2002;92:825–8. [PubMed] [Google Scholar]
- 14.Bright RK, Kimchi ET, Shearer MH, Kennedy RC, Pass HI. SV40 Tag-specific cytotoxic T lymphocytes generated from the peripheral blood of malignant pleural mesothelioma patients. Cancer Immunol Immunother. 2002;50:682–90. doi: 10.1007/s00262-001-0240-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.O’Neill FJ, Carney H, Hu Y. Host range analysis of simian virus 40, BK virus and chimeric SV40/BKV: relative expression of large T-antigen and Vp1 in infected and transformed cells. Dev Biol Stand. 1998;94:191–205. [PubMed] [Google Scholar]
- 16.O’Neill FJ, Carroll D. Amplification of papovavirus defectives during serial low multiplicity infections. Virology. 1981;112:800–3. doi: 10.1016/0042-6822(81)90330-5. [DOI] [PubMed] [Google Scholar]
- 17.Bocchetta M, et al. Human mesothelial cells are unusually susceptible to simian virus 40-mediated transformation and asbestos cocarcinogenicity. Proc Natl Acad Sci USA. 2000;97:10214–9. doi: 10.1073/pnas.170207097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cacciotti P, et al. SV40 replication in human mesothelial cells induces HGF/Met receptor activation: a model for viral-related carcinogenesis of human malignant mesothelioma. Proc Natl Acad Sci USA. 2001;98:12032–7. doi: 10.1073/pnas.211026798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Shay JW, Wright WE. Quantitation of the frequency of immortalization of normal human diploid fibroblasts by SV40 large T-antigen. Exp Cell Res. 1989;184:109–18. doi: 10.1016/0014-4827(89)90369-8. [DOI] [PubMed] [Google Scholar]
- 20.Huschtscha LI, Holliday R. Limited and unlimited growth of SV40-transformed cells from human diploid MRC-5 fibroblasts. J Cell Sci. 1983;63:77–99. doi: 10.1242/jcs.63.1.77. [DOI] [PubMed] [Google Scholar]
- 21.Hara H, Kaji H. Random integration of SV40 in SV40-transformed, immortalized human fibroblasts. Exp Cell Res. 1987;168:531–8. doi: 10.1016/0014-4827(87)90025-5. [DOI] [PubMed] [Google Scholar]
- 22.Huang KC, Yamasaki EF, Snapka RM. Maintenance of episomal SV40 genomes in GM637 human fibroblasts. Virology. 1999;262:457–69. doi: 10.1006/viro.1999.9952. [DOI] [PubMed] [Google Scholar]
- 23.Forsman ZH, et al. Phylogenetic analysis of polyomavirus simian virus 40 from monkeys and humans reveals genetic variation. J Virol. 2004;78:9306–16. doi: 10.1128/JVI.78.17.9306-9316.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Morelli C, Barbisan F, Iaccheri L, Tognon M. Simian virus 40 persistent infection in long-term immortalized human fibroblast cell lines. J Neurovirol. 2004;10:250–4. doi: 10.1080/13550280490441185. [DOI] [PubMed] [Google Scholar]
- 25.Barbanti-Brodano G, et al. Reactivation of infectious simian virus 40 from normal human tissues. J Neurovirol. 2004;10:199–205. doi: 10.1080/13550280490441112. [DOI] [PubMed] [Google Scholar]
- 26.Martini F, et al. Different simian virus 40 genomic regions and sequences homologous with SV40 large T antigen in DNA of human brain and bone tumors and of leukocytes from blood donors. Cancer. 2002;94:1037–48. [PubMed] [Google Scholar]
- 27.Clever J, Yamada M, Kasamatsu H. Import of simian virus 40 virions through nuclear pore complexes. Proc Natl Acad Sci USA. 1991;88:7333–7. doi: 10.1073/pnas.88.16.7333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Martini F, et al. SV40 early region and large T antigen in human brain tumors, peripheral blood cells, and sperm fluids from healthy individuals. Cancer Res. 1996;56:4820–5. [PubMed] [Google Scholar]
- 29.Carbone M, Rizzo P, Pass HI. Simian virus 40, poliovaccines and human tumors: a review of recent developments. Oncogene. 1997;15:1877–88. doi: 10.1038/sj.onc.1201375. [DOI] [PubMed] [Google Scholar]
- 30.Vastag B. Sewage yields clues to SV40 transmission. JAMA. 2002;288:1337–8. doi: 10.1001/jama.288.11.1337. [DOI] [PubMed] [Google Scholar]
- 31.Sainis L, Angelidis C, Pagoulatos GN, Lazaridis L. HSC70 interactions with SV40 viral proteins differ between permissive and nonpermissive mammalian cells. Cell Stress Chaperones. 2000;5:132–8. doi: 10.1379/1466-1268(2000)005<0132:hiwsvp>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Klawitz I, Preuss U, Scheidtmann KH. Interaction of SV40 large T antigen with components of the nucleo/cytoskeleton. Int J Oncol. 2001;19:1325–32. doi: 10.3892/ijo.19.6.1325. [DOI] [PubMed] [Google Scholar]
- 33.Li PP, Nakanishi A, Clark SW, Kasamatsu H. Formation of transitory intra-chain and interchain disulfide bonds accompanies the folding and oligomerization of simian virus 40 Vp1 in the cytoplasm. Proc Natl Acad Sci USA. 2002;99:1353–8. doi: 10.1073/pnas.032668699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Nakanishi A, Shum D, Morioka H, Otsuka E, Kasamatsu H. Interaction of the Vp3 nuclear localization signal with the importin alpha 2/beta heterodimer directs nuclear entry of infecting simian virus 40. J Virol. 2002;76:9368–77. doi: 10.1128/JVI.76.18.9368-9377.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pelkmans L, Puntener D, Helenius A. Local actin polymerization and dynamin recruitment in SV40-induced internalization of caveolae. Science. 2002;296:535–9. doi: 10.1126/science.1069784. [DOI] [PubMed] [Google Scholar]
- 36.Gordon-Shaag A, Ben-Nun-Shaul O, Roitman V, Yosef Y, Oppenheim A. Cellular transcription factor Sp1 recruits simian virus 40 capsid proteins to the viral packaging signal, ses. J Virol. 2002;76:5915–24. doi: 10.1128/JVI.76.12.5915-5924.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Milavetz BI. SP1 and AP-1 elements direct chromatin remodeling in SV40 chromosomes during the first 6 hours of infection. Virology. 2002;294:170–9. doi: 10.1006/viro.2001.1308. [DOI] [PubMed] [Google Scholar]
- 38.Cripe TP, Delos SE, Estes PA, Garcea RL. In vivo and in vitro association of hsc70 with polyomavirus capsid proteins. J Virol. 1995;69:7807–13. doi: 10.1128/jvi.69.12.7807-7813.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chromy LR, Pipas JM, Garcea RL. Chaperone-mediated in vitro assembly of polyomavirus capsids. Proc Natl Acad Sci USA. 2003;100:10477–82. doi: 10.1073/pnas.1832245100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lednicky JA, Butel JS. Simian virus 40 regulatory region structural diversity and the association of viral archetypal regulatory regions with human brain tumors. Semin Cancer Biol. 2001;11:39–47. doi: 10.1006/scbi.2000.0345. [DOI] [PubMed] [Google Scholar]