

Abstract
Endogenous 2-arachidonoylglycerol (2-AG) inhibits invasion of androgen-independent prostate cancer cells. Blocking cellular hydrolysis of 2-AG to increase its endogenous concentration results in a decrease in cell invasion. A series of compounds containing a trifluoromethyl ketone (TFK) moiety or the methyl analog (known to inhibit carboxylesterases) were investigated for their ability to inhibit 2-AG hydrolysis and prostate cancer cell invasion. Compounds containing a thioether β to a TFK moiety inhibited 2-AG hydrolysis as well as cell invasion in a concentration-dependent manner. Inhibition of 2-AG hydrolysis increased concomitantly with inhibitor alkyl chain length from 4- to 12-carbons while inhibition of cell invasion exhibited a maximum at 8- to 10-carbons of the compounds. These results demonstrate a new series of 2-AG hydrolysis inhibitors as a potential therapeutic approach for prostate cancer.
Keywords: 2-Arachidonoylglycerol, Cell invasion, Endocannabinoid, Carboxylesterase inhibitors, Prostate cancer
2-Arachidonoylglycerol (2-AG), an endocannabinoid, is an endogenous ligand for the central cannabinoid (CB1) receptors [1] and for the peripheral cannabinoid (CB2) receptors in some cells [2]. It is synthesized from arachidonic acid (AA)-enriched membrane phospholipids [3,4] with the final-step metabolism of diacylglycerol by diacylglycerol lipase [5]. 2-AG can be hydrolyzed to AA and glycerol by two characterized enzymes, fatty acid amide hydrolase (FAAH) [6-9], and monoacylglycerol lipase (MGL) [10-12]. This hydrolysis is a terminating step to end the actions or functions of 2-AG [12]. 2-AG is endogenously produced at high concentrations in androgen-independent prostate cancer (PC-3, DU-145) cells and it possesses an anti-invasive effect in these prostate cancer cells through a CB1-dependent pathway [13]. Blocking 2-AG synthesis to reduce its endogenous concentration results in an increase in cell invasion. On the other hand, maintaining and/or increasing endogenous 2-AG concentrations by inhibiting 2-AG hydrolysis results in a decrease in cell invasion [13]. These results suggest that inhibitors of enzymes involved in the degradation of 2-AG could be of importance in the regulation of prostate cancer cell invasion. In this study, a new series of compounds were examined for their ability to inhibit 2-AG hydrolysis and invasion in PC-3 and DU-145 cells. This group of small molecule inhibitors was rationally selected to examine their physicochemical properties in 2-AG hydrolysis inhibition. They included some of the most potent carboxylesterase inhibitors identified to date [14-19].
Materials and methods
Materials
Carboxylesterase inhibitors (Fig. 1) were synthesized as previously described [16,17]. Androgen-independent prostate cancer (PC-3 and DU-145) cells were obtained from the American Type Culture Collection (ATCC, Rockville, Maryland). 2-Arachidonoylglycerol (2-AG) and [2H8]2-AG were obtained from Cayman Chemical (Ann Arbor, Michigan). [methyl-3H]Thymidine (1 μCi/ml) was obtained from Applied Biosystems (Foster City, California). SR141716 (SR-1) was obtained from Research Triangle Institute (Research Triangle Park, North Carolina). 2-Oleoyl-[3H]glycerol (2-OG, 20 Ci/mmol) was obtained from American Radiolabeled Chemical (St Louis, Missouri). A positive control for caveolin-1, non-stimulated A431 cell lysate (human carcinoma cell line), and monoclonal anti-caveolin-1 were obtained from Upstate (Lake Placid, New York). Secondary antibody, goat anti-mouse IgG-HRP was obtained from Zymed Laboratories (South San Francisco, California). Transwells with 8.0 μm pore polyvinylpyrrolidone-free polycarbonate filters were obtained from Corning (Corning, New York). Matrigel was obtained from BD Biosciences (Bedford, Massachusetts). All other chemicals and solvents were of analytical or highest purity grades. Distilled, deionized water was used in all experiments.
Fig. 1.
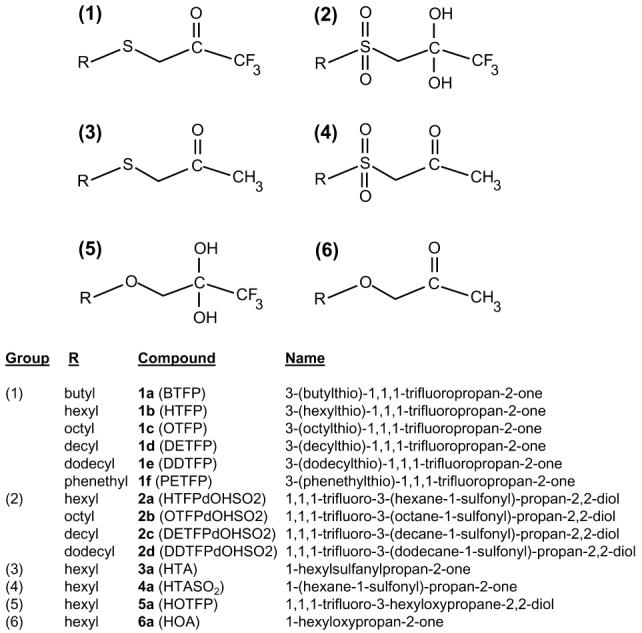
Carboxylesterase inhibitors used in this study.
Cell culture
Cells were maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum, l-glutamine (2 mM), streptomycin (100 μg/ml), and penicillin (100 U/ml). Cells were grown in 75 cm2 polystyrene tissue culture flasks at 37 °C in 5% CO2 in air to about 60–70% confluency before use.
Cell invasion assay
Cell invasion was determined as previously described [13]. Briefly, cells were incubated overnight with [methyl-3H]thymidine (1 μCi/ml) in the medium containing 10% fetal bovine serum, rinsed with fresh complete media to remove unbound [methyl-3H]thymidine. Cells were detached, centrifuged, and resuspended in 3 ml of serum-free RPMI. The experiments were performed in 24-well plates containing Transwells with 8.0 μm pore polyvinylpyrrolidone-free polycarbonate filters coated with Matrigel on the top compartments. A cell suspension of 50,000 cells containing the vehicle control or carboxylesterase inhibitors and other pharmacological agents at various concentrations was added to each upper compartment. Fibroblast-conditioned media (400 μl) were added in the bottom compartment of the well as a chemoattractant. Cells were incubated at 37 °C in the incubator for 5 h. The cells passed into the lower compartment media were detached with Pucks buffer containing 0.75% trypsin, placed in vials, and counted for radioactivity. The invasion was reported as the percentage of the invasion of the control cells.
Determination of inhibition of 2-oleoylglycerol (2-OG) hydrolysis by carboxylesterase inhibitors
2-Oleoyl-[3H]glycerol (2-OG) was used as a substrate for determining hydrolase activity. The hydrolysis of 2-OG by cytosolic and membrane proteins was determined as previously described [11,13]. Briefly, cells were grown to 60–70% confluence, washed with PBS buffer, and homogenized by pulling through a 27G needle a few times. The membrane and cytosolic proteins were separated by centrifugation. Protein concentration was measured by Bio-Rad Protein Assay (Bio-Rad, Hercules, California). The membrane protein pellet was resuspended in 150 μl PBS buffer (100 mM phosphate-buffered saline, pH 7.4). The 0.5 ml assay solution consisting of PBS buffer, pH 7.4, containing 100 μg proteins in the absence and presence of inhibitors at various concentrations and 2-OG (20 Ci/mmol) at 10,000 dpm were incubated at 37 °C for 30 min. The reaction was stopped by the addition of 2.0 ml of 1:2 chloroform:methanol and vortexed. Next, 0.60 ml of water was added to the reaction mixture, followed by vortexing and extraction with 0.67 ml chloroform. The radioactivity in the organic phase represents the unreacted 2-OG and the radioactivity in the aqueous phase represents the hydrolyzed glycerol. The hydrolysis was normalized to the percentage of the control (100%).
Determination of 2-AG in prostate cancer cells by liquid chromatography-electrospray ionization-mass spectrometry (LC-ESI-MS)
Cells were grown in T-75 flasks, rinsed with 5 ml Hepes buffer (containing 20 mM Hepes, 130 mM NaCl, 5 mM KCl, 1 mM CaCl2,2mM MgCl2, and 30 mM glucose, pH 7.4), treated with inhibitors or vehicle in Hepes buffer at 37 °C for 15 min. The cells were lyzed, scraped, and [2H8]2-AG was added as an internal standard. Samples were extracted by solid phase extraction as previously described [20], redissolved in 20 μl acetonitrile, and analyzed or kept at −80 °C. Samples were analyzed by using LC-ESI-MS (Agilent 1100 LC-MSD, SL model) as previously described [20]. The detection was made in the positive ion mode. For quantitative measurement, the m/z 379 and 387 were used for 2-AG and [2H8]2-AG, respectively. The concentrations of 2-AG were calculated by comparing their ratios of peak areas to the standard curves. The results were normalized to the protein content.
Western blot analysis of caveolin-1
To demonstrate the separation of membrane proteins from the cytosolic fraction of PC-3 cells, the caveolin-1 (a membrane protein) was determined by Western blot analysis. Cells were lyzed as described above in the presence of a protease inhibitor. Membrane and cytosolic proteins (50 μg), and lysate of human A431 carcinoma cells (a positive control) were separated by SDS–PAGE (Ready Gels) and transferred to a 0.7-μm nitrocellu-lose (Bio-Rad, Hercules, California) membrane. Proteins were incubated with the monoclonal anti-caveolin-1 antibody at 1:500 dilution. Then, goat anti-mouse IgG-HRP at 1:5000 dilution was used to complex with the primary antibody. The detection was made by using Western Lightning Chemiluminescence Reagent (Perkin-Elmer, Boston, Massachusetts) and captured by Fuji film X-ray (Tokyo, Japan).
Results and discussion
Hydrolysis of 2-oleoylglycerol
Exogenously added 2-AG to prostate cancer cells did not significantly inhibit cell invasion possibly due to its rapid metabolism to AA and glycerol (data not shown). Two characterized hydrolases, FAAH, and MGL [6-12], metabolize 2-AG to AA and glycerol. To investigate 2-AG hydrolysis, the hydrolysis of 2-oleoyl-[3H]glycerol (2-OG) (a readily available radiolabeled analog of 2-AG) by cytosolic and membrane proteins of PC-3 cells was determined. Several groups of inhibitors known to inhibit carboxylesterases (see Fig. 1) were tested as potential inhibitors of 2-OG (or 2-AG) hydrolysis. Only the compounds containing a thioether β to a TFK moiety (group 1) inhibited the 2-OG hydrolysis in the cytosolic fraction (Fig. 2A). The inhibition increased with the length of the carbon chain from HTFP (1b) < OTFP (1c) < DETFP (1d) < DDTFP (1e) (92.4 ± 3.3%, 71.8 ± 5.4%, 67.7 ± 3.3%, and 56.3 ± 4.1% of control, respectively). Inhibitors with a short carbon chain length such as BTFP (1a) or a bulky group such as PETFP (1f) did not exhibit any inhibitory effect on 2-OG hydrolysis. Only the long chain analog of these compounds, DDTFP (1e), slightly inhibited the hydrolysis of 2-OG in the membrane fraction (87.71 ± 1.95% of control) (Fig. 2B). All other compounds examined did not inhibit 2-OG hydrolysis by cytosolic or membrane proteins (data not shown). To illustrate that membrane proteins were separated from the cytosolic fraction, the caveolin-1 was analyzed in both fractions of PC-3 cells using human caveolin-1 as a positive control. Western blot shows an intense immunoreactive band in the membrane fraction which corresponds to the caveolin-1 positive control (25 kDa) but a very faint band in the cytosolic fraction (Fig. 2C), indicating a relatively good separation. More importantly, the hydrolysis inhibition of the enzyme(s) was observed in the cytosolic fraction and not the membrane fraction. This indicates that the incomplete separation of the membrane enzyme(s) should not account for the hydrolysis inhibition in the cytosolic fraction.
Fig. 2.
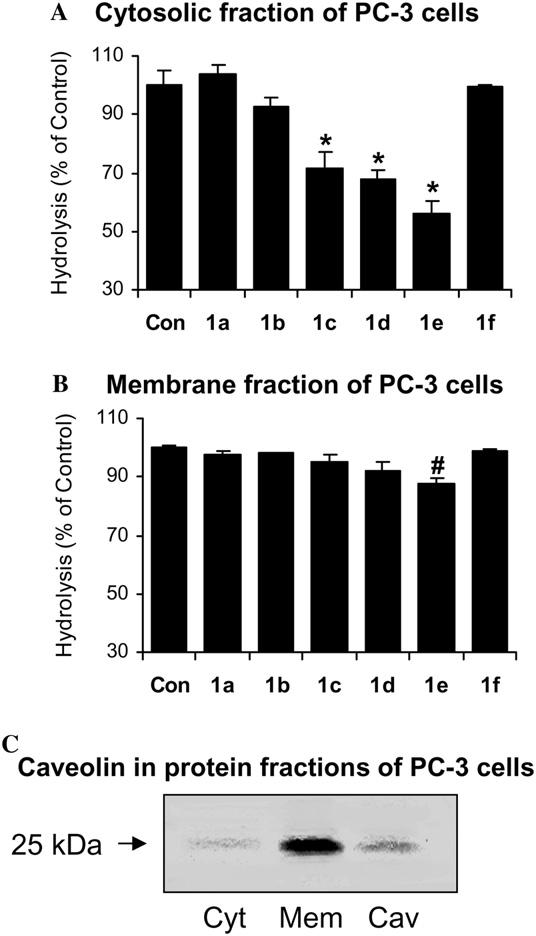
Inhibition of 2-oleoyl-[3H]glycerol (2-OG) hydrolysis by carboxylesterase inhibitors (1 μM) in cytosolic and membrane fractions of PC-3 cells. (A) Inhibition of 2-OG hydrolysis in cytosolic fraction by β-thioether trifluoromethyl ketone analogs (group 1) as compared with control. Values are means ± SEM (n = 6). (B) Inhibition of 2-OG hydrolysis in membrane fraction by β-thioether trifluoromethyl ketone analogs (group 1) as compared with control. Values are means ± SEM (n = 3). *Significantly different from control with p < 0.005 and #significantly different from control with p < 0.05. (C) Immunoreactive bands of proteins from cytosolic (lane 1) and membrane (lane 2) fractions of PC-3 cells and caveolin-1 (membrane protein as a positive control, lane 3) correspond to the molecular weight markers of 25 kDa.
Inhibitors containing a TFK moiety have been shown to inhibit FAAH with aliphatic TFK-containing inhibitors (oleamide analogs) exhibiting nanomolar Ki values [21,22]. The inhibition of 2-OG hydrolysis observed following treatment with β-thioether-containing TFK carboxylesterase inhibitors is different from that resulting from studies with FAAH and MGL inhibitors such as methylarachidonyl fluorophosphonate (MAFP) and diazomethylarachidonyl ketone (DAK). The former compound is thought to phosphorylate the enzyme with fluorine as the leaving group while DAK is a reactive photolabel. We previously found that PC-3 and DU-145 cells exhibited similar 2-OG hydrolysis in both membrane and cytosolic fractions, and MAFP and DAK inhibited the hydrolysis of 2-OG or 2-AG in both fractions [13]. These results suggest that enzymes hydrolyzing 2-AG in cytosolic fraction are probably different from the enzymes in the membrane fraction. The results also indicate that carboxylesterase inhibitors may be more specific inhibitors of cytosolic hydrolases.
Effects of carboxylesterase inhibitors on 2-AG concentration
Prostate cancer PC-3 cells produced 2-AG at high concentrations and treatment with the β-thioether-containing TFKs resulted in increased 2-AG concentrations as a function of inhibitor carbon chain length (Fig. 3A). The highest concentration of endogenous 2-AG was obtained with compound DETFP (1d), which incorporated a decyl alkyl chain attached to the thioether. In contrast, the greatest inhibition of 2-OG hydrolysis occurred with the inhibitor with the dodecyl analog DDTFP (1e) (Fig. 2A). This difference in activity may be attributed to different substrates as the hydrolysis of 2-OG was determined in the cytosol, and the 2-AG concentration was determined in intact cells. Inhibitors with increased chain length have greater lipophilicity and would be expected to have greater ability to diffuse through cellular membranes. However, the difference in activity of inhibitors may also be attributed to the uptake efficiency of the inhibitors into the cells which is limited by the optimal carbon chain length of the molecules. Other inhibitors which did not inhibit 2-OG hydrolysis such as HTFP (1b), HOA (6a) and OTFPOHSO2 (2b) also did not increase the cellular concentration of 2-AG.
Fig. 3.
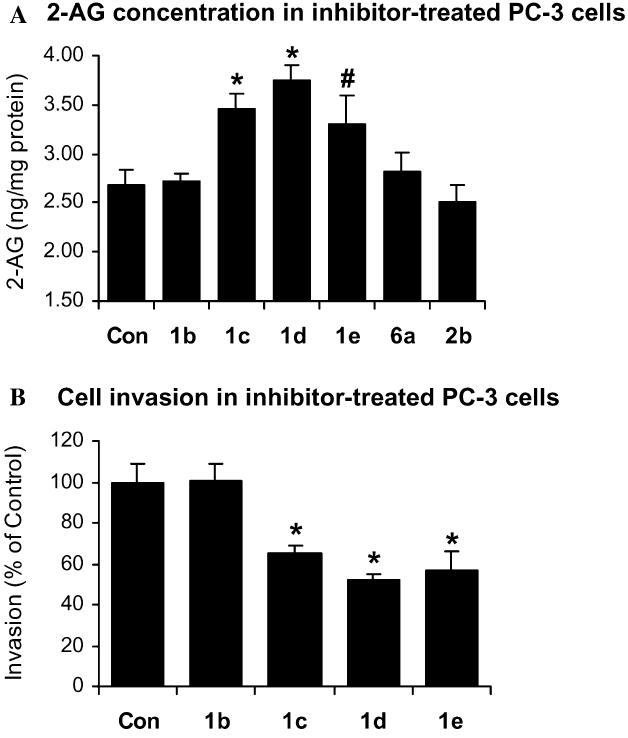
Effects of carboxylesterase inhibitors (1 μM) on 2-AG concentrations and cell invasion of PC-3 cells. (A) 2-AG concentrations in PC-3 cells treated with 1 μM of compounds HTFP (1b), OTFP (1c), DETFP (1d), DDTFP (1e), HOA (6a), and OTFPOHSO2 (2b). Values are means ± SEM (n = 3). (B) Cell invasion of PC-3 cells treated with compounds HTFP (1b), OTFP (1c), DETFP (1d), and DDTFP (1e). Cell invasion was normalized to the control cells (as 100%). Values are means ± SEM (n = 12). *Significantly different from control with p < 0.005 and #significantly different from control with p < 0.01.
Effects of inhibition of 2-AG hydrolysis on cell invasion
PC-3 cells were incubated with the β-thioether-containing TFKs to inhibit 2-AG hydrolysis, and cell invasion was measured. The inhibitors OTFP (1c), DETFP (1d), and DDTFP (1e) inhibited cell invasion (64.77 ± 4.34%, 52.00 ± 3.30%, and 56.96 ± 9.24% of control, respectively) (Fig. 3B). The inhibition by these compounds correlates well with the increase in cellular 2-AG concentrations in treated PC-3 cells. Again, DDTFP (1e) did not exhibit the inhibition of cell invasion greater than DETFP (1d). Other compounds such as HTFP (1b) (Fig. 3B) which did not increase cellular 2-AG concentration did not significantly inhibit cell invasion of PC-3 cells.
When DU-145 cells were incubated with OTFP (1c) and DETFP (1d) at different concentrations, cell invasion decreased in a concentration-dependent manner (Figs. 4A and B). OTFP also significantly inhibited cell invasion of PC-3 cells in a concentration-dependent manner (Fig. 4C). Exogenously added 2-AG did not decrease invasion of PC-3 cells [13] but it augmented the inhibition of cell invasion by OTFP (1.0 μM) (Fig. 4C). These results indicate that the blockade of 2-AG hydrolysis inhibits cell invasion and further supports the hypothesis that endogenous 2-AG is a negative regulator of invasion in androgen-independent prostate cancer cells. Furthermore, SR141716 (SR-1), a selective CB1 antagonist, at 500 nM increases invasion of PC-3 cells [13]. SR141716 reversed the inhibition of cell invasion by OTFP (1.0 μM) (Fig. 4C). These results further indicate that 2-AG is an endogenous inhibitor of prostate cancer cell invasion and its inhibition is CB1-receptor dependent as previously reported [13].
Fig. 4.
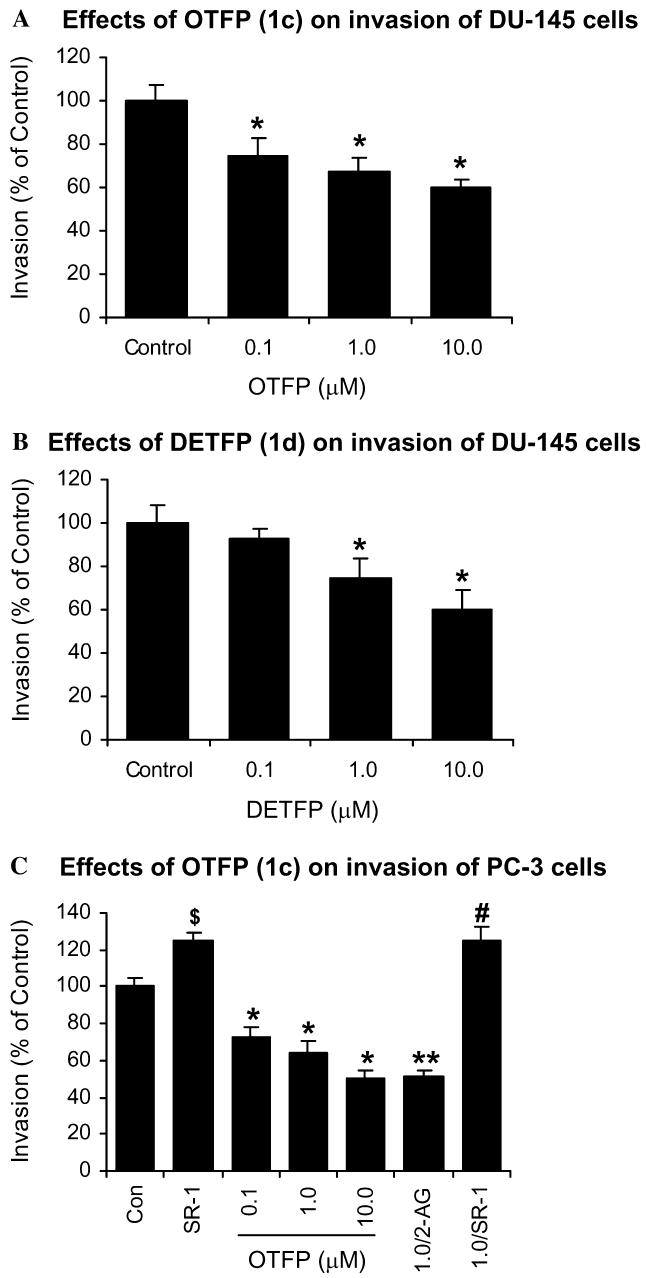
Invasion of PC-3 and DU-145 cells. (A) Cell invasion of DU-145 cells treated with OTFP (1c) (0.1, 1.0, and 10.0 μM). Values are means ± SEM (n = 12). (B) Cell invasion of DU-145 cells treated with DETFP (1d) (0.1, 1.0, and 10.0 μM). Values are means ± SEM (n = 12). (C) Cell invasion of PC-3 cells treated with SR141716 (SR-1) (500 nM), OTFP (0.1, 1.0, and 10.0 μM), OTFP (1.0 μM) + 2-AG (1.0 μM), and OTFP (1.0 μM) + SR141716 (SR-1) (500 nM). Values are means ± SEM (n = 18). $Significantly higher than control with p < 0.05; *significantly lower than control with p < 0.01; **significantly lower than OTFP (1.0 μM) with p < 0.05; #significantly higher than OTFP (1.0 μM) with p < 0.005.
In conclusion, β-thioether-containing TFKs inhibited 2-OG hydrolysis in the cytosolic fraction of prostate cancer (PC-3) cells. The observed structure–activity relationships indicate that inhibitor lipophilicity is a major factor in the inhibition of 2-OG hydrolysis. The oxidation of the thioether to the corresponding sulfone essentially removes all biological activity. This observation could be due to a number of factors including steric and/or electronic effects from the sulfone moiety, reduction in the overall lipophilicity of the inhibitors, or effects upon the electrophilicity of the ketone. In addition, aromatic moieties appear to be unfavorable for inhibition of hydrolysis activity, although it is difficult to make conclusions with only one compound. Of particular interest is the 2-AG synthesis and inhibition of cell invasion in intact cells exhibited a maximum at alkyl chain length of 8- to 10- carbons of inhibitors. This indicates that the uptake efficiency of inhibitors into intact cells may be limited by their molecular structures. Further studies could explore the observed structure–activity relationships by expanding the aromatic-containing inhibitors to include larger aromatic systems and heteroatoms, as well as testing the sulfoxide analogs of the thioethers. The results from this study suggest that maintaining and/or increasing the endogenous concentration of 2-AG is critical for controlling cell invasion. Therefore, specific inhibitors of enzymes responsible for the degradation of 2-AG may be important therapeutic targets for treating prostate cancer.
Acknowledgments
This work was supported in part by the Wisconsin Breast Cancer Showhouse for a Cure, the Cancer Center of the Medical College of Wisconsin, and the National Institute of Health (DA 09155). C.E.W. was supported by NIH Post Doctoral Training Grant T32 DK07355-22. Partial support was also provided by NIEHS Grant R37 ES02710, NIESH Superfund Grant P42 ES04699, and NIESH Center for Environmental Health Sciences Grant P30 ES05707.
Footnotes
Abbreviations: 2-AG, 2-arachidonoylglycerol; CB receptor, cannabinoid receptor; DAG, diacylglycerol; DAK, diazomethylarachidonyl ketone; FAAH, fatty acid amide hydrolase; LC-ESI-MS, liquid chromatography-electrospray ionization-mass spectrometry; MAFP, arachidonyl fluorophosphonate; MGL, monoacylglycerol lipase; 2-OG, 2-oleoylglycerol, TFK, trifluoromethyl ketone.
References
- 1.Sugiura T, Nakane S, Kishimoto S, Waku K, Yoshioka Y, Tokumura A, Hanahan DJ. Occurrence of lysophosphatidic acid and its alkyl ether-linked analog in rat brain and comparison of their biological activities toward cultured neural cells. Biochim. Biophys. Acta. 1999;1440:194–204. doi: 10.1016/s1388-1981(99)00127-4. [DOI] [PubMed] [Google Scholar]
- 2.Mechoulam R, Ben-Shabat S, Hanus L, Ligumsky M, Kaminski NE, Schatz AR, Gopher A, Almog S, Martin BR, Compton DR, et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995;50:83–90. doi: 10.1016/0006-2952(95)00109-d. [DOI] [PubMed] [Google Scholar]
- 3.Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K, Yamashita A, Waku K. 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 1995;215:89–97. doi: 10.1006/bbrc.1995.2437. [DOI] [PubMed] [Google Scholar]
- 4.Bisogno T, Berrendero F, Ambrosino G, Cebeira M, Ramos JA, Fernandez-Ruiz JJ, Di Marzo V. Brain regional distribution of endocannabinoids: implications for their biosynthesis and biological function. Biochem. Biophys. Res. Commun. 1999;256:377–380. doi: 10.1006/bbrc.1999.0254. [DOI] [PubMed] [Google Scholar]
- 5.Stella N, Schweitzer P, Piomelli D. A second endogenous cannabinoid that modulates long-term potentiation. Nature. 1997;388:773–778. doi: 10.1038/42015. [DOI] [PubMed] [Google Scholar]
- 6.Goparaju SK, Ueda N, Yamaguchi H, Yamamoto S. Anandamide amidohydrolase reacting with 2-arachidonoylglycerol, another cannabinoid receptor ligand. FEBS Lett. 1998;422:69–73. doi: 10.1016/s0014-5793(97)01603-7. [DOI] [PubMed] [Google Scholar]
- 7.Di Marzo V, Bisogno T, De Petrocellis L, Melck D, Orlando P, Wagner JA, Kunos G. Biosynthesis and inactivation of the endocannabinoid 2-arachidonoylglycerol in circulating and tumoral macrophages. Eur. J. Biochem. 1999;264:258–267. doi: 10.1046/j.1432-1327.1999.00631.x. [DOI] [PubMed] [Google Scholar]
- 8.Di Marzo V, Bisogno T, Sugiura T, Melck D, De Petrocellis L. The novel endogenous cannabinoid 2-arachidonoylglycerol is inactivated by neuronal- and basophil-like cells: connections with anandamide. Biochem. J. 1998;331(Pt 1):15–19. doi: 10.1042/bj3310015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ueda N, Puffenbarger RA, Yamamoto S, Deutsch DG. The fatty acid amide hydrolase (FAAH) Chem. Phys. Lipids. 2000;108:107–121. doi: 10.1016/s0009-3084(00)00190-0. [DOI] [PubMed] [Google Scholar]
- 10.Goparaju SK, Ueda N, Taniguchi K, Yamamoto S. Enzymes of porcine brain hydrolyzing 2-arachidonoylglycerol, an endogenous ligand of cannabinoid receptors. Biochem. Pharmacol. 1999;57:417–423. doi: 10.1016/s0006-2952(98)00314-1. [DOI] [PubMed] [Google Scholar]
- 11.Dinh TP, Carpenter D, Leslie FM, Freund TF, Katona I, Sensi SL, Kathuria S, Piomelli D. Brain monoglyceride lipase participating in endocannabinoid inactivation. Proc. Natl. Acad. Sci. USA. 2002;99:10819–10824. doi: 10.1073/pnas.152334899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dinh TP, Freund TF, Piomelli D. A role for monoglyceride lipase in 2-arachidonoylglycerol inactivation. Chem Phys. Lipids. 2002;121:149–158. doi: 10.1016/s0009-3084(02)00150-0. [DOI] [PubMed] [Google Scholar]
- 13.Nithipatikom K, Endsley MP, Isbell MA, Falck JR, Iwamoto Y, Hillard CJ, Campbell WB. 2-Arachidonoylglycerol: a novel inhibitor of androgen-independent prostate cancer cell invasion. Cancer Res. 2004;64:8826–8830. doi: 10.1158/0008-5472.CAN-04-3136. [DOI] [PubMed] [Google Scholar]
- 14.Hammock B, Wing K, McLaughlin J, Lovell V, Sparks T. Trifluoromethylketones as possible trasition state analog inhibitors of juvenile hormone esterase. Pestic. Biochem. Physiol. 1982;17:76–88. [Google Scholar]
- 15.Hammock B, Abdel-Aal Y, Mullin C, Hanzlik T, Roe R. Substituted thiotrifluoropropanones as potent selective inhibitors of juvenile horomne esterase. Pestic. Biochem. Physiol. 1984;22:209–223. [Google Scholar]
- 16.Wheelock CE, Severson TF, Hammock BD. Synthesis of new carboxylesterase inhibitors and evaluation of potency and water solubility. Chem. Res. Toxicol. 2001;14:1563–1572. doi: 10.1021/tx015508+. [DOI] [PubMed] [Google Scholar]
- 17.Wheelock CE, Colvin ME, Uemura I, Olmstead MM, Sanborn JR, Nakagawa Y, Jones AD, Hammock BD. Use of ab initio calculations to predict the biological potency of carboxylesterase inhibitors. J. Med. Chem. 2002;45:5576–5593. doi: 10.1021/jm020072w. [DOI] [PubMed] [Google Scholar]
- 18.Kamita SG, Hinton AC, Wheelock CE, Wogulis MD, Wilson DK, Wolf NM, Stok JE, Hock B, Hammock BD. Juvenile hormone (JH) esterase: Why are you so JH specific? Insect Biochem. Mol. Biol. 2003;33:1261–1273. doi: 10.1016/j.ibmb.2003.08.004. [DOI] [PubMed] [Google Scholar]
- 19.Stok JE, Huang H, Jones PD, Wheelock CE, Morisseau C, Hammock BD. Identification, expression, and purification of a pyrethroid-hydrolyzing carboxylesterase from mouse liver microsomes. J. Biol. Chem. 2004;279:29863–29869. doi: 10.1074/jbc.M403673200. [DOI] [PubMed] [Google Scholar]
- 20.Nithipatikom K, Isbell MA, Lindholm PF, Kajdacsy-Balla A, Kaul S, Campbell WB. Requirement of cyclooxygenase-2 expression and prostaglandins for human prostate cancer cell invasion. Clin. Exp. Metastasis. 2002;19:593–601. doi: 10.1023/a:1020915914376. [DOI] [PubMed] [Google Scholar]
- 21.Boger DL, Sato H, Lerner AE, Austin BJ, Patterson JE, Patricelli MP, Cravatt BF. Trifluoromethyl ketone inhibitors of fatty acid amide hydrolase: a probe of structural and conformational features contributing to inhibition. Bioorg. Med. Chem. Lett. 1999;9:265–270. doi: 10.1016/s0960-894x(98)00734-3. [DOI] [PubMed] [Google Scholar]
- 22.Boger DL, Sato H, Lerner AE, Hedrick MP, Fecik RA, Miyauchi H, Wilkie GD, Austin BJ, Patricelli MP, Cravatt BF. Exceptionally potent inhibitors of fatty acid amide hydrolase: the enzyme responsible for degradation of endogenous oleamide and anandamide. Proc. Natl. Acad. Sci. USA. 2000;97:5044–5049. doi: 10.1073/pnas.97.10.5044. [DOI] [PMC free article] [PubMed] [Google Scholar]