

Abstract
Several chemokines or chemokine receptors are involved in atherogenesis. CCR1 is expressed by macrophages and lymphocytes, two major cell types involved in the progression of atherosclerosis, and binds to lesion-expressed ligands. We examined the direct role of the blood-borne chemokine receptor CCR1 in atherosclerosis by transplanting bone marrow cells from either CCR1+/+ or CCR1−/− mice into low-density lipoprotein-receptor (LDLr)-deficient mice. After exposure to an atherogenic diet for 8 weeks, no differences in fatty streak size or composition were detected between the 2 groups. After 12 weeks of atherogenic diet, however, an unexpected 70% increase in atherosclerotic lesion size in the thoracic aorta was detected in the CCR1−/− mice, accompanied by a 37% increase in the aortic sinus lesion area. CCR1−/− mice showed enhanced basal and concanavalin A–stimulated IFN-γ production by spleen T cells and enhanced plaque inflammation. In conclusion, blood-borne CCR1 alters the immuno-inflammatory response in atherosclerosis and prevents excessive plaque growth and inflammation.
INTRODUCTION
Chemokines play key roles in the maturation, homing, and activation of leukocytes at sites of inflammation (1). Although various chemokines have been shown to be expressed in atherosclerotic arteries, so far only CCL2 (MCP-1) and CX3CL1 (Fractalkine) chemokines have been shown to play a direct role in atherosclerotic plaque development (2,3). Signaling through G protein–coupled chemokine receptors is responsible for chemokine effects on leukocyte development, recruitment, and activation (1). Three chemokine receptors, CCR2, CXCR2, and CX3CR1, appear to play key roles in leukocyte recruitment into the atherosclerotic vessels (4–7). Many other chemokine receptors have been identified, however, and may play different roles in atherosclerosis (8). CCR1 is a chemokine receptor for CCL3 and CCL5 and is expressed on macrophages and T cells (8), two major cell types involved in atherosclerosis (9). In this study, we examined the direct role of blood-borne CCR1 in the development and composition of atherosclerotic plaques in low-density lipoprotein receptor-deficient (LDLr−/−) mice. We chose the irradiation/bone marrow transplantation model because CCR1 is mainly expressed on bone marrow–derived cells that give rise to the circulating leukocytes involved in atherogenesis (10).
METHODS
Generation of Chimeric Mice
CCR1−/− mice on a C57BL/6J background were used (11). Female C57BL/6J LDLr−/− (Jackson Laboratory) were 7 months old. Mice were irradiated and reconstituted with CCR1+/+ or CCR1−/− (n = 10) bone marrow cells (5). After 4 weeks of recovery, mice were put on a diet containing 15% fat, 1.25% cholesterol, and 0% cholate for 8 weeks (n = 6 CCR1+/+; n = 7 CCR1−/−) to study fatty streak development or 12 weeks (n = 9 CCR1+/+; n = 10 CCR1−/−) to examine the development of large and advanced atherosclerotic plaques.
Assessment of the Extent of Atherosclerosis in Aortas and Aortic Sinus
Plasma total and HDL cholesterol levels were measured with a commercially available cholesterol kit (Sigma). Morphometric and immunohistochemical studies were performed in the aortic sinus and the thoracic aorta as previously described (12). Goat polyclonal antibodies against mouse CCL3 and CCR1 (Santa Cruz) were used at a dilution of 1:30 and 1:50, respectively, to assess CCL3 and CCR1 expression in atherosclerotic plaques. A goat anti-mouse IFN-γ antibody (R&D Systems) was used at 10 μg/mL, and a biotinylated EO6 antibody (provided by J. Witztum) that recognizes oxidized phospholipids was used at 8 μg/mL. Irrelevant immunoglobulins were used as negative controls. A semiquantitative score was established to compare IFN-γ staining between the two groups of mice. Maximal staining was scored as 3 and no staining as 0.
Cytokine Assays
Murine cytokines (IL-2, IL-4, IFNγ, and TNFα) were measured in supernatants of mouse splenocytes and in plasma using the mouse Th1/Th2 cytokine cytometric bead array kit according to the manufacturer’s instructions (BD Biosciences).
RT-Multiplex PCR
Total RNA was extracted from splenocytes using the QIAamp RNA Blood Mini kit (Qiagen, Courtaboeuf, France) and cDNA generated with Superscript IITM Rnase H (Invitrogen, Carlsbad, CA, USA) following standard procedures. Amplification of the chemokine receptor transcripts was performed with kits for detecting the transcripts coding for the mouse CCR receptors CCR1, CCR2, CCR3, CCR4, and CCR5 and GAPDH (BioSource Europe) as previously described (13).
Statistical Analysis
Data are expressed as means ± SE. Statistical significance was determined by ANOVA. A value of P < 0.05 was considered to be statistically significant.
RESULTS
Animal weights (not shown), plasma total cholesterol (14.1 ± 0.6 g/L vs. 14.6 ± 0.5 at 8 weeks and 13.3 ± 0.9 g/L vs. 14.6 ± 1.3 g/L at 12 weeks in CCR1+/+ and CCR1−/− mice g/L, respectively), and HDL cholesterol levels (not shown) were similar in the two groups. We verified the success of bone marrow transplantation using RT-PCR on splenocytes at the time mice were killed, 12 weeks after the high-fat diet. CCR1 transcripts were detected in the spleens of mice reconstituted with CCR1+/+ bone marrow but were absent from the spleens of mice reconstituted with CCR1−/− bone marrow (Figure 1a). Other chemokine receptors showed similar expression between the two groups of mice (Figure 1a). CCR1 protein expression was assessed using immunohistochemistry and was detectable in macrophage-rich plaque areas of CCR1+/+ mice but was absent in CCR1−/− mice (Figure 1c). CCL3, a major ligand for CCR1, was detectable in both CCR1+/+ and CCR1−/− plaques (Figure 1b and data not shown). We found no difference in fatty streak size (134,501 ± 22,850 μm2 vs. 162,197 ± 17,718 μm2, in CCR1−/− and CCR1+/+ mice, respectively, P = 0.4) or composition (data not shown) between the two groups after 8 weeks of atherogenic diet. After 12 weeks of atherogenic diet, however, we found an unexpected 70% increase in the extent of surface area stained with oil red O in thoracic aortas of CCR1−/− mice compared with controls (16.86 ± 2.91% vs. 9.89 ± 0.98%, respectively, P < 0.05) (Figure 2). These results were confirmed and extended in analyses from the aortic sinus showing a highly significant 37% increase in atherosclerotic plaque area in CCR1−/− mice compared with controls (573,985 ± 41,074 μm2 vs. 418,126 ± 26,053 μm2, respectively, P = 0.006) (Figure 3). The effects on plaque size were associated with a significant 30% increase in macrophage infiltration (MOMA-2 staining) in CCR1−/− mice (48.1 ± 4.7% of plaque area vs. 37.4 ± 1.0%, respectively, P < 0.05) (Figures 4a and b) and a persistent infiltration of lesions by T cells (89.11 ± 13.35 CD3+ cells/mm2 of plaque area in CCR1−/− mice vs. 77.63 ± 14.25 CD3+ cells/mm2 in CCR1+/+ mice, P = 0.5) (Figures 4c and d). Interestingly, atherosclerotic plaques of CCR1−/− mice showed a substantially lower accumulation of smooth muscle cells (1.5 ± 0.5% vs. 6.2 ± 1.8%, respectively, P < 0.05) (Figures 5a and b) and collagen (16.3 ± 2.1% vs. 23.1 ± 2.7%, respectively, P = 0.06) in comparison with controls (Figures 5c and d), suggesting reduced plaque healing. No differences were found in Ki-67 or TUNEL staining between the two groups (data not shown). Thus, CCR1 deficiency in bone marrow cells enhances the development of atherosclerotic lesions and promotes an inflammatory and unstable plaque phenotype.
Figure 1.

(a) Chemokine receptor expression after bone marrow reconstitution of irradiated mice. Expression of CCR1 transcripts and other CC chemokine receptors was followed by multiplex RT-PCR in splenocytes of LDLr−/− mice reconstituted with bone marrow from either CCR1−/− or control CCR1+/+ mice. The data are representative of 3 mice in each group. (b) Representative photomicrograph showing specific CCL3 staining (red) in a macrophage-rich plaque from CCR1−/− mice. Similar staining was observed in CCR1+/+ plaques (not shown). (c) Representative positive staining (red) for CCR1 protein in macrophage-rich plaques obtained from CCR1+/+ mice and its absence in plaques of CCR1−/− mice. Original magnifications × 200 (b), × 100 (c).
Figure 2.
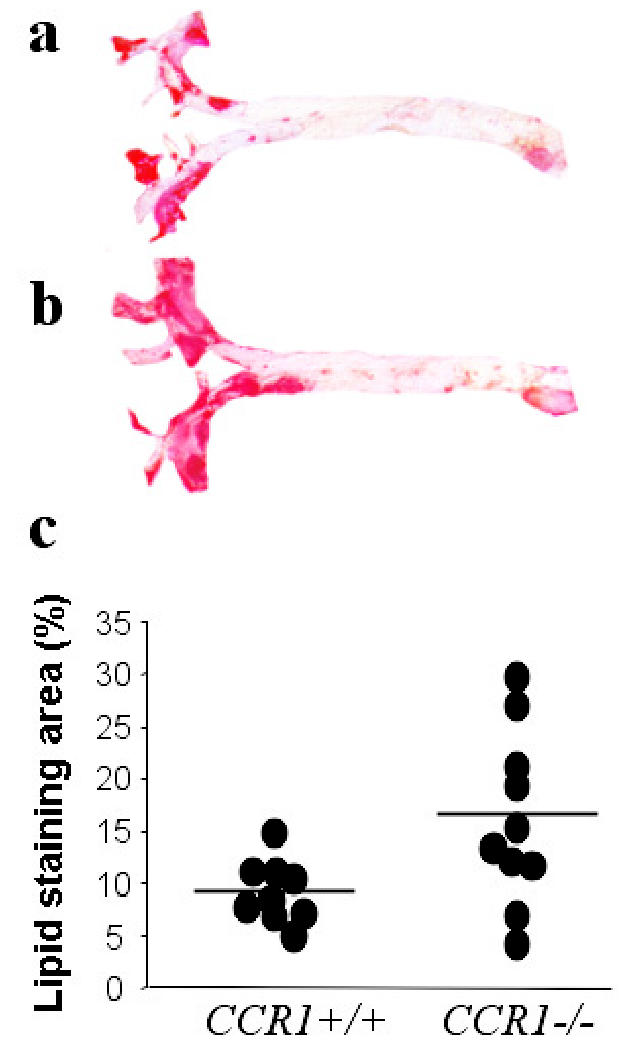
Representative photomicrographs showing oil red O staining in the thoracic aorta of CCR1+/+ (a) and CCR1−/− (b) mice. Using computer-assisted quantitative analysis (c), we found a 70% increase in the percentage of lipid area in CCR1−/− mice.
Figure 3.

Representative photomicrographs showing oil red O staining in transversal sections from the aortic sinus of CCR1+/+ (a) and CCR1−/− (b) mice. Using computer-assisted quantitative analysis (c), we found a 37% increase in plaque size in CCR1−/− mice. Original magnifications × 40.
Figure 4.

Representative photomicrographs showing immunohistochemical stainings for MOMA-2 (macrophage) (dark red in a, b) and CD3 (lymphocytes) (red dots in c, d) in transversal sections from the aortic sinus of CCR1+/+ (a, c) and CCR1−/− (b, d) mice. We found a 30% increase in macrophage content and a persistent substantial accumulation of T cells in lesions of CCR1−/− mice. Original magnifications × 200.
Figure 5.

Representative photomicrographs showing immunostaining for α-actin (smooth muscle cells) (dark blue in a, b) and staining using Sirius red (collagen) (red in c, d) in transversal sections from the aortic sinus of CCR1+/+ (a, c) and CCR1−/− (b, d) mice. We found a 76% decrease in smooth muscle cell accumulation and a 30% decrease in collagen content in lesions of CCR1−/− mice. Original magnifications × 200.
To obtain additional insights into the potential mechanisms of increased lesion formation and inflammation in the presence of CCR1 deficiency, we assessed the immuno-inflammatory response in CCR1+/+ and CCR1−/− mice. Atherosclerosis is an immuno-inflammatory disease that develops in response to lipid oxidation (9), and systemic activation of immuno-inflammatory cells in murine models correctly reflects cell activation within the lesions (14,15). We studied the spleen cytokine profile of mice killed after 12 weeks of atherogenic diet after stimulation with concanavalin A, which promotes T lymphocyte activation. As expected, the spleen T cell cytokine profile in wild-type mice was clearly of the Th1 type, with high levels of IFN-γ (3.8 ± 0.7 ng/mL; n = 3) and very low levels of IL-4 (50.5 ± 6.5 pg/mL; n = 3). CCR1 deficiency resulted in a more powerful Th1 response as revealed by the substantial increase in IFN-γ production (9.7 ± 0.4 ng/mL, P = 0.01) and the persistently low levels of IL-4 (43.0 ± 5.0 pg/mL). We also found a significant increase in IFN-γ staining in vivo in spleen cryostat sections collected from CCR1−/− mice after 12 weeks of high-fat diet compared with wild-type mice (immunohistochemical score: 1.74 ± 0.44 vs. 0.53 ± 0.22, respectively, n = 5; P < 0.05). Interestingly, IFN-γ staining in spleen sections was exclusively detected in areas showing accumulation of oxidized lipids (Figure 6), suggesting an adaptive immune response against modified lipoproteins. We found no difference in spleen CD3, CD4, or CD8 staining between the two groups of mice. Finally, no difference in basal circulating plasma levels of IFN-γ, IL-4, or TNF-α was found between the two groups (n = 4; data not shown).
Figure 6.

Representative photomicrographs showing immunostaining for oxidized phospholipids recognized by a biotinylated EO6 antibody (a), for CD3 (T lymphocytes in b), and IFN-γ (c, d) in spleen sections from the two groups of mice at the time of death. Staining with EO6 and CD3 is shown for CCR1−/− mice and was highly similar to staining observed in CCR1+/+ mice. Areas showing accumulation positive EO6 staining also showed positive staining for IFN-γ. IFN-γ staining was much more pronounced in CCR1−/− (d) than in CCR1+/+ (c) mice.
DISCUSSION
The chemokines CCL3 and CCL5 are expressed in atherosclerotic lesions (16) and are thought to be major candidates for leukocyte recruitment into diseased arteries (8). CCL3 and CCL5 have two major receptors, CCR1 and CCR5 (1). A recent study has shown that CCR5 deficiency does not affect early atherosclerotic lesion development in apoE knockout mice (17). Thus, we hypothesized that deficiency in CCR1 expression may alter CCL3- and/or CCL5-induced monocyte recruitment and lead to reduction in lesion formation. Unexpectedly, we found that CCR1 deficiency in circulating leukocytes did not affect early fatty streak development, but rather promoted the progression toward larger and inflammatory atherosclerotic plaques at two different atherosclerosis-prone sites. These results suggest a novel and important role for CCR1 in the protection against excessive plaque growth.
Examining plaque composition, we found a significant and unexpected increase in the accumulation of macrophages within lesions of CCR1−/− mice after 12 weeks of atherogenic diet, suggesting that these cells were recruited through at least one of the other chemokine/chemokine receptor pathways previously reported to play a key role in leukocyte trafficking during this disease. At the advanced stages of lesion development, T lymphocyte accumulation within the lesion gradually decreases (14) as the plaque phenotype switches from an inflammatory toward a healing stage. However, despite the increase of plaque size in CCR1−/− mice, we found a persistent T cell accumulation within the lesions, testifying to the important immune activation in the absence of CCR1. This activated state was supported by our finding of enhanced conA-induced IFN-γ production in splenocytes of CCR1−/− mice and a lipid-associated increase in IFN-γ staining in spleen sections. Taken together, our results argue for an enhanced systemic Th1 immune response initiated by oxidized lipids that may have contributed to the exaggerated plaque inflammation and to the reduction in smooth muscle cell and collagen accumulation in advanced lesions of CCR1−/− mice.
Finally, total-body irradiation per se has been shown to differentially affect plaque size at two different atherosclerosis-prone sites (18). However, this is unlikely to affect our interpretation of the results, because both groups of mice received total-body irradiation. Moreover, differences in atherosclerotic lesion size were observed at the level of both the aortic sinus and the thoracic aorta.
In conclusion, we showed that rather than preventing leukocyte recruitment in atherosclerosis, the absence of CCR1 expression in leukocytes led to exaggerated IFN-γ production and promoted both lesion progression and inflammation, revealing novel roles for chemokine receptors in this complex disease.
Acknowledgments
This study was supported by the “Nouvelle Société Française d’Athérosclérose” (S.P.) and the “Fondation de France” (C.C.). We are grateful to Joseph Witztum and La Jolla SCOR program (Department of Medicine, University of California San Diego, La Jolla, CA, USA) for providing EO6 antibody.
Footnotes
Online address: http://www.molmed.org
REFERENCES
- 1.Rossi D, Zlotnik A. The biology of chemokines and their receptors. Annu Rev Immunol. 2000;18:217–42. doi: 10.1146/annurev.immunol.18.1.217. [DOI] [PubMed] [Google Scholar]
- 2.Gu L, et al. Absence of monocyte chemoattractant protein-1 reduces atherosclerosis in low density lipoprotein receptor-deficient mice. Mol Cell. 1998;2:275–81. doi: 10.1016/s1097-2765(00)80139-2. [DOI] [PubMed] [Google Scholar]
- 3.Teupser D, et al. Major reduction of atherosclerosis in fractalkine (CX3CL1)-deficient mice is at the brachiocephalic artery, not the aortic root. Proc Natl Acad Sci USA. 2004;101:17795–800. doi: 10.1073/pnas.0408096101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Boring L, Gosling J, Cleary M, Charo IF. Decreased lesion formation in CCR2−/− mice reveals a role for chemokines in the initiation of atherosclerosis. Nature. 1998;394:894–7. doi: 10.1038/29788. [DOI] [PubMed] [Google Scholar]
- 5.Boisvert WA, Santiago R, Curtiss LK, Terkeltaub RA. A leukocyte homologue of the IL-8 receptor CXCR-2 mediates the accumulation of macrophages in atherosclerotic lesions of LDL receptor-deficient mice. J Clin Invest. 1998;101:353–63. doi: 10.1172/JCI1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Combadiere C, et al. Decreased atherosclerotic lesion formation in CX3CR1/apolipoprotein E double knockout mice. Circulation. 2003;107:1009–16. doi: 10.1161/01.cir.0000057548.68243.42. [DOI] [PubMed] [Google Scholar]
- 7.Lesnik P, Haskell CA, Charo IF. Decreased atherosclerosis in CX3CR1−/− mice reveals a role for fractalkine in atherogenesis. J Clin Invest. 2003;111:333–40. doi: 10.1172/JCI15555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Charo IF, Taubman MB. Chemokines in the pathogenesis of vascular disease. Circ Res. 2004;95:858–66. doi: 10.1161/01.RES.0000146672.10582.17. [DOI] [PubMed] [Google Scholar]
- 9.Hansson GK. Inflammation, atherosclerosis, and coronary artery disease. N Engl J Med. 2005;352:1685–95. doi: 10.1056/NEJMra043430. [DOI] [PubMed] [Google Scholar]
- 10.Sata M, et al. Hematopoietic stem cells differentiate into vascular cells that participate in the pathogenesis of atherosclerosis. Nat Med. 2002;8:403–9. doi: 10.1038/nm0402-403. [DOI] [PubMed] [Google Scholar]
- 11.Gao JL, et al. Impaired host defense, hematopoiesis, granulomatous inflammation and type 1-type 2 cytokine balance in mice lacking CC chemokine receptor 1. J Exp Med. 1997;185:1959–68. doi: 10.1084/jem.185.11.1959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mallat Z, et al. Interleukin-18/interleukin-18 binding protein signaling modulates atherosclerotic lesion development and stability. Circ Res. 2001;89:E41–5. doi: 10.1161/hh1901.098735. [DOI] [PubMed] [Google Scholar]
- 13.Nitcheu J, et al. Perforin-dependent brain-infiltrating cytotoxic CD8+ T lymphocytes mediate experimental cerebral malaria pathogenesis. J Immunol. 2003;170:2221–8. doi: 10.4049/jimmunol.170.4.2221. [DOI] [PubMed] [Google Scholar]
- 14.Pinderski LJ, et al. Overexpression of interleukin-10 by activated T lymphocytes inhibits atherosclerosis in LDL receptor-deficient mice by altering lymphocyte and macrophage phenotypes. Circ Res. 2002;90:1064–71. doi: 10.1161/01.res.0000018941.10726.fa. [DOI] [PubMed] [Google Scholar]
- 15.Zhou X, Paulsson G, Stemme S, Hansson GK. Hypercholesterolemia is associated with a T helper (Th) 1/Th2 switch of the autoimmune response in atherosclerotic apo E-knockout mice. J Clin Invest. 1998;101:1717–25. doi: 10.1172/JCI1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Reape TJ, Groot PH. Chemokines and atherosclerosis. Atherosclerosis. 1999;147:213–25. doi: 10.1016/s0021-9150(99)00346-9. [DOI] [PubMed] [Google Scholar]
- 17.Kuziel WA, et al. CCR5 deficiency is not protective in the early stages of atherogenesis in apoE knockout mice. Atherosclerosis. 2003;167:25–32. doi: 10.1016/s0021-9150(02)00382-9. [DOI] [PubMed] [Google Scholar]
- 18.Schiller NK, Kubo N, Boisvert WA, Curtiss LK. Effect of gamma-irradiation and bone marrow transplantation on atherosclerosis in LDL receptor-deficient mice. Arterioscler Thromb Vasc Biol. 2001;21:1674–80. doi: 10.1161/hq1001.096724. [DOI] [PubMed] [Google Scholar]