

Abstract
G-protein-coupled receptors (GPCRs) have recently joined the list of cell surface receptors that dimerize. Dimerization has been shown to alter the ligand-binding, signaling, and trafficking properties of these receptors. Recent studies have shown that GPCRs heterodimerize with closely related members, resulting in the modulation of their function. In this study, we have attempted to determine whether members of GPCR superfamilies that couple to different families of G-proteins can associate and form oligomers. We chose the β2 adrenergic receptor that couples to stimulatory G-proteins and δ & κ opioid receptors that couple to inhibitory G-proteins. β2 and δ receptors undergo robust agonist-mediated endocytosis, whereas κ receptors do not. We find that when coexpressed, β2 receptors can form heteromeric complexes with both δ and κ receptors. This heterooligomerization does not significantly alter the ligand binding or coupling properties of the receptors. However, it affects the trafficking properties of the receptors. For example, we find that δ receptors, when coexpressed with β2 receptors, undergo isoproterenol-mediated endocytosis. Conversely, β2 receptors in these cells undergo etorphine-mediated endocytosis. However, β2 receptors, when coexpressed with κ receptors, undergo neither opioid- nor isoproterenol-mediated endocytosis. Moreover, these cells exhibit a substantial decrease in the isoproterenol-induced phosphorylation of mitogen-activated protein kinases. Taken together, these results provide direct evidence of heteromerization of GPCRs that couple to different types of G-proteins, which results in the modulation of receptor trafficking and signal transduction.
Keywords: receptor subtypes, G-protein-coupled receptor, endocytosis, dimers, heterodimerization
G-protein-coupled receptors (GPCRs), the largest and most diverse family of transmembrane receptors, are involved in the transduction of signals in response to a wide variety of stimuli. Until recently they were thought to function as monomers (1). However, a growing number of biochemical, biophysical, and functional studies suggest that GPCRs form functional, SDS-stable dimers (2). Heteromeric assembly of the nonfunctional γ-aminobutyric acid receptor B R1a and γ-aminobutyric acid receptor B R2 is necessary for γ-aminobutyric acid receptor-mediated signaling in cultured cells and in rat superior ganglion neurons (3–8). In contrast, fully functional opioid receptors associate with each other, resulting in the generation of heterodimeric receptors with unique properties (9–11). Heterodimerization of fully functional somatostatin receptor 5 with somatostatin receptor 1 has also been found to alter the pharmacology and signaling of both receptors (12). Although there is increasing evidence for receptor dimerization between members of the same subfamily of GPCRs, not much is known about dimerization between members of different subfamilies.
β2-Adrenergic receptors are prototypical receptors of the rhodopsin family that undergo rapid and robust agonist-mediated endocytosis. They have been shown to exist as dimers via noncovalent interactions, and dimerization favors receptor/G-protein coupling efficiency (13–15). Recent studies have shown that δ and κ opioid receptors exist as dimers (9, 16). These receptors differ in their agonist-mediated internalization properties; δ receptors undergo rapid and robust internalization (16, 17), whereas κ receptors do not (18). Moreover, a dimerization defective mutant of the δ receptor does not undergo agonist-mediated internalization, suggesting a role for dimerization in δ receptor internalization (16).
In the present study, we attempted to determine whether β2-adrenergic receptors that couple to stimulatory G-proteins and undergo robust internalization are able to associate with δ or κ opioid receptors that couple to inhibitory G-proteins but exhibit distinct internalization properties. In cells coexpressing δ and β2 (δ-β2 cells) or κ and β2 receptors (κ-β2 cells), we find biochemical evidence for receptor heteromerization at the cell surface. This phenomenon does not affect ligand binding or functional coupling but alters the trafficking properties. In δ-β2 cells, δ receptors undergo isoproterenol-mediated internalization and β2 receptors undergo opioid-mediated internalization. In κ-β2 cells, β2 receptors undergo neither isoproterenol- nor opioid-mediated internalization. Furthermore, there is a significant decrease in the level of isoproterenol-mediated MAP kinase phosphorylation in κ-β2 cells. These data support a role for GPCR oligomerization in modulating trafficking and signaling properties.
Materials and Methods
Cell Culture and Transfections.
Chinese hamster ovary cells, maintained in F-12 containing 10% FBS, were transfected by using lipofectin (GIBCO) reagent with equal amounts of the Flag-tagged human β2-adrenergic receptor cDNA in a geneticin-selectable vector pcDNA3 (generous gift from J. Javitch, Columbia University, New York, NY) and myc-tagged mouse δ or myc-tagged rat κ receptor cDNA in a hygromycin-selectable vector pCDNA 3.1. Cell lines stably expressing both receptors were selected in media containing 500 μg/ml each of geneticin and hygromycin (GIBCO). The cell surface expression of both receptors was confirmed by flow cytometry with monoclonal anti-Flag (M1; Sigma) and polyclonal anti-myc (c-Myc A14; Santa Cruz Biotechnology) antibodies. For transient expression, human embryonic kidney-293 (HEK-293) cells, maintained in DMEM containing 10% FBS, were transfected with receptor cDNAs (individually or in combination) by the use of calcium phosphate precipitation. Cells were collected for further experiments 48–72 h after transfection. The relative levels of receptor expression were approximately 1–3 pmol/mg protein.
Coimmunoprecipitation and Western Blotting.
HEK-293 cells expressing receptors were lysed for 1 h in buffer G (1% Triton X-100/10% glycerol/300 mM NaCl/1.5 mM MgCl2/1 mM CaCl2/50 mM Tris⋅Cl, pH 7.4), containing 100 mM iodoacetamide and a protease inhibitor mixture (9). For immunoprecipitation, 100–200 μg of protein was incubated with 1–2 μg of the polyclonal anti-myc antibody overnight at 4°C. Immunocomplexes were isolated by incubation with 10% vol/vol protein A-Sepharose (Sigma) for 2–3 h. The beads were washed three times with buffer G, resolved by nonreducing (without the addition of DTT to the sample buffer) 8% SDS/PAGE and subjected to Western blotting as described, with monoclonal anti-Flag antibody (16). In some cases the immunoprecipitate was treated with 100 mM DTT in the sample buffer (reducing conditions) and subjected to SDS/PAGE and Western blotting.
Detection of Cell Surface Heterooligomers.
Cell surface heteromeric complexes were visualized essentially as described (19). Briefly, HEK cells transiently expressing epitope-tagged receptors were washed with ice-cold PBS and incubated with 5 μg/ml of anti-flag antibody for 2 h at 4°C. Cells were washed and lysed with buffer G containing a protease inhibitor mixture (9) for 1 h at 4°C. The lysate was incubated with 15 μl of protein A-Sepharose 4B beads for 16 h at 4°C. The immunoprecipitates (containing cell surface receptors) were collected, washed twice with buffer G, and eluted with 30 μl of nonreducing sample buffer. Ten microliters of eluate was subjected to SDS/PAGE, and receptor proteins were detected by Western blotting analysis with the anti-myc antibody as described (16).
Membrane-Binding Assay.
Preparation of membranes and binding assays were carried out essentially as described previously (9). For most assays, membranes from cells transiently expressing individual or a combination of receptors were incubated with 0.3 nM [3H]CGP12177 or 2 nM [3H]diprenorphine in 50 mM Tris⋅Cl (pH 7.4) for 60 min at 37°C in the absence or presence of 8–10 concentrations of unlabeled ligands in a final volume of 1 ml. One micromolar unlabeled CGP12177 or diprenorphine was used to obtain nonspecific binding. The membranes were then collected on Whatman GF-B filters and washed, and the radioactivity was determined. IC50 values were determined from displacement curves by using graphpad prism 2.0.
cAMP Assays.
HEK cells transiently expressing 1–3 pmol/mg protein of epitope-tagged receptors were treated for 5 min with the indicated concentrations of isoproterenol or etorphine (after a 60-min pretreatment with forskolin) as described (20) (Fig. 2). The levels of intracellular cAMP after the stimulation of cells with ligands was determined by a RIA with antiserum to cAMP from Biomedical Technologies (Stoughton, MA) (20).
Figure 2.

Agonist-mediated accumulation of cAMP in cells individually expressing epitope-tagged δ, κ, and β2 receptors or cells coexpressing δ-β2 or κ-β2 receptors. Cells were treated with indicated concentrations of etorphine (Left) or isoproterenol (Right) for 5 min at 37°C, and the intracellular cAMP level was measured by RIA as described under Materials and Methods. The amount of cAMP in cells without agonist treatment is taken as 100%. The data represent means ± SEM (n = 3–5).
Internalization Assays.
Chinese hamster ovary or HEK cells expressing approximately equal levels of receptors were treated for 60 min with the indicated concentrations of etorphine (Fig. 3) or 100 μM isoproterenol (Fig. 4). The levels of cell surface receptors were examined by using anti-Flag or anti-myc antibodies by ELISA (21).
Figure 3.

Agonist-mediated internalization of receptors in HEK cells expressing δ, β2, δ-β2 (Left) or κ, β2, κ-β2 (Right) receptors. Cells were treated with indicated concentrations of etorphine for 60 min at 37°C, stained, and analyzed by ELISA, with the use of anti-Flag and anti-myc antibodies. The mean absorbance without agonist treatment is taken as 100%. The data represent means ± SEM (n = 5–7).
Figure 4.
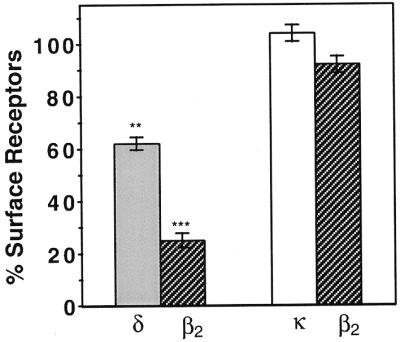
Agonist-mediated internalization of receptors in cells coexpressing κ-β2 or δ-β2 receptors. Cells were treated with indicated concentrations of 100 μM isoproterenol for 60 min at 37°C, stained, and analyzed by ELISA as described under Materials and Methods. The mean absorbance without agonist treatment is taken as 100%. Significant differences from untreated controls are indicated. **, P < 0.01, ***, P < 0.005 (n = 3–5).
Detection of the Phosphorylated MAP Kinase (pMAPK).
The level of pMAPK was determined essentially as described (22). Briefly, Chinese hamster ovary cells expressing δ-β2 or κ-β2 receptors were incubated with different concentrations of ligands for 5 min as described (9). Detection of pMAPK was by Western blotting with pMAPK antiserum (Cell Signaling Technologies/New England Biolabs) directed against phospho-extracellular regulated kinases (p42 and p44). Standardization was with tubulin measured in the same blots with antitubulin antibody (Sigma). NIH image 1.61 software was used to densitize and quantify the amount of pMAPK.
Results
δ or κ Opioid Receptors Oligomerize with β2-Adrenergic Receptors.
Heterodimerization between closely related GPCRs has been increasingly reported in recent years. In contrast, heterodimerization between distant family members is not well explored. We addressed this issue by examining the ability of peptide receptors that couple to inhibitory G-proteins (opioid receptors) to dimerize with monoamine (nonpeptide) receptors that couple to stimulatory G-proteins (β2-adrenergic receptors). For these studies, opioid receptors were tagged with a myc epitope, and β2 receptors were tagged with a Flag epitope. These receptors were expressed individually or in combination in heterologous cells, and the cell lysates were immunoprecipitated by using an anti-myc antibody. The resulting material was subjected to SDS/PAGE under nonreducing conditions and blotted with an anti-Flag antibody to visualize the β2 receptors. We find that we could coprecipitate fully glycosylated β2 dimers (≈130-kDa band) and tetramers (>200 kDa band); these oligomers are insensitive to DTT, inasmuch as treatment of immunoprecipitates with 100 mM DTT does not significantly alter their levels (Fig. 1A). We also see partially glycosylated monomers. These results confirm an interaction between opioid receptors and β2-adrenergic receptors. Opioid–β2 receptor interactions could not be detected when a mixture of cells individually expressing opioid and β2 receptors was subjected to similar immunoprecipitation/Western blotting analysis (Fig. 1A). These data suggest that oligomerization of δ or κ receptors with β2 receptors is not induced during the solubilization/immunoprecipitation procedures and that β2 receptors interact with δ and κ receptors only in cells coexpressing both receptors.
Figure 1.

(A) Myc-tagged δ or κ receptors interact with Flag-tagged β2 receptors to form heteromers. Immunoprecipitation of cell lysates from a mixture of HEK-293 cells individually expressing mycδ and Flagβ2 receptors (Flag β2 + mycδ) or Flagβ2 and mycκ receptors (Flagβ2 + mycκ) or cells coexpressing Flagβ2 and mycδ receptors (Flagβ2-mycδ) or Flagβ2 and mycκ receptors (Flagβ2-mycκ) was carried out with the use of anti-myc antibodies. Western blotting of the immunocomplexes with anti-Flag antibodies shows heteromers only in cells coexpressing the opioid and adrenergic receptors. IP, immunoprecipitation; WB, Western blotting. + DTT, immunoprecipitates treated with100 mM DTT in the sample buffer. (B) δ-β2 and κ-β2 heteromers exist on the cell surface. Cells individually expressing Flagβ2 or coexpressing Flagβ2-mycδ or Flagβ2-mycκ were incubated with anti-myc antibodies before solubilization and immunoprecipitation, as described under Materials and Methods. Western blotting of the immunocomplexes with anti-Flag antibodies shows the presence of the heteromers only in cells coexpressing both receptors. The 50-kDa band corresponds to a crossreactivity of the secondary Ab with the monoclonal Ab used for IP.
To determine whether these receptors exist on the cell surface, we prelabeled and immunoprecipitated cell surface β2 receptors with anti-Flag antibody and assayed for the presence of opioid receptors by probing blots with an anti-myc antibody. As seen in Fig. 1B, we can coprecipitate glyocosylated δ and κ dimers (≈130 kDa and ≈150 kDa) and their respective glycosylated monomers (≈70 kDa and ≈80 kDa). These results demonstrate the existence of opioid-β2 heteromers on the cell surface.
Ligand Binding and Functional Coupling Properties of Opioid-β2 Oligomers.
To determine whether physical interactions lead to changes in receptor pharmacology, the ligand binding properties were examined. We find that the properties of δ or κ receptors in membranes expressing δ and β2 or κ and β2 receptors are similar to the properties of these receptors expressed alone (Table 1). In all cases κ and δ receptors exhibited high affinity for their selective ligands (Table 1). Similarly, the binding properties of β2 receptors in membranes expressing heteromers are similar to the properties of β2 receptors expressed alone (Table 1). β2 receptors in all cases exhibited high affinity for β2-selective ligands. Thus it appears that the heteromerization between δ or κ receptors and β2-adrenergic receptors does not lead to alterations in their pharmacological properties.
Table 1.
Ligand-binding properties of the δ–β2, κ–β2, δ, κ, and β2 receptors
| Ligand |
Ki
|
||||||
|---|---|---|---|---|---|---|---|
| [3H]Diprenorphine
binding
|
[3H]CGP binding
|
||||||
| δβ2 | κβ2 | δ | κ | δβ2 | κβ2 | β2 | |
| CGP | >10 μM | >10 μM | >10 μM | >10 μM | 0.2 ± 0.3 nM | 0.9 ± 0.1 nM | 0.17 ± 0.03 nM |
| Isoproterenol | >10 μM | >10 μM | >10 μM | >10 μM | 1.1 ± 0.6 μM | 1.2 ± 0.1 μM | 1.17 ± 0.6 μM |
| Epinephrine | >10 μM | >10 μM | >10 μM | >10 μM | 1.2 ± 0.4 μM | 3.4 ± 0.6 μM | 3.12 ± 0.7 μM |
| DPDPE | 60 ± 6.4 nM | >10 μM | 82 ± 6.3 nM | — | >10 μM | — | >10 μM |
| U-69593 | — | 70.6 ± 6.4 nM | — | 65.7 ± 0.3 nM | — | >10 μM | >10 μM |
| nor-BNI | — | 0.7 ± 0.2 nM | — | 0.4 ± 0.3 nM | — | >10 μM | >10 μM |
Data represent means ± SEM (n = 3–7); —, not determined.
Next we attempted to determine whether heteromerization leads to alterations in functional coupling. The activation of opioid receptors results in an inhibition of adenylate cyclase activity, leading to a decrease in the level of intracellular cAMP, whereas the activation of β2 receptors results in the activation of adenylate cyclase, leading to an increase in the level of intracellular cAMP. The level of intracellular cAMP in response to etorphine, a universal opioid agonist, was examined in cells expressing individual or a combination of receptors. We find that this treatment results in a significant decrease in the level of cAMP in cells expressing κ receptors alone or with β2 receptors (Fig. 2). The level of decrease in cAMP in cells expressing heteromers is statistically indistinguishable from those expressing only κ receptors (Fig. 2). Similarly, there are no significant differences between cAMP levels in cells expressing δ receptors alone as compared with cells expressing δ-β2 receptors (not shown). Treatment of cells with a β2 agonist causes a dose-dependent increase in cAMP in all cells expressing β2 receptors; there are no significant differences between the cells expressing heteromers and the cells expressing individual receptors (Fig. 2). Treatment with cholera toxin completely abolishes the isoproterenol-induced increase in cAMP, and treatment with pertussis toxin abolished the etorphine-induced decrease in intracellular cAMP (not shown). These results support the notion that heteromerization with opioid receptors does not significantly affect the functional coupling properties (i.e., the adenylate cyclase activity) of β2 receptors.
Trafficking Properties of the Opioid-β2 Heteromer: EtorphineMediated Internalization.
To determine whether heteromerization affects the trafficking properties of these receptors, the extent of agonist-induced redistribution of receptors from the cell surface (i.e., internalization) was examined. Opioid-mediated internalization of δ receptors (expressed alone or with β2 receptors) was examined with the use of etorphine, a universal opioid agonist. Treatment with etorphine results in significant internalization of δ receptors both in cells expressing only δ receptors and in cells expressing δ-β2 receptors (Fig. 3). Surprisingly, β2 receptors, in δ-β2 cells, undergo a dose-dependent internalization in response to etorphine, as evidenced by a significant decrease in the level of cell surface β2 receptors (Fig. 3). Approximately 40% of β2 receptors are internalized within 60 min on treatment with 100 nM etorphine (Fig. 3). Etorphine does not cause internalization of β2 receptors when expressed alone (Fig. 3). Thus it appears that the coexpression of β2 receptors with δ receptors affects the trafficking properties of β2 receptors. We have previously shown that κ receptors do not undergo agonist-mediated internalization when expressed alone or together with δ receptors (9). As expected, neither κ nor β2 receptors, when expressed alone, could internalize in response to etorphine (Fig. 3). Furthermore, etorphine does not promote a significant internalization of β2 receptors in κ-β2 cells (Fig. 3).
Trafficking Properties of the Opioid-β2 Heterodimer: Isoproterenol-Mediated Internalization.
The extent of isoproterenol-mediated internalization of opioid and β2 receptors was also examined. We find that treatment with β2 agonists, as expected, results in a significant extent of internalization of β2 receptors in cells expressing δ and β2 receptors (Fig. 4). Surprisingly, this treatment also causes internalization of δ receptors in these cells; approximately 40% of δ receptors are internalized in 60 min by 100 μM isoproterenol (Fig. 4). Isoproterenol does not cause internalization of δ receptors in cells expressing only these receptors (not shown). Epinephrine, the endogenous β2 receptor agonist, is also able to internalize δ receptors in cells coexpressing δ and β2 receptors. This internalization could be blocked by β2 receptor-selective antagonists, but not by delta-selective antagonists (not shown). In cells expressing κ-β2 oligomers, treatment with a β2 agonist results in no significant internalization of β2 receptors, even at the highest dose of isoproterenol (Fig. 4). Thus it appears that heterodimerization with κ receptors drastically affects the trafficking properties of β2 receptors. Taken together, these results suggest that the coexpression of opioid receptors with β2 receptors has differential effects on β2 receptor trafficking. Expression of β2 receptors with δ opioid receptors results in the migration of the δ-β2 receptor complex to an intracellular compartment on activation of either one of the receptors. In contrast, expression of β2 receptors with κ receptors results in the retention of the β2 receptors on the cell surface.
Isoproterenol-Induced pMAPK Levels in Cells Expressing Opioid-β2 Receptors.
To explore the functional consequences of the altered trafficking properties of β2 receptors, the isoproterenol-mediated phosphorylation of MAP kinases was compared between cells expressing δ-β2 and κ-β2 receptors. We find that treatment with isoproterenol results in a dose-dependent increase in the level of phosphorylated MAP kinases in cells expressing δ-β2 heteromers (Fig. 5A Left). In contrast, treatment with fairly high doses of isoproterenol is not able to mediate a significant phosphorylation of the MAP kinases in cells expressing κ-β2 heteromers (Fig. 5A Right). The opioid-mediated phosphorylation of MAP kinases is not significantly altered in these cells, indicating that the ability to signal through opioid receptors is intact in these cells (Fig. 5B). These data suggest that dimerization with κ but not with δ receptors leads to a significant loss of β2 receptor-mediated activation of the MAP kinase pathway. Taken together, this observation supports the notion that agonist-mediated endocytosis plays a role in the activation of the MAP kinase pathway by β2 receptors.
Figure 5.

Heterodimerization with κ receptors affects the level of β2 receptor-mediated phosphorylation of pMAP kinase. Cells expressing δ-β2 or κ-β2 receptors were treated with indicated concentrations of isoproterenol (A) or etorphine (B) for 5 min. The extent of phospho-MAP kinase (pMAPK) was determined by Western blotting analysis with antiphospho-MAP kinase antibody; standardization was with tubulin measured in the same blots, with the use of antitubulin antibody.
Discussion
The existence of receptor homo- and heterodimers has been demonstrated for several GPCRs (23). However, the question of whether GPCRs from different subclasses are capable of heterodimerization has not been extensively examined. Two very recent studies have provided some evidence that receptors from different GPCR families interact through oligomerization (24, 25). However, these studies did not examine the effect of dimerization on the trafficking properties and the resulting modulation of function of the receptors. In the present study we explored the functional consequences of a physical interaction between a peptide receptor (opioid receptor) that couples to inhibitory G-proteins with a nonpeptide receptor (β2-adrenergic receptor) that couples to stimulatory G-proteins. We find that opioid receptors are able to form oligomers with β2-adrenergic receptors. The immunoprecipitation of δ receptors results in the coprecipitation of β2 tetramers and dimers. These homomeric forms of the β2 receptor are SDS stable and resistant to reducing agents. That we could also coprecipitate β2 monomers with opioid receptors suggests that the heteromeric interactions are SDS sensitive. Thus it appears that homooligomeric and heterooligomeric interactions exhibit differences in their physical properties.
In this study, no significant changes in ligand binding properties are observed. This is in contrast to the properties of other GPCR heterodimers, in which dimerization results in altered ligand binding properties (9–12, 24, 25). Thus it appears that heterodimerization differentially affects the properties of receptors, and this is unique for each heteromeric complex. Although the pharmacological properties are not significantly altered, the trafficking properties of these receptors are substantially modulated by heterodimerization. δ receptors coexpressed with β2 receptors gain the ability to internalize in response to β2 receptor-specific agonists. As shown by radioligand binding, none of the β2 receptor ligands bind directly to the δ receptor, suggesting that δ receptor internalization could be mainly because of interactions with β2 receptors. It is also possible that β2 agonist-induced internalization of the δ opioid receptor results from a heterologous (nonspecific) activation of these receptors because of their proximity with closely located active β2 receptors. The fact that δ receptor antagonists fail to block its internalization favors a role for heterodimerization in the internalization process. Furthermore, coexpression with the noninternalizing κ opioid receptor leads to a loss of agonist-mediated internalization of β2 receptors; this correlation strongly supports a role of heteromerization in GPCR trafficking. Heterodimerization between closely related GPCRs has been shown to modulate trafficking properties (6, 9, 12). In the case of GABAB receptors, heterodimerization is required for the maturation and translocation of the receptors to the cell surface (6). In the case of somatostatin and opioid receptors, heterodimerization regulates the agonist-mediated endocytosis and redistribution of the receptors from the cell surface to an intracellular compartment (9, 12). Thus it appears that physical interactions between GPCRs play an important role in the localization and trafficking of these receptors.
The internalization of GPCRs has been implicated in a variety of biological processes. This process is thought to be important for the resensitization of desensitized β2 receptors and has been implicated as a necessary step for the activation of MAPK by β2 agonists (26, 27). The present study shows that the loss of isoproterenol-mediated internalization is accompanied by a loss of MAPK activation in κ-β2 cells. However, the lack of β2 trafficking in these cells does not alter their ability to stimulate adenylyl cyclase. Thus it is possible that physical interactions between GPCRs can differentially alter the signal transduction by different pathways.
In vivo, opioid receptors and β2 receptors have been shown to coexist and functionally interact in cardiac myocyte sarcolemma (28, 29). Studies have shown that low doses of selective opioids are able to inhibit norepinephrine-mediated functions (28–30). Our studies demonstrate that opioid-selective agonists alter β2 receptor internalization, thus providing a possible molecular mechanism for the lack of β2 agonist mediated effects after opioid receptor activation. It is possible that bradycardia and other cardiac effects mediated by opioids may be due, in part, to direct physical interactions between these two receptors in cardiac tissue. In this case the activation of opposing pathways (opioids decrease intracellular cAMP and β2 receptors increase cAMP) and the physical regulation of these receptors may contribute to the resulting effects.
The interaction between opioid and β2 receptors raises the question of just how promiscuous these associations can be. So far, a wide variety of GPCRs have been shown to interact, each resulting in different functional consequences. Opioid heterodimers affect ligand binding, whereas other heterodimer pairs do not (31). Although it is likely that in their endogenous settings receptors are sorted and localized to specific areas, physical interactions could be mediated and/or regulated by “accessory” proteins that could further tailor the response to ligands. Oligomerization could thus represent a novel regulatory mechanism that could restrict or enhance receptor function.
Acknowledgments
We thank Jonathan Javitch and Brian Kobilka for the gift of Flag-tagged human β2-adrenergic receptor cDNA and Achla Gupta for help with the functional assays. This work is supported in part by Grants DA-08863 and DA-00458 (to L.A.D.) and postdoctoral training Grant DA-07254 (to B.A.J. and R.N.).
Abbreviations
- GPCRs
G-protein-coupled receptors
- MAP
mitogen-activated protein
- MAPK
MAP kinase
- pMAPK
phosphorylated MAPK
- HEK
human embryonic kidney
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.011384898.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.011384898
References
- 1.Gether U. Endocr Rev. 2000;21:90–113. doi: 10.1210/edrv.21.1.0390. [DOI] [PubMed] [Google Scholar]
- 2.Hebert I E, Bouvier M. Biochem Cell Biol. 1998;76:1–11. doi: 10.1139/bcb-76-1-1. [DOI] [PubMed] [Google Scholar]
- 3.Marshall F H, Jones K A, Kaupmann K, Bettler B. Trends Pharmacol Sci. 1999;20:396–399. doi: 10.1016/s0165-6147(99)01383-8. [DOI] [PubMed] [Google Scholar]
- 4.Jones K A, Borowsky B, Tamm J A, Craig D A, Durkin M M, Dai M, Yao W-J, Johnson M, Gunwaldsen C, Huang L-Y, et al. Nature (London) 1998;396:674–679. doi: 10.1038/25348. [DOI] [PubMed] [Google Scholar]
- 5.Kaupmann K, Malitschek B, Schuler V, Heid J, Froestl W, Beck P, Mosbacher J, Bischoff S, Kulik A, Shigemoto R, et al. Nature (London) 1998;396:683–687. doi: 10.1038/25360. [DOI] [PubMed] [Google Scholar]
- 6.White J H, Wise A, Main M J, Green A, Fraser N J, Disney G H, Barnes A A, Emson P, Foord S M, Marshall F H. Nature (London) 1998;396:679–682. doi: 10.1038/25354. [DOI] [PubMed] [Google Scholar]
- 7.Kuner R, Köhr G, Grünewald S, Eisenhardt G, Bach A, Kornau H-C. Science. 1999;283:74–77. doi: 10.1126/science.283.5398.74. [DOI] [PubMed] [Google Scholar]
- 8.Sullivan R, Chateauneuf A, Coulombe N, Kolakowski L F, Jr, Johnson M P, Hebert T E, Ethier N, Belley M, Metters K, Abramovitz M, et al. J Pharmacol Exp Ther. 2000;293:460–467. [PubMed] [Google Scholar]
- 9.Jordan B A, Devi L A. Nature (London) 1999;399:697–700. doi: 10.1038/21441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gomes I, Jordan B A, Gupta A, Trapaidze N, Nagy V, Devi L A. J Neurosci. 2000;20:RC110. doi: 10.1523/JNEUROSCI.20-22-j0007.2000. : 1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.George S R, Fan T, Xie Z, Tse R, Tam V, Varghese G, O'Dowd B F. J Biol Chem. 2000;275:26128–26135. doi: 10.1074/jbc.M000345200. [DOI] [PubMed] [Google Scholar]
- 12.Rocheville M, Lange D, Kumar U, Sasi R, Patel R C, Patel Y C. J Biol Chem. 2000;275:7862–7869. doi: 10.1074/jbc.275.11.7862. [DOI] [PubMed] [Google Scholar]
- 13.Angers S, Salahpour A, Joly E, Hilairet S, Chelsky D, Dennis M, Bouvier M. Proc Natl Acad Sci USA. 2000;97:3684–3689. doi: 10.1073/pnas.060590697. . (First Published March 21, 2000; 10.1073/pnas.060590697) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hebert T E, Moffett S, Morello J-P, Loisel T P, Bichet D G, Barret C, Bouvier M. J Biol Chem. 1996;271:16384–16392. doi: 10.1074/jbc.271.27.16384. [DOI] [PubMed] [Google Scholar]
- 15.Hebert T E, Loisel T P, Adam L, Ethier N, Onge S S, Bouvier M. Biochem J. 1998;330:287–293. doi: 10.1042/bj3300287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cvejic S, Devi L A. J Biol Chem. 1997;272:26959–26964. doi: 10.1074/jbc.272.43.26959. [DOI] [PubMed] [Google Scholar]
- 17.Trapaidze N, Keith D E, Cvejic S, Evans C J, Devi L A. J Biol Chem. 1996;271:29279–29285. doi: 10.1074/jbc.271.46.29279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jordan B A, Cvejic S, Devi L A. DNA Cell Biol. 2000;19:19–27. doi: 10.1089/104454900314672. [DOI] [PubMed] [Google Scholar]
- 19.Zeng F-Y, Wess J. J Biol Chem. 1999;274:19487–19497. doi: 10.1074/jbc.274.27.19487. [DOI] [PubMed] [Google Scholar]
- 20.Cvejic S, Trapaidze N, Cyr C, Devi L A. J Biol Chem. 1996;271:4073–4076. doi: 10.1074/jbc.271.8.4073. [DOI] [PubMed] [Google Scholar]
- 21.Trapaidze N, Gomes I, Bansinath M, Devi L A. DNA Cell Biol. 2000;19:195–204. doi: 10.1089/104454900314465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Trapaidze N, Gomes I, Cvejic S, Bansinath M, Devi L A. Brain Res Mol Brain Res. 2000;76:220–228. doi: 10.1016/s0169-328x(00)00002-4. [DOI] [PubMed] [Google Scholar]
- 23.Salahpour A, Angers S, Bouvier M. Trends Endocrinol Metab. 2000;11:163–168. doi: 10.1016/s1043-2760(00)00260-5. [DOI] [PubMed] [Google Scholar]
- 24.Rocheville M, Lange D, Kumar U, Patel S C, Patel R C, Patel Y C. Science. 2000;288:154–157. doi: 10.1126/science.288.5463.154. [DOI] [PubMed] [Google Scholar]
- 25.Gines S, Hillion J, Torvinen M, Le Crom S, Casado V, Canela E I, Rondin S, Lew J Y, Watson S, Zoli M, et al. Proc Natl Acad Sci USA. 2000;97:8606–8611. doi: 10.1073/pnas.150241097. . (First Published July 11, 2000; 10.1073/pnas.150241097) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lefkowitz R J. Nat Biotechnol. 1996;14:283–286. doi: 10.1038/nbt0396-283. [DOI] [PubMed] [Google Scholar]
- 27.Lefkowitz R J. J Biol Chem. 1998;273:18677–18680. doi: 10.1074/jbc.273.30.18677. [DOI] [PubMed] [Google Scholar]
- 28.Pepe S, Xiao R P, Hohl C, Altschuld R, Lakatta E G. Circulation. 1997;95:2122–2129. doi: 10.1161/01.cir.95.8.2122. [DOI] [PubMed] [Google Scholar]
- 29.Xiao R P, Pepe S, Spurgeon H A, Capogrossi M C, Lakatta E G. Am J Physiol. 1997;272:H797–H805. doi: 10.1152/ajpheart.1997.272.2.H797. [DOI] [PubMed] [Google Scholar]
- 30.Yu X C, Wang H X, Pei J M, Wong T M. J Mol Cell Cardiol. 1999;31:1809–1819. doi: 10.1006/jmcc.1999.1014. [DOI] [PubMed] [Google Scholar]
- 31.Devi L A, Brady L S. Neuropsychopharmacology. 2000;23:S3–S64. doi: 10.1016/S0893-133X(00)00154-8. [DOI] [PubMed] [Google Scholar]