

RANDOM union of gametes (RUG) is the null model central to theoretical population genetics. In this model, all male and female parents contribute their gametes equally to male and female gametic pools, respectively. Zygotes (offspring) are produced by random union of gametes, each from the male and female gametic pools. One assumption behind this model is that individuals mate an infinite number of times. Some organisms reproduce more or less in a fashion like the RUG model. For example, some aquatic animals release large numbers of gametes into the ocean, and some wind-pollinated plants release vast numbers of male gametes (Falconer 1989). However, this mating system is in reality rather rare. In most animal mating systems, individuals (rather than gametes) come together to mate. Even in some plant systems, this is effectively the case whenever a mass of pollen from a single donor plant is transferred to another plant (Nunney 1993).
Recently, Balloux and Lehmann (2003) relaxed the assumption of the RUG model, by allowing females to mate an arbitrary number of times. From a consideration of coalescence times, they derived an equation of the effective population size (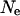 ) with an arbitrary number of matings. In a similar context, Nomura (2002a) independently developed an equation of the effective population size. As shown in the following, the equation of Balloux and Lehmann (2003) is a special form of the more general result given by Nomura (2002a).
) with an arbitrary number of matings. In a similar context, Nomura (2002a) independently developed an equation of the effective population size. As shown in the following, the equation of Balloux and Lehmann (2003) is a special form of the more general result given by Nomura (2002a).
Consider a population of dioecious species, in which 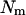 males mate randomly with
males mate randomly with 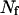 females. Subscript s (denoting m or f) is used to specify sex. Let
females. Subscript s (denoting m or f) is used to specify sex. Let 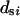 be the number of matings of parent
be the number of matings of parent 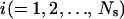 of sex s, with the mean
of sex s, with the mean 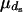 and variance
and variance 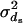 . There are
. There are 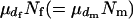 matings in the population. Then,
matings in the population. Then, 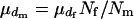 and
and 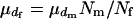 . Assuming a Poisson distribution of litter size (the number of newborns per mating), the equation given by Nomura (2002a) reduces to
. Assuming a Poisson distribution of litter size (the number of newborns per mating), the equation given by Nomura (2002a) reduces to
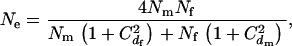 |
(1) |
where 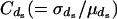 is the coefficient of variation of
is the coefficient of variation of 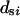 .
.
In the model of Balloux and Lehmann (2003), females are assumed to mate with a fixed number of randomly sampled males. Thus, 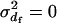 and
and 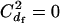 . For males, the number of matings (
. For males, the number of matings (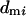 ) will follow a binomial distribution with the mean
) will follow a binomial distribution with the mean 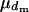 and variance
and variance 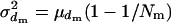 , giving
, giving
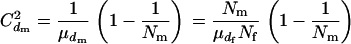 |
Substituting these expressions into Equation 1 leads to
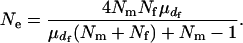 |
This agrees with Equation 7 in Balloux and Lehmann (2003), but with the neglect of two minor terms in their equation. It is more realistic to allow a variation in the number of matings for females (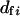 ). If
). If 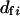 follows a Poisson distribution,
follows a Poisson distribution,  . Thus, the effective population size is expressed as
. Thus, the effective population size is expressed as
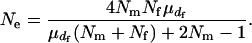 |
More generally, assuming that the number of matings follows a Poisson distribution within each sex, Equation 1 is written as
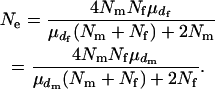 |
One of the most interesting applications of Equation 1 is that to harem polygamy in animals. In traditional works (e.g., Chepko-Sade et al. 1987), the effective population size under harem polygamy has been estimated with a well-known formula of Wright (1931):
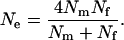 |
(2) |
However, this formula is based on the RUG model and holds under random mating with a finite number of matings only when there are no variations in the mating numbers within each sex; i.e., 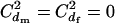 . A more appropriate equation, but with a simplicity comparable to Equation 2, is obtained from Equation 1. Under harem polygamy, successful males generally mate with most or all of the females in their harem, and the females generally mate with only one male. Thus,
. A more appropriate equation, but with a simplicity comparable to Equation 2, is obtained from Equation 1. Under harem polygamy, successful males generally mate with most or all of the females in their harem, and the females generally mate with only one male. Thus, 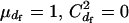 , and
, and 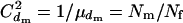 if the number of matings per male (harem size) follows a Poisson distribution. Then, from Equation 1, the effective population size is estimated by
if the number of matings per male (harem size) follows a Poisson distribution. Then, from Equation 1, the effective population size is estimated by
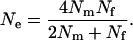 |
Other applications including the extension to sex-linked locus or haplo-diploid species are shown in Nomura (2002a).
Finally, the difference between the RUG model and random mating with a finite number of matings is specially important for considering the effect of heritable variation in fitness on the effective population size. The expected fitness of a parent under the RUG model is not affected by the genotypes of mates, because they are randomly sampled from the population. Thus, fitness in this model should be defined for individual parents. In contrast, under random mating with a finite number of matings, fitness should be defined for couples in monogamy and families in polygamy or polyandry. Further discussion on this topic and on the formulation of effective population size can be seen in Nomura (2002b).
Acknowledgments
This work was supported in part by grants-in-aid for scientific research (nos. 14560241 and 15658076) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan.
References
- Balloux, F., and L. Lehmann, 2003. Random mating with a finite number of matings. Genetics 165: 2313–2315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chepko-Sade, B. D., W. M. Shields, J. Berger, Z. T. Halpin, W. T. Jones et al., 1987. The effects of dispersal and social structure on effective population size, pp. 287–321 in Mammalian Dispersal Patterns: The Effects of Social Structure on Population Genetics, edited by B. D. Chepko-Sade and Z. T. Halpin. University of Chicago Press, Chicago.
- Falconer, D. S., 1989. Introduction to Quantitative Genetics, Ed. 3. Longman Harlow, Essex, UK.
- Nomura, T., 2002. a Effective size of populations with unequal sex ratio and variation in mating success. J. Anim. Breed. Genet. 118: 297–310. [Google Scholar]
- Nomura, T., 2002. b Effective size of populations with heritable variation in fitness. Heredity 89: 413–416. [DOI] [PubMed] [Google Scholar]
- Nunney, L., 1993. The influence of mating system and overlapping generations on effective population size. Evolution 47: 1329–1341. [DOI] [PubMed] [Google Scholar]
- Wright, S., 1931. Evolution in Mendelian populations. Genetics 16: 97–159. [DOI] [PMC free article] [PubMed] [Google Scholar]