

Abstract
Coupling of Rab GTPase activation and SNARE complex assembly during membrane fusion is poorly understood. The homotypic fusion and vacuole protein sorting (HOPS) complex links these two processes: it is an effector for the vacuolar Rab GTPase Ypt7p and is required for vacuolar SNARE complex assembly. We now report that pure, active HOPS complex binds phosphoinositides and the PX domain of the vacuolar SNARE protein Vam7p. These binding interactions support HOPS complex association with the vacuole and explain its enrichment at the same microdomains on docked vacuoles as phosphoinositides, Ypt7p, Vam7p, and the other SNARE proteins. Concentration of the HOPS complex at these microdomains may be a key factor for coupling Rab GTPase activation to SNARE complex assembly.
Keywords: HOPS, phosphoinositide, PX, Rab, SNARE
Introduction
Many of the proteins and lipids involved in membrane traffic are well-conserved (Jahn et al, 2003). Rab GTPases are ras-like proteins required for membrane tethering (Allan et al, 2000; Pfeffer, 2001). Diverse Rab effectors, which interact with GTP-bound Rab proteins, have been identified (Segev, 2001). Diverse multiprotein complexes also act in intracellular transport pathways (Peterson and Emr, 2001; Hsu et al, 2004; Babst, 2005; Oka et al, 2005); two, the conserved oligomeric Golgi complex and transport particle I complex, have been purified in active form (Walter et al, 1998; Sacher et al, 2001). SNARE proteins are integral or peripheral membrane proteins that bind each other to form four-helical SNARE complexes (Poirier et al, 1998) that act at a late stage of membrane fusion (Weber et al, 1998). Sec1/munc18 proteins bind SNAREs and SNARE complexes, promoting SNARE complex assembly (Pevsner et al, 1994; Carr et al, 1999; Peng and Gallwitz, 2002). Lipids also play a role in membrane trafficking. Ergosterol is required for yeast vacuole fusion (Kato and Wickner, 2001), and cholesterol is involved in localization of SNAREs (Lang et al, 2001). Phosphoinositides are required for vacuole fusion (Mayer et al, 2000) and regulate the localization of both Rabs and SNAREs (Boeddinghaus et al, 2002; Fratti et al, 2004).
Vacuoles from Saccharomyces cerevisiae are used to study membrane tethering and fusion. Fusion of purified vacuoles can be assayed in vitro (Haas et al, 1994). This reaction requires many of the same lipids and protein families as other fusion reactions, and thus uses the same mechanisms (Wickner and Haas, 2000). Vacuole fusion occurs in three stages: priming, docking, and mixing of lipid bilayers and lumenal contents. During priming, SNARE complexes are disassembled by Sec18p and its co-chaperone Sec17p (Mayer et al, 1996; Ungermann et al, 1998a). In the first phase of docking, vacuoles tether; this requires the Rab GTPase Ypt7p (Mayer and Wickner, 1997). Proteins and lipids important for fusion then become enriched in a ring encircling the apposed patches of membrane on each tethered vacuole. This microdomain is termed the vertex ring (Wang et al, 2002, 2003; Fratti et al, 2004). After the vertex ring has formed, SNAREs from the apposed membranes bind each other to form trans-SNARE complexes (Ungermann et al, 1998b; Wang et al, 2003; Dietrich et al, 2005). Finally, the membranes fuse around the vertex ring, internalizing the apposed membrane from each vacuole as a lumenal vesicle (Wang et al, 2002).
The homotypic fusion and vacuole protein sorting (HOPS) complex is a six-subunit complex that acts during the docking stage of yeast vacuole fusion (Price et al, 2000b). HOPS complex subunits are also involved in transport between endocytic compartments, as well as transport to the vacuole (Cowles et al, 1997; Rehling et al, 1999; Srivastava et al, 2000; Peterson and Emr, 2001). The HOPS complex is also termed the Class C vacuolar protein sorting (Vps) complex because it is composed of Vps proteins, which are encoded by genes required for proper protein sorting to the yeast vacuole (Raymond et al, 1992). The first proteins shown to be in the HOPS complex were Vps11p/Pep5p, Vps16p, Vps18p/Pep3p, and Vps33p (Rieder and Emr, 1997). Later, Vps39p/Vam6p and Vps41p/Vam2p were found in a complex with these four proteins (Seals et al, 2000; Wurmser et al, 2000). Vps11p, Vps16p, Vps18p, and Vps33p also form a complex, lacking Vps39p and Vps41p, with Vps8p (Subramanian et al, 2004). We will refer here to the complex of Vps11p, Vps16p, Vps18p, Vps33p, Vps39p, and Vps41p as the HOPS complex.
The HOPS complex interacts functionally and physically with Ypt7p and SNAREs. Vps39p is a nucleotide exchange factor for Ypt7p (Wurmser et al, 2000). HOPS binds to GTPγS-bound Ypt7p, and thus is also a Ypt7p effector (Seals et al, 2000). The HOPS complex is required for SNARE complex assembly and stably associates with SNARE complexes on vacuoles (Price et al, 2000a; Sato et al, 2000; Collins et al, 2005). This interaction may involve Vps33p, a member of the Sec1/munc18 family (Rieder and Emr, 1997; Dulubova et al, 2001).
The specific functions of the HOPS complex and the mechanisms by which it couples activated Ypt7p to SNARE complex assembly are unknown. To directly investigate HOPS complex functions, we have developed a method to purify HOPS from yeast, and an assay to show that pure HOPS complex is active. We now report that Rab GTPase-dependent tethering of vps11-1ts vacuoles requires pure HOPS complex. Rab GTPase, Rho GTPase, and SNARE catalysts of vacuole docking cannot complete their functions on vps11-1ts vacuoles without pure HOPS complex. Pure HOPS complex binds directly to phosphoinositides and to the phox homology (PX) domain of the soluble SNARE Vam7p. Thus, HOPS, phosphoinositides, and Vam7p each have mutual affinities, explaining their interdependent assembly into the vertex ring with Ypt7p. This high local concentration of HOPS, SNAREs, and Ypt7p may be a key element in coupling active Ypt7p to SNARE complex assembly during vacuole docking.
Results
We purified the HOPS complex using a streptavidin-binding peptide (SBP) fused to Vps33p (Keefe et al, 2001). Purified vacuoles (Seals et al, 2000) bearing Vps33-SBP were lysed in detergent and subjected to streptavidin affinity chromatography. Only the other five known HOPS complex subunits co-purified at high levels with SBP-tagged Vps33 (Figure 1). Ypt7p interacts with the HOPS complex in the presence of GTPγS (Seals et al, 2000), but no Ypt7p was detected in our HOPS preparations, even when GTPγS was added during Vps33p-SBP purification (not shown). Membrane association of Ypt7p may be required for HOPS–Ypt7p interactions at native Ypt7p levels.
Figure 1.
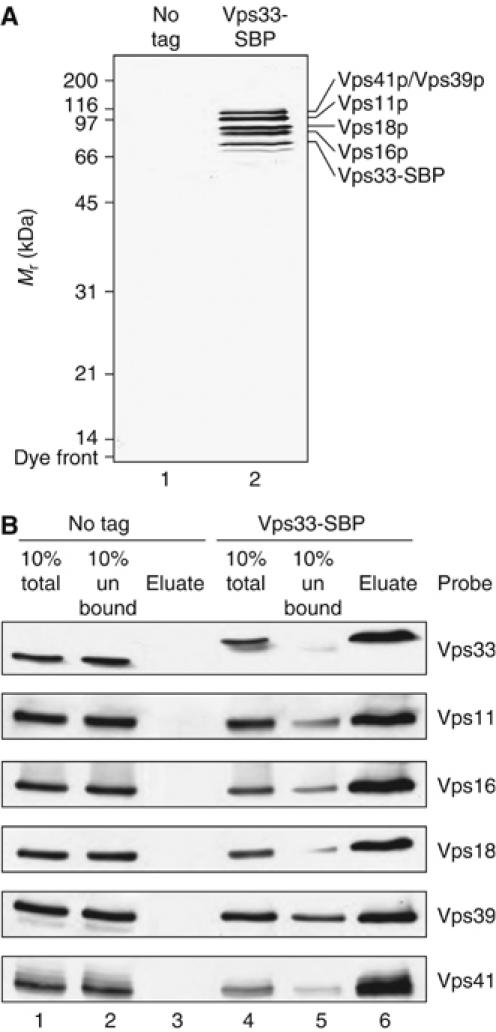
HOPS complex purification. (A) Lysates from BJ2168 (no tag) and CSY12 (Vps33-SBP) vacuoles were incubated with streptavidin Sepharose. Bound material was eluted in buffer containing Triton X-100 as described in Materials and methods and subjected to SDS–PAGE and silver staining. Lane 1, material purified from BJ2168 vacuoles; lane 2, material purified from CSY12 vacuoles. (B) Lysates from BJ3505 (lanes 1–3) and CSY12 (lanes 4–6) vacuoles were incubated with streptavidin Sepharose. Bound material was eluted in the presence of BSA and subjected to SDS–PAGE and immunoblotting. Lanes 1 and 4, vacuole lysate (representative of 10% of sample); lanes 2 and 5, material not bound to streptavidin Sepharose (representative of 10% of sample); lanes 3 and 6, material eluted from streptavidin Sepharose (representative of 100% of sample).
In addition to the other HOPS complex subunits, two proteins co-purify at low levels with Vps33-SBP in detergent solution (Figure 1A). Mass spectrometry analysis shows that the band below Vps33-SBP contains two peptides from Pho8p, the vacuolar alkaline phosphatase, and one peptide from the heat-shock protein Ssb1/2p. Because of the apparent molecular mass, we believe that this band is largely Pho8p. However, our final HOPS complex preparation is eluted in buffer lacking detergent and containing BSA, and contains neither the integral-membrane Pho8p protein (Supplementary Figure S1A) nor Pho8p activity (Supplementary Figure S1B). The faint band just above Vps33-SBP contains 56 peptides from three HOPS subunits and one peptide from the ER-lumenal chaperone Kar2p, and thus likely represents a breakdown product of HOPS complex subunits.
To test whether pure HOPS complex was functional, we used an in vitro assay of homotypic vacuole fusion. This assay (Haas, 1995) uses vacuoles from two yeast strains. One strain has normal vacuolar proteases but is deleted in PHO8. The other is deleted in PEP4, which encodes vacuolar proteinase A, and thus contains inactive pro-Pho8p. In vitro fusion of these vacuoles allows proteolytic activation of pro-Pho8p. We introduced a temperature-sensitive VPS11 allele, vps11-1ts (Peterson and Emr, 2001), into these strains. Fusion of vacuoles from vps11-1ts strains was stimulated by pure HOPS (Figure 2A, squares). Fusion was further stimulated by a low concentration (3.7 nM) of recombinant Vam7p (rVam7p; Figure 2A, triangles). This low level of rVam7p stimulated fusion of vps11-1ts vacuoles only slightly in the absence of pure HOPS complex (Figure 2A and C). High levels (2.7 μM) of rVam7p only partially restored fusion without pure HOPS complex (Figure 2A, open arrow; Figure 2C, squares). Pure HOPS complex did not stimulate fusion of vacuoles from strains bearing the wild-type VPS11 allele (Figure 2B and D). The modest (but statistically significant) inhibition of the fusion of VPS11 vacuoles by ∼4 nM HOPS complex in the presence of 2.7 μM rVam7p (Figure 2B, diamonds) may be due to their binding and sequestering other fusion factors. Thus, our pure HOPS complex is functionally active.
Figure 2.
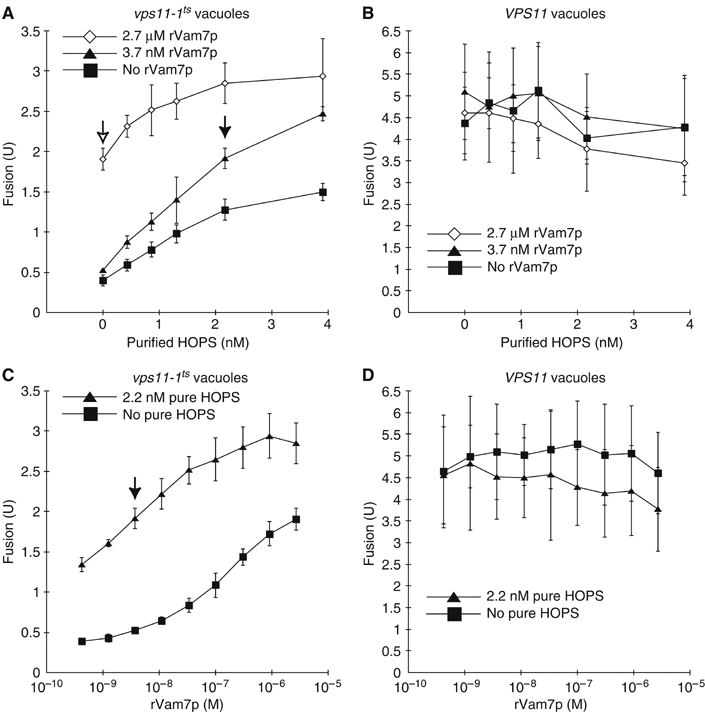
HOPS complex activity assay. Vps11-1ts (CSY9 and CSY10) and VPS11 (BY4742 pep4Δ∷kanMX6 and BY4742 pho8Δ∷kanMX6) vacuoles were assayed for fusion in the presence of pure HOPS complex and rVam7p. HOPS buffer was added, so the total volume of pure HOPS and HOPS buffer was 9 μl. Filled arrows indicate the fusion signal observed with the concentrations of pure HOPS complex (2.2 nM) and rVam7p was (3.7 nM) used in the fusion assays in the remainder of this paper. Means and standard deviations for fusion signals observed in three independent experiments are shown. (A) Pure HOPS added to vps11-1ts vacuoles, with the indicated concentrations of rVam7p. (B) Pure HOPS added to VPS11 vacuoles, with the indicated concentrations of rVam7p. (C) rVam7p added to vps11-1ts vacuoles, with or without pure HOPS. (D) rVam7p added to VPS11 vacuoles, with or without pure HOPS.
To determine whether the HOPS complex-dependent fusion of vps11-1ts vacuoles uses the normal fusion pathway, we tested the effects of well-characterized inhibitors of vacuole fusion (Figure 3). Antibodies to Sec17p and Sec18p blocked fusion in the absence of rVam7p and partially inhibited fusion in the presence of rVam7p, consistent with the finding that rVam7p can bypass the priming stage of vacuole fusion (Thorngren et al, 2004). Fusion was inhibited by excess recombinant Sec17p (rSec17p). Antibodies directed against Ypt7p, Vps33p, or the SNARE Vam3p also inhibited fusion. Thus, HOPS complex-dependent fusion of vps11-1ts vacuoles uses the same catalysts as fusion of VPS11 vacuoles.
Figure 3.
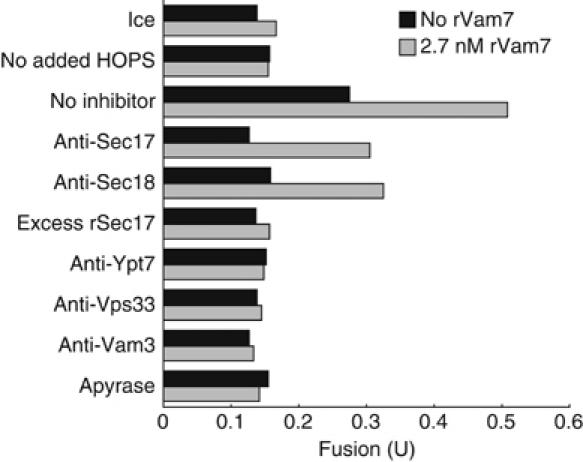
Authentic fusion of vps11-1ts vacuoles. Vps11-1ts vacuoles supplemented with pure HOPS complex were assayed for fusion in the absence (black bars) or presence (gray bars) of rVam7p and with the indicated inhibitors. All reactions received HOPS except for the reactions labeled ‘no added HOPS', which received an equal volume of HOPS buffer.
Vacuole tethering and docking require active HOPS complex
Which fusion catalysts can fulfil their function before HOPS complex action? To address this question, we incubated vps11-1ts vacuoles with rVam7p but without pure HOPS complex for 30 min, and then added inhibitors followed by pure HOPS complex and continued the incubations for 55 min. Reactions achieved resistance to anti-Sec17p antibodies prior to HOPS addition, and thus had completed their requirement for priming (Figure 4). Without pure HOPS complex, however, reactions did not become resistant to excess rSec17p, which inhibits vacuole fusion during docking and reduces HOPS association with SNARE complexes (Wang et al, 2000; Collins et al, 2005), or to anti-Ypt7p antibodies, anti-Vps33p antibodies, Rho GDP dissociation inhibitor (RDI), or anti-Vam3p antibodies (Figure 4). The reduced fusion signal after incubation without pure HOPS complex may be due to proteolytic sensitivity of vps11-1ts vacuoles (Supplementary Figure S1B, sets 14 and 16); also, reactions in the third set contained HOPS for only 55 min, whereas reactions in the first two sets contained HOPS for 90 min. Reactions with pure HOPS complex added at the start of the incubation became resistant to each of these inhibitors within 30 min (Figure 4). Thus, Ypt7p, Vps33p (which is present on vps11-1ts vacuoles; data not shown), Vam3p, and Rho GTPases cannot fulfil their function in the absence of HOPS complex activity. All of these fusion catalysts are needed for vacuole docking (Mayer and Wickner, 1997; Nichols et al, 1997; Seals et al, 2000; Eitzen et al, 2001). The HOPS complex is therefore needed for catalysts of docking, but not of priming, to complete their functions.
Figure 4.
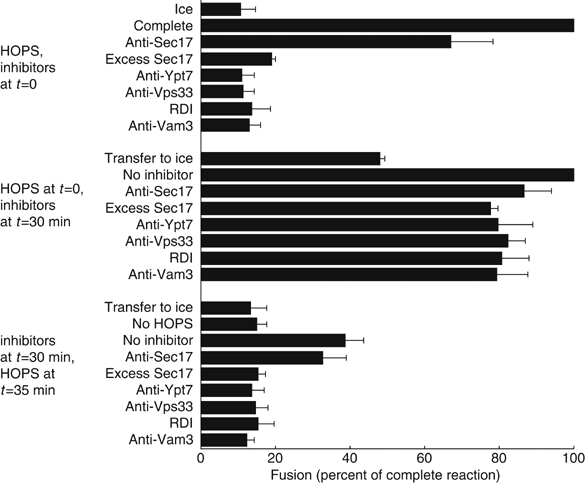
Staging of HOPS complex function. Reactions with vps11-1ts vacuoles and 3.7 nM rVam7p were divided into three sets. One set received inhibitors and pure HOPS at the beginning of the assay and HOPS buffer at 35 min. A second set received pure HOPS at the beginning of the assay, inhibitors at 30 min, and HOPS buffer at 35 min. The third set received HOPS buffer at the beginning of the assay, inhibitors at 30 min, and pure HOPS at 35 min. All reactions were assayed for fusion at 90 min. Means and standard deviations of the fusion in three independent experiments, expressed as a percentage of the fusion signal of each ‘complete' reaction (no inhibitor) before averaging, are shown.
It has been proposed that HOPS may support membrane tethering (Price et al, 2000a; Srivastava et al, 2000; Richardson et al, 2004; LaGrassa and Ungermann, 2005), but the lack of pure, active HOPS complex has precluded a direct test of this model. We therefore incubated vps11-1ts vacuoles, in the presence or absence of pure HOPS complex, under conditions that permit assay of membrane tethering (Mayer and Wickner, 1997; Wang et al, 2002). Pure HOPS complex increased the median number of vacuoles in each cluster. This is shown in representative fields (Figure 5A), in a cumulative distribution plot of all scored fields (Figure 5B), and in determination of median cluster size (Figure 5C). To test whether this increase required a Rab GTPase, we also incubated tethering reactions with Rab GDP dissociation inhibitor (GDI) and Gyp1-46, a catalytic fragment of a Rab GTPase activating protein (Garrett et al, 1994; Will et al, 2001). Gyp1-46/GDI treatment abolished the increase in median cluster size due to pure HOPS complex. Tethering did not occur in the absence of pure HOPS complex, as the median cluster size without added HOPS was not significantly decreased by Gyp1-46/GDI (Figure 5C). Thus, the HOPS complex is required for Rab GTPase-dependent vacuole tethering.
Figure 5.
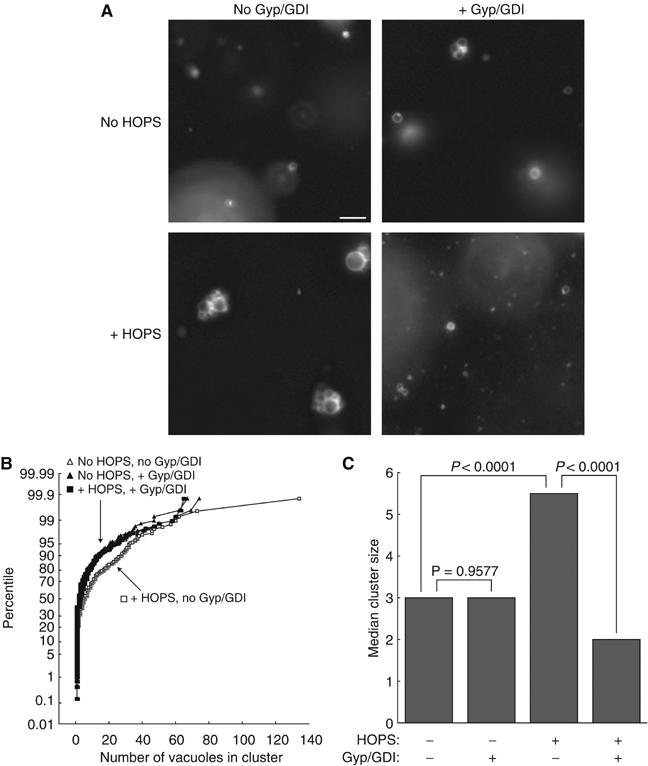
The HOPS complex is required for vacuole tethering. Vps11-1ts vacuoles (CSY10) were incubated in the presence or absence of pure HOPS and Gyp1-46 and GDI where indicated; no rVam7p was added. Samples were randomized and mounted on slides for fluorescence microscopy. Random fields were imaged. For each sample, the number of vacuoles in ∼100 vacuole clusters was counted. Data from three independent experiments were pooled for analysis. Bar, 5 μm. (A) Representative images of vacuoles from each sample in one experiment. The enlarged vacuoles in samples with HOPS but without Gyp1-46/GDI reflect fusion, and cause an underestimation of the number of vacuoles that had initially tethered in these clusters. (B) Cumulative distribution plot of the pooled data. (C) Median cluster sizes for the four treatments.
The HOPS complex binds phosphoinositides
Phosphoinositides are required for enrichment of the HOPS complex at the vertex ring during docking (Fratti et al, 2004). To determine whether a direct interaction between the HOPS complex and phosphoinositides might contribute to this localization, we performed protein–lipid overlay assays using pure HOPS. Pure HOPS complex was incubated with nitrocellulose strips bearing adsorbed phospholipid spots. Bound HOPS complex was detected using antibodies to each HOPS subunit (Figure 6A). Each HOPS complex subunit associated with the same phosphoinositides; since pure HOPS complex, not individual subunits, was used in these assays, this suggests that the intact complex interacts with phosphoinositides. The HOPS complex interacted most strongly with singly phosphorylated phosphoinositides. The HOPS complex did not associate strongly with phosphatidylinositol (PI) or other acidic phospholipids such as phosphatidylserine (PS) or phosphatidic acid (PA) (Figure 6A). Removal of trace amounts of Vam7p, a HOPS-binding protein (see below) that interacts with PI-3-phosphate (PI(3)P; Cheever et al, 2001), did not reduce the amount of HOPS complex bound to PI(3)P (Supplementary Figure S3).
Figure 6.
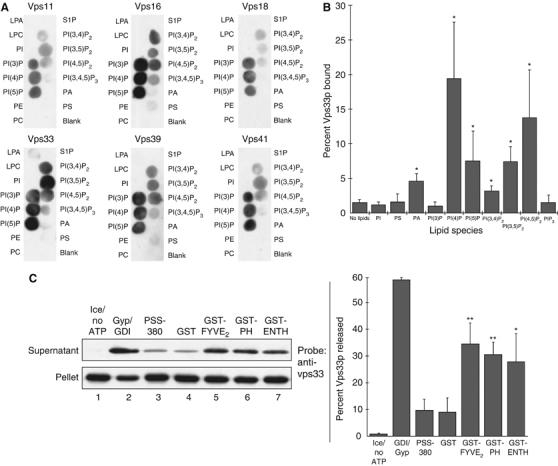
The HOPS complex binds phosphoinositides. (A) PIP MicroStrips (Echelon) were blocked with HBSTG (20 mM NaHEPES, pH 7.8, 3 mg/ml defatted BSA, 200 mM NaCl, 0.1% Tween-20, 5% glycerol) and incubated with pure HOPS (0.5 ml of 0.33 nM HOPS in HBSTG) at 4°C. Strips were washed four times with HBSTG and probed with antibodies in HBSTG, then washed four times with HBSTG; secondary detection was carried out with HRP-conjugated donkey anti-rabbit antibodies (GE). Strips were washed four times with HBSTG and bound antibody was detected by enhanced chemiluminescence (GE). (B) HOPS binding to liposomes was assayed by a modification of a described method (Matsuoka et al, 1998). Liposomes with PI were made by mixing CHCl3 solutions of soy PC, soy PE, soy PI (Avanti), and rhodamine-DHPE (Invitrogen) at molar ratios of 50:25:25:0.025 and drying with N2 gas and vacuum. Lipids were resuspended in 250 μl RB (20 mM NaHEPES, pH 7.8, 100 mM NaCl, 5% glycerol, 250 mM sorbitol; 8 mM total lipids), and subjected to freeze–thaw cycles (liquid N2/bath sonication) until the suspension was clear. Liposomes with DOPA, brain PS (Avanti), or phosphoinositides (Echelon; in 1:2:0.8 CHCl3:MeOH:H2O) were prepared in the same way, with 20–25% of the PI replaced by the appropriate phospholipid. Liposomes (20 μl) or RB were mixed in 7 × 20 mm tubes (Beckman) with 10 μl pure HOPS (2.6 nM final) and 10 μl of HBSG (20 mM NaHEPES, pH 7.8, 200 mM NaCl, 5% glycerol, 250 mM sorbitol) with 1.8 M sucrose and 2 mg/ml defatted BSA, then incubated on ice for 30 min. This mixture was overlayed with 75 μl of HBSG with 0.5 M sucrose, 75 μl of HBSG with 0.3 M sucrose, and 50 μl of HBSG, then centrifuged (50 000 r.p.m., 1 h, 4°C, TLS-55 rotor). Liposomes (40 μl) were taken from the border between the top layers, and bound HOPS was quantified with a ChemiDoc system (UVP) after SDS–PAGE and immunoblotting for Vps33p, with a standard curve of pure HOPS. Liposome recovery was estimated using rhodamine-DHPE fluorescence (λex/λem 510/580 nm); reported bound HOPS was corrected for liposome recovery. Mean percentages and standard deviations for at least three independent experiments are shown. *P<0.05 for differences relative to binding to PI liposomes, by one-way ANOVA. (C) HOPS release assay: fusion assays were supplemented as indicated and incubated on ice or at 27°C for 90 min, then sedimented (16 000 g, 15 min, 4°C). Supernatants and pellets were subjected to SDS–PAGE and immunoblotting for Vps33p. Blots were quantified by densitometry. Left, representative immunoblot. Right, mean percentages and standard deviations of Vps33p released in three independent experiments. **P<0.01, *P<0.05, for differences relative to release in the presence of GST, by one-way ANOVA.
To determine whether the HOPS complex can interact with phosphoinositides in a phospholipid bilayer, we performed binding assays using liposomes that contained each phosphoinositide, as well as liposomes bearing PS, PA, or unphosphorylated PI. Liposomes were mixed with pure HOPS complex and floated in a sucrose gradient, then harvested; bound HOPS was quantified by immunoblotting. HOPS complex bound to liposomes containing PI-4-phosphate (PI(4)P), PI-3,5-bisphosphate (PI(3,5)P2), or PI-4,5-bisphosphate (PI(4,5)P2), but interacted only weakly with liposomes bearing PS, PA, or PI (Figure 6B). (P<0.05 by one-way ANOVA for PI(4)P, PI(3,5)P2, or PI(4,5)P2-containing liposomes relative to PI or PA liposomes.) One difference between the results of the liposome assay and the overlay assay is that HOPS did not bind liposomes bearing PI(3)P. A PI(3)P-binding FYVE domain construct (Gillooly et al, 2000), however, did bind these liposomes (Supplementary Figure S4). This difference may be due to an interaction between the HOPS complex and the hydrophobic portion of the bilayer. This phenomenon, seen in the FYVE domain of Early Endosomal Antigen 1 (EEA1; Kutateladze and Overduin, 2001), may force HOPS into an orientation that prevents specific interactions with PI(3)P.
To determine whether the affinity of the HOPS complex for phosphoinositides contributes to its association with vacuoles, we incubated vacuole fusion reactions with ligands to specific phospholipids, then sedimented the vacuoles and assayed for HOPS complex release by SDS–PAGE and immunoblotting. Little HOPS complex was released when fusion reactions were incubated on ice, with PSS-380 (a PS ligand) or with GST (Figure 6B). However, HOPS complex was released when fusion reactions were incubated with ligands specific for phosphoinositides. GST fusions to a tandem FYVE domain specific for PI(3)P (Gillooly et al, 2000), a pleckstrin homology (PH) domain specific for PI(4)P (Weixel et al, 2005), or an ENTH domain specific for PI(4,5)P2 (Rosenthal et al, 1999) caused HOPS complex release from vacuoles (Figure 6C). HOPS complex release may be caused by competition of these lipid ligands with HOPS for phosphoinositide binding. However, HOPS complex release may be an indirect effect due to release of HOPS-binding proteins, such as Vam7p (see below), which also bind phosphoinositides (Cheever et al, 2001). Release of Vam7p correlated almost perfectly with release of HOPS complex in these experiments (not shown), even when a PI(4)P or PI(4,5)P2 ligand was present. Release of HOPS by these ligands may cause Vam7p release, as the Vam7p PX domain has no affinity for PI(4)P or PI(4,5)P2 (Cheever et al, 2001).
The HOPS complex binds the PX domain of the soluble SNARE Vam7p
The HOPS complex associates with SNARE complexes (Collins et al, 2005), and GST fusions to Vam3p and its SNARE domain interact with HOPS (Sato et al, 2000; Dulubova et al, 2001). However, it is not clear whether other individual SNAREs can also associate with the HOPS complex. We therefore incubated vacuole lysates with purified GST-tagged vacuolar SNAREs and examined bound proteins by immunoblotting. HOPS complex from vacuole lysates showed the strongest interaction with GST-Vam7p (Figure 7A, lane 13). Only a weak association was seen between the HOPS complex and GST–Vam3p or GST–Nyv1p, and no association between HOPS and GST–Vti1p was seen (Figure 7A and longer exposures of the same immunoblots, not shown).
Figure 7.
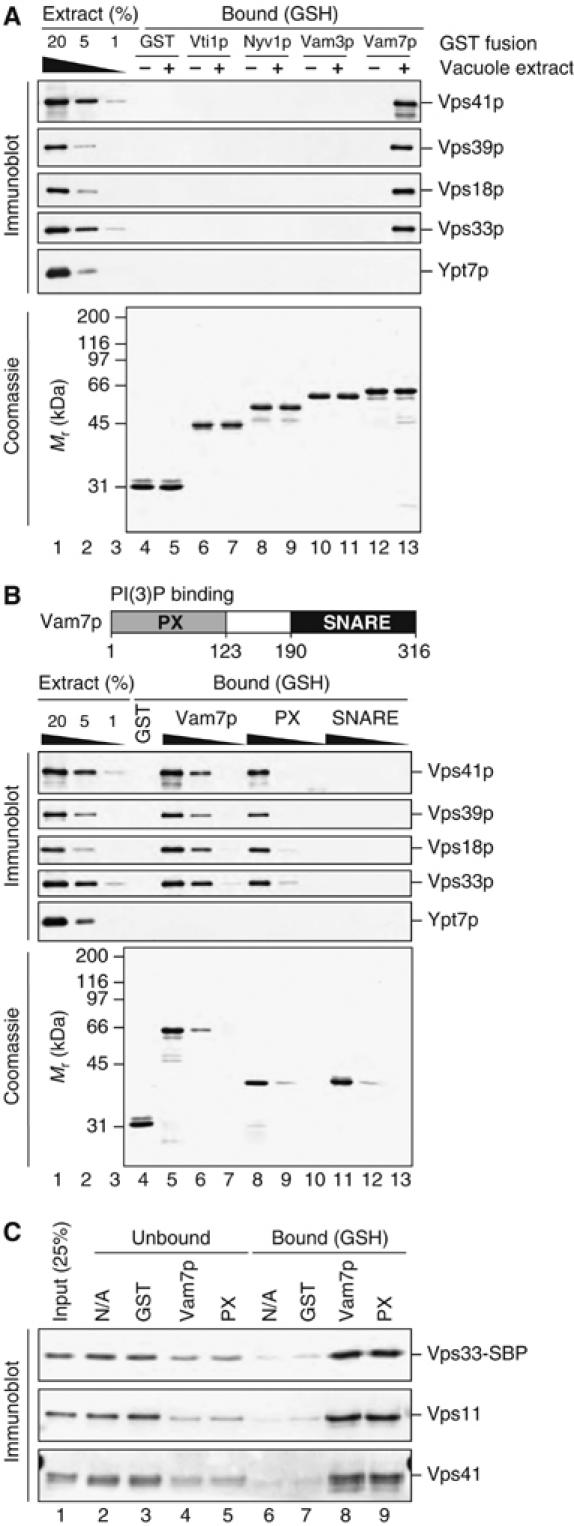
The HOPS complex binds Vam7p and the Vam7p PX domain. (A) HOPS binds recombinant Vam7p. Vacuoles (1.2 mg) from BJ3505 yeast were sedimented (16 000 g, 10 min, 4°C), resuspended in 2 ml SB (20 mM HEPES–KOH, pH 7.4, 100 mM NaCl, 2 mM EDTA, 0.5% Triton X-100, 20% glycerol, 0.46 μg/ml leupeptin, 3.5 μg/ml pepstatin, 2.4 μg/ml pefabloc-SC, 1 mM DTT) and incubated on ice for 20 min. Insoluble material was removed by centrifugation (TLA 120.2, 52 000 r.p.m., 11 min, 4°C). A sample of lysate was removed, and 200 μl of SB (lanes 4, 6, 8, 10, and 12) or vacuole lysate (lanes 5, 7, 9, 11, and 13) was incubated with 5 μg each of pure GST (lanes 4 and 5) or GST fused to the cytoplasmic residues of indicated SNAREs (lanes 6 and 7: Vti1p; lanes 8 and 9: Nyv1p; lanes 10 and 11: Vam3p; lanes 12 and 13: Vam7p). Lanes 1–3 represent 20, 5, and 1% of each lysate added. Fusion proteins were retrieved with glutathione Sepharose and bound material was analyzed by SDS–PAGE and immunoblotting (top) or staining with Coomassie blue (40% of eluted material, bottom). (B) The HOPS complex interacts with the Vam7p PX domain. GST (lane 4) or GST fusions to full-length Vam7p (lanes 5–7), the PX domain (lanes 8–10), or the SNARE domain (lanes 11–13) were added (5 μg for GST alone, and 5, 1, and 0.2 μg for Vam7p, PX, and SNARE domains) to vacuole lysates prepared in parallel to those in (A). Lanes 1–3 represent 20, 5, and 1% of each lysate added. Fusion proteins were retrieved with glutathione Sepharose (GE) and bound material was analyzed by SDS–PAGE. (C) The HOPS complex binds directly to the Vam7p PX domain. Pure HOPS (0.65 nM; lane 1) was incubated with glutathione Sepharose and buffer (lanes 2 and 6) or 5 μg GST (lanes 3 and 7), GST-Vam7 (lanes 4 and 8), or GST-Vam7 PX domain (lanes 5 and 9), in a total volume of 200 μl of SB; bound material was eluted by heating with SDS. Unbound (lanes 2–5) and bound (lanes 6–9) material was analyzed by SDS–PAGE and immunoblotting.
To determine which domain of Vam7p mediates the interaction between GST-Vam7p and the HOPS complex, we incubated vacuole lysates with GST fusions to full-length Vam7p, the amino-terminal PX domain of Vam7p, or the carboxy-terminal SNARE domain of Vam7p. HOPS complex from the lysates bound fusions of GST to full-length Vam7p or the PX domain of Vam7p but not a GST fusion to the Vam7p SNARE domain (Figure 7B).
Does the HOPS complex interact directly with the PX domain of Vam7p, or does this interaction require other proteins? To address this question, we incubated pure HOPS complex with purified fusions of GST to Vam7p or to the PX domain from Vam7p. HOPS complex, containing only the six known subunits (Figure 1A), bound efficiently to GST fusions to either full-length Vam7p or the Vam7p PX domain on glutathione beads (Figure 7C, lanes 8 and 9).
Discussion
The HOPS complex is at the center of vacuole docking mechanisms. It is an exchange factor and an effector for Ypt7p (Seals et al, 2000; Wurmser et al, 2000). It is required for SNARE complex assembly and remains bound to SNARE complexes after vacuole fusion (Price et al, 2000b; Sato et al, 2000; Collins et al, 2005). Purification of functional HOPS complex (Figures 1, 2 and 3) has allowed us to show that HOPS is required for vacuole tethering (Figure 5). We have also shown that the HOPS complex directly binds phosphoinositides (Figure 6) and the PX domain of Vam7p (Figure 7), crucial components of the vertex docking ring that are required for vertex ring assembly (Wang et al, 2003; Fratti et al, 2004). The HOPS complex is required for other catalysts of docking, including SNAREs, Ypt7p, and Rho GTPases, to complete their functions (Figure 4).
The physical and functional interactions of the HOPS complex suggest a model of how it links Ypt7p activation to SNARE complex assembly. After Vps39p activates Ypt7p by catalyzing the exchange of bound GDP for GTP, HOPS may remain associated with GTP-bound Ypt7p. The HOPS complex and Ypt7p then act together to promote tethering. The isoprenyl anchors of Ypt7p and the affinity of the HOPS complex for phosphoinositides and the PX domain of Vam7p may promote enrichment of HOPS, Ypt7p, and Vam7p at the vertex ring. Both the HOPS complex and PI(3)P bind Vam7p through its PX domain, which may initiate the assembly of a SNARE complex associated with HOPS. HOPS remains associated with newly assembled SNARE complexes, where it may regulate or contribute to their roles in hemifusion and fusion. Finally, Sec17p displaces the HOPS complex from the SNARE complex, allowing HOPS to engage in another cycle of SNARE complex assembly and preparing the SNARE complex for disassembly by Sec17p and Sec18p.
Our finding that the HOPS complex binds phosphoinositides and the Vam7p PX domain suggests that HOPS is enriched at the vertex ring through direct interactions with Ypt7p, Vam7p, and PI(4,5)P2. All of these HOPS-binding partners are enriched at the vertex ring, and inhibitors directed at Ypt7p, Vam7p, or PI(4,5)P2 block HOPS enrichment at the vertex ring (Wang et al, 2002, 2003; Fratti et al, 2004; data not shown). Some of the effect of phosphoinositide ligands on HOPS vertex enrichment may be indirect: both the PI(4,5)P2-binding ENTH domain and the PX domain of Vam7p inhibit Vam3p vertex enrichment (Wang et al, 2003; Fratti et al, 2004), and anti-Vam3p Fab fragments block enrichment of HOPS at the vertex ring (Wang et al, 2003).
The interaction between the HOPS complex and the Vam7p PX domain may initiate the assembly of SNARE complexes, possibly in combination with an interaction between HOPS and Vam3p. We have shown that the HOPS complex binds GST-tagged Vam7p preferentially over GST fusions to other SNAREs (Figure 7A). However, HOPS can also bind GST fusions to both full-length Vam3p and its SNARE domain (Sato et al, 2000; Dulubova et al, 2001). These results are not mutually exclusive: monomeric Vam7p and Vam3p may bind simultaneously or sequentially to HOPS before their assembly into a SNARE complex (Ungermann et al, 1999; Sato et al, 2000; Collins et al, 2005). This hypothesis is supported by the finding that a SNARE complex can assemble on a Sed5p–Sly1p complex (Peng and Gallwitz, 2002). Sed5p is a Vam3p homolog, and Sly1p is a Vps33p homolog, suggesting that Vps33p may be involved directly in SNARE complex assembly.
The multiple interactions of the HOPS complex are reminiscent of tethering factors that interact with proteins and lipids that can reside on different membranes. EEA1 tethers endosomal membranes by binding both Rab5 and PI(3)P (Lawe et al, 2002). p115 binds GM130, a Golgi matrix protein, and giantin, a protein from COPI vesicles (Nakamura et al, 1997; Sonnichsen et al, 1998). The exocyst complex binds the vesicular Rab Sec4p and the plasma membrane GTPase Rho1p (Boyd et al, 2004). This common feature of the HOPS complex and tethering factors, the ability to engage in multiple interactions, suggests that HOPS tethers membranes directly by binding to proteins and lipids in the apposed lipid bilayers.
The interaction between the HOPS complex and Pho8p is consistent with the requirement for Vps41p for formation of AP-3-dependent vesicles, which transport Pho8p from the Golgi apparatus to the vacuole (Rehling et al, 1999). Vps41p has been proposed to homo-oligomerize and act as a coat, perhaps stimulating vesicle budding (Darsow et al, 2001). If the HOPS complex is part of a coat for AP-3-dependent vesicles, then a HOPS–Pho8p interaction may contribute to sorting of Pho8p into these vesicles. Such an activity would be analogous to the COPII-dependent packaging of cargo into ER-derived vesicles, which requires interaction between the coat and the cargo protein Bet1p (Miller et al, 2003).
How does the HOPS complex bind phosphoinositides? The six HOPS complex subunits do not have any known lipid-binding motifs, although Vps11p and Vps18p have ‘really interesting new gene' (RING) motifs (Schultz et al, 1998). RING domains are involved in protein–protein interactions by ubiquitin-protein ligases (Joazeiro and Weissman, 2000). However, RING domains resemble plant homeodomain domains (Aravind et al, 2003), one of which binds PI-5-phosphate (Gozani et al, 2003). It is also possible that the phosphoinositide-binding site in the HOPS complex contains residues from more than one subunit; it has been shown that a lipid-binding PH domain is formed by partial PH domains from PLC-γ1 and TRPC3 (van Rossum et al, 2005).
It is likewise not known how the HOPS complex binds the Vam7p PX domain. The RING domain of Vps11p or Vps18p may mediate this interaction by interacting with the proline-rich loop between the α1 and α2 helices of the Vam7p PX domain (Cheever et al, 2001). The RING domain of the c-Cbl ubiquitin-protein ligase recognizes two proline-rich loops in the UbcH7 ubiquitin-conjugating enzyme (Zheng et al, 2000). However, the α1−α2 loop of the Vam7p PX domain may be involved in phosphoinositide binding (Cheever et al, 2001). Also, Vps18p and Vps41p contain clathrin heavy chain repeats (Schultz et al, 1998), which mediate clathrin self-association (Ybe et al, 1999). These motifs may be involved in HOPS binding to Vam7p. Another possible Vam7p ligand is a WD40 repeat near the Vps41 amino terminus (Schultz et al, 1998). This motif is required for Vps41p self-association and binding of Vps41p to Apl5p (Darsow et al, 2001), and may also interact with Vam7p.
The availability of pure, active HOPS complex will be essential for uncovering the mechanisms by which HOPS carries out its membrane tethering activity and couples Ypt7p activation to SNARE complex assembly. New in vitro assays for intermediate steps of membrane fusion that require pure HOPS may reveal currently unknown activities for this complex. Pure, active HOPS complex will be invaluable for reconstitution of the docking reactions in yeast vacuole fusion.
Materials and methods
Reagents
Anti-Sec18p (Haas and Wickner, 1996), anti-Sec17p (Haas and Wickner, 1996), anti-Ypt7p (Eitzen et al, 2001), anti-Vps33p (Seals et al, 2000), anti-Vam3p (Nichols et al, 1997), GST-RDI (Eitzen et al, 2001), Gyp1-46 (Will et al, 2001), GDI (Haas et al, 1995), PSS-380 (Fratti et al, 2004), GST-ENTH (Rosenthal et al, 1999), GST-FYVE2 (Gillooly et al, 2000), GST-PH (Weixel et al, 2005), GST-Vam7p and rVam7p (Merz and Wickner, 2004), GST-PX (Merz and Wickner, 2004), I2B (Slusarewicz et al, 1997), his6-Sec17p (Haas and Wickner, 1996), and his6-Sec18p (Thorngren et al, 2004) were described previously. Apyrase was from Sigma-Aldrich. A plasmid encoding a GST fusion to the Vam7p SNARE domain (residues 190–316) was the generous gift of Dr Alexey Merz.
Vacuole fusion assay
Vacuoles were purified and assayed for fusion as described (Haas, 1995). Vacuoles lacking Pep4p or Pho8p (3 μg each) were in a volume of 30 μl of 20 mM PIPES–KOH, pH 6.8, 200 mM sorbitol, 125 mM KCl, 5 mM MgCl2, 15 μM coenzyme A, 1 μg/ml his6-Sec18p, 7 μg/ml I2B, 3.3 ng/ml leupeptin, 3.3 ng/ml 1,10 phenanthroline, 16.7 ng/ml pepstatin A, 0.8 ng/ml pefabloc-SC, 1 mg/ml creatine kinase, 1 mM MgATP, and 40 mM creatine phosphate. Reactions were incubated on ice or at 27°C for 90 min and Pho8p activity was measured (Haas, 1995). Fusion inhibitors were at the following concentrations: anti-Sec18p, 39 μg/ml; anti-Sec17p, 113 μg/ml; excess rSec17p, 4 μg/ml; anti-Ypt7p, 100 μg/ml; anti-Vps33p, 28 μg/ml; anti-Vam3p, 133 μg/ml; GST-RDI, 833 μg/ml; apyrase, 1 U per reaction; GDI, 500 μg/ml; his6-Gyp1-46, 467 μg/ml.
HOPS complex purification
A plasmid based on pFA6-GFP(S65T)-TRP1 (Longtine et al, 1998), with the streptavidin-binding peptide coding sequence (Keefe et al, 2001) in place of the GFP(S65T) coding sequence, was the generous gift of Dr John Flanagan. The yeast strain CSY12 was prepared by transforming BJ2168 cells (Zubenko et al, 1980; Gietz and Schiestl, 1995) with a PCR product amplified from this plasmid using primers (Wang et al, 2002) for carboxy-terminal tagging of Vps33p.
For purification of HOPS complex, 12 mg CSY12 vacuoles (Seals et al, 2000) were thawed, diluted in 25 ml of 20 mM NaHEPES, pH 7.8, 200 mM sorbitol, and reisolated by centrifugation (13 000 r.p.m., JA-20 rotor, 20 min, 4°C). Vacuoles were resuspended to 0.6 mg/ml in lysis buffer (20 mM NaHEPES, pH 7.8, 400 mM NaCl, 5% glycerol, 0.5% Triton X-100, 5 mM β-mercaptoethanol, 0.46 μg/ml leupeptin, 3.5 μg/ml pepstatin, 2.4 μg/ml pefabloc-SC, 1 mM PMSF) and incubated on ice for 20 min. Lysate was clarified (50 000 r.p.m., Type 60 Ti rotor, 30 min, 4°C) and incubated with streptavidin Sepharose High Performance (2 ml; GE Healthcare) for 3 h at 4°C. The resin was drained and washed four times with 20 ml lysis buffer, four times with 20 ml BSA buffer (20 mM NaHEPES, pH 7.8, 400 mM NaCl, 5% glycerol, 5 mM β-mercaptoethanol, 2 mg/ml BSA), and eluted with 0.5 ml fractions of HOPS buffer (BSA buffer with 2 mg/ml biotin). Fractions were tested for HOPS complex activity (see below). Fractions with activity were pooled and frozen in 52 μl aliquots in liquid N2. HOPS was quantified as described in Supplementary Figure S2. For SDS–PAGE/silver stain analysis, bound proteins were eluted with 2 mg/ml biotin in lysis buffer after washing with lysis buffer.
HOPS complex activity assay
VPS11 was deleted from strains BY4742 pep4Δ∷kanMX6 and BY4742 pho8Δ∷kanMX6 (ATCC) by transformation with a PCR product amplified from pFA6-His3MX6 (Longtine et al, 1998) to make strains that were then transformed with pMP160, which bears the vps11-1ts allele (Peterson and Emr, 2001), to make CSY9 and CSY10, respectively.
Vacuoles were purified from CSY9 and CSY10 cells as described (Haas, 1995). Cultures were grown overnight in CSM-ura at 30°C. Cells (CSY9, 7 ml at OD600=1; CSY10, 5 ml at OD600=1) from these cultures were used to start cultures in 2% YPD, grown for ∼24 h at 25°C, and used for vacuole purification.
To assay pure HOPS, vacuole fusion reactions with vps11-1ts vacuoles were performed with pure HOPS and/or HOPS buffer. Sorbitol was added at 0.2 M to aliquots of pure HOPS complex or HOPS buffer before being added to fusion reactions. The [KCl] in the reactions was adjusted, so the sum of [NaCl] (from the HOPS buffer) and [KCl] was 125 mM. MgGTP was added at 10 μM; this stimulates fusion of vps11-1ts vacuoles by 10–20%, but does not stimulate fusion of VPS11 vacuoles (not shown). Where indicated, rVam7p was added.
Vacuole tethering assay
Tethering reactions were as described (Mayer and Wickner, 1997), except that they contained CSY10 vacuoles, pure HOPS (2.2 nM) or HOPS buffer, and 33 mM KCl (to keep the sum of [KCl] and [NaCl] at 100 mM). Reactions contained 6 μg CSY10 vacuoles, 20 mM PIPES–KOH, pH 6.8, 200 mM sorbitol, 33 mM KCl, 0.5 mM MgCl2, 20 μM coenzyme A, 1 μg/ml his6-Sec18p, 2.7 μM FM4-64 (Invitrogen), 0.7 mg/ml creatine kinase, 0.3 mM MgATP, 6 mM creatine phosphate, and 5 μl HOPS buffer. Reactions were randomized and incubated at 27°C for 30 min. Low melting point agarose in 20 mM PIPES–KOH, 200 mM sorbitol (50 μl of 0.6% solution) at 42°C was added and reactions were vortexed (3 s at medium setting). Aliquots (15 μl) were placed on glass slides, overlayed with coverslips and immersion oil (nd 1.516; Olympus), and imaged.
Imaging used an Olympus BX51 microscope with a 100 W mercury arc lamp, a Plan Apochromat × 60 objective (1.4 NA; Olympus), and a Sensicam QE CCD camera (Cooke). A U-25ND25 neutral density filter and a U-RSL6 UV/IR filter (Olympus) were used to reduce incident light. A TRITC/Dil filter set (Chroma Technologies) was used to collect FM4-64 images. Images were recorded using IPLab (Scanalytics) and vacuole clusters were counted manually using ImageJ (NIH). Wilcoxon–Mann–Whitney tests on data pooled from three independent experiments were carried out using KaleidaGraph (Synergy Software).
Supplementary Material
Figure S1
Figure S2
Figure S3
Figure S4
Acknowledgments
We thank S Emr, J Flanagan, and A Merz for sharing plasmid constructs, N Margolis for vacuole purification, and Y Jun, S Seeley, and N Thorngren for reagents. This work was supported by a grant (GM23377) from the NIH. SCS was supported by a fellowship (GM65710) from the NIH. KMC received support from the NIH (GM08704). RAF is supported by a fellowship from the Helen Hay Whitney Foundation.
References
- Allan BB, Weissman J, Aridor M, Moyer B, Chen CD, Yoo JS, Balch WE (2000) Stage-specific assays to study biosynthetic cargo selection and role of SNAREs in export from the endoplasmic reticulum and delivery to the Golgi. Methods 20: 411–416 [DOI] [PubMed] [Google Scholar]
- Aravind L, Iyer LM, Koonin EV (2003) Scores of RINGS but no PHDs in ubiquitin signaling. Cell Cycle 2: 123–126 [DOI] [PubMed] [Google Scholar]
- Babst M (2005) A protein's final ESCRT. Traffic 6: 2–9 [DOI] [PubMed] [Google Scholar]
- Boeddinghaus C, Merz AJ, Laage R, Ungermann C (2002) A cycle of Vam7p release from and PtdIns 3-P-dependent rebinding to the yeast vacuole is required for homotypic vacuole fusion. J Cell Biol 157: 79–89 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyd C, Hughes T, Pypaert M, Novick P (2004) Vesicles carry most exocyst subunits to exocytic sites marked by the remaining two subunits, Sec3p and Exo70p. J Cell Biol 167: 889–901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr CM, Grote E, Munson M, Hughson FM, Novick PJ (1999) Sec1p binds to SNARE complexes and concentrates at sites of secretion. J Cell Biol 146: 333–344 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheever ML, Sato TK, de Beer T, Kutateladze TG, Emr SD, Overduin M (2001) Phox domain interaction with PtdIns(3)P targets the Vam7 t-SNARE to vacuole membranes. Nat Cell Biol 3: 613–618 [DOI] [PubMed] [Google Scholar]
- Collins KM, Thorngren NL, Fratti RA, Wickner WT (2005) Sec17p and HOPS, in distinct SNARE complexes, mediate SNARE complex disruption or assembly for fusion. EMBO J 24: 1775–1786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowles CR, Snyder WB, Burd CG, Emr SD (1997) Novel Golgi to vacuole delivery pathway in yeast: identification of a sorting determinant and required transport component. EMBO J 16: 2769–2782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darsow T, Katzmann DJ, Cowles CR, Emr SD (2001) Vps41p function in the alkaline phosphatase pathway requires homo-oligomerization and interaction with AP-3 through two distinct domains. Mol Biol Cell 12: 37–51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dietrich LE, Peplowska K, Lagrassa TJ, Hou H, Rohde J, Ungermann C (2005) The SNARE Ykt6 is released from yeast vacuoles during an early stage of fusion. EMBO Rep 6: 245–250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dulubova I, Yamaguchi T, Wang Y, Sudhof TC, Rizo J (2001) Vam3p structure reveals conserved and divergent properties of syntaxins. Nat Struct Biol 8: 258–264 [DOI] [PubMed] [Google Scholar]
- Eitzen G, Thorngren N, Wickner W (2001) Rho1p and Cdc42p act after Ypt7p to regulate vacuole docking. EMBO J 20: 5650–5656 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fratti RA, Jun Y, Merz AJ, Margolis N, Wickner W (2004) Interdependent assembly of specific regulatory lipids and membrane fusion proteins into the vertex ring domain of docked vacuoles. J Cell Biol 167: 1087–1098 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrett MD, Zahner JE, Cheney CM, Novick PJ (1994) GDI1 encodes a GDP dissociation inhibitor that plays an essential role in the yeast secretory pathway. EMBO J 13: 1718–1728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gietz RD, Schiestl RH (1995) Transforming yeast with DNA. Meth Mol Cell Biol 5: 255–269 [Google Scholar]
- Gillooly DJ, Morrow IC, Lindsay M, Gould R, Bryant NJ, Gaullier JM, Parton RG, Stenmark H (2000) Localization of phosphatidylinositol 3-phosphate in yeast and mammalian cells. EMBO J 19: 4577–4588 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gozani O, Karuman P, Jones DR, Ivanov D, Cha J, Lugovskoy AA, Baird CL, Zhu H, Field SJ, Lessnick SL, Villasenor J, Mehrotra B, Chen J, Rao VR, Brugge JS, Ferguson CG, Payrastre B, Myszka DG, Cantley LC, Wagner G, Divecha N, Prestwich GD, Yuan J (2003) The PHD finger of the chromatin-associated protein ING2 functions as a nuclear phosphoinositide receptor. Cell 114: 99–111 [DOI] [PubMed] [Google Scholar]
- Haas A (1995) A quantitative assay to measure homotypic vacuole fusion in vitro. Methods Cell Sci 17: 283–294 [Google Scholar]
- Haas A, Conradt B, Wickner W (1994) G-protein ligands inhibit in vitro reactions of vacuole inheritance. J Cell Biol 126: 87–97 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas A, Scheglmann D, Lazar T, Gallwitz D, Wickner W (1995) The GTPase Ypt7p of Saccharomyces cerevisiae is required on both partner vacuoles for the homotypic fusion step of vacuole inheritance. EMBO J 14: 5258–5270 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas A, Wickner W (1996) Homotypic vacuole fusion requires Sec17p (yeast α-SNAP) and Sec18p (yeast NSF). EMBO J 15: 3296–3305 [PMC free article] [PubMed] [Google Scholar]
- Hsu SC, TerBush D, Abraham M, Guo W (2004) The exocyst complex in polarized exocytosis. Int Rev Cytol 233: 243–265 [DOI] [PubMed] [Google Scholar]
- Jahn R, Lang T, Sudhof TC (2003) Membrane fusion. Cell 112: 519–533 [DOI] [PubMed] [Google Scholar]
- Joazeiro CA, Weissman AM (2000) RING finger proteins: mediators of ubiquitin ligase activity. Cell 102: 549–552 [DOI] [PubMed] [Google Scholar]
- Kato M, Wickner W (2001) Ergosterol is required for the Sec18/ATP-dependent priming step of homotypic vacuole fusion. EMBO J 20: 4035–4040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keefe AD, Wilson DS, Seelig B, Szostak JW (2001) One-step purification of recombinant proteins using a nanomolar-affinity streptavidin-binding peptide, the SBP-Tag. Protein Expr Purif 23: 440–446 [DOI] [PubMed] [Google Scholar]
- Kutateladze T, Overduin M (2001) Structural mechanism of endosome docking by the FYVE domain. Science 291: 1793–1796 [DOI] [PubMed] [Google Scholar]
- LaGrassa TJ, Ungermann C (2005) The vacuolar kinase Yck3 maintains organelle fragmentation by regulating the HOPS tethering complex. J Cell Biol 168: 401–414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang T, Bruns D, Wenzel D, Riedel D, Holroyd P, Thiele C, Jahn R (2001) SNAREs are concentrated in cholesterol-dependent clusters that define docking and fusion sites for exocytosis. EMBO J 20: 2202–2213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawe DC, Chawla A, Merithew E, Dumas J, Carrington W, Fogarty K, Lifshitz L, Tuft R, Lambright D, Corvera S (2002) Sequential roles for phosphatidylinositol 3-phosphate and Rab5 in tethering and fusion of early endosomes via their interaction with EEA1. J Biol Chem 277: 8611–8617 [DOI] [PubMed] [Google Scholar]
- Longtine MS, McKenzie A III, Demarini DJ, Shah NG, Wach A, Brachat A, Philippsen P, Pringle JR (1998) Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 14: 953–961 [DOI] [PubMed] [Google Scholar]
- Matsuoka K, Orci L, Amherdt M, Bednarek SY, Hamamoto S, Schekman R, Yeung T (1998) COPII-coated vesicle formation reconstituted with purified coat proteins and chemically defined liposomes. Cell 93: 263–275 [DOI] [PubMed] [Google Scholar]
- Mayer A, Scheglmann D, Dove S, Glatz A, Wickner W, Haas A (2000) Phosphatidylinositol 4,5-bisphosphate regulates two steps of homotypic vacuole fusion. Mol Biol Cell 11: 807–817 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer A, Wickner W (1997) Docking of yeast vacuoles is catalyzed by the Ras-like GTPase Ypt7p after symmetric priming by Sec18p (NSF). J Cell Biol 136: 307–317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer A, Wickner W, Haas A (1996) Sec18p (NSF)-driven release of Sec17p (α-SNAP) can precede docking and fusion of yeast vacuoles. Cell 85: 83–94 [DOI] [PubMed] [Google Scholar]
- Merz AJ, Wickner WT (2004) Trans-SNARE interactions elicit Ca2+ efflux from the yeast vacuole lumen. J Cell Biol 164: 195–206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller EA, Beilharz TH, Malkus PN, Lee MC, Hamamoto S, Orci L, Schekman R (2003) Multiple cargo binding sites on the COPII subunit Sec24p ensure capture of diverse membrane proteins into transport vesicles. Cell 114: 497–509 [DOI] [PubMed] [Google Scholar]
- Nakamura N, Lowe M, Levine TP, Rabouille C, Warren G (1997) The vesicle docking protein p115 binds GM130, a cis-Golgi matrix protein, in a mitotically regulated manner. Cell 89: 445–455 [DOI] [PubMed] [Google Scholar]
- Nichols BJ, Ungermann C, Pelham HR, Wickner WT, Haas A (1997) Homotypic vacuolar fusion mediated by t- and v-SNAREs. Nature 387: 199–202 [DOI] [PubMed] [Google Scholar]
- Oka T, Vasile E, Penman M, Novina CD, Dykxhoorn DM, Ungar D, Hughson FM, Krieger M (2005) Genetic analysis of the subunit organization and function of the conserved oligomeric golgi (COG) complex: studies of COG5- and COG7-deficient mammalian cells. J Biol Chem 280: 32736–32745 [DOI] [PubMed] [Google Scholar]
- Peng R, Gallwitz D (2002) Sly1 protein bound to Golgi syntaxin Sed5p allows assembly and contributes to specificity of SNARE fusion complexes. J Cell Biol 157: 645–655 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peterson MR, Emr SD (2001) The class C Vps complex functions at multiple stages of the vacuolar transport pathway. Traffic 2: 476–486 [DOI] [PubMed] [Google Scholar]
- Pevsner J, Hsu SC, Scheller RH (1994) n-Sec1: a neural-specific syntaxin-binding protein. Proc Natl Acad Sci USA 91: 1445–1449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeffer SR (2001) Rab GTPases: specifying and deciphering organelle identity and function. Trends Cell Biol 11: 487–491 [DOI] [PubMed] [Google Scholar]
- Poirier MA, Xiao W, Macosko JC, Chan C, Shin YK, Bennett MK (1998) The synaptic SNARE complex is a parallel four-stranded helical bundle. Nat Struct Biol 5: 765–769 [DOI] [PubMed] [Google Scholar]
- Price A, Seals D, Wickner W, Ungermann C (2000a) The docking stage of yeast vacuole fusion requires the transfer of proteins from a cis-SNARE complex to a Rab/Ypt protein. J Cell Biol 148: 1231–1238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price A, Wickner W, Ungermann C (2000b) Proteins needed for vesicle budding from the Golgi complex are also required for the docking step of homotypic vacuole fusion. J Cell Biol 148: 1223–1229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raymond CK, Howald-Stevenson I, Vater CA, Stevens TH (1992) Morphological classification of the yeast vacuolar protein sorting mutants: evidence for a prevacuolar compartment in class E vps mutants. Mol Biol Cell 3: 1389–1402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rehling P, Darsow T, Katzmann DJ, Emr SD (1999) Formation of AP-3 transport intermediates requires Vps41 function. Nat Cell Biol 1: 346–353 [DOI] [PubMed] [Google Scholar]
- Richardson SC, Winistorfer SC, Poupon V, Luzio JP, Piper RC (2004) Mammalian late vacuole protein sorting orthologues participate in early endosomal fusion and interact with the cytoskeleton. Mol Biol Cell 15: 1197–1210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rieder SE, Emr SD (1997) A novel RING finger protein complex essential for a late step in protein transport to the yeast vacuole. Mol Biol Cell 8: 2307–2327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenthal JA, Chen H, Slepnev VI, Pellegrini L, Salcini AE, Di Fiore PP, De Camilli P (1999) The epsins define a family of proteins that interact with components of the clathrin coat and contain a new protein module. J Biol Chem 274: 33959–33965 [DOI] [PubMed] [Google Scholar]
- Sacher M, Barrowman J, Wang W, Horecka J, Zhang Y, Pypaert M, Ferro-Novick S (2001) TRAPP I implicated in the specificity of tethering in ER-to-Golgi transport. Mol Cell 7: 433–442 [DOI] [PubMed] [Google Scholar]
- Sato TK, Rehling P, Peterson MR, Emr SD (2000) Class C Vps protein complex regulates vacuolar SNARE pairing and is required for vesicle docking/fusion. Mol Cell 6: 661–671 [DOI] [PubMed] [Google Scholar]
- Schultz J, Milpetz F, Bork P, Ponting CP (1998) SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci USA 95: 5857–5864 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seals DF, Eitzen G, Margolis N, Wickner WT, Price A (2000) A Ypt/Rab effector complex containing the Sec1 homolog Vps33p is required for homotypic vacuole fusion. Proc Natl Acad Sci USA 97: 9402–9407 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segev N (2001) Ypt and Rab GTPases: insight into functions through novel interactions. Curr Opin Cell Biol 13: 500–511 [DOI] [PubMed] [Google Scholar]
- Slusarewicz P, Xu Z, Seefeld K, Haas A, Wickner WT (1997) I2B is a small cytosolic protein that participates in vacuole fusion. Proc Natl Acad Sci USA 94: 5582–5587 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnichsen B, Lowe M, Levine T, Jamsa E, Dirac-Svejstrup B, Warren G (1998) A role for giantin in docking COPI vesicles to Golgi membranes. J Cell Biol 140: 1013–1021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srivastava A, Woolford CA, Jones EW (2000) Pep3p/Pep5p complex: a putative docking factor at multiple steps of vesicular transport to the vacuole of Saccharomyces cerevisiae. Genetics 156: 105–122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian S, Woolford CA, Jones EW (2004) The Sec1/Munc18 protein, Vps33p, functions at the endosome and the vacuole of Saccharomyces cerevisiae. Mol Biol Cell 15: 2593–2605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorngren N, Collins KM, Fratti RA, Wickner W, Merz AJ (2004) A soluble SNARE drives rapid docking, bypassing ATP and Sec17/18p for vacuole fusion. EMBO J 23: 2765–2776 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungermann C, Nichols BJ, Pelham HR, Wickner W (1998a) A vacuolar v-t-SNARE complex, the predominant form in vivo and on isolated vacuoles, is disassembled and activated for docking and fusion. J Cell Biol 140: 61–69 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungermann C, Sato K, Wickner W (1998b) Defining the functions of trans-SNARE pairs. Nature 396: 543–548 [DOI] [PubMed] [Google Scholar]
- Ungermann C, von Mollard GF, Jensen ON, Margolis N, Stevens TH, Wickner W (1999) Three v-SNAREs and two t-SNAREs, present in a pentameric cis-SNARE complex on isolated vacuoles, are essential for homotypic fusion. J Cell Biol 145: 1435–1442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Rossum DB, Patterson RL, Sharma S, Barrow RK, Kornberg M, Gill DL, Snyder SH (2005) Phospholipase Cγ1 controls surface expression of TRPC3 through an intermolecular PH domain. Nature 434: 99–104 [DOI] [PubMed] [Google Scholar]
- Walter DM, Paul KS, Waters MG (1998) Purification and characterization of a novel 13 S hetero-oligomeric protein complex that stimulates in vitro Golgi transport. J Biol Chem 273: 29565–29576 [DOI] [PubMed] [Google Scholar]
- Wang L, Merz AJ, Collins KM, Wickner W (2003) Hierarchy of protein assembly at the vertex ring domain for yeast vacuole docking and fusion. J Cell Biol 160: 365–374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Seeley ES, Wickner W, Merz AJ (2002) Vacuole fusion at a ring of vertex docking sites leaves membrane fragments within the organelle. Cell 108: 357–369 [DOI] [PubMed] [Google Scholar]
- Wang L, Ungermann C, Wickner W (2000) The docking of primed vacuoles can be reversibly arrested by excess Sec17p (α-SNAP). J Biol Chem 275: 22862–22867 [DOI] [PubMed] [Google Scholar]
- Weber T, Zemelman BV, McNew JA, Westermann B, Gmachl M, Parlati F, Sollner TH, Rothman JE (1998) SNAREpins: minimal machinery for membrane fusion. Cell 92: 759–772 [DOI] [PubMed] [Google Scholar]
- Weixel KM, Blumental-Perry A, Watkins SC, Aridor M, Weisz OA (2005) Distinct Golgi populations of phosphatidylinositol 4-phosphate regulated by phosphatidylinositol 4-kinases. J Biol Chem 280: 10501–10508 [DOI] [PubMed] [Google Scholar]
- Wickner W, Haas A (2000) Yeast homotypic vacuole fusion: a window on organelle trafficking mechanisms. Annu Rev Biochem 69: 247–275 [DOI] [PubMed] [Google Scholar]
- Will E, Albert S, Gallwitz D (2001) Expression, purification, and biochemical properties of Ypt/Rab GTPase-activating proteins of Gyp family. Methods Enzymol 329: 50–58 [DOI] [PubMed] [Google Scholar]
- Wurmser AE, Sato TK, Emr SD (2000) New component of the vacuolar class C-Vps complex couples nucleotide exchange on the Ypt7 GTPase to SNARE-dependent docking and fusion. J Cell Biol 151: 551–562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ybe JA, Brodsky FM, Hofmann K, Lin K, Liu SH, Chen L, Earnest TN, Fletterick RJ, Hwang PK (1999) Clathrin self-assembly is mediated by a tandemly repeated superhelix. Nature 399: 371–375 [DOI] [PubMed] [Google Scholar]
- Zheng N, Wang P, Jeffrey PD, Pavletich NP (2000) Structure of a c-Cbl-UbcH7 complex: RING domain function in ubiquitin-protein ligases. Cell 102: 533–539 [DOI] [PubMed] [Google Scholar]
- Zubenko GS, Mitchell AP, Jones EW (1980) Mapping of the proteinase b structural gene PRB1, in Saccharomyces cerevisiae and identification of nonsense alleles within the locus. Genetics 96: 137–146 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1
Figure S2
Figure S3
Figure S4