

Abstract
On many cyclic-interval schedules, animals adjust their postreinforcement pause to follow the interval duration (temporal tracking). Six pigeons were trained on a series of square-wave (2-valued) interval schedules (e.g., 12 fixed-interval [FI] 60, 4 FI 180). Experiment 1 showed that pigeons track square-wave schedules, except those with a single long interval per cycle. Experiments 2 and 3 established that tracking and nontracking are learned and both can transfer from one cyclic schedule to another. Experiment 4 demonstrated that pigeons track a schedule with a single short interval per cycle, suggesting that a dual process—cuing and tracking—is necessary to explain behavior on these schedules. These findings suggest a potential explanation for earlier results that reported a failure to track square-wave schedules.
Hungry animals will readily learn to adjust their behavior to the typical time in between periodic food deliveries. Under some conditions, these adjustments take place rapidly. These rapid adjustments have been studied by manipulating the time between consecutive food deliveries. On cyclic-interval (CI) reinforcement schedules, for example, successive interfood intervals (IFIs) increase and then decrease progressively. On these schedules, animals typically adjust their postreinforcement pause according to the duration of the preceding interval—a generally adaptive behavior termed temporal tracking (Innis & Staddon, 1971).
On a typical CI schedule, the criterion interval duration is varied by successive small increments and decrements. Innis and Staddon (1971), for example, used a cycle that was an ascending arithmetic progression of interval durations followed by a descending one: 2t, 3t, 4t, 5t, 6t, 7t, 8t, 7t, 6t, 5t, 4t, 3t, 2t and so on. The value of t determined the absolute durations of the IFIs, as well as the size of the step between consecutive intervals. In their experiments, t ranged from 2 to 40 s for the different conditions. During each session, the same cycle was repeatedly presented throughout, and pigeons were run for many (>20) sessions under identical conditions. Only data from the last few sessions were analyzed. Under these conditions, they found that postreinforcement pause times for each pigeon showed cyclicity, with a period that corresponded to the IFI cycle.
Subsequent research established that animals track a wide range of interval sequences. Temporal tracking has been found when the progression between successive IFIs is arithmetic (Crystal, Church, & Broadbent, 1997; Innis, 1981; Innis & Staddon, 1970), logarithmic (Innis, 1981), sinusoidal (Higa, Wynne, & Staddon, 1991; Keller, 1973), or even random ascending (Church & Lacourse, 1998). When the IFIs in a cycle change at a geometric rate, tracking is weaker than with smaller jumps, indicating that the size of the change between successive intervals does influence the ability to track. Nonetheless, absolute interval ranges as varied as 1–512 s with rats (Church & Lacourse, 1998), and 5–15 s (Higa et al., 1991) to 40–320 s with pigeons (Innis & Staddon, 1971), with steps as large as 40 s are found to be reliably tracked.
One candidate explanation for this rapid temporal tracking is the linear-waiting hypothesis (Staddon & Higa, 1991; Wynne & Staddon, 1988; Wynne, Staddon, & Delius, 1996). In its simplest, one-back version, this hypothesis claims that the current wait time is entirely determined by the previous IFI. That is, postreinforcement pause (P) on trial n + 1 is a fixed proportion (A) of the interfood interval (I) on the previous trial (n):
| (1) |
where A is a constant less than 1 and B is small. What this simple equation captures is the intuition that an animal’s behavior is entirely elicited by the duration of the previous interval. For a CI schedule, linear waiting predicts that animals will show progressively longer pauses on the ascending leg of the cycle and progressively shorter pauses on the descending leg of the cycle, trailing the actual interval duration with a lag of 1. Most recent work on dynamic timing schedules gives strong support to this account. For instance, on a sinusoidal CI schedule, Higa et al. (1991) found the phase lag between intervals and pausing to be 1, indicating that pigeons’ adjusting pauses trailed the changing interval. The linear-waiting hypothesis is also supported by evidence from autocatalytic schedules (Wynne & Staddon, 1988), as well as single-impulse (Higa et al., 1991) and single-transition experiments (Higa, Thaw, & Staddon, 1993).
A striking failure of the linear-waiting hypothesis, however, is its inability to account for pigeons’ failure to track most square-wave (SW) cyclic schedules. A SW schedule is merely a CI schedule with two interval durations—short and long (Kello & Staddon, 1974; Staddon, 1967, 1969). On these SW schedules, pigeons do not show tracking behavior and respond at a higher rate during those periods when they are reinforced less frequently. Staddon (1967, 1969), for example, repeatedly presented pigeons with cycles of the form 12 fixed-interval (FI) 60, 4 FI 180: blocks of 12 short (60-s) intervals followed by blocks of 4 long (180-s) intervals. The linear-waiting hypothesis predicts longer pausing following all longer intervals and shorter pausing following all shorter intervals, but in fact, pigeons paused for almost the same duration following each reinforcer before starting to respond at a steady rate, showing no tracking whatsoever. As a result, this cyclic SW schedule produced the seemingly maladaptive and counterintuitive finding of higher overall response rates during the longer IFIs—that is, when the reinforcement density was actually lower.
Other features of behavior on these dynamic schedules hint at further limitations to simple linear waiting. In Innis and Staddon’s (1971) initial CI work, they found that the phase lag between the interval presented and pigeons’ pausing was typically zero, indicating that pigeons were anticipating the upcoming interval. Because the cycle began at the same point every session, they speculated that pigeons were able to learn the full cycle. More recent work, however, has failed to find clear evidence in support of this idea. For example, Higa et al. (1991) showed that on a sinusoidal CI, if several cycles at the end of a session are replaced by a simple FI, no wait-time cyclicity is seen on these ultimate intervals. On the other hand, there is evidence for anticipation in the experiments of Church and Lacourse (1998), who found that rats presented with a repeating ascending staircase of intervals showed pausing behavior appropriate to intervals 1 or 2 ahead in the cycle. There is also growing evidence for the influence on pausing of intervals further back than the immediately preceding one. For instance, Higa (1996) presented animals with a series of multiple short “impulse” FIs in a series of long FIs and found that pausing returned to baseline more slowly following more short impulses.
Thus, the simple, one-back linear-waiting account cannot be the whole story for responding on dynamic timing schedules. An animal’s behavior is sometimes controlled by more than the duration of the previous interval. Some of these findings have led researchers to consider models that use a greater number of previous intervals to determine the pause (e.g., Wynne et al., 1996; see General Discussion for an expanded treatment of this idea) or even more comprehensive theoretical frameworks for timing in general (Staddon & Higa, 1999).
A simple possibility raised by anticipatory effects is that the duration of interval n may, under some conditions, serve as a discriminative stimulus for the subsequent interval, n + 1. Perhaps each interval plays two roles in determining subsequent behavior: as a duration that elicits a certain wait time and as a discriminative cue that predicts the upcoming interval duration. This combination offers a possible explanation for why pigeons and rats sometimes show pausing behavior tailored to the current or even an upcoming interval on some cyclic schedules (Church & Lacourse, 1998; Innis & Staddon, 1971). Further support for such a two-fold process comes from experiments that used very long (720-s) intervals as the longer interval in the SW schedule. Pigeons exposed to a 12 FI 60, 2 FI 720 schedule actually showed the converse of tracking, with shorter pauses following longer intervals and vice versa (Innis & Staddon, 1970; Kello & Staddon, 1974). This result makes sense if pigeons learned that the very long 720-s interval served as a cue for the shorter 60-s interval.
In this article, we present results from four different experiments that studied the necessary and sufficient conditions for temporal tracking. We looked at the effects of the number of consecutive intervals (Experiments 1, 2, 3, and 4), the duration of those intervals (Experiment 2), and whether tracking occurs invariably and immediately on a given schedule or depends on the animal’s schedule history (Experiments 2 and 3). A more general aim was to explore interactions between other forms of learning (i.e., cuing or discriminative stimulus effects) and temporal tracking (Experiments 2, 3 and 4). In the course of these experiments, we also hoped to resolve the anomalous nontracking behavior observed many years ago on SW schedules (Staddon, 1967, 1969).
Experiment 1
Experiment 1 evaluated the effect of the number of consecutive short and long intervals in each cycle. Blocks of short (30-s) and long (90-s) intervals consisting of 1, 4, or 12 repetitions were presented in six different combinations. The linear-waiting account predicts tracking on all six of these SW schedules. Previous results with slightly longer intervals (60 s and 180 s), however, have failed to find tracking (Staddon, 1967, 1969). In addition, two of these conditions contained only a single long interval per cycle; as a result, this long interval was always followed by a shorter interval. If pigeons are indeed able to learn that the long interval duration can serve as a discriminative cue for the subsequent shorter interval duration, then tracking may be diminished (or even abolished) on these two schedules.
Method
Subjects
Six male pigeons (Columba livia) obtained from the Palmetto Pigeon Plant served as subjects. All subjects were between 2 and 4 years old and were kept at 85% of their ad-libitum weight throughout the experiment. Subjects were housed individually and received continuous access to water in their home cages. Four pigeons were experimentally naive (P7227, P7257, P7272, P7207), and 2 pigeons (P411, P440) had previous experience in an autoshaping procedure. All subjects were treated in accordance with the ethical guidelines outlined by the American Psychological Association.
Materials
Three operant chambers were built out of 24-gallon plastic storage containers (Rubbermaid Roughneck) with a height of 31 cm and a plastic grid floor measuring 37 cm × 46 cm. A touch-screen-equipped computer monitor was located in front of a 20 cm × 27 cm hole in the front wall of the chamber. Reinforcers consisting of mixed grain (Purina ProGrain for Pigeons) were delivered using a Coulbourn Instruments feeder (Allentown, PA, Model #E14-10) through a 5.0 cm × 4.5 cm hole on the right side of the container, 8.0 cm off the floor, and 8.5 cm from the front of the chamber. A fan mounted on the back wall of the chamber provided ventilation and masking noise.
Stimuli were presented on a 13-inch VGA Multisync A500+ monitor located ~1 cm behind a Carroll Touch Technology 13-inch infrared touch screen. Stimuli consisted of a white disk (radius = 40 pixels—termed the key) on a black background that appeared on the monitor 18 cm off the ground with a diameter of 2.5 cm. Responses to a rectangle with sides measuring 120 × 140 pixels around the key were recorded; all other responses were discarded. Three NEC Pentium-II computers (one per chamber) running the Microsoft Windows 95 operating system were used to control presentation of stimuli and recording of responses.
Procedure
Response training.
Pigeons were trained to peck at the key for food reinforcement using a shaping procedure that consisted of two simultaneous schedules on the single white key: an FR1 and an FTx schedule. On a given trial, reinforcement on the FR1 schedule terminated the FT schedule. With these contingencies in place, pigeons were reinforced for both the first peck they emitted on the key, and should they not peck, following a fixed amount of time. All pigeons readily learned to peck the key for food reinforcement under these contingencies.
FI training.
Reinforcers consisted of 2-s access to grain in a food hopper. Each session began with a 100-s start delay during which no stimulus was presented, followed by a single “free” (response-independent) reinforcer. Including this first reinforcer, there were 101 reinforcers available in each session. Intervals (30 s and 90 s) were all signaled by the presentation of the white key on the computer screen and timed from the end of the food access period; following expiration of the interval, the first peck on the key produced a food reinforcer. The white key stimulus was not displayed during the food reinforcer. Table 1 shows a list of all the conditions used in this first experiment. In the first phase of the experiment (1A), all 6 pigeons were initially trained for 10 days on an FI 30-s schedule. In the second phase (1B), half the animals (P411, P7272, and P7227) continued to be reinforced on an FI 30-s schedule, and the other three pigeons (P7257, P440, and P7207) were switched to an FI 90-s schedule for 25 sessions.
Table 1.
Conditions and Results From Each Phase in Experiment 1
| Mean pause (s) ± SEM |
||||||||
|---|---|---|---|---|---|---|---|---|
| Phase | Schedule | Sessions | Results | After FI 30 s | After FI 90 s | magnitude (s) | t | p |
| 1A | FI 30 | 10 | ||||||
| 1B | FI 30 or FI 90 | 25 | ||||||
| 1C | 12 FI 30, 12 FI 90 | 20 | Tracking | 8.3 ± 1.3 | 14.4 ± 1.9 | 6.1 | 7.61 | .0016 |
| 1D | 12 FI 30, 4 FI 90 | 25 | Tracking | 12.5 ± 1.7 | 16.3 ± 1.8 | 3.8 | 15.94 | .0001 |
| 1E | 4 FI 30, 4 FI 90 | 20 | Tracking | 11.9 ± 1.5 | 15.9 ± 1.8 | 4.0 | 5.97 | .004 |
| 1F/1G | 4 FI 30, 1 FI 90 | 15 | No tracking | 13.1 ± 1.8 | 14.3 ± 1.9 | 1.2 | 1.65 | .17 |
| 1F/1G | 4 FI 30, 12 FI 90 | 15 | Tracking | 12.2 ± 1.8 | 18.1 ± 1.7 | 5.9 | 8.51 | .0010 |
| 1H | 12 FI 30, 1 FI 90 | 12 | No tracking | 13.3 ± 1.7 | 12.6 ± 1.7 | −0.7 | −2.28 | .084 |
Note. Italics indicate statiscally reliable effects. FI = fixed interval.
SW schedules.
As indicated in Table 1, in the first phase with SW schedules (1C), lasting 20 sessions, all pigeons were placed on a 12 FI 30, 12 FI 90 schedule—a SW schedule of reinforcement that consisted of 12 short intervals (FI 30 s) followed by 12 long intervals (FI 90 s) in a repeating cycle. For all SW schedules, sessions started at a random point in the cycle and continued looping through the cycle until all reinforcers for that session had been delivered. To further explore the relation between run length (number of consecutive intervals) and tracking, in the next two phases, all 6 pigeons were then placed on a 12 FI 30, 4 FI 90 schedule for 25 sessions (1D), followed by a 4 FI 30, 4 FI 90 for 20 sessions (1E). During Condition 1E, two different hardware malfunctions disrupted the schedules received by P411. On consecutive days (Days 5 and 6), the feeder mechanism jammed and the screen failed to display the stimulus. The other pigeons were unaffected.
The remaining three conditions (1F–1H) explored whether a long run length of only 1 interval would result in unique behavioral patterns. These schedules are different from other SW cycles in that a long interval is always followed by a short interval, so that tracking the longer interval is maladaptive, and cuing—changing wait time in anticipation of the upcoming interval—is adaptive. These conditions should maximize the ability of the longer interval to act as a discriminative stimulus for the subsequent shorter interval. In the first two phases (1F/1G), pigeons were again split into two groups of 3 pigeons. The first group (P411, P7272, and P7227) was presented with a 4 FI 30, 12 FI 90 schedule followed by a 4 FI 30, 1 FI 90 schedule, and the second group (P7257, P440, and P7207) received the same two schedules in reverse order for 15 sessions each. In the final phase of this experiment, all 6 pigeons were placed on a 12 FI 30, 1 FI 90 schedule for 12 sessions.
Data analysis.
All simple calculations (e.g., means, t tests) were performed with Microsoft Excel. More advanced calculations (e.g., analysis of variance [ANOVA]) were performed with the STATISTICA Version 6.0 software package (StatSoft, 2001). The first four intervals of each session were considered “warm-up” trials and discarded from all analyses. In addition, any pauses that exceeded the longer criterion interval by more than 30 s were deemed outliers and excluded. An alpha level of .05 was selected for all inferential statistics.
Results
The primary dependent measure in this experiment was postreinforcement pause—the duration from the end of a period of food hopper access until the first key peck emitted by a pigeon. Only the data from the last 5 days of training in each condition were analyzed. We also looked at other dependent measures, including break point or start time (Cheng & Westwood, 1993; Church, Meck, & Gibbon, 1994; Schneider, 1969), quarter-life (Dukich & Lee, 1973; Freeman & Lattal, 1992; Herrnstein & Morse, 1957), overall response rate, and run rates, but none were found to have reliable and interpretable relations with the independent variables (see Discussion for more details). Over the last5 days of training with only FI 30 s (1A), pigeons paused for approximately one third of the interval duration (M = 10.8 s). Over the last 5 days of differential FI training (1B), pigeons trained with the FI 90 s paused for a greater absolute duration (M = 29.6 s), but a smaller relative proportion of the prevailing interval than those that continued to receive an FI 30 s (M = 14.2 s).
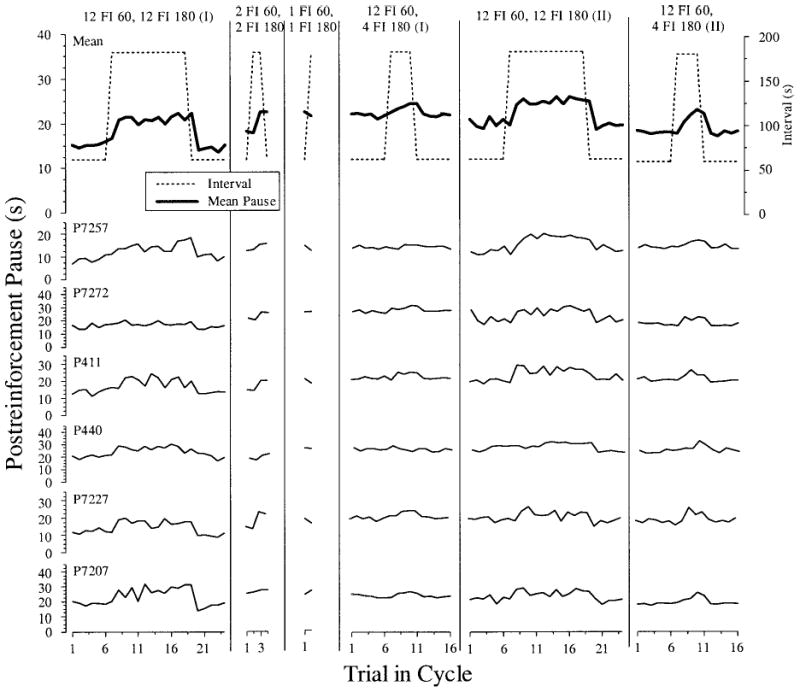
Figure 1 shows the results from the final five daily sessions for each of the six SW conditions in Experiment 1 (Phases 1C–1H in Table 1). The data points are the mean pause for all trials that occurred in that cycle position across all days of SW training. The top row represents the mean pause for all 6 pigeons as well as the scheduled interval duration for each trial in the SW cycle. Rows below show data from individual pigeons. All 6 pigeons in both groups (FI 30 and FI 90 pretraining from Phase 1B) showed clear one-back temporal tracking for the 12 FI 30, 12 FI 90 condition (1C). Following a longer interval, pigeons paused for a longer time and following a shorter interval, pigeons paused for a shorter time. This change in pause duration was evident following the first short interval or the first long interval in a cycle. Across successive long intervals, the pause gradually increased.
Figure 1.
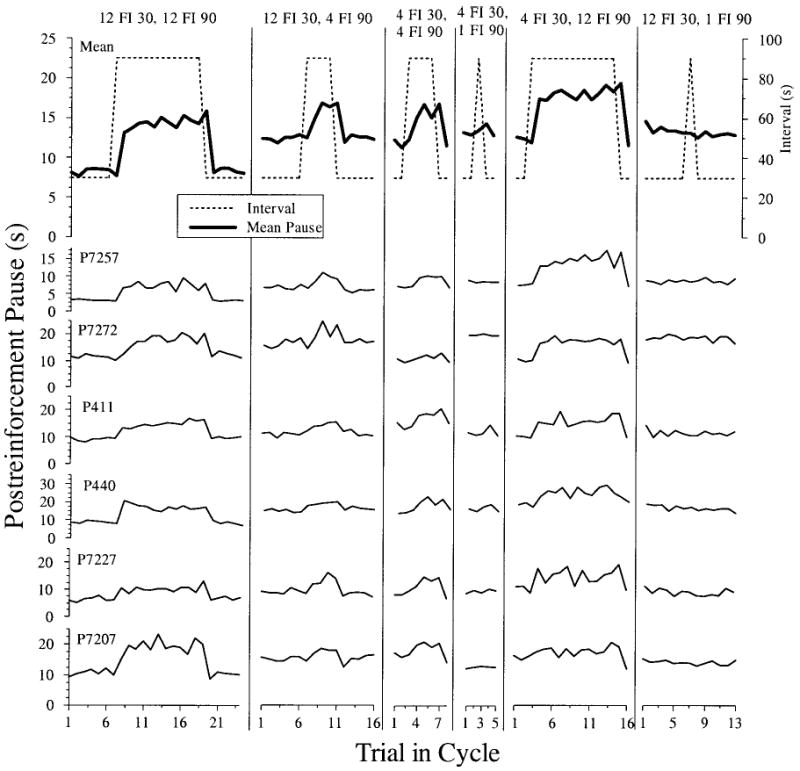
Postreinforcement pause as a function of trial into square-wave cycle for Conditions 1C–1H. The top row displays the mean pause for all pigeons for each trial in the cycle superimposed with the actual interval values, plotted on the right axis (see inset legend). The next six rows are the averaged responding across the final five experimental sessions in each condition for each individual pigeon. FI = fixed interval.
Table 1 summarizes these data, showing that the mean pause across all pigeons and all intervals in a cycle was substantially longer following the longer 90-s intervals for this 12 FI 30, 12 FI 90 schedule. The same tracking pattern was observed under the 12 FI 30, 4 FI 90; 4 FI 30, 4 FI 90; and 4 FI 30, 4 FI 90 conditions (Columns 2, 3, and 5 in Figure 1). This temporal tracking was attenuated in the remaining two conditions when the number of consecutive long intervals was only 1. With the 4 FI 30, 1 FI 90 schedule, only 2 pigeons (P440 and P411) showed any tendency toward a longer pause following the longer interval, but the increase was only a fraction of the increase seen on the other schedules. For the 12 FI 30, 1 FI 90 condition, none of the pigeons showed even the faintest evidence of tracking the single long interval; if anything, they showed a decrease in pausing following the longer interval.
To evaluate the statistical reliability of these effects, we conducted a multifactor, repeated measures ANOVA (Group × FI × Schedule) for all the SW conditions in Experiment 1. There was a main effect of FI, F(1, 4) = 187.0, p < .001, and an FI × Schedule interaction, F(5, 20) = 18.84, p < .0001. There was absolutely no effect of group nor any interactions (maximum F = 1.0, all ps > .40); thus, for all subsequent analyses, pigeons were collapsed into a single group. The final two columns of Table 1 summarize the results of planned comparisons of the two levels of FI (after FI 30 and after FI 90) that were conducted at the different levels of schedule (the different conditions). For all conditions with more than one consecutive long (90-s) interval, pauses were significantly longer following an FI 90 than following an FI 30. For the two conditions with only one consecutive long interval per cycle (4 FI 30, 1 FI 90 and 12 FI 30, 1 FI 90), there was no significant difference in pausing. For the 12 FI 30, 1 FI 90, there was even a nonsignificant trend toward shorter pausing following the longer interval.
The differential pausing following short and long FIs was statistically more robust and appeared much larger for those conditions with multiple consecutive long intervals. As a result, a direct comparison of the magnitude of tracking in the different conditions was conducted. As can be seen in Table 1, the mean difference in pausing following short and long intervals (pause after FI 90 – pause after FI 30) ranged from 6.1 s for the 12 FI 30, 12 FI 90 condition (1C) to −0.7 s for the 12 FI 30, 1 FI 90 condition (1H). A one-way repeated measures ANOVA confirmed a significant effect of SW schedule, F(5, 25) = 18.83, p < .0001. We followed up this analysis with a planned comparison, which indicated that the four conditions with multiple long intervals (12 FI 30, 12 FI 90; 12 FI 30, 4 FI 90; 4 FI 30, 4 FI 90; and 4 FI 30, 12 FI 90) produced a larger difference in pausing than the two conditions with a single long interval per cycle (4 FI 30, 1 FI 90 and 12 FI 30, 1 FI 90), t(5) = 18.30, p < .0001. Thus, we conclude that pigeons show much greater tracking following conditions with multiple consecutive long intervals than those with only a single long interval per SW cycle.
For those conditions with multiple long FIs, a key issue is whether tracking begins following the first long interval of a cycle. If temporal tracking is indeed one-back, as simple linear waiting predicts, then an increase (and decrease) in pause should be observed following the very first long (and short) interval in a cycle. As Figure 1 clearly shows, in the four conditions with blocks of multiple long FIs (Phases 1C, 1D, 1E, and 1F/G), pigeons showed an increase in pausing following the very first long FI (90 s) in a cycle (and a sharp decrease in pausing following the very first short FI [30 s] in a cycle).
A similar question can be asked of the statistical comparisons reported above that confirmed a reliable difference in the magnitude of tracking between single and multiple long FI conditions. Because of the varying number of long FIs, the difference in tracking magnitude may be due to differential averaging. By averaging only one long interval for those conditions with one long FI (1F/G, 1H) and by averaging together many long intervals per cycle for the remaining conditions, we might find a reliable difference in tracking magnitude, even if pigeons always paused for the same time following the first long FI of a cycle on any schedule. From visual inspection of the individual and mean data in Figure 1, the clear and immediate tracking displayed renders this possibility unlikely. Nonetheless, to confirm the statistical reliability of our results, we explored a potentially more accurate measure of the magnitude of tracking: subtracting the average pause following all 30-s intervals from the average pause following only the first FI 90 in a cycle. Figure 2 shows the mean magnitude of tracking, using this revised measure, for the four conditions with many long intervals and the two conditions with a single long FI per cycle. Tracking was reliably greater following conditions with many long FIs versus conditions with a single long FI per cycle, t(5) = 8.67, p < .001. Thus, pigeons clearly show much greater tracking in conditions with multiple consecutive long intervals than in those conditions with only a single long FI per cycle, even when only the pause in the interval following the first long FI is examined.
Figure 2.
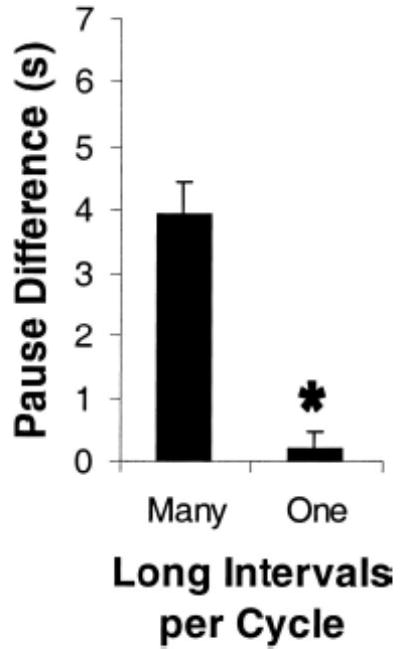
Mean (+ SEM) pause difference, computed by subtracting the average pause following an fixed-interval (FI) 30 from the average pause following the first FI 90 per cycle, for Conditions 1C–1H. The first column represents the average of all conditions with many long intervals per cycle (Phases 1C, 1D, 1E, and 4 FI 30, 12 FI 90 from 1F/1G). The second column depicts the average of both conditions in which there was only a single long interval per cycle (4 FI 30, 1 FI 90 and 12 FI 30, 1 FI 90). Data are from the final five sessions from each condition. *p < .001.
Discussion
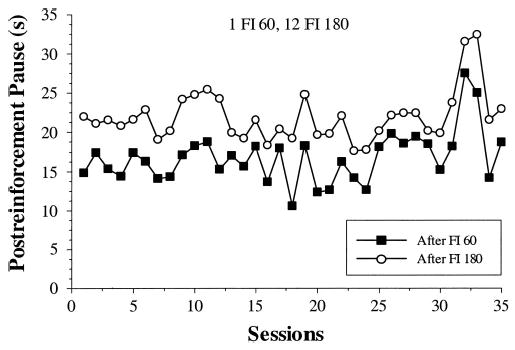
We found that pigeons clearly track a wide variety of SW interval schedules of reinforcement. Whenever the longer interval appeared more than once in succession per cycle, all pigeons showed clear temporal tracking, adjusting their postreinforcement pause in accordance with the duration of the preceding IFI. This tracking was apparent following the very first interval after a transition (see Figure 2). When the longer interval appeared only once per cycle, however, tracking was completely eliminated. These results are consistent with a two-process account, whereby behavior is determined by a combination of linear waiting and discriminative cuing. Under most conditions, pigeons show temporal tracking, exhibiting the effects of an obligatory linear-waiting process. For conditions in which the longer interval is always followed by a shorter interval, and can thus act as an effective discriminative stimulus, pigeons do not show tracking behavior. Our main finding that pigeons do indeed track certain SW schedules conflicts with the prior experimental results of Staddon (1967, 1969) who found a failure to track under similar conditions (12 FI 60, 4 FI 180). Two possible sources for this discrepancy are the different absolute interval durations (30/90 vs. 60/180) in the two experiments and the different experimental histories of the pigeons before the experiments began. We return to these issues in the next two experiments.
In drawing our conclusions, we used postreinforcement pause (or wait time) as our dependent variable because other measures, including break point, quarter-life (QL), overall response rate, and run rate are problematic in any condition in which multiple interval durations occur in the same session. For example, in a mix of 30- and 90-s intervals, measures that take into account responses during the 90-s interval that occur after 30 s may misrepresent differences in behavior during the two intervals. In the 12 FI 30, 1 FI 90 condition (1H), even if behavior during the FI 90 were identical to the FI 30 for the first 30 s, the pigeons’ behavior in the final 60 s of the longer interval might differ and thus generate differences in all these measures. Using the differential behavior as indicated by one of these measures, one would draw the false conclusion that pigeons can exactly anticipate the occurrence of a long interval every 13th interval.
Consider the QL measure, which is the time at which 25% of responses in a FI have been emitted (Dukich & Lee, 1973; Freeman & Lattal, 1992; Herrnstein & Morse, 1957). Suppose the pigeon responds at a constant rate of one response/s at all times, regardless of interval duration or any other variable. Now, expose this pigeon to a schedule of reinforcement containing two different FI values (e.g., 30 and 90 s). If one were to then use the QL as a dependent measure, one would get a QL of 7.5 s for the 30-s interval and a QL of 22.5 s for the 90-s one, despite the fact that the pigeon is not responding differently during the two intervals. The QL may be a useful measure when comparing varying performance with the same FI value (e.g., in a drug study), but not when multiple FI values are involved. The break point or start time (Cheng & Westwood, 1993; Church et al., 1994; Schneider, 1969) also suffers from the same logical (and confirmed empirical) defect whenever responding does not closely approximate a break-and-run pattern and multiple interval durations are being compared (see also Staddon, 2001, chap. 13, Figure B13.1.1). In contrast, in this hypothetical situation, the postreinforcement pauses would be identical, thereby accurately depicting the animal’s behavior in this situation. Thus, measures that include responding late in the interval were not considered any further in this study.
By examining only the final few days of responding on these SW schedules, we have ignored a potentially important element of the dynamics of pausing. If the longer interval is serving as a discriminative stimulus for the shorter interval, pigeons would need to learn this relationship; thus, tracking should initially be apparent when a schedule that does not produce tracking follows a reinforcement schedule that does. Unfortunately, the design of Experiment 1 did not allow us to accurately evaluate these sequential effects because of the counterbalancing across schedules in Phases 1F/1G. Experiment 2 thus attempted to determine whether temporal tracking (and nontracking) is immediately apparent when a pigeon is exposed to a particular schedule of reinforcement or whether the behavior is learned across sessions.
Experiment 2
In Experiment 1, we observed that pigeons can and do display tracking behavior on SW schedules, except when there is only a single long interval per cycle. To what extent are these two patterns, tracking and nontracking, learned behaviors that change across time? To what extent do these two patterns depend on previous training? In an experiment with a three-valued cyclic schedule (5, 15, and 45 s), Higa et al. (1993) found that pigeons initially showed tracking behavior, but this pattern disappeared after a few sessions. These results suggest that pigeons can learn to change their pausing pattern in response to the reinforcement schedule.
The present experiment was designed to examine these two learning effects by looking at the initial sessions following a transition from a tracking condition to a nontracking condition and vice versa. In the four phases that comprised this experiment, we first administered a schedule with blocks of multiple long intervals, followed this phase with two different conditions containing only a single long interval per cycle, and ended with a second condition with blocks of multiple long intervals (see Table 2 for details). If pigeons are learning to use the interval duration as a discriminative stimulus during the one-long conditions, then the nontracking behavior should show a learning curve at the outset of those conditions. As well, we would expect to see residual effects of this learned discriminative cue for the first few sessions of the second tracking condition.
Table 2.
Conditions and Results From Each Phase in Experiment 2
| Mean pause (s) ± SEM |
||||||||
|---|---|---|---|---|---|---|---|---|
| Phase | Schedule | Sessions | Results | After Short FI | After Long FI | Tracking magnitude (s) | t | p |
| 2A | 12 FI 30, 20 FI 90 | 10 | Tracking | 13.2 ± 2.3 | 19.6 ± 2.8 | 6.4 | 6.11 | .002 |
| 2B | 1 FI 30, 1 FI 90 | 26 | Disappearing tracking | 13.9 ± 1.8 | 14.7 ± 1.6 | 0.8 | 0.95 | .38 |
| 2C | 1 FI 60, 1 FI 180 | 30 | No tracking | 15.5 ± 1.2 | 14.8 ± 0.9 | −0.7 | −0.92 | .41 |
| 2D | 12 FI 60, 12 FI 180 | 25 | Appearing tracking | 16.6 ± 2.0 | 21.0 ± 2.1 | 4.4 | 5.99 | .002 |
Note. Italics indicate statistically reliable effects. FI = fixed interval.
A secondary aim was to determine whether absolute interval duration (30 and 90 s) is critical for the pattern of responding observed in the first experiment. Hence, this experiment also used 60 s and 180 s as the short and long interval durations respectively in the SW schedule.
Method
Subjects
The same 6 pigeons (P7257, P7272, P411, P440, P7227, and P7207) from Experiment 1 participated in this experiment.
Materials
The same three operant chambers, run by the same hardware and software, were used.
Procedure
The basic procedure was identical to the one used in Experiment 1. All sessions began with a 100-s start delay, followed by a 2-s food reinforcer. A further 100 reinforcers were available according to a specified SW interval schedule of reinforcement. Table 2 summarizes the four phases that comprised this experiment. In the first phase of this experiment (2A), pigeons were presented with a 12 FI 30, 20 FI 90 schedule for 10 sessions. In the second phase (2B), pigeons were switched to a simple-alternation 1 FI 30, 1 FI 90 schedule for 26 sessions. On the basis of results from Experiment 1, pigeons were expected to track the first condition (2A), but not to show temporal tracking in the second condition (2B) because the long interval was only presented once per cycle. The development across days of pigeons’ pausing behavior in the second condition should indicate whether nontracking behavior is a learned response or an immediate reaction to the simple-alternation schedule.
The third phase of the experiment (2C) kept the simple-alternation pattern in place, but altered the absolute interval durations for a 1 FI 60, 1 FI 180 schedule for 30 sessions. With these longer interval durations, the total number of food reinforcers per session was reduced from 101 to 65, keeping session lengths at approximately 2 hr. This condition immediately followed the first simple-alternation condition (2B) to determine whether the anticipated nontracking behavior of the pigeons would transfer to another simple-alternation schedule with different interval durations. The fourth, final phase switched back to the 12–12 pattern, which produced tracking with 30- and 90-s intervals in Experiment 1, but with 60- and 180-s intervals, for 25 sessions. This manipulation should reveal whether tracking behavior is expressed immediately (or even at all) following a condition in which pigeons do not track. These final two conditions also test whether using a different pair of absolute interval durations (with the same 3:1 ratio) as the two components in the SW cycle influences the pattern of results found in Experiment 1.
Results
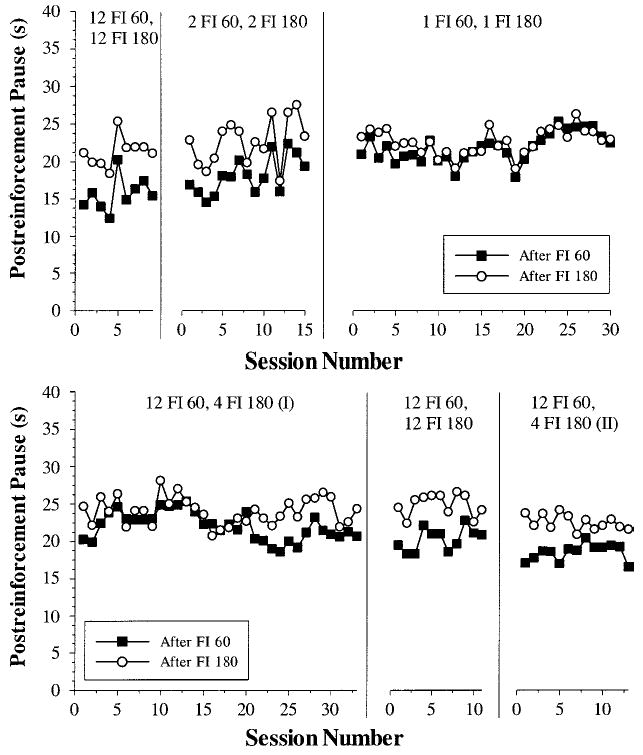
The main dependent measure in this experiment was once again the postreinforcement pause. Figure 3 illustrates the pause as a function of trial in the SW cycle for each of the pigeons, averaged across all daily sessions. As was the case in Experiment 1, for the two conditions with multiple consecutive long intervals (the first and last columns), all pigeons showed temporal tracking. (The first two columns contain data from 6 pigeons, and the second two columns contain data from only 5 pigeons, as P440 became ill at the end of phase 2B and was unable to complete the remainder of this experiment.) For the two simple-alternation schedules (center two columns), pigeons showed very little difference in pausing following long or short interval durations. Under the 1 FI 30, 1 FI 90 schedule (2B), 4 pigeons (P7257, P411, P440, and P7227) paused minimally longer following the longer intervals, whereas the other two pigeons showed the opposite effect. For the 1 FI 60, 1 FI 180 condition (2C), the same pattern held with very little effect of previous FI duration on pausing in the subsequent interval. If anything, the mean for all pigeons showed the opposite (anticipatory) effect with longer pauses following the shorter intervals and vice versa.
Figure 3.
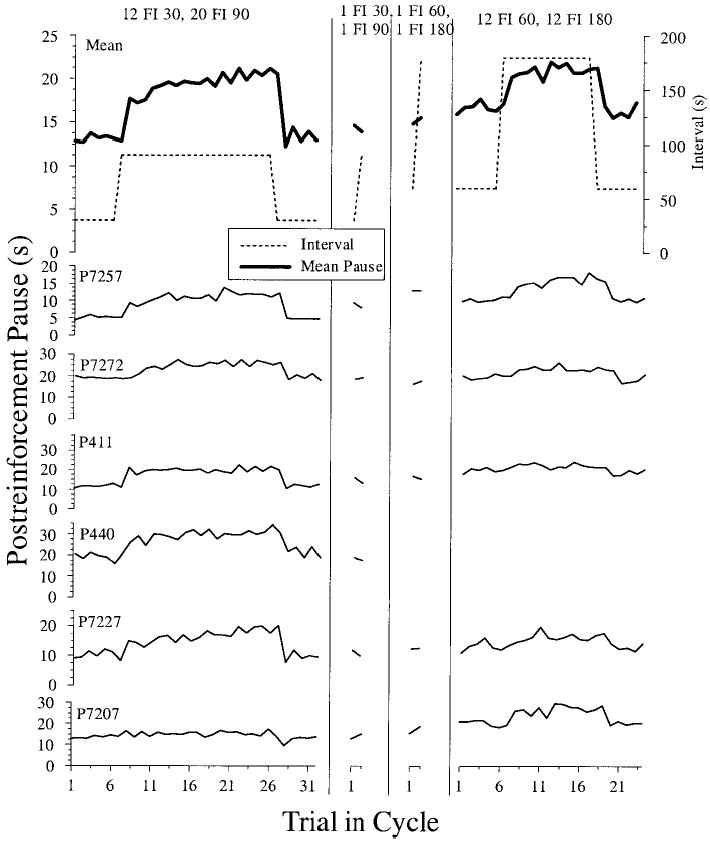
Postreinforcement pause as a function of trial into square-wave cycle for Conditions 2A–2D. The top row displays the mean pause for all pigeons for each trial in the cycle superimposed with the actual interval values, plotted on the right axis (see inset legend). The next six rows are the averaged responding across all experimental sessions for each individual pigeon. Pigeon P440 became ill after the second condition and did not complete this experiment. FI = fixed interval.
Table 2 summarizes the pausing data, by averaging across all trials that follow either a short or a long FI for all pigeons. The same pattern of results was again evident with pigeons showing robust temporal tracking when multiple consecutive intervals occur per cycle (top and bottom rows), but not in the simple-alternation conditions (middle two rows). To confirm these visible trends statistically, two multifactor, repeated measures ANOVAs (FI × Schedule) were conducted on the dataset—one for the pair of conditions that used 30 s and 90 s as the interval durations (2A and 2B) and another for the pair of conditions that used 60 s and 180 s as the interval durations (2C and 2D). For the two conditions that used 30- and 90-s interval durations, there was a main effect of FI, F(1, 5) = 16.61, p < .01, but no main effect of schedule, F(1, 5) = 3.71, p = .11. A significant FI × Schedule interaction was found as well, F(1, 5) = 89.61, p < .001, indicating that the effect on pausing of the different FI values (30 or 90 s) was different for the two different SW schedules. Planned comparisons of the simple effects of FI for the two different levels of schedule (12/20 and 1/1) teased out the details of this interaction and are displayed in the final columns of Table 2. These tests confirmed that pausing was reliably longer following FI 90 s for the schedule with multiple consecutive long intervals (2A), but there was no effect of FI on pausing for the simple-alternation schedule (2B). A comparison of the magnitude of tracking (mean pause after FI 90 – mean pause after FI 30) confirmed much greater tracking for the schedule with many long intervals than for the simple-alternation schedule, one-tailed t(5) = 9.47, p < .001. Taken together, these results confirm that pigeons showed longer pauses following longer intervals in the 12 FI 30, 20 FI 90 condition (2A), but did not show this tracking behavior in the 1 FI 30, 1 FI 90 condition (2B).
For the two conditions that used 60- and 180-s interval durations (2C and 2D), there were main effects of FI and schedule as well as an FI × Schedule interaction (all Fs > 4.3, ps < .05). We conducted planned comparisons of the simple effects of FI for the different levels of schedule to further evaluate this interaction. These comparisons (see Table 2) indicated that there was no effect of FI duration on pausing for the simple-alternation (1/1) condition (2C), but pausing was significantly longer following the long FI 180 s for the condition with multiple consecutive long intervals (2D). A comparison of the magnitude of tracking (mean pause after FI 180 – mean pause after FI 60) also indicated greater tracking for the schedule with many long intervals than for the simple-alternation schedule, one-tailed t(4) = 3.64, p < .05. These statistical results confirm the pattern observed in the individual pigeon (Figure 3) and mean data (Table 2) that pigeons pause longer following the longer interval when multiple consecutive long intervals occur per cycle (2D) but not in the simple-alternation condition (2C). This pattern of temporal tracking results held for both the 30–90 schedule and the 60–180 pair, indicating that within this range the absolute interval duration did not make a difference.
To examine how quickly the pigeons’ pauses adjusted to the interval changes, we also compared pausing after the first trial in each cycle of the two different interval durations. The differences detailed above between conditions with multiple consecutive identical intervals (2A and 2D) and those with a single long and short interval per cycle (2B and 2C) may result because many trials per cycle are averaged together in the former conditions but not the latter. Even when tracking does occur, the effects may not be apparent on the first trial following a change. As was the case with the mean of all the intervals, for the 12 FI 30, 20 FI 90 condition (2A), the pause following only the first long (90-s) interval of a cycle was significantly longer than the pause following only the first short (30-s) interval of a cycle, one-tailed t(5) = 3.99, p < .01. This pattern also held for the 12 FI 60, 12 FI 180 condition (2D); pauses following the first 180-s interval of a cycle were reliably longer than pauses following the first FI 60 s, one-tailed t(4) = 2.55, p < .05.
Having confirmed that tracking occurs when blocks of multiple long intervals are cycled with blocks of multiple short intervals, we now turn to the questions of learning and history-dependence. Figure 4 illustrates the development of the pattern of pausing behavior across sessions for Phases 2B–2D. In each panel, the two curves represent the mean pause across pigeons for all trials that followed either a short (30 or 60 s) or a long (90 or 180 s) interval plotted as a function of SW training day. The first panel of Figure 4 shows that in the first simple-alternation phase (2B), which followed a condition that pigeons tracked (2A), tracking behavior (a difference between the two curves) occurred in the initial few sessions, but disappeared after approximately 9 days. The second panel in Figure 4 shows the second single-alternation phase (2C), which followed a nontracking condition (2B). In this case, pigeons did not track right from the outset. In the final phase, a tracking condition (2D; third panel), which also followed a nontracking condition (2C), tracking was not observed at first, but appeared after approximately 9 days.
Figure 4.
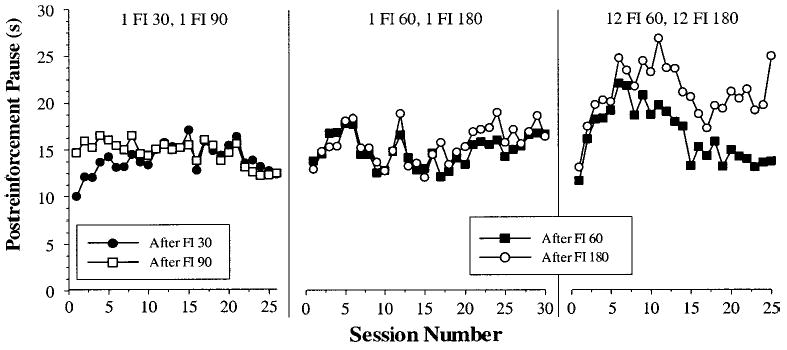
Session-by-session changes in mean postreinforcement pause for all pigeons in Conditions 2B–2D. From left to right, the panels represent mean daily values for 1 fixed-interval (FI) 30, 1 FI 90 (2B); 1 FI 60, 1 FI 180 (2C); and 12 FI 60, 12 FI 180 (2D). The data in the third panel represent an average of all trials (not only the first per cycle) that followed an FI 60 or an FI 180. The first panel is an average of 6 pigeons; the second and third panels are averages of 5 pigeons.
To statistically confirm these learning effects, we conducted three two-way (FI × Session Blocks) ANOVAs: one for each of the final three phases in this experiment (2B–2D). For the first single-alternation condition (2B), we divided the 26 days of this condition into four blocks of 5 days and a final block of 6 days and found no main effect of FI or sessions (Fs < 1.2, ps > .3), but a significant interaction, F(4, 20) = 13.56, p < .001. A pair of planned comparisons confirmed that there was reliable tracking during the first two blocks (10 days), t(5) = 3.90, p < .05, but no reliable effects of previous FI duration for the final three blocks (16 days), t(5) = 0.31, p = .76. For the subsequent single-alternation condition (2C), we used blocks of 10 sessions and found no main effects or interactions (all Fs < 3.3, ps > .05), confirming that pigeons consistently did not show any tracking behavior throughout this condition. Finally, for the 12 FI 60, 12 FI 180 condition (2D), we again used five blocks of five sessions each and found that there were significant main effects of FI and sessions and an FI × Sessions interaction (all Fs > 4.3, ps < .05). Another pair of planned comparisons confirmed that pigeons initially (for the first two blocks or 10 days) did not show tracking behavior on this schedule, t(4) = 1.54, p = .2, but did display this tracking pattern for the final three blocks (15 days), t(4) = 4.93, p < .05. These statistical results all corroborate the visible trends in Figure 4 that both tracking and nontracking are influenced by the previous schedule and are learned across sessions.
Discussion
Once again, pigeons showed temporal tracking behavior in the conditions that had blocks of multiple short and long intervals in a repeating cycle. In the two single-alternation conditions, pigeons paused similarly regardless of the previous interval duration. This pattern held for both the 30- and 90-s intervals and the 60- and 180-s intervals. Of greatest interest to the main goal of this experiment, tracking (and nontracking) seems to be a learned behavior, taking about nine daily sessions to develop following a nontracking condition (2C→2D) and about the same number of sessions to disappear following a tracking condition (2A→2B). Both of these results can be interpreted in terms of a dual-process account. When pigeons first encounter the simple-alternation schedule (2B), they learn that long intervals cue short intervals (and/or vice versa). When pigeons are subsequently presented with a schedule with multiple long intervals (2D), they need to un-learn this discriminative cue before they display tracking again.
The learning across sessions of temporal tracking suggests a promising avenue for resolving the incongruity between the Staddon (1967, 1969) failure-to-track results and the current set of experiments that clearly demonstrate tracking on SW schedules. The pigeons in Staddon’s (1967) failure-to-track results and the current set of experiments that clearly demonstrate tracking on SW schedules. The pigeons in Staddon’s (1969) experiments were not naive; perhaps some contingency in their previous reinforcement history trained them not to track the subsequent SW schedules. Experiment 3 investigated this possibility.
Experiment 3
One question that remains unanswered by the first two experiments is why Staddon (1967, 1969) found that pigeons failed to track a 12 FI 60, 4 FI 180 schedule. Some minor procedural details do differ between the first two experiments here and the Staddon research. The duration of reinforcement was 3 s in the earlier experiments and only 2 s in the current experiments. In the present experiments, the sessions started at a random point in the cycle, whereas, in the older work, cycles started at the same point in the cycle every day. Although we replicated the run lengths (12 and 4) and interval-duration ratio (3:1) of the earlier studies in Experiment 1, we used different interval durations (30 and 90 vs. 60 and 180). It remains possible, therefore, that these apparently modest differences may account for the differences in tracking we observed. The likelihood of this prospect is limited as pigeons did show tracking with a 12:4 short–long ratio when the intervals were 30 s and 90 s (see Phase 1D in Table 1), and pigeons also tracked when the intervals were 60 s and 180 s if the short–long ratio was 12:12 (see Phase 2D in Table 2). Nonetheless, the possibility still exists that those exact parameters for the SW schedule would produce different behavior.
In Experiment 3, therefore, we explored the behavior of pigeons on the same 12 FI 60, 4 FI 180 schedule used by Staddon (1967, 1969) but following two different historical contexts. Our hypothesis, based on the results of Experiment 2, was that the differential reinforcement history of pigeons can determine the pattern of tracking/nontracking that they display on this SW schedule of reinforcement. The 12 FI 60, 4 FI 180 schedule was first administered following a condition (1 FI 60, 1 FI 180) in which pigeons showed no tracking behavior in Experiment 2. The 12 FI 60, 4 FI 180 schedule was then administered a second time following a condition (12 FI 60, 12 FI 180) in which pigeons do show tracking. If the immediate reinforcement history of the animals determines their subsequent tracking behavior, then pigeons should not show tracking when the same 12 FI 60, 4 FI 180 follows a nontracking condition, but they should show tracking when that schedule follows a tracking condition.
Prior to this primary manipulation, the first three SW conditions in this experiment (3C–3E in Table 3) also tested a secondary hypothesis that the occasional reinforcement of the first response on a given trial (i.e., following a reinforcer) would result in reduced tracking. The first response on a trial is only reinforced if the pause exceeds the current interval criterion. Under these circumstances, the pigeons “miss” an opportunity to receive food earlier which, to avoid having reinforcement delayed in the future, may result in shortened pausing across the board (and eliminated tracking). To prevent a possible first-response reinforced effect, a slightly modified schedule was used for the first three SW conditions in this experiment: If the pigeon’s pause exceeded the scheduled interval, the first response was not reinforced and a second response was required for food delivery. Under most circumstances, this modified schedule was, from the pigeon’s point of view, indistinguishable from a standard FI schedule.
Table 3.
Conditions and Results From Each Phase in Experiment 3
| Mean pause (s) ± SEM |
||||||||
|---|---|---|---|---|---|---|---|---|
| Phase | Schedule | Sessions | Result | After FI 60 s | After FI 180 s | Tracking magnitude (s) | t | p |
| 3A | FI 60 | 7 | ||||||
| 3B | FI 180 | 15 | ||||||
| 3C | 12 FI 60, 12 FI 180a | 9 | Tracking | 15.0 ± 1.7 | 21.1 ± 2.2 | 6.1 | 6.26 | .0015 |
| 3D | 2 FI 60, 2 FI 180a | 15 | Tracking | 18.0 ± 2.3 | 22.6 ± 2.2 | 4.6 | 4.73 | .0052 |
| 3E | 1 FI 60, 1 FI 180a | 30 | No tracking | 21.8 ± 2.5 | 22.6 ± 1.9 | 0.8 | 0.91 | .406 |
| 3F | 12 FI 60, 4 FI 180 | 33 | Reduced tracking | 22.0 ± 1.9 | 24.0 ± 2.1 | 2.0 | 3.48 | .018 |
| 3G | 12 FI 60, 12 FI 180 | 11 | Tracking | 19.8 ± 1.7 | 25.0 ± 1.7 | 5.2 | 9.76 | .00019 |
| 3H | 12 FI 60, 4 FI 180 | 13 | Tracking | 18.5 ± 1.4 | 22.5 ± 1.6 | 4.0 | 10.57 | .00013 |
Note. Italics indicate statistically reliable effects. FI = fixed interval.
a On these schedules, the first response on a given trial was not reinforced. When the first response occurred after expiration of the criterion interval, a second response was required for reinforcement.
In the first two experiments, the shortest block of long intervals (apart from 1) that was used in any of the experiments was four, leaving open the possibility that repeats of two or three long intervals per cycle would also have resulted in a behavioral pattern without tracking. Thus, the fourth condition of this experiment (3D) presented pigeons with two short and two long intervals per cycle. This manipulation tests whether a single long interval per cycle is critical for the elimination of tracking, as in the first two experiments, or whether a block of two long intervals is also sufficient. The double-alternation condition also provides a particularly interesting environment for studying the interaction of discriminative cuing and temporal tracking. Each interval in this sequence provides an ambiguous cue as to the duration of the next interval—long intervals are followed by short intervals exactly half the time and vice versa. If pigeons learn this double-alternation sequence, we might see a wholly different effect (i.e., neither tracking nor nontracking) whereby the two long intervals are followed by pauses of different durations.
Method
Subjects
The same 6 pigeons (P7257, P7272, P411, P440, P7227, and P7207) from Experiments 1 and 2 participated in this experiment. P411 was returned to the experimental sequence after recovering from its illness.
Materials
The same three operant chambers, run by the same hardware and software, were used.
Procedure
FI training.
The basic procedure was identical to the one used in Experiments 1 and 2. All sessions began with a 100-s start delay followed by a 2-s food reinforcer. A further 64 reinforcers were available according to a specified FI or SW interval schedule of reinforcement. Table 3 details a full list of all the conditions in this experiment. Pigeons were first trained with a standard FI 60-s schedule for 7 sessions (Phase 3A). They were then exposed to an FI 180-s schedule for 15 daily sessions (Phase 3B).
SW training.
Pigeons were then exposed to six different SW schedules of reinforcement (see Table 3). Food was available to the pigeon for the first key peck that followed the expiration of the currently scheduled interval. For the first three SW conditions (Phases 3C to 3E), if the pigeon’s pause exceeded the scheduled interval, then the first response was not reinforced and a second response was required for food delivery. If a pigeon responded before the end of the current interval, this procedure was identical to a standard FI schedule. On those trials in which a pigeon paused for longer than the interval duration, the pigeon received what was, in effect, a tandem FT x, FR 1 schedule. These three conditions tested whether the occasional reinforcement of the first response influenced the pattern of temporal tracking. For the final three conditions (Phases 3F–3H), this additional contingency was removed and pigeons were reinforced for the first response after the criterion interval elapsed regardless of whether that peck was the first postreinforcement response.
In the first SW condition (Phase 3C), pigeons were trained with a 12 FI 60, 12 FI 180 schedule. This condition was followed by the double-alternation 2 FI 60, 2 FI 180 schedule (Phase 3D). The final four phases (Phases 3E–3H in Table 3) are the crux of this experiment. Pigeons were first trained with a nontracking condition (Phase 3E: 1 FI 60, 1 FI 180) and then transferred to the condition with previously conflicting results (Phase 3F: 12 FI 60, 4 FI 180). In the ensuing, parallel, condition pigeons were trained with a tracking condition (Phase 3G: 12 FI 60, 12 FI 180) and then transferred back to the very same condition that produced the conflicting results (Phase 3H: 12 FI 60, 4 FI 180). If the immediately preceding reinforcement history is critical in determining the subsequent pattern of tracking or nontracking behavior displayed by the pigeons, then pigeons should not track the first repeat of the 12 FI 60, 4 FI 180 schedule (3F) but should track the second repeat (3H).
Results
The primary dependent measure in this experiment was once again the postreinforcement pause. During the first period of FI 60 training (Phase 3A), pigeons paused on average for approximately one third of the interval (M = 19.7), before responding at a gradually accelerating rate through the remainder of the interval. During the subsequent FI 180 training (Phase 3B), pigeons paused longer on average, but for a smaller proportion (~one sixth) of the overall prevailing interval (M = 29.0).
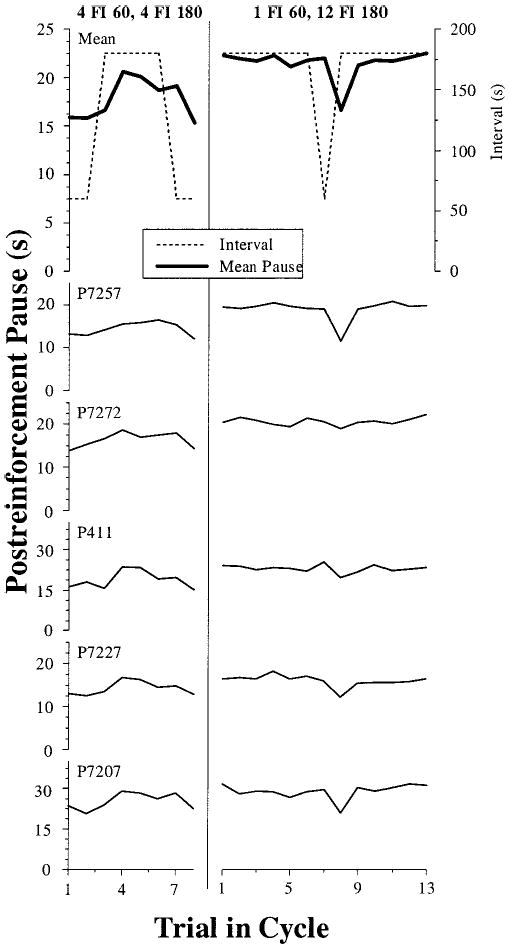
The mean postreinforcement pause as a function of trial in cycle for all six SW conditions is presented in Figure 5. The top row shows the mean; the remaining rows show data from each of the 6 individual pigeons. For the first two SW conditions (3C and 3D; Columns 1 and 2), all the pigeons showed clear temporal tracking, pausing longer following the longer intervals (FI 180) and shorter following the shorter intervals (FI 60). The robust tracking on the 2 FI 60, 2 FI 180 schedule (3D) again points to a single long interval per cycle as the critical variable for the elimination of tracking behavior. The tracking of the prevailing interval was evident following the first trial after a switch from short to long intervals and was similar following both of the longer (180-s) intervals.
Figure 5.
Postreinforcement pause as a function of trial into cycle for all 6 pigeons in all six square-wave conditions in Experiment 3. The top row plots the interval duration (dashed line) and the mean pause for all pigeons (solid line). Subsequent rows plot the average cycles for each individual pigeon. Each column represents the data from a different condition (Phases 3C–3H). FI = fixed interval.
As in Experiment 2, in the simple-alternation condition (3E), there was no difference in pausing following either short or long intervals (Column 3). On subsequent exposure to a 12 FI 60, 4 FI 180 schedule (3F), which pigeons usually track, the pigeons displayed rather limited tracking (Column 4 in Figure 5). When the pigeons were then switched to the 12 FI 60, 12 FI 180 schedule, all pigeons resumed tracking (Column 5). Finally, on the second exposure to the 12 FI 60, 4 FI 180, all 6 pigeons showed clear and pronounced tracking of the most recently experienced IFI (compare Columns 4 and 6). The tracking was less apparent following the first long interval of a cycle, but grew with repeated presentations of the longer interval.
Table 3 contains a summary of these data, showing the mean pause following all 60-s and 180-s intervals for all pigeons. The same trends that were evident in the trial-by-trial individual pigeon data recur in the mean data: Pigeons show temporal tracking on most conditions, but do not track the simple-alternation condition (3E) and show limited tracking on the subsequent 12 FI 60, 4 FI 180 schedule (3F). To check these effects statistically, we conducted a two-way repeated measures ANOVA (FI × Schedule) for the six SW conditions in this experiment. There was a main effect of FI, a main effect of schedule, and an FI × Schedule interaction (all Fs > 4.8, ps < .01). The final two columns of Table 3 display the results of planned comparisons between the two levels of FI (after FI 60 or after FI 180) for each of the six SW schedules administered in this experiment. For four of the conditions with multiple consecutive long FIs in a cycle (3C, 3D, 3G, and 3H), pigeons showed very robust temporal tracking. In the condition with a single interval of each duration per cycle (3E), pigeons did not show reliable tracking. When the 12 FI 60, 4 FI 180 followed this simple-alternation condition (3F), there was significant though less statistically robust tracking than for the other conditions with multiple long intervals per cycle. This mixed finding was probably the result of learning across sessions whereby pigeons initially did not show tracking behavior but did for the final few days of training (see Figure 6 below for more details).
Figure 6.
Session-by-session changes in mean postreinforcement pause for all pigeons in Conditions 3C–3H. On the top row, from left to right, the panels represent mean daily values for 12 fixed-interval (FI) 60, 12 FI 180 (3C); 2 FI 60, 2 FI 180 (3D); and 1 FI 60, 1 FI 180 (3E). On the bottom row, from left to right, the panels represent mean daily values for 12 FI 60, 4 FI 180 (3F); 12 FI 60, 12 FI 180 (3G); and 12 FI 60, 4 FI 180 (3H). All data points are an average of all trials (not only the first per cycle) that followed an FI 60 or an FI 180 on a given day.
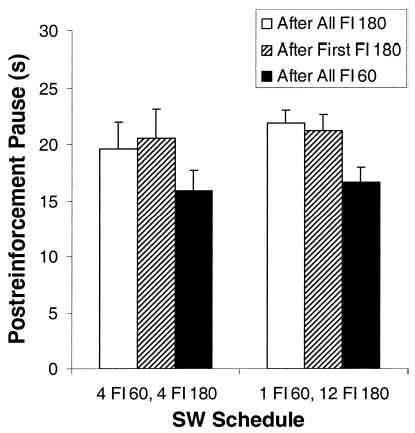
As can be seen in Table 3, tracking magnitude (mean pause after FI 180 – mean pause after FI 60) was greatest for four of the conditions with repeating blocks of multiple short and long intervals, reduced for the first repeat of the 12 FI 60, 4 FI 180 schedule and virtually nonexistent for the single-alternation condition. A one-way repeated measures ANOVA indicated a main effect of schedule, F(5, 25) = 7.36, p < .001. A planned comparison between the two repeats of the 12 FI 60, 4 FI 180 condition (3F and 3H) indicated that the magnitude of tracking was significantly greater when this condition followed a tracking condition (3H) than when it followed a nontracking condition (3F), t(5) = 3.36, p < .05.
Figure 6 depicts the development of tracking across sessions for all six SW conditions in this experiment. Each panel represents the results from a single phase and displays one line for the mean pause following an FI 60 and another line for the mean pause following an FI 180. For the first two and last two conditions (3C, 3D, 3G and 3H), in which pigeons showed very robust tracking, they displayed this tracking immediately in the first session and consistently tracked in all subsequent sessions. In the simple-alternation condition (3E), pigeons initially showed minimal tracking behavior, which disappeared after about 8–9 days and did not reappear over then next 21 sessions. Of particular note, pigeons never displayed antitracking or pausing longer after the short intervals (i.e., during the longer intervals) and vice versa, despite 30 days of training.
In the first repeat of the 12 FI 60, 4 FI 180 condition, pigeons showed little evidence of any differential pausing for the first 22 sessions. After 3 weeks, the mean pause did start to show a slight difference, perhaps explaining why pigeons did show a minimally (albeit significantly) longer pause following the longer interval duration. To determine the extent that tracking behavior changed across sessions, we divided performance on this schedule into three blocks of 11 sessions and conducted a two-way ANOVA (FI Value × Session Blocks) on the resultant data. There was a main effect of FI and an FI × Session interaction (both Fs > 4.4, ps < .05), but no main effect of session, F(2, 10) = 0.62, p = .55. A pair of planned comparisons confirmed that during the first two blocks (22 sessions), pigeons showed no tracking whatsoever, t(5) = 1.36, p = .17, whereas during the final block (11 sessions), pausing was significantly longer following the 180-s intervals, t(5) = 5.38, p < .01. In contrast, in the second repeat of the 12 FI 60, 4 FI 180 schedule (3H) that followed the tracking condition, pigeons showed clear tracking from the very first day and continued this pattern of differential pausing for the full 13 days in this phase.
Following up on these learning effects, we examined the magnitude of tracking separately during the first 13 sessions and the final 8 days (Sessions 26–33) of the first repeat of the 12 FI 60, 4 FI 180 when tracking seemed most apparent from the learning curves of Figure 6. The magnitude of tracking was significantly larger for all 13 days of the second repeat (following a tracking condition) of the 12 FI 60, 4 FI 180 schedule than for the first 13 days of the first repeat, t(5) = 2.95, p < .05. There was no difference, however, in the magnitude of tracking between the final 8 days of the first repeat and performance during the second repeat of the 12 FI 60, 4 FI 180 schedule, t(5) = 0.80, p = .45. So, although the effects of prior training with a nontracking schedule persisted for at least 13 days (and probably 22 days), they did dissipate given sufficient time.
The reinforcement schedules during the first three SW conditions (3C–3E) were modified such that if the first response following the end of the criterion interval was also the first response in a trial, then a second response was required for reinforcement. Whether this condition was detectably different from a regular SW schedule depended on the frequency of long pausing, especially during the shorter (60-s) interval. During the 9 days of exposure to the modified 12 FI 60, 12 FI 180 condition (3C), 3 pigeons never paused for longer than 60 s during an FI 60, and the remaining 3 pigeons did so once, once, and three times. The percent of all FI 60 trials that had a pause longer than 60 s was only 0.32%. Long pausing was not any more frequent during the FI 60-s components in the other two modified SW schedules (3D and 3E); the percent of trials with pauses longer than 60 s was 0.30% for the 2 FI 60, 2 FI 180 condition (3D) and 0.19% for the 1 FI 60, 1 FI 180 condition (3E). Because the pigeons paused longer than the short FI so rarely, our manipulation had practically no effect on the schedule the pigeons received and these conditions were treated as though they were ordinary SW interval schedules.
Discussion
In Experiment 3, pigeons showed temporal tracking on four different conditions (3C, 3D, 3G, and 3H) in which blocks of multiple short and long intervals were cyclically repeated. As in Experiment 2, they did not show tracking on the simple-alternation condition (3E). Moreover, as hypothesized, the immediate reinforcement history of the pigeons clearly influenced the degree of tracking on the 12 FI 60, 4 FI 180 schedule. When this schedule followed a tracking condition, pigeons showed considerable tracking; when this schedule followed a nontracking condition, pigeons showed more limited tracking. Tracking failure was apparent for the first 22 days of SW training (see Figure 6), but tracking began to emerge over the final few days. These results establish that the particular contingencies in an animal’s immediate reinforcement history can virtually eliminate tracking behavior for at least 3 weeks on the 12 FI 60, 4 FI 180 schedule that Staddon (1967, 1969) used.
In addition, the tracking behavior on the double-alternation condition (3D) confirms that the key event in triggering tracking or nontracking behavioral patterns is the presence of a single long interval in a cycle. Given the presence of history effects (see Phases 2B, 2D and 3E–3H) on these SW schedules, there remains the possibility that the tracking observed on the double-alternation condition was a carryover from the tracking observed during the previous 12 FI 60, 12 FI 180 schedule (3C). Perhaps, given long enough (or a different immediate reinforcement history), tracking on this condition might cease. Two features of the current dataset militate against this possibility. First, under all previous nontracking conditions, the effects were apparent within the first 15 days (see Phases 1F/1G, 2B, 2C, and 3E). Thus, any changes in the animal’s pausing pattern should have been apparent in the 15 days for which the double-alternation condition was run. Moreover, on the ensuing single-alternation schedule, pigeons continued to display tracking behavior for 8–9 days (compare Phase 2B), suggesting that pigeons were not learning or performing any differently on the 2 FI 60, 2 FI 180 schedule than on other schedules with multiple long intervals.
One point that we have not established so far is whether the critical single interval for discriminative cue learning need be the longer interval. Experiment 1 showed that a long interval can serve as a discriminative cue for a shorter interval, but the converse point has yet to be established. The final experiment takes up this thread and explores whether a single short interval in a cycle is sufficient to eliminate tracking.
Experiment 4
In the first three experiments, we established that temporal tracking behavior occurs on a wide variety of SW schedules, yet pigeons do not track schedules with a single long interval. On these schedules, pigeons stop tracking over the course of several days (see Phases 1F/G, 1H, 2B, 2C, 3E in Tables 1–3). When multiple short intervals are cycled with a single long interval, pigeons always show the learned, nontracking behavior. One way to conceive of these results is that this behavioral pattern reflects a balance between two processes: temporal tracking (as described by the linear-waiting equation) and discriminative cuing. Experiment 4 attempts to answer the following question: Can pigeons learn that short intervals follow long intervals and/or long intervals follow short intervals?
In this experiment, we first presented pigeons with a 4 FI 60, 4 FI 180 schedule to reestablish that these pigeons can and do show temporal tracking when confronted with this SW schedule. These pigeons then encountered a 1 FI 60, 12 FI 180 schedule. This manipulation should establish whether pigeons show temporal tracking when there is only a single short interval per cycle. If pigeons can learn that short intervals cue longer ones, then pigeons should initially show strong tracking behavior on the 1 FI 60, 12 FI 180 schedule, before a gradual attenuation to nontracking behavior across sessions. On the other hand, research on other dynamic timing schedules has indicated that transitions to shorter intervals produce much stronger tracking behavior on impulse (one anomalous interval/session) and step schedules (one transition/session) than transitions to longer intervals (Higa et al., 1991, 1993). On the basis of these asymmetrical results, the hypothesis follows that a short→long cue is more difficult for pigeons to learn. As a result, pigeons should continue to show tracking throughout exposure to the 1 FI 60, 12 FI 180 schedule, despite showing the learned nontracking behavior on the converse schedule (when 1 = long; see Phase 1H in Table 1).
Method
Subjects
The same 6 pigeons from Experiments 1–3 served as subjects in this experiment. One pigeon (P440) became ill again in the middle of the experiment and was unable to complete the two conditions. The data from this pigeon were excluded from all analyses.
Materials
The same operant chambers and operating software were used.
Procedure
The basic procedure was the same as in the previous experiments. After a 100-s start delay, pigeons received 65 reinforcers according to a SW interval schedule of reinforcement. There were two phases in this experiment. In the first condition (Phase 4A), pigeons received 10 daily sessions of a 4 FI 60, 4 FI 180 schedule—a block of 4 FI 60 s followed by a block of 4 FI 180 s in a repeating sequence. All sessions began at a random point in this sequence. In the second condition (Phase 4B), pigeons were exposed to a 1 FI 60, 12 FI 180 schedule for 35 daily sessions. All statistical summaries and tests were conducted using Microsoft Excel. A minor programming glitch during Phase 4A resulted in sessions sometimes beginning with five or six repeats of the same FI; the remainder of the sessions were unaffected. As a result, the first six (instead of four as in previous experiments) trials of each daily session were considered warm-up trials and were excluded from all analyses.
Results
Figure 7 shows the mean pause for each position in the SW cycle. The dark line in the top row represents the mean of all 5 pigeons, and the individual pigeon data are contained in the subsequent five rows. In the 4 FI 60, 4 FI 180 condition (Phase 4A: left column), all 5 pigeons show clear temporal tracking, pausing longer following the longer intervals and shorter following the shorter intervals (see also Figure 8). As can be seen in the right column, in the 1 FI 60, 12 FI 180 condition (4B), the pigeons also show a temporal tracking pattern. Three pigeons (P7227, P7257, and P7207) show very pronounced short pauses on the trial immediately after the single short interval, and the remaining 2 pigeons (P411 and P7272) show a smaller effect.
Figure 7.
Mean postreinforcement pause as a function of trial into cycle for Conditions 4A and 4B, averaged across all sessions. The top row contains the mean for all pigeons, and subsequent rows depict the data from individual pigeons. FI = fixed interval.
Figure 8.
Mean (+SEM) postreinforcement pause following fixed-interval (FI) 60 or FI 180 for Experiment 4, averaged across all sessions. The white columns represent pauses following all FI 180 s, the hatched columns represent pauses following only the first FI 180 s per trial, and the black columns represent pauses following all FI 60 s. Data from the 4 FI 60, 4 FI 180 schedule (4A) are on the left, and data from the 1 FI 60, 12 FI 180 condition (4B) are on the right. SW = square wave.
Figure 8 summarizes these data, by averaging across all trials that follow either all 60-s and 180-s intervals or the first 180-s interval of a cycle. In both conditions, pigeons show very clear temporal tracking behavior. A pair of t tests confirms the statistical reliability of this visible trend for both the 4 FI 60, 4 FI 180 schedule and the 1 FI 60, 12 FI 180 schedule (both ts > 4.2, ps < .05). This tracking effect persists when only the first trial following an interval switch is examined. As can be seen from the hatched columns of Figure 8, mean pauses during that first trial are longer following the first FI 180 s for both the 4 FI 60, 4 FI 180 condition and the 1 FI 60, 12 FI 180 condition (both ts > 3.35, ps < .05).
The development of this pausing pattern for the 1 FI 60, 12 FI 180 condition is shown in Figure 9. The two lines represent the mean pause after FI 60 or after FI 180 for every session; each line is an average of all 5 pigeons. Throughout all 35 days of training, pigeons showed temporal tracking, pausing longer following the 180-s intervals than following the 60-s interval. There seems to be little or no change in the magnitude of this pause difference over the course of sessions. A two-way ANOVA (FI Value × Session Blocks) confirmed these observations. Using seven blocks of five sessions each, we found a main effect of FI, F(1, 4) = 17.74, p < .05, and no effect of sessions or interaction (Fs < 1.6, ps > .2). Thus, pigeons reliably tracked the single short 60-s interval across all 35 days in this 1 FI 60, 12 FI 180 condition (4B).
Figure 9.
Mean postreinforcement pause following the short (60-s) and long (180-s) fixed interval (FI) for each of 35 daily sessions for the 1 FI 60, 12 FI 180 condition (Phase 4B).
Discussion
In this experiment, we showed that pigeons track a SW condition with multiple long intervals and a single short interval per cycle. Pigeons tracked the short interval from the first session, and continued to do so through five subsequent weeks of training. Unlike the converse condition with one long interval per cycle (1H), pigeons did not cease to track even after multiple encounters with this single anomalous interval duration. This finding further isolates the single long interval as the key event for producing nontracking behavior in the pigeons. The asymmetry suggests that a long interval can serve as a discriminative stimulus for a subsequent shorter cue, but the reverse does not hold true. These results also mesh nicely with previous research on the dynamics of temporal control that illustrated an asymmetry between short and long intervals with impulse (one long interval per session) and step (one switch from short to long per session) procedures (Higa et al., 1991, 1993).
General Discussion
There are four novel findings in this quartet of experiments. First, in contrast to earlier results in the literature, pigeons do show temporal tracking on many SW schedules of reinforcement. These results represent the first clear demonstration of tracking behavior on a SW schedule of reinforcement. Second, the main exception to the tracking pattern seems to occur whenever there is only a single long interval per cycle. In this unique circumstance, longer intervals are always followed by shorter ones. Under these conditions, tracking is not adaptive, and it does not occur. Third, both the tracking and nontracking seem to be learned, developing across days when pigeons are switched from a schedule that produces one of these behaviors to a schedule that produces the opposite pattern. Finally, the immediate reinforcement history of an animal can determine the extent to which it shows tracking behavior on a given schedule, thus providing a plausible explanation for why the pigeons in Staddon’s (1967, 1969) experiments failed to show temporal tracking.
According to the linear-waiting hypothesis, pigeons pause for a fixed proportion of the previously experienced IFI. The simplest, one-back version of the linear-waiting equation predicts complete temporal tracking in all conditions, no matter how few or how many consecutive intervals there are per SW cycle. This prediction accords with results from conditions in which pigeons show tracking, but fails for all conditions (see Phases 1F/1G, 1H, 2B, 2C, and 3E in Tables 1–3) in which pigeons do not show tracking. Noting other limitations to the one-back linear waiting hypothesis, Wynne et al. (1996) discuss several modifications of the linear-waiting notion that may account for a wider range of timing dynamics. One of the variants they consider is a moving-average linear-waiting model in which pausing is controlled by an average of the previous M intervals. Following their lead, we consider a linear-waiting model expanded to include the average of the previous two intervals:
| (2) |
In this version, the pause (P) on a given trial (n + 1) is equal to a fixed proportion (A) of an unweighted average of the interval durations (I) of the previous two trials (n, n − 1) plus a small constant (B). This simple equation predicts tracking whenever multiple consecutive intervals occur per cycle, but predicts attenuated tracking on the first trial after a switch (when one of the two previous trials is short and the other one is long). Both these predictions are in agreement with the current empirical results. Furthermore, this equation accounts for the nontracking behavior exhibited on simple-alternation schedules (2B, 2C, and 3E) when the previous two trials always consist of one long trial and one short trial. In this circumstance, an average of the previous two trials is always the same no matter which interval duration occurred most recently.
Although in better agreement with the data than the one-back equation, this version of the linear-waiting hypothesis also fails to account for all of the existing data. When multiple short intervals are cycled with a single long interval, the two-back model would predict some tracking—a prediction not borne out by the data (see the 4 FI 30, 1 FI 90 and 12 FI 30, 1 FI 90 conditions). Equation 2 also predicts that a single short interval in a series of longer ones will affect pausing equally in two subsequent intervals, rather than only one, as the data show (4B, see also Higa et al., 1991). As well, pigeons should show immediate tracking (in the first session) when they are switched from the 1 FI 60, 1 FI 180 schedule to the 12 FI 60, 12 FI 180 schedule in Experiment 2D, which they do not.
One potential solution within this linear-waiting framework would consist of taking into account an even larger window of past intervals (i.e., a larger M). For instance, to accommodate the nontracking behavior exhibited on the 12 FI 30, 1 FI 90 schedule (1H), this sort of model would require a window extending back at least 13 intervals. This fix runs into problems of its own, however, as any window size greater than 8 intervals back would predict no tracking on the 4 FI 30, 4 FI 90 (1E, 4A) or 2 FI 60, 2 FI 180 (3D) schedules—conditions that pigeons do indeed track. In fact, there does not exist a window size that can simultaneously account for tracking behavior on the 2 FI 60, 2 FI 180 (3D) and the nontracking behavior on the 12 FI 30, 1 FI 90 (1H) and 4 FI 30, 1 FI 90 (1F/G) schedules. In addition, the sequential learning effects (Experiment 2) and history effects (Experiment 3) are beyond the scope of this account. Thus, this dataset definitively rules out any simple N-back model of temporal tracking for interval schedules.
Perhaps what pigeons are learning when they are exposed to these schedules is to shift from one-back tracking to many-back tracking and use a larger sample of previous intervals in determining their pause on a given cycle. Such a two-fold formulation would offer the benefits of both linear-waiting models considered so far. In some sense, this way of thinking about these data is equivalent to assuming that the pigeons operate in one of two modes that can be likened to one-back linear waiting and long-term averaging. When the interval duration is changing very rapidly (e.g., every trial in the simple-alternation situations), pigeons use the long-term averaging strategy. When there is relative stability in the pattern of IFIs (with blocks of more than one identical interval), pigeons use the one-back strategy. Under certain conditions, pigeons will switch between these two pausing strategies. The history effects demonstrated in Experiment 3 can then be understood as an instance when, because of the reinforcement history, pigeons stayed in the long-term averaging mode in a situation in which they ordinarily would have followed the linear-waiting strategy. For this model to provide a full account of tracking, a rigorous specification of the transition rules that determine which strategy a pigeon would use is necessary. Two other empirical problems faced by this account are (a) the asymmetry between the “one-short” and “one-long” conditions (Phases 1H and 4B), which does not fit either one-back tracking or long-term averaging and (b) the evident fact that animals can anticipate (i.e., show phase lag zero) under some conditions (Church & Lacourse, 1998; Innis & Staddon, 1971).
As an alternative to these variants on linear waiting, we suggested that behavior on these cyclic schedules may be controlled by a combination of a linear-waiting process and the discriminative cuing effects of the interval. The asymmetry between pausing when there is either a single short (Phase 4B) or a single long interval (Phase 1H, 1F/1G) per cycle strongly suggests that pigeons are learning a discriminative cuing strategy with two independent cues. To account for the asymmetrical patterns of responding seen under these two conditions, the key assumption that we need to make is that cuing “long is followed by short” is more salient (learnable) than cuing “short is followed by long.” This supposition is defensible because when the short interval ends and the reinforcer is received, the previous reinforcer (i.e., the end of the previous interval) is more proximate in time. Under this framework, pigeons are readily able to learn that a long interval cues the subsequent short interval and thus stop tracking (as the cuing counteracts the tracking) the 12 FI 60, 1 FI 180 schedule (1H). In fact, with sufficient training, or with a small enough absolute interval duration of the shorter interval, pigeons might also be expected to show anticipatory behavior on a 12/1 or 4/1 SW schedule. In contrast, pigeons have greater difficulty learning that the short interval cues the subsequent long interval, and thus they persist on tracking the 1 FI 60, 12 FI 180 indefinitely (4B).
A particularly interesting outcome in Experiments 2 and 3 is that, on the three single-alternation schedules (Phases 2B, 2C and 3E), pigeons never seem to fully learn the single-alternation pattern. That is, they never perform what would seem to be the optimal behavior: pause longer following the short interval (i.e., during the long interval) and shorter following the long interval (i.e., during the short interval). Under other conditions, pigeons do show single-alternation learning for spatial location or color (Hearst, 1962; Williams, 1971). Keller (1973) even showed single-alternation learning for interval duration when the short and long intervals are 20 s and 180 s respectively. According to the discriminative-cue account that we are offering, the lack of tracking on these 1/1 schedules reflects a balance between an obligatory linear-waiting process and discriminative effects of the longer interval. As a result, any manipulation that increases the strength of the discriminative stimulus effect should decrease, abolish, or reverse tracking. Increasing the ratio between the short and the long stimulus would likely enhance the cuing effect and this discriminative stimulus account would expect reverse tracking (i.e., full anticipation) in that situation.
Two particular features of our dataset pose the greatest problems to this two-process (cuing/waiting) account. First, there is a perfect transition from Phase 2B to 2C; pigeons begin the 1 FI 60, 1 FI 180 condition right where they left off the 1 FI 30, 1 FI 90 condition—with no tracking behavior (see Figure 4). Even with some transfer/generalization between the two schedules, we would expect the new discriminative cue (the 180-s interval) to require some learning before affecting behavior. For conditions with the same pairs of intervals, the discriminative cue remains the same across successive conditions, thus explaining the residual effects observed in Phase 2C and perhaps Phase 3F. With a new pair of intervals, however, a discriminative cue should require some new learning and thus a learning curve (albeit truncated) would be expected for the first handful of sessions on Condition 2C. On the other hand, an averaging account would expect perfect transfer to the new interval durations.
Second, the gradual increase in pausing sometimes seen across multiple long intervals (see Phases 1A, 1F/G, 2A, and 3H in Tables 1–3) is not easily reconciled with this approach. This difficulty arises because it is not clear what sort of discriminative cue (if any), pigeons might learn in this situation. One possibility is that pigeons learn that longer intervals cue other longer intervals, and thus discriminative cuing and tracking would act in concert. The gradual increase in pause across long intervals could then be attributed to decreased memory interference from recent short intervals. However, we know from the condition with only a single short interval per cycle (1 FI 30, 12 FI 90; Phase 4B) that pigeons seem unable to learn that shorter intervals can serve as cues for longer intervals, even when a longer interval follows 100% of the time. As a result, it seems improbable that pigeons would be able to learn a predictive cue for the longer interval in this partially ambiguous situation (long intervals only follow long intervals 11 times out of 12). From the asymmetry observed in the two 1/12 conditions (1H and 4B), a more likely discriminative cue for the pigeons to be able to learn is that short intervals occasionally follow longer intervals. On a 12 FI 60, 12 FI 180 schedule, for example, presuming the animals cannot count to 12 exactly, the discriminative cue effect should increase across successive long intervals, thereby reducing (not increasing) the pause. In other words, as more long intervals pass, the more likely the next one is to be short, and the shorter the pause should be. This prediction clearly does not accord with the data (see Figures 1 and 3). Any sort of moving average of previous intervals is a better candidate explanation for these two features of the data (full transfer and gradual increase) than a combination of discriminative effects and linear waiting.
The dependency of temporal tracking on the immediately preceding SW schedule demonstrated in Experiment 3 offers insight into the role history effects can play in determining behavior on schedules of reinforcement. Behavior analysts often allude to the imperviousness of schedule effects to particular reinforcement histories (e.g., Ferster & Skinner, 1957), but rarely is evidence adduced to substantiate that point. There is currently a small body of mixed evidence for reinforcement history effects on FI performance (Baron & Leinenweber, 1995; Cole, 2001; Freeman & Lattal, 1992; Tatham, Wanchisen, & Yasenchack, 1993; Wanchisen, Sutphin, & Balogh, 1998; Wanchisen, Tatham, & Mooney, 1989). For example, Wanchisen et al. (1989) found reduced scalloping on an FI 30-s schedule that was preceded by a VR 20 schedule, and Tatham et al. (1993) found increased postreinforcement pausing on an FI 30 s that followed a differential-reinforcement-of-low-rate (DRL) 60-s schedule. In contrast, Cole (2001) argued that all previously demonstrated history effects were the result of insufficient training; Cole showed that after 80–100 sessions of exposure to an FI schedule, all effects of previous schedules (FR or DRL) had dissipated. In the condition in which we found history effects (3F), we ran the experiment for only 33 days. Thus, there is the definite possibility that after 80–100 sessions, all history effects might be abolished. This extrapolation is further supported by our observation that the pausing pattern for the final eight sessions of this condition was no different than during the second repeat of the 12 FI 60, 4 FI 180 schedule. Nevertheless, there was still a clear difference in the pattern of responding for the first 3 weeks of training, establishing that a reinforcement history with one variant on an FI schedule can produce relatively long-lasting effects on performance on a different type of multiple FI schedule.
On the basis of the data collected in these 4 experiments, we have proposed two different explanatory accounts for performance on these SW interval schedules: a cuing/tracking balance and a transition between one-back tracking and long-term averaging. Each of these ideas better explains a certain subset of the dataset, but neither can account for the whole range of phenomena observed here. We have managed, however, to bring performance on SW interval schedules—performance that was previously thought of as anomalous—into better accord with the data from CI schedules and other dynamic timing schedules. Finally, the learning curves for temporal tracking behavior demonstrate how some forms of temporal control can be obscured or even negated by discriminative cuing, sequence learning, or other nontemporal factors. These findings suggest that “timing” is not the whole story when examining behavior on interval schedules of reinforcement.
Acknowledgments
We thank Dan Cerutti, Jérémie Jozefiewiez, Sasha Hart, Ron Parr, Karen Skinazi, Ted Hall, and Güven Güzeldere for insightful comments on earlier versions of this article.
References
- Baron A, Leinenweber A. Effects of a variable-ratio conditioning history on sensitivity to fixed-interval contingencies in rats. Journal of the Experimental Analysis of Behavior. 1995;63:97–110. doi: 10.1901/jeab.1995.63-97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng K, Westwood R. Analysis of single trials in pigeons’ timing performance. Journal of Experimental Psychology: Animal Behavior Processes. 1993;19:56–67. [Google Scholar]
- Church RM, Lacourse DM. Serial pattern learning of temporal intervals. Animal Learning & Behavior. 1998;26:272–289. [Google Scholar]
- Church RM, Meck WH, Gibbon J. Application of scalar timing theory to individual trials. Journal of Experimental Psychology: Animal Behavior Processes. 1994;20:135–155. doi: 10.1037//0097-7403.20.2.135. [DOI] [PubMed] [Google Scholar]
- Cole MR. The long-term effect of high- and low-rate responding histories on fixed-interval responding in rats. Journal of the Experimental Analysis of Behavior. 2001;75:43–54. doi: 10.1901/jeab.2001.75-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crystal JD, Church RM, Broadbent HA. Systematic nonlinearites in the memory representation of time. Journal of Experimental Psychology: Animal Behavior Processes. 1997;23:267–282. doi: 10.1037//0097-7403.23.3.267. [DOI] [PubMed] [Google Scholar]
- Dukich TD, Lee AE. A comparison of measures of responding under fixed-interval schedules. Journal of Experimental Analysis of Behavior. 1973;20:281–290. doi: 10.1901/jeab.1973.20-281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferster, C. B., & Skinner, B. F. (1957). Schedules of reinforcement Acton, MA: Copley Publishing Group.
- Freeman TJ, Lattal KA. Stimulus control of behavioral history. Journal of the Experimental Analysis of Behavior. 1992;57:5–15. doi: 10.1901/jeab.1992.57-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hearst E. Delayed alternation in the pigeon. Journal of the Experimental Analysis of Behavior. 1962;5:225–228. doi: 10.1901/jeab.1962.5-225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrnstein RJ, Morse WH. Effects of pentobarbitol on intermittently reinforced behavior. Science. 1957 May 10;125:929–931. doi: 10.1126/science.125.3254.929-a. [DOI] [PubMed] [Google Scholar]
- Higa JJ. Dynamics of time discrimination: II. The effects of multiple impulses. Journal of the Experimental Analysis of Behavior. 1996;66:117–134. doi: 10.1901/jeab.1996.66-117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higa JJ, Thaw JM, Staddon JER. Pigeons’ wait-time responses to transitions in interfood-interval duration: Another look at cyclic schedule performance. Journal of the Experimental Analysis of Behavior. 1993;59:529–541. doi: 10.1901/jeab.1993.59-529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higa JJ, Wynne CDL, Staddon JER. Dynamics of time discrimination. Journal of Experimental Psychology: Animal Behavior Processes. 1991;17:281–291. doi: 10.1037//0097-7403.17.3.281. [DOI] [PubMed] [Google Scholar]
- Innis, N. K. (1981). Reinforcement as input: Temporal tracking on cyclic interval schedules. In M. L. Commons & J. A. Nevin (Eds.), Quantitative analysis of behavior (Vol. 1, pp. 257–286). Cambridge, MA: Ballinger.
- Innis NK, Staddon JER. Sequential effects in cyclic-interval schedules. Psychonomic Science. 1970;19:313–315. [Google Scholar]
- Innis NK, Staddon JER. Temporal tracking on cyclic-interval reinforcement schedules. Journal of the Experimental Analysis of Behavior. 1971;16:411–423. doi: 10.1901/jeab.1971.16-411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keller, J. (1973). Responding maintained by sinusoidal cyclic-interval schedules of reinforcement: A control-systems approach to operant behavior Unpublished doctoral thesis, University of Maryland, College Park.
- Kello JE, Staddon JER. Control of long-interval performance on mixed cyclic-interval schedules. Bulletin of the Psychonomic Society. 1974;4:1–4. [Google Scholar]
- Schneider BA. A two-state analysis of fixed-interval responding in the pigeon. Journal of the Experimental Analysis of Behavior. 1969;12:677–687. doi: 10.1901/jeab.1969.12-677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staddon JER. Attention and temporal discrimination: Factors controlling responding under a cyclic-interval schedule. Journal of the Experimental Analysis of Behavior. 1967;10:349–359. doi: 10.1901/jeab.1967.10-349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staddon JER. Multiple fixed-interval schedules: Transient contrast and temporal inhibition. Journal of the Experimental Analysis of Behavior. 1969;12:583–590. doi: 10.1901/jeab.1969.12-583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staddon, J. E. R. (2001). Adaptive dynamics: The theoretical analysis of behavior. Cambridge, MA: MIT Press/Bradford.
- Staddon JER, Higa JJ. Temporal learning. Psychology of Learning and Motivation. 1991;27:265–294. [Google Scholar]
- Staddon JER, Higa JJ. Time and memory: Towards a pacemaker-free theory of interval timing. Journal of the Experimental Analysis of Behavior. 1999;71:215–251. doi: 10.1901/jeab.1999.71-215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- StatSoft. (2001). STATISTICA (Version 6.0) [Computer Software]. Tulsa, OK: Author.
- Tatham TA, Wanchisen BA, Yasenchack MP. Effects of differential-reinforcement-of-low-rate schedule history on fixed-interval responding. Psychological Record. 1993;43:289–297. [Google Scholar]
- Wanchisen BA, Sutphin GE, Balogh SA. Lasting effects of behavioral history of low-rate responding in rats. Learning and Motivation. 1998;29:220–235. [Google Scholar]
- Wanchisen BA, Tatham TA, Mooney SE. Variable-ratio conditioning history produces high- and low-rate fixed-interval performance in rats. Journal of the Experimental Analysis of Behavior. 1989;52:167–179. doi: 10.1901/jeab.1989.52-167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams BA. Color alternation learning in the pigeon under fixed-ratio schedules of reinforcement. Journal of the Experimental Analysis of Behavior. 1971;15:129–140. doi: 10.1901/jeab.1971.15-129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wynne CDL, Staddon JER. Typical delay determines waiting time on periodic-food schedules: Static and dynamic tests. Journal of the Experimental Analysis of Behavior. 1988;50:197–210. doi: 10.1901/jeab.1988.50-197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wynne CDL, Staddon JER, Delius JD. Dynamics of waiting in pigeons. Journal of the Experimental Analysis of Behavior. 1996;65:603–618. doi: 10.1901/jeab.1996.65-603. [DOI] [PMC free article] [PubMed] [Google Scholar]