

Abstract
The oomycetous, fungus-like, aquatic organism Pythium insidiosum is the etiologic agent of pythiosis, a life-threatening infectious disease of humans and animals that has been increasingly reported from tropical, subtropical, and temperate countries. Human pythiosis is endemic in Thailand, and most patients present with arteritis, leading to limb amputation and/or death, or cornea ulcer, leading to enucleation. Diagnosis of pythiosis is time-consuming and difficult. Radical surgery is the main treatment for pythiosis because conventional antifungal drugs are ineffective. The aims of this study were to evaluate the use of Western blotting for diagnosis of human pythiosis, to identify specific immunodominant antigens of P. insidiosum, and to increase understanding of humoral immune responses against the pathogen. We performed Western blot analysis on 16 P. insidiosum isolates using 12 pythiosis serum samples. These specimens were derived from human patients with pythiosis who had different forms of infection and lived in different geographic areas throughout Thailand. We have identified a 74-kDa immunodominant antigen in all P. insidiosum isolates tested. The 74-kDa antigen was also recognized by sera from all patients with pythiosis but not by control sera from healthy individuals, patients with thalassemia, and patients with various infectious diseases, indicating that Western blot analysis could facilitate diagnosis of pythiosis. Therefore, the 74-kDa antigen is a potential target for developing rapid serodiagnostic tests as well as a therapeutic vaccine for pythiosis. These advances could lead to early diagnosis and effective treatment, crucial factors for better prognosis for patients with pythiosis.
Pythiosis is an emerging and fatal infectious disease caused by the oomycetous fungus-like, aquatic organism Pythium insidiosum. It is the only Pythium species of the kingdom Chromista (Stramenopila) known to infect humans and animals, such as horses, dogs, cats, cattle, and sheep (8, 13, 25). The disease has been reported from tropical, subtropical, and temperate areas of the world (8, 13, 21). Phylogenic analysis has shown that P. insidiosum is more closely related to diatom and golden-brown algae than to fungi (11, 29). P. insidiosum exists in two stages: biflagellate zoospore and right-angle-branching hyphae (3). The zoospore is the infective unit, and it swims to attach and germinate as hyphae in host tissue (14).
The first 2 cases of human pythiosis were discovered in Thailand in 1985 (3, 7). Since then, cases have been increasingly reported (2, 7, 10, 20, 23, 26-28, 31, 32). Three forms of human pythiosis have been described: (i) cutaneous/subcutaneous pythiosis affecting face or limbs as a granulomatous, ulcerating, or cellulitic lesion, (ii) systemic (vascular) pythiosis affecting arteries of lower extremities resulting in arterial occlusion or aneurysm, and (iii) ocular pythiosis causing corneal ulcers or keratitis (7, 27). Systemic and ocular pythiosis are the most common forms of infection. Most of the systemic patients had the affected limb amputated. Likewise, most of the ocular patients had the infected eye removed. Moreover, many systemic patients died from a ruptured aneurysm. Thalassemias and an agriculture-related career in a swampy area are the predisposing factors (7, 8, 23, 32). Diagnosis by culture identification is time-consuming and requires expertise (17). Serodiagnostic (enzyme-linked immunosorbent assay and immunodiffusion) and PCR-based diagnostic tests have been developed, but diagnostic materials are not widely available (5, 9, 19, 30). Since conventional antifungal drugs are not effective in treating pythiosis, surgery is then the main treatment option. However, postsurgical relapse can occur and cause further damage and death. Recently, immunotherapy with the P. insidiosum vaccine prepared from crude extracts of P. insidiosum increased a curative rate to 72% of horses with pythiosis but just 33% and 50% of dogs and humans with pythiosis, respectively (15, 31). Therefore, improvements in the diagnosis and treatment of pythiosis are still important healthcare goals.
In this study, we performed Western blot analyses of P. insidiosum extracts using sera from Thai patients with pythiosis to evaluate the potential use of Western blots for the diagnosis of human pythiosis, to identify specific immunodominant antigens of P. insidiosum, and to increase understanding of humoral immune responses against this pathogen. Since early diagnosis and effective treatment are crucial factors for better outcome of patients with pythiosis, the specific immunodominant antigens identified could be targets for developing rapid serodiagnostic tests as well as a more effective vaccine.
MATERIALS AND METHODS
Microorganisms.
Sixteen culture-proven Pythium insidiosum isolates (P1 to P16) from Thai patients with pythiosis (Table 1; Fig. 1) were included for Western blot analysis. In addition, Pythium aphanidermatum (MCC19-1) and Pythium deliense (MCC19-2), which are closely related to P. insidiosum and nonpathogenic species, as well as Conidiobolus coronatus (MCC19-3) and Rhizopus sp. (MCC19-4), which are zygomycetous fungi that share microscopic features with P. insidiosum, were also included in this study.
TABLE 1.
Pythium insidiosum isolates used in this study
| Isolatea | Reference no. | Infected tissue |
|---|---|---|
| P1 | RAPI-0201 | Artery |
| P2 | SIMI149-41 | Artery |
| P3 | SIMI7874 | Artery |
| P4 | SIMI8659 | Artery |
| P5 | SIMI8727 | Artery |
| P6 | MMC44P21-1 | Artery |
| P7 | CUMF45 | Artery |
| P8 | MMC45P21-1 | Brain tissue |
| P9 | MMC44P21-2 | Cutaneous, artery |
| P10 | CBS673.85 | Cutaneous |
| P11 | CUMF42 | Cutaneous |
| P12 | MMC45P21-2 | Cornea |
| P13 | SIMI3306-4 | Cornea |
| P14 | RAPI-0202 | Cornea |
| P15 | SIMI9642 | Cornea |
| P16 | RAPI-0001 | Stomach, ileum |
Isolates P1, P14, and P16 were provided by the Department of Pathology, Faculty of Medicine, Ramathibodi Hospital, Mahidol University; isolates P2 to P5, P10, P13, and P15 were provided by the Department of Microbiology, Faculty of Medicine, Siriraj Hospital, Mahidol University; isolates P6, P8, P9, and P12 were provided by the Department of Microbiology, Faculty of Medicine, Chiang Mai University; and isolates P7 and P11 were provided by the Department of Microbiology, Faculty of Medicine, Chulalongkorn University.
FIG. 1.
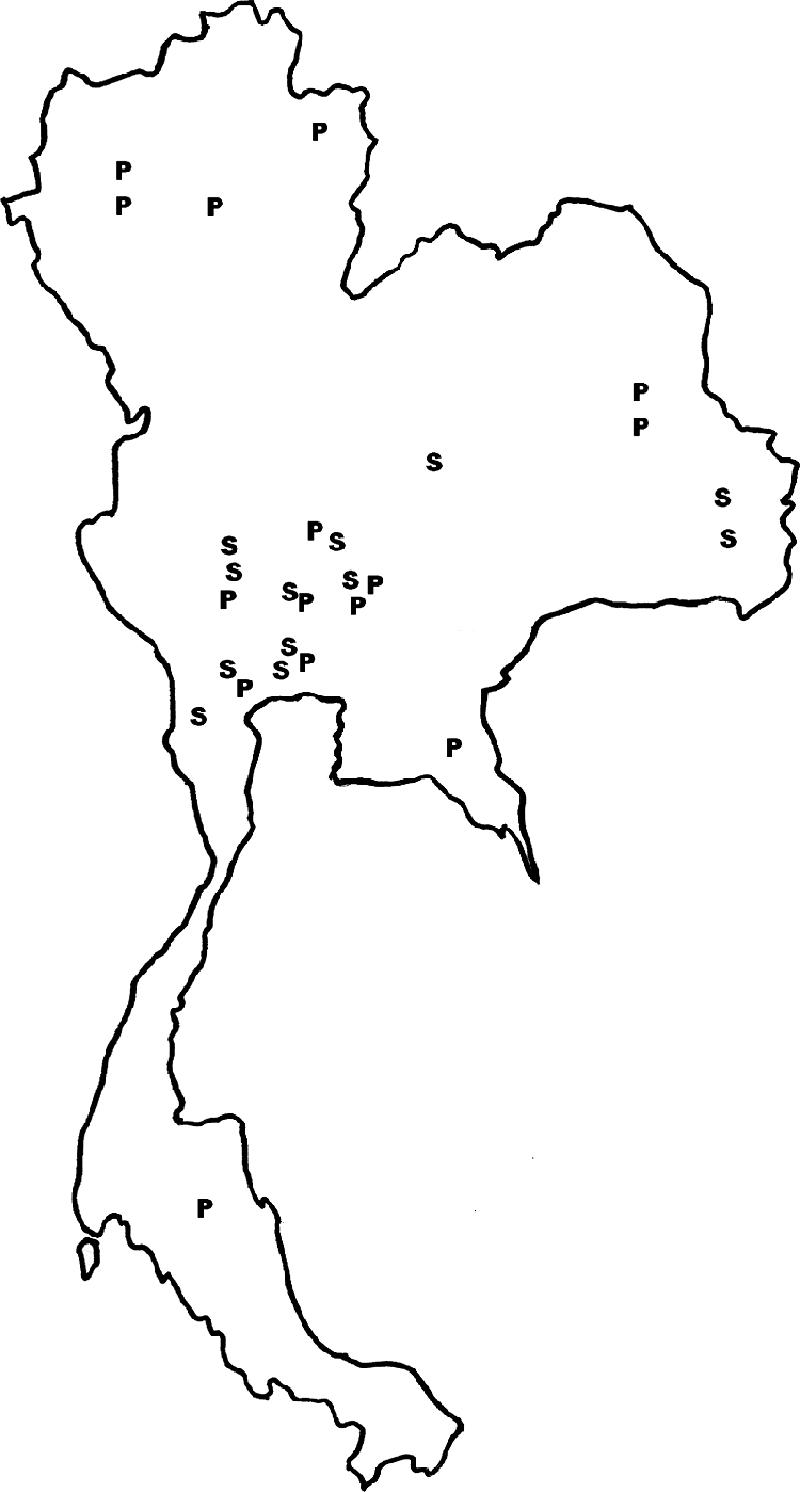
Map of Thailand showing geographic distribution of patients with pythiosis as sources of Pythium insidiosum isolates (P) and sera (S).
Antigen preparations.
Each isolate of P. insidiosum was subcultured on Sabouraud dextrose agar and incubated at 37°C for 2 days. Several small agar pieces, containing hyphal elements from the growing culture, were transferred to 200 ml of Sabouraud dextrose broth and shaken (100 rpm) at 37°C for 1 week. Merthiolate (final concentration, 0.02% [wt/vol]) was added to kill the cultures before they were filtered through a Durapore membrane filter (0.22-μm pore size; Millipore, County Cork, Ireland). The filtered broth containing phenylmethylsulfonyl fluoride (PMSF; 0.1 mg/ml) and EDTA (0.3 mg/ml) is referred to as culture filtrate antigen (CFA). The retained hyphal mass was washed once with distilled water and then quickly transferred to a prechilled mortar in the presence of 25 ml of cold sterile distilled water containing PMSF (0.1 mg/ml) and EDTA (0.3 mg/ml). The hyphal mass was ground on ice to produce 100% ruptured cells before being centrifuged at 4,800 × g, 4°C for 10 min. The resulting supernatant, referred to as soluble antigen from broken hyphae (SABH), was filtered through a Durapore membrane filter (0.22-μm pore size). SABHs, CFAs, and sterile Sabouraud dextrose broth were then concentrated ∼80-fold using an Amicon Ultra 15 centrifugal filter (10,000 nominal molecular weight limit; Millipore, Bedford, MA).
Culture portions of well-grown P. aphanidermatum, P. deliense, C. coronatus, and Rhizopus sp. were washed from Sabouraud dextrose agar slants with 10 ml of sterile distilled water and transferred to 500 ml of Sabouraud dextrose broth and shaken (100 rpm) for up to a month at 37°C (except P. deliense cultures were grown at room temperature). Merthiolate (final concentration, 0.02% [wt/vol]) was added, and culture broths were filtered through a Durapore membrane filter (0.22-um pore size). Then, 25 ml of filtrated broth containing PMSF (0.1 mg/ml) and EDTA (0.3 mg/ml) was mixed with the corresponding hyphal mass in a prechilled mortar for grinding until 100% rupture was achieved. The homogenates were then centrifuged at 4,800 × g, 4°C for 10 min. The resulting supernatant was filtered through a Durapore membrane filter (0.22-μm pore size) and concentrated ∼80-fold by using an Amicon Ultra-15 centrifugal filter (10,000 nominal molecular weight limit) and is referred to as concentrated antigen mixture (CAM). Protein concentration was determined using the Bradford assay (1). All CFAs, SABHs, and CAMs were stored at −20°C until use.
Sera.
Sera from 12 Thai patients with pythiosis were used in this study (Table 2; Fig. 1). Sera from 3 patients who received a full course of P. insidiosum vaccine (2 injections of 100 to 200 μl [each] of 2-mg/ml vaccine at a 14-day interval) (31) were collected before and after (3 weeks for S12, 1 month for S4, and 3 months for S7) vaccinations. Another 23 sera were included as controls: 6 serum samples from healthy blood donors who came to the Blood Bank Division, Ramathibodi Hospital; 5 serum samples from healthy thalassemic patients; 1 serum sample (each) from patients with aspergillosis, zygomycosis, histoplasmosis, candidiasis, cryptococcosis, leptospirosis, amoebiasis, and toxoplasmosis; and 1 serum sample (each) from anti-hepatitis B virus-positive, anti-hepatitis C virus-positive, anti-human immunodeficiency virus-positive, and highly positive antinuclear antibody individuals. All sera were stored at −20°C until use.
TABLE 2.
Clinical data for pythiosis patient sera used in this study
| Patient | Infected tissue | Duration of symptoms | Treatment | Clinical outcome |
|---|---|---|---|---|
| S1 | Artery | 5 mo | Blood clot retraction, vaccination | Disease progressed |
| S2 | Artery | 3 wk | Vaccination | Initial improvement, relapse |
| S3 | Artery | 1 mo | BKa amputation | Cured |
| S4 | Artery | 3 mo | AKb amputation, vaccination | Dead |
| S5 | Cutaneous | 4 days | Debridement | Lost in follow-up |
| S6 | Artery | 5 mo | AK amputation | Being followed up |
| S7 | Artery | 1 mo | AK amputation, vaccination | Dead |
| S8 | Artery | 3 mo | AK amputation | Cured |
| S9 | Artery | 2 wk | AK amputation | Cured |
| S10 | Artery | 7 wk | AK amputation | Cured |
| S11 | Artery | 2 mo | BK amputation, vaccination | Cured |
| S12 | Artery | 10 days | BK amputation, vaccination | Cured |
BK, below the knee.
AK, above the knee.
SDS-PAGE.
Ten microliters of SABH, CFA, or CAM concentrate, containing up to 20 μg protein, was mixed with loading buffer (60 mM Tris-HCl, pH 6.8, 25% glycerol, 2% sodium dodecyl sulfate [SDS], 14.4 mM mercaptoethanol, and 0.1% bromophenol blue), boiled for 5 min, and centrifuged at 13,800 × g for 5 min. Supernatant proteins were separated at 100 V in an SDS-polyacrylamide gel (4% stacking gel, 8 or 12% separating gel) on a minigel apparatus (Bio-Rad, Hercules, CA). Prestained SDS-polyacrylamide gel electrophoresis (PAGE) molecular weight standards (Bio-Rad) were run in parallel. The gel was stained with Coomassie brilliant blue.
Western blotting.
Separated antigens were transferred from SDS-PAGE gels to polyvinylidene difluoride membranes (Bio-Rad) for 1 h at 100 V. Membranes were blocked with 5% gelatin in Tris-buffered saline (TBS), pH 7.5, and incubated overnight at room temperature with sera diluted 1:1,000 with antibody buffer (TBS, pH 7.5, 0.05% Tween 20, 1% gelatin). The membranes were washed twice with washing buffer (TBS, pH 7.5, 0.05% Tween 20) and incubated at room temperature for 2 h with goat anti-human immunoglobulin G (heavy plus light chains) conjugated with horseradish peroxidase (Bio-Rad) at a 1:3,000 dilution with antibody buffer. The membranes were washed with washing buffer three times. Signals were developed by adding a fresh mixture of 10 ml of 0.3% 4-chloro-1-naphthol in methanol and 50 ml of TBS with 30 μl of 30% H2O2. Reactions were stopped by immersing the membranes in distilled water. Color-developed membranes were protected from light and immediately photographed.
India ink staining.
Blotted membranes were washed two times in TBS (pH 7.5) with 0.5% Tween 20 for 5 min and incubated at room temperature for 3 h in India ink diluted 1:1,000 with distilled water. The membrane was rinsed several times with distilled water.
RESULTS
Identification of immunodominant antigens of P. insidiosum.
We studied the protein profile and immunogenicity of SABHs as cytoplasmic antigens and CFAs as secretory/culture supernatant antigens of P. insidiosum isolated from patients who had different forms of infection (Table 1) and live in different areas throughout Thailand (Fig. 1). Proteins of SABHs from 16 P. insidiosum isolates appeared mostly below 120 kDa in the Coomassie blue-stained SDS-PAGE gel (Fig. 2A). Most of the proteins were transferred onto a polyvinylidene difluoride membrane during blotting and were visualized by India ink staining (Fig. 2B). Blotted membranes were immunostained with S2 (Fig. 2C), S12 (Fig. 2D), and S4 (data not shown) sera. An 87-kDa immunoreactive antigen was observed in SABHs of 15, 13, and 15 isolates after probing with S2, S12, and S4 sera, respectively. When probed with S4 or S12 serum (Fig. 2D), several immunoreactive antigens ranging from 43 to 34 kDa were also noted in all SABHs, except for P13 SABH from an patient who lived in the south of Thailand (Fig. 1). However, when probed with S2 serum, these 43- to 34-kDa antigens were faint or undetectable (Fig. 2C). In contrast to the 87- and 43- to 34-kDa antigens, which were found in many but not all isolates, a 74-kDa antigen was strongly detected in all 16 SABHs tested. Western blots of CFAs from 6 P. insidiosum isolates (P1, P6 to P8, P10, and P15) were probed with S12 serum, and strong bands were detected at 74 kDa and 43 to 34 kDa (Fig. 3).
FIG. 2.

Coomassie brilliant blue-stained SDS-PAGE gel (A), India ink-stained blotted membrane (B), and Western blot analysis (C, D) of Pythium insidiosum P1 to P16 SABH (lane 1 to 16, respectively) probed with S2 (C) and S12 (D) pythiosis serum. Molecular markers (A, B) or predicted sizes (C, D) are shown in kDa on the left.
FIG. 3.
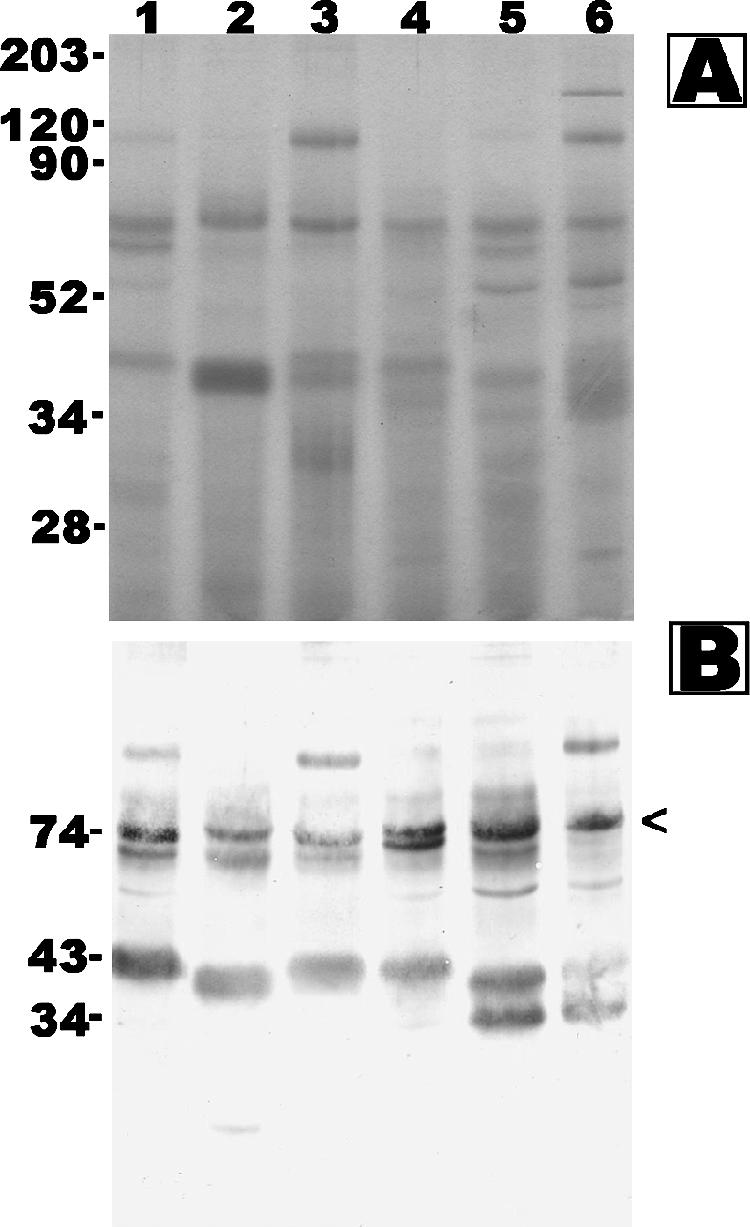
Coomassie brilliant blue-stained SDS-PAGE gel (A) and Western blot analysis (B) of Pythium insidiosum CFAs P1 (lane 1), P15 (lane 2), P10 (lane 3), P6 (lane 4), P8 (lane 5), and P7 (lane 6) probed with S12 pythiosis serum. Molecular markers (A) or predicted sizes (B) are shown in kDa on the left.
Specificity and sensitivity of immunodominant antigens of P. insidiosum.
The immunodominant antigens of P. insidiosum were also evaluated for specificity using 23 control sera from healthy individuals, patients with thalassemia, and patients with various infectious diseases. When SABH and CFA of the P1 isolate were probed with the serum from a patient with either thalassemia or zygomycosis at a 1:1,000 dilution, the 87-, 74-, 32-, and 28-kDa antigens of P1 SABH reacted weakly, while no such cross-reaction was observed in P1 CFA, even when probed at a 1:100 dilution (data not shown). To test whether the immunodominant antigens of P. insidiosum can be broadly recognized by sera from other patients with pythiosis, sera from 12 patients were used to probe against P1 CFA (Fig. 4). All 12 pythiosis sera recognized the 74-kDa antigen, while only 5 sera (S1, S4, S6, S8, and S12) weakly recognized antigens ranging from 43 to 34 kDa. In addition, the 74-kDa antigen was the only one recognized by S5, S9, and diluted S12.
FIG. 4.

Western blot analysis of Pythium insidiosum P1 CFA probed with S1 to S12 pythiosis sera (lane 1 to 12, respectively) at 1:1,000 dilution. Lanes 13 and 14 represent S12 serum at 1:10,000 and 1:100,000 dilutions, respectively. Predicted sizes are shown in kDa on the left.
Comparison of cytoplasmic and secretory immunogens of P. insidiosum.
The S12 serum appears to detect a subset of proteins in the P1 CFA preparation relative to the P1 SABH sample (Fig. 5, lanes 1 and 2). Additionally, the intensity of detection of the 74-kDa antigen is higher for the P1 CFA sample than for P1 SABH. Although the 74-kDa antigen of P1 SABH and P1 CFA appears to be the same size in the 12% SDS-PAGE gel (Fig. 5, lanes 1 and 2), only the 74-kDa antigen of P1 SABH showed cross-reaction with the two control sera (above). This suggests that the 74-kDa band, present in Western blots of P1 SABH and P1 CFA, may represent different antigens. To test this hypothesis, we increased the resolution of protein separation by decreasing the gel concentration to 8% and increasing the separation time for SDS-PAGE and found that the 74-kDa antigens of P1 CFA migrated slightly faster than those of P1 SABH (Fig. 5, lanes 3 and 4). In contrast, the antigens ranging from 43 to 34 kDa of P1 SABH and P1 CFA had banding patterns similar to that seen in the 12% gel.
FIG. 5.
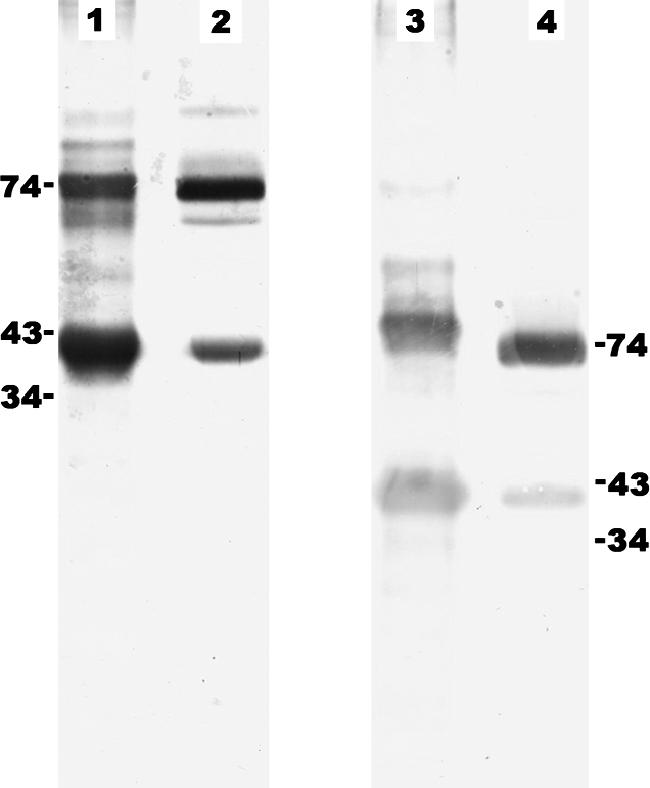
Western blot analysis of Pythium insidiosum P1 SABH (lanes 1 and 3) and Pythium insidiosum P1 CFA (lanes 2 and 4) probed with S12 pythiosis serum. Proteins were separated in 12% (lanes 1 and 2) and 8% (lanes 3 and 4) SDS-PAGE gels. Predicted sizes (kDa) are shown at both sides.
Comparison of immunogenicity of Pythium spp. and zygomycetous fungi.
Protein profiles and immunogenicity of nonpathogenic Pythium species and zygomycetous fungi were also studied and compared with those of P. insidiosum. Protein profiles of both SABH and CFA of the P. insidiosum P1 isolate were different from CAMs (mixture of cytoplasmic and culture supernatant proteins) of P. deliense, P. aphanidermatum, C. coronatus and Rhizopus sp. (Fig. 6A). Western blots probed with S12 demonstrated at least nine antigens (>203, 100, 87, 74, 66, 54, and 43 to 34 kDa) of P1 SABH, at least four antigens (100, 74, 66, and 42 kDa) of P1 CFA, at least two antigens (138 and 100 kDa) of P. aphanidermatum CAM, and at least six antigens (>203, 165, 109, 94, 85, and 32 kDa) of P. deliense CAM (Fig. 6B). A 100-kDa antigen was observed in extracts of both P. aphanidermatum and P. insidiosum. No immunoreactive antigen was found in concentrated Sabouraud dextrose broth or in CAMs of C. coronatus and Rhizopus sp.
FIG. 6.
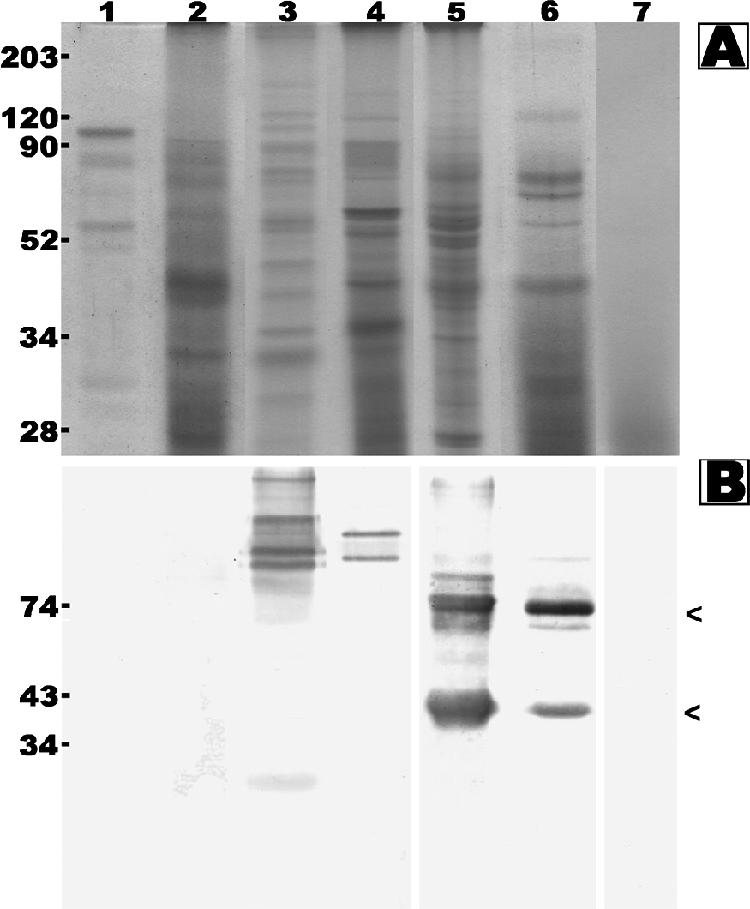
Coomassie brilliant blue-stained SDS-PAGE gel (A) and Western blot analysis (B) of C. coronatus CAM (lane 1), Rhizopus sp. CAM (lane 2), Pythium deliense CAM (lane 3), Pythium aphanidermatum CAM (lane 4), Pythium insidiosum P1 SABH (lane 5), Pythium insidiosum P1 CFA (lane 6), and concentrated Sabouraud dextrose broth (lane 7) probed with S12 pythiosis serum. Molecular markers (A) and predicted sizes (B) are shown in kDa on the left.
Investigation of a P. insidiosum vaccine mechanism.
The P. insidiosum vaccine is prepared from a mixture of SABH and CFA of P. insidiosum isolated from a horse with pythiosis and shows a curative effect in treating humans with pythiosis (28, 31). It has been proposed that the vaccine works by introducing previously unexposed antigens, which stimulate host protective immunity (4, 18). If this immunity results from the production of antibodies not present during an infection, then it may be possible to detect the antigens by Western blotting. Therefore, we probed SABH and CFA of the P1 isolate with preimmunized versus postimmunized sera from 3 patients who received full courses of the vaccine and found that the positions and numbers of bands were similar, although the band intensity was higher with postimmunized sera (data not shown).
DISCUSSION
Mendoza and coworkers used sera from five Costa Rican horses with pythiosis and three horses cured of pythiosis by immunotherapy, together with P. insidiosum isolates from four horses and one Thai patient (CBS 673.85), in Western blot analyses and demonstrated 32-, 30-, and 28-kDa immunodominant antigens (16). However, Santurio et al., in a study of the serological response to P. insidiosum antigens in a rabbit model of pythiosis, failed to detect these three antigens (22). Recently, Vanittanakom et al. showed that 110-, 73-, 56-, 42- to 35-, 30- to 28-, 26-, and 23-kDa antigens were immunoreactive with serum from a human with pythiosis, with strong reactivity against the 42- to 35-kDa antigens (30). In our study using 16 P. insidiosum isolates (including CBS 673.85) and 12 sera from Thai patients with pythiosis (Table 1; Fig. 1), we have identified the 74-kDa immunodominant antigen in all isolates tested (Fig. 2 and 3). We also found immunodominant antigens ranging from 43 to 34 kDa, but the molecular size and intensity varied among the isolates and sera tested (Fig. 2, 3, and 4). These results together suggest that the humoral immune response to P. insidiosum antigens could vary from host to host (human, horse, or rabbit) or among strains of the pathogen.
Schurko et al. performed phylogenetic analysis using the internal transcribed spacer and 5.8S rRNA sequences of 23 P. insidiosum isolates from around the world (24). They found that all of the isolates were more closely related to each other than to other Pythium species. P. insidiosum was classified into 3 distinct clades according to geographic distribution: clade I consisted of isolates from the Americas, clade II consisted of isolates from Asia and Australia, and clade III consisted of isolates from Thailand and the United States. Interestingly, clade III appeared to be the most distantly related of the three clades, which may indicate that it forms a distinct subspecies (24). Recently, Rivierre et al. reported the first case of pythiosis from Africa (21). They showed that the causative organism's internal transcribed spacer and 5.8S rRNA sequences were phylogenetically far different from other oomycetous organisms but closest to the P. insidiosum clades, suggesting a new Pythium species. Therefore, the spectrum of mammalian pathogenic Pythium species could be broader than just P. insidiosum. The different subgroups of the pathogenic Pythium species found in the phylogenetic analysis could explain differences in immunogenicity, as demonstrated by Mendoza et al. (16) and by our group. Additionally, no animal cases have been reported from Thailand, where P. insidiosum is widespread (Fig. 1). These observations could be hints that strain variability of the pathogen may explain why human pythiosis specifically occurs in Thailand. Molecular analysis of P. insidiosum Thai isolates is under way to further clarify the relationship between the phylogeny of the pathogen, geographic distribution, and clinical presentation of its hosts.
Although close in size, the 74-kDa antigen of CFA is slightly smaller than that of SABH (Fig. 5). Since the 74-kDa antigen of CFA mainly accumulated in cultured broth, it is possible that the 74-kDa antigen of CFA was a secretory or shedding molecule of a truncated 74-kDa antigen of SABH. On the other hand, they could be different proteins. Either one of these hypotheses could explain why control sera from thalassemic and zygomycosis patients weakly cross-react with the 74-kDa antigen of SABH but not CFA. Because of the cross-reaction, the 74-kDa antigen of SABH would not be suitable for developing serodiagnostic tests. In contrast, the 74-kDa antigen of CFA had no cross-reaction with any of the control sera (even at 1:100 dilution), it was recognized by all of the pythiosis sera, and it was the only antigen recognized by S5, S9 (from patients with a short history of symptoms, 4 days and 2 weeks, respectively), and S12 sera (at a dilution as high as 1:100,000) (Table 2; Fig. 4). These results demonstrate that the 74-kDa antigen of CFA is a sensitive and specific antigen and that identifying this antigen by Western blot analysis could facilitate diagnosis of human pythiosis. Moreover, this antigen could be a candidate for developing convenient and rapid serodiagnostic tests, such as latex agglutination or paper chromatographic strip tests.
Although P. insidiosum has hyphal morphology, it is not a true fungus. Its ultrastructure, biochemistry, and phylogeny are quite different from those of true fungi (11, 29). Zygomycetous fungi (C. coronatus and Rhizopus sp.) were included in our study because their morphologies (broad and right-angle branching hyphae) are similar to that of Pythium species, which make it difficult to differentiate P. insidiosum from these fungi in infected tissue. In our study, proteins in CAMs of these two fungi did not cross-react with the serum from a human patient with pythiosis (Fig. 6), while a 44-kDa antigen cross-reaction with C. coronatus to sera from horses with pythiosis has been reported by Mendoza et al. (16). There are at least two possibilities to explain why we did not find a cross-reaction with C. coronatus. First, the C. coronatus strains used in our study were different from the one used by Mendoza et al., and antigens prepared from different strains could be recognized by host humoral immunity differently. Second, human and horse antibodies could respond differently to the same proteins. Nevertheless, developing and evaluating immunoperoxidase- or immunofluorescent-based tissue staining assays using anti-P. insidiosum antibodies may prove useful for facilitating histodiagnosis of human pythiosis.
Among Pythium spp., P. aphanidermatum and P. deliense are phylogenetically close to P. insidiosum (12, 24). Moreover, P. aphanidermatum can grow at 34 to 36°C, which is close to human body temperature (24). To our knowledge, no non-P. insidiosum Pythium spp. causes infection in mammals. Interestingly, the 74-kDa band and bands ranging from 43 to 34 kDa in both SABH and CFA of P. insidiosum were distinct from these 2 Pythium species (Fig. 6). This observation raises the possibility that these immunoreactive antigens of P. insidiosum may be associated with pathogenesis of P. insidiosum. In other eukaryotic pathogens, virulence factors were identified as immunoreactive antigens and some play important roles in pathogenesis or in modulating the host immune response (6). Therefore, study of the structure, function, and encoding genes could elucidate the role that these immunodominant antigens play in P. insidiosum infection.
In horses with pythiosis, infected animals often die if left untreated. However, an immunotherapeutic vaccine prepared from CFA of a P. insidiosum strain isolated from a horse surprisingly cured ∼50% of horses (4). Later on, Mendoza et al. found that the vaccine efficacy in horses with pythiosis was ∼70% after adding 32-, 30-, and 28-kDa immunodominant antigens to the original CFA vaccine (4). In humans with pythiosis, just half of all cases had favorable outcomes after vaccination (31). The results from our study suggest that enriching the 74- and 43- to 34-kDa antigens in the original vaccine could increase its efficacy in treating human pythiosis. Two hypotheses have been proposed regarding the mechanism of the vaccine effect. First, the vaccine, as an exogenous antigenic source, could introduce previously unexposed antigens which stimulate host protective immunity against the pathogen (4, 18). However, from our findings, there was no evidence to support this hypothesis, as no additional bands were observed with the postimmunized sera. Second, the vaccine could modulate an immune pattern switching from a T helper 2 to a T helper 1 response, which could lead to the effective host defense against the infection (15, 31). Characterization of differences in levels of certain subclasses of antibodies and cytokines in the pre- and postimmunized sera could be a first step in elucidating such a mechanism.
In conclusion, human pythiosis has high morbidity and mortality. The lack of early detection for new cases and an effective treatment for affected patients leads to a poor prognosis for affected individuals. In this study, we demonstrated that Western blot analysis can facilitate diagnosis of human pythiosis by identifying a specific 74-kDa immunodominant antigen of P. insidiosum. Future work could take advantage of this finding, so that the 74-kDa antigen could form the basis for rapid serodiagnostic tests and for improving the efficacy of the P. insidiosum vaccine. In addition, this 74-kDa antigen could open the door to exploration of the biology and pathogenesis of P. insidiosum as well as host responses to infection by P. insidiosum.
Acknowledgments
This work was supported by the Ramathibodi Foundation (T.K.), by the Faculty of Medicine, Ramathibodi Hospital, Mahidol University (research grant ID09-44-17/02; T.K.), and by the Thailand Research Fund (research grant TRG4580031; T.K.). T.K. was a recipient of the 2002 UNESCO and American Society for Microbiology Traveling Award toward a Pythium insidiosum study hosted by Leonel Mendoza, Michigan State University.
We thank Bruce S. Klein and Thomas D. Sullivan for reviewing the manuscript. We also thank Malai Vorachit, Pimpan Tadthong, Wanla Kulwichit, Chusana Suankratay, Poj Intalapaporn, Sawet Srisiri, Sineenart Kalnauwakul, Wanchai Wanachiwanawin, Mawin Wongsaisuwan, Sansanee Chaiyaroj, Runglawan Chawengkittikul, Porntip Poonswad, Vithaya Meevoothisom, Kavi Ratanabanangkoon, Kanda Kaslo, Lertlakana Bhoopat, Peerayut Sitthichaitakul, and Kanong Angkananukul for suggestions and material support.
REFERENCES
- 1.Bollag, D., M. Rozycki, and S. Edelstein. 1996. Protein concentration determination, p. 62-67. In D. Bollag, M. Rozycki, and S. Edelstein (ed.), Protein methods, 2nd ed. Wiley-Liss, Inc., New York, N.Y.
- 2.Chetchotisakd, P., C. Pairojkul, O. Porntaveevudhi, B. Sathapatayavongs, P. Mairiang, K. Nuntirooj, B. Patjanasoontorn, O. T. Saew, A. K. Chaiprasert, and M. R. Haswell-Elkins. 1992. Human pythiosis in Srinagarind Hospital: one year's experience. J. Med. Assoc. Thai. 75:248-254. [PubMed] [Google Scholar]
- 3.De Cock, A. W., L. Mendoza, A. A. Padhye, L. Ajello, and L. Kaufman. 1987. Pythium insidiosum sp. nov., the etiologic agent of pythiosis. J. Clin. Microbiol. 25:344-349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dixon, D. M., A. Casadevall, B. Klein, L. Mendoza, L. Travassos, and G. S. Deepe, Jr. 1998. Development of vaccines and their use in the prevention of fungal infections. Med. Mycol. 36:57-67. [PubMed] [Google Scholar]
- 5.Grooters, A. M., and M. K. Gee. 2002. Development of a nested polymerase chain reaction assay for the detection and identification of Pythium insidiosum. J. Vet. Intern. Med. 16:147-152. [DOI] [PubMed] [Google Scholar]
- 6.Hogan, L., B. Klein, and S. Levitz. 1996. Virulence factors of medically important fungi. Clin. Microbiol. Rev. 9:469-488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Imwidthaya, P. 1994. Human pythiosis in Thailand. Postgrad. Med. J. 70:558-560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kaufman, L. 1998. Penicilliosis marneffei and pythiosis: emerging tropical diseases. Mycopathologia 143:3-7. [DOI] [PubMed] [Google Scholar]
- 9.Krajaejun, T., M. Kunakorn, S. Niemhom, P. Chongtrakool, and R. Pracharktam. 2002. Development and evaluation of an in-house enzyme-linked immunosorbent assay for early diagnosis and monitoring of human pythiosis. Clin. Diagn. Lab. Immunol. 9:378-382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Krajaejun, T., R. Pracharktam, S. Wongwaisayawan, M. Rochanawutinon, M. Kunakorn, and S. Kunavisarut. 2004. Ocular pythiosis: is it under-diagnosed? Am. J. Ophthalmol. 137:370-372. [DOI] [PubMed] [Google Scholar]
- 11.Kwon-Chung, K. J. 1994. Phylogenetic spectrum of fungi that are pathogenic to humans. Clin. Infect. Dis. 19:S1-7. [DOI] [PubMed] [Google Scholar]
- 12.Martin, F. N. 2000. Phylogenetic relationships among some Pythium species inferred from sequence analysis of the mitochondrially encoded cytochrome oxidase II gene. Mycologia 92:711-727. [PubMed] [Google Scholar]
- 13.Mendoza, L., L. Ajello, and M. R. McGinnis. 1996. Infection caused by the oomycetous pathogen Pythium insidiosum. J. Mycol. Med. 6:151-164. [Google Scholar]
- 14.Mendoza, L., F. Hernandez, and L. Ajello. 1993. Life cycle of the human and animal oomycete pathogen Pythium insidiosum. J. Clin. Microbiol. 31:2967-2973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mendoza, L., W. Mandy, and R. Glass. 2003. An improved Pythium insidiosum-vaccine formulation with enhanced immunotherapeutic properties in horses and dogs with pythiosis. Vaccine 21:2797-2804. [DOI] [PubMed] [Google Scholar]
- 16.Mendoza, L., V. Nicholson, and J. F. Prescott. 1992. Immunoblot analysis of the humoral immune response to Pythium insidiosum in horses with pythiosis. J. Clin. Microbiol. 30:2980-2983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mendoza, L., and J. Prendas. 1988. A method to obtain rapid zoosporogenesis of Pythium insidiosum. Mycopathologia 104:59-62. [DOI] [PubMed] [Google Scholar]
- 18.Miller, R. I. 1981. Treatment of equine phycomycosis by immunotherapy and surgery. Aust. Vet. J. 57:77-382. [DOI] [PubMed] [Google Scholar]
- 19.Pracharktam, R., P. Changtrakool, B. Sathapatayavongs, P. Jayanetra, and L. Ajello. 1991. Immunodiffusion test for diagnosis and monitoring of human pythiosis insidiosi. J. Clin. Microbiol. 29:2661-2662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Prasertwitayakij, N., W. Louthrenoo, N. Kasitanon, K. Thamprasert, and N. Vanittanakom. 2003. Human pythiosis, a rare cause of arteritis: case report and literature review. Semin. Arthritis Rheum. 33:204-214. [DOI] [PubMed] [Google Scholar]
- 21.Rivierre, C., C. Laprie, O. Guiard-Marigny, P. Bergeaud, M. Berthelemy, and J. Guillot. 2005. Pythiosis in Africa. Emerg. Infect. Dis. 11:479-481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Santurio, J. M., A. T. Leal, A. B. Leal, R. Festugatto, I. Lubeck, E. S. Sallis, M. V. Copetti, S. H. Alves, and L. Ferreiro. 2003. Three types of immunotherapics against pythiosis insidiosi developed and evaluated. Vaccine 21:2535-2540. [DOI] [PubMed] [Google Scholar]
- 23.Sathapatayavongs, B., P. Leelachaikul, R. Prachaktam, V. Atichartakarn, S. Sriphojanart, P. Trairatvorakul, S. Jirasiritham, S. Nontasut, C. Eurvilaichit, and T. Flegel. 1989. Human pythiosis associated with thalassemia hemoglobinopathy syndrome. J. Infect. Dis. 159:274-280. [DOI] [PubMed] [Google Scholar]
- 24.Schurko, A. M., L. Mendoza, C. A. Levesque, N. L. Desaulniers, A. W. de Cock, and G. R. Klassen. 2003. A molecular phylogeny of Pythium insidiosum. Mycol. Res. 107:537-544. [DOI] [PubMed] [Google Scholar]
- 25.Tabosa, I. M., F. Riet-Correa, V. M. Nobre, E. O. Azevedo, J. L. Reis-Junior, and R. M. Medeiros. 2004. Outbreaks of pythiosis in two flocks of sheep in northeastern Brazil. Vet. Pathol. 41:412-415. [DOI] [PubMed] [Google Scholar]
- 26.Tanphaichitra, D. 1989. Tropical disease in the immunocompromised host: melioidosis and pythiosis. Rev. Infect. Dis. 11:S1629-S1643. [DOI] [PubMed] [Google Scholar]
- 27.Thianprasit, M., A. Chaiprasert, and P. Imwidthaya. 1996. Human pythiosis. Curr. Top. Med. Mycol. 7:43-54. [PubMed] [Google Scholar]
- 28.Thitithanyanont, A., L. Mendoza, A. Chuansumrit, R. Pracharktam, J. Laothamatas, B. Sathapatayavongs, S. Lolekha, and L. Ajello. 1998. Use of an immunotherapeutic vaccine to treat a life-threatening human arteritic infection caused by Pythium insidiosum. Clin. Infect. Dis. 27:1394-1400. [DOI] [PubMed] [Google Scholar]
- 29.Tyler, B. 2001. Genetics and genomics of the oomycete-host interfact. Trends Genet. 17:611-614. [DOI] [PubMed] [Google Scholar]
- 30.Vanittanakom, N., J. Supabandhu, C. Khamwan, J. Praparattanapan, S. Thirach, N. Prasertwitayakij, W. Louthrenoo, S. Chiewchanvit, and N. Tananuvat. 2004. Identification of emerging human-pathogenic Pythium insidiosum by serological and molecular assay-based methods. J. Clin. Microbiol. 42:3970-3974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wanachiwanawin, W., L. Mendoza, S. Visuthisakchai, P. Mutsikapan, B. Sathapatayavongs, A. Chaiprasert, P. Suwanagool, W. Manuskiatti, C. Ruangsetakit, and L. Ajello. 2004. Efficacy of immunotherapy using antigens of Pythium insidiosum in the treatment of vascular pythiosis in humans. Vaccine 22:3613-3621. [DOI] [PubMed] [Google Scholar]
- 32.Wanachiwanawin, W., M. Thianprasit, S. Fucharoen, A. Chaiprasert, N. Sudasna, N. Ayudhya, N. Sirithanaratkul, and A. Piankijagum. 1993. Fatal arteritis due to Pythium insidiosum infection in patients with thalassaemia. Trans. R. Soc. Trop. Med. Hyg. 87:296-298. [DOI] [PubMed] [Google Scholar]