

As Shakespeare wrote in Much Ado About Nothing, “…there was never yet philosopher, That could endure the toothache patiently…” Indeed, perhaps the fundamental existential human experience is suffering. Under unbearable pain or distress, even the staunchest advocates of solipsism will eventually rescind their conviction that the world is only an illusion. What makes suffering so powerful in convincing us that we and the world are real is the fact that we experience it; we do not merely observe it. “I experience, therefore I am” would thus describe the indisputable human condition more appropriately than the classical statement “I think, therefore I am.”
Under unbearable pain or distress, even the staunchest advocate of solipsism will eventually rescind their conviction that the world is only an illusion
The latter statement was the foundation on which the seventeenth century French philosopher René Descartes built his conception of the world. For Descartes (also known as Cartesius), thinking was the exclusive capacity of the incorporeal mind, the soul, which he defined as a thinking substance. Moreover, he claimed that only humans have a soul, and only they can reason; all other organisms, including monkeys as he explicitly posited, were mere automatons built of flesh and bones. Descartes did not deny life to animals or equate them with human made machines, as has often been imputed to him. In contrast to inanimate machines, animals had corporeal spirits, which humans had too, and exhibited sensations and passions. Yet in Descartes' view, the behaviour of non human animals could easily be explained solely by the function and constitution of their organs; in contrast to humans, they did not have any capacity for pure thought free from natural impulses and passions. In his argument, he referred to the ancient Greek philosopher Epiktetos, who stated that you are not your body—your body is just finely moulded clay. To explain how the soul caused human reasoning, Descartes proposed an intriguing mechanism: the human mind commands the body. For some time, this dualism of mind and body was the intellectual basis on which philosophers, and later biologists, tried to explain consciousness and free will (Rose, 2005).
Descartes preceded Isaac Newton by 50 years and Charles Darwin by 230 years. His mechanistic explanation of how the body works and how the mind communicates with the brain was ingenious for his time. To understand the impact of Descartes' theory, it is worthwhile remembering that chemistry as a science did not exist in his day. At that time, the clock was the most sophisticated instrument and thus provided the best model for the workings of the body. There was little effective medicine, and analgesia and anaesthesia had not yet been invented. Neglect, contempt and rejection of the body, and the identification of the ‘I', of oneself, with a pure soul, might have been a cunning way for the noblest and wisest humans to belittle the daily humiliation imparted on them by bodily troubles, and to preserve their serenity and dignity. This attitude can be traced back as far as the Greek philosopher Plato. Classical rationalism, which is a major current of European thought and the procreator of Western science, might have had one of its sources here.
Neglect, contempt and rejection of the body, and the identification of the ‘I', of oneself, with a pure soul, might have been a cunning way for the noblest and wisest humans…to preserve their serenity and dignity
A modern incarnation of Descartes' dualism might be the ‘computational model' of cognition: the mind is like a software program working on the hardware of the brain. This model has dominated cognitive science—or, as George Miller (2003) prefers to call it, cognitive sciences, in the plural—for the past few decades. A large part of the community of cognitive scientists still regards cognition as the exclusive property of humans, with the human mind being the organ of conscious perception, thinking and memory, busily processing information. The mind is no longer incorporeal, the brain is its “structural and functional realization” (Miller, 2003); however, in principle, there is no reason why its representational and computational capacities cannot be embodied in other kinds of ‘hardware', including the computer. This is why the fields of computer science and artificial intelligence have become an important part of cognitive sciences. The clock has been replaced by the computer as the model for the brain and even for life as a whole.
At least three contemporary conceptual shifts could represent radical departures from Cartesian dualism. The first is the notion of ‘affective revolution', which assumes that emotions are neither unimportant nor impeding contaminations of cognition, but rather are an inseparable or even central part of it. Consciousness might therefore be more a matter of emotion than of reasoning; the brain is no longer seen as a ‘computer', but, instead, the entire body is regarded as the organ of cognition. One of the leading protagonists of affective revolution, Antonio Damasio, entitled his book Descartes' Error, as it essentially does away with Descartes' body–soul dualism (Damasio, 1994). Second, the role of unconsciousness, which was once accentuated by Sigmund Freud, is being reinterpreted: conscious deliberation might represent just a ‘monomolecular layer' on the immense ‘ocean' of what Wilson (2002) called ‘the adaptive unconscious'. Third, cognition is being analysed with an evolutionary perspective, under the name of ‘evolutionary epistemology'. This approach states that cognition is not an exclusive capacity of humans, but that life itself is equated with cognition, and biological evolution as a whole is the evolution of cognition (Radnitzky & Bartley, 1987).
…conscious deliberation might represent just a ‘monomolecular layer' on the immense ‘ocean' of…‘the adaptive unconscious'
Evolutionary epistemology represents a ‘horizontal' approach to understanding cognition, based on the assumption that cognition arose together with the origins of life—as was famously expressed by one of its founders, Karl Popper. “From the amoeba to Einstein, the growth of knowledge is always the same…” (Popper, 1979). How about adopting a ‘vertical' approach that is orthogonal to that of evolutionary epistemology? This would start with a conscious human, and ascend to community, society and eventually the entire universe (Fig 1). Conversely, it would also proceed in the opposite direction, descending through tissues and cells to single molecules. At which level would the concept of cognition—with its inventory of consciousness, cogitation, feeling, perception, sensing and living—lose meaning and correspond only to lifeless interactions?
Figure 1.
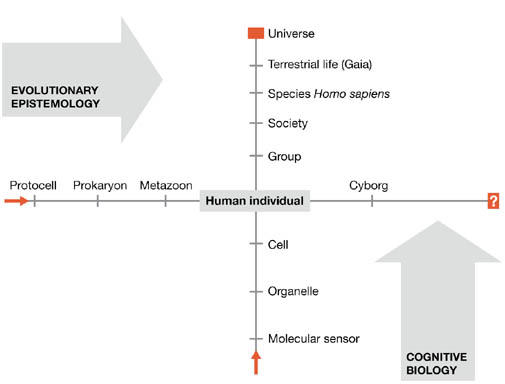
The various levels of cognition. The horizontal axis represents the levels described by evolutionary epistemology, and the vertical axis represents the levels described by cognitive biology. Cognitive biology is therefore an approach to understanding cognition that is orthogonal to evolutionary epistemology.
This vertical approach to analysing cognition has been named ‘cognitive biology' (Kovác, 2000) and could complement evolutionary epistemology. One of its fundamental postulates is the principle of minimum complexity or Delbrück's principle—named in honour of Max Delbrück, who laid the foundation of modern genetics not by studying complex human heredity but by investigating simple bacteriophages. Incidentally, Delbrück also studied cognition and behaviour using simple fungi (Phycomyces) as model organisms.
Brian Goodwin, who is reported to have invented the term ‘cognitive biology' (Goodwin, 1978), descended even further down the scale and regarded cognition as arising from the purposeful interaction of molecules. But it was probably Jacques Monod who was the first to propose that cognition can take place at the level of single molecules—specifically proteins, as their ability to discriminate between substrates or ligands might represent a form of cognition (Monod, 1970).
The current fashion of modelling cognition, or even life as a whole, has often seduced cognitive scientists into believing that they need not worry about details such as energy and physical appearance—instead, they have dispensed with the austere universe of thermodynamic constraints and focused on what they consider to be fundamental questions of information flow. Only recently has the idea gained ground that cognition is embodied, and that embodiment cannot be bracketed. For cognitive biology, this means that cognition is not only embodied, but that this embodiment has a specific form: life as we know it, natural life, is based on the rules of chemistry, and cognition is therefore a property of a chemical system. Logical possibility does not necessarily equal thermodynamic feasibility. Even virtual life, which thrives on the computer screens of those who model natural life, is embodied, although its representation is not chemical but electromagnetic. This also applies to potential artificial life, which might soon be invented by humans: it does not need to be founded on chemical principles, but might well function in the form of electromagnetic and even purely mechanical systems. Some science fiction authors have described such a mechanic artificial life based on Charles Babbage's difference engine (Gibson & Sterling, 1991).
In an authoritative review of the history of cognitive sciences, Miller (2003) presented an instructive picture of six disciplines, which presumably formed the field of cognitive sciences. Chemistry was absent from his scheme. But, in fact, it would be appropriate to place it at the centre, if we describe natural life and cognition as one of its attributes (Fig 2). To paraphrase Galileo, who said that nature is written in the language of mathematics, biologists should keep in mind that life is written in the language of chemistry.
Figure 2.
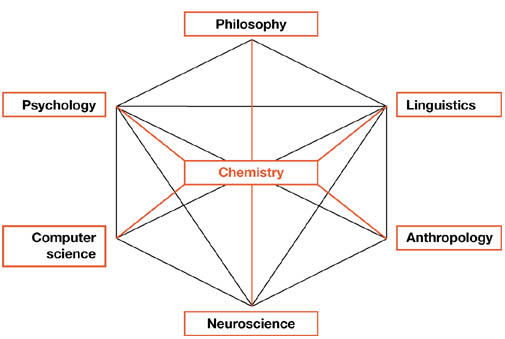
Scientific disciplines involved in cognitive sciences. Modified from Miller (2003). The original scheme of Miller is shown in black, and the modifications are shown in red.
As thermodynamics textbooks state, “chemical processes are the simplest of various natural processes … fluxes are directed to the state of equilibrium and do not flow in space coordinates but in coordinates of the composition of the system” (Bazarov, 1983). Thus, chemical processes are scalar. Centuries ago, alchemists used to say that compounds do not react unless dissolved (“Corpora non agunt nisi soluta”), and even one of the founding fathers of modern biochemistry, Otto Warburg, was convinced that “where structure begins, biochemistry ends” (Kornberg, 1989).
However, this is not the case. In contrast to chemical reactions, biochemical processes are not scalar, but are intrinsically vectorial. As Peter Mitchell described (1961), “vectorial metabolism is represented by a network of spatio-temporal pathways along which ligands (including solutes, ions, chemical groups, electrons, catalytic compounds and complexes) are conducted by articulated movements that occur in the direction of the thermodynamically natural escaping tendency, corresponding to the vectorial (or higher tensorial order) resultants of the thermodynamic and field-effect forces acting on the ligands.” Mitchell's accomplishment might have been a breakthrough in biology that was no less important than Charles Darwin's theory of evolution.
Biochemical vectoriality resides in the nature and structure of proteins. A native protein is a spatially asymmetric construction, in which many electrostatic, hydrogen, hydrophobic and van der Waals attractions add up to a strong internal attractive force that determines its structure. Owing to these forces, a typical protein would be an extremely rigid substance, perhaps stronger than steel. However, these strong chemical forces are balanced by the equally-strong dispersive force of entropy. This energy–entropy compensation makes a protein a uniquely labile and, at the same time, remarkably robust structure. Frauenfelder et al (2003) explained that “a protein does not exist in a unique conformation but can assume a very large number of somewhat different conformations or conformational substates.” In fact, “if a protein had just a single conformation, it could not function and would be dead like a stone.” The conformational sub-states are not random. Each of the proteins we encounter in nature is a product of evolution; it has been selected to perform a goal-directed teleonomic function (Monod, 1970).
For most natural proteins, the function begins with the-specific binding of a ligand. But it is not the protein molecule as a rigid structure that selects the appropriate ligand. There are constant structural changes between sub-states, even in the absence of a ligand. When the ligand is present, it binds to one particular sub-state that the protein molecule is able to adopt—it is therefore possible to say that a protein molecule exhibits exploratory ‘behaviour'. This intrinsic goal-directed plasticity of the protein molecule can be dubbed ‘molecular sentience', and it is this sentience that makes a protein a ‘living' molecule. Traditionally, this privilege has been attributed to DNA, but, according to Lewontin (1992), DNA is a ‘dead' molecule, one of the least reactive. Robert Rosen (1991) anchored biology entirely in what August Weismann called the soma: he stated that the soma is what is alive. Sentience—the-capacity to exhibit a variety of potential internal states, which respond to the immediate state of the environment—might therefore constitute the essence of life.
It is this purpose, built into the protein structure, which allows us to call protein–ligand interaction ‘molecular recognition'. Because of this intrinsic teleonomy, a protein gives meaning and significance to its environment—that is, to its ligand. By contrast, nomic interactions of individual atoms and molecules, such as chemical reactions in the inanimate world, with no evolutionary history, are inevitable, deterministic, timeless and do not represent cognition.
However, molecular recognition by a protein molecule is only part of the story. Recognition is followed by an action. A ligand is a signal. In contrast to standard chemical interactions, binding energy is not fully dissipated as heat, but is used partly for molecular work—a specific pre-programmed change in the conformation of the protein. In this way, the signal is transmitted from one site on the protein to another. The transmission takes place in four-dimensional space, as it involves time as a coordinate, and this process gives biochemistry its vectoriality. The exploitation of binding energy was originally recognized in enzymatic catalysis, in which a portion of the binding energy acts to lower the activation energy of a reaction (Jencks, 1975), but it can be expanded to explain the work of translocators, receptors and transcription factors.
By receiving and transmitting signals, proteins execute a complete working cycle in an ‘all or nothing' fashion. It is appropriate to regard most protein molecules as molecular engines. Hence, molecular cognition consists of molecular sensation—which has two inseparable aspects, recognition and signifaction—and molecular action. As Monod (1970) pointed out, by binding two or more ligands, some proteins can bring them together not on thermodynamic, but exclusively on logical, grounds—the principle of gratuity. By selective binding, proteins also associate with each other to form purposeful protein networks. In addition, owing to their structural asymmetry, protein molecules can use thermal energy from the environment to perform work as Brownian ratchets. It is essential to acknowledge that all the activities of a protein reside in its structure, which is built in by evolution. In terms of Shannon's communication theory (Shannon, 1948), the exploratory behaviour of a protein molecule—its constant change between sub-states—is a manifestation of its information entropy. The appropriate ligand triggers pre-programmed responses; the whole process is nothing more than a one-bit information transaction.
The principles of cognition at the basic molecular level seem to apply to cognition at all other levels. Cellular cognition is based on the operation of a set of molecular sensors as modules. Cellular cognitive devices constitute cognition at the level of the individual organism. There are numerous lower-level entities within higher-level individuals (Gould, 2002). Life appears as a Russian doll; it comprises several loosely bound modules and exhibits multi-layered nestedness. At higher levels, sentience means sensitivity and excitability, but still largely explores pre-programmed alternatives.
During bacterial chemotaxis, movements are modulated by ligand binding, which results in a biased random walk towards a higher or lower ligand concentration. The immune system is an example of sentience at a higher organismal level. Stochastic gene expression, which is not random in the common statistical sense but is circumscribed by other constraints, might be another example. So is synaptogenesis, in which many possible neural connections are displayed, but some—those ‘approved' by the input from the environment—are retained. Still, at these higher levels, the same principle applies as for a single protein: the signal merely triggers a response but does not contain information about what the response should be (Kirschner & Gerhart, 2005). Therefore, the use of terms from the theory of communication, including ‘channel capacity' or ‘information processing', and analogies from linguistics, might be misleading at all levels, with the exception of genuine human communication systems.
According to Feinberg (2001), “the brain is arranged in the same nested hierarchy as all biological systems. From this structure emerges the unified self.” As studies on the visual system in mammals have shown, specific neurons might show spontaneous patterns of activity that resemble representations of visual stimuli; such intrinsic cortical states might embody the brain's ‘hypotheses' about the state of the external world, which are continuously updated by visual data (Kenet et al, 2003). Conditioning is based on exploring various pre-programmed behaviours and stabilizing one of them. Scaruffi (2000) went so far as to state that emotions might be produced randomly and that the environment ‘selects' which one survives.
At all levels, cognition is a dual process of sensation and action. In humans, acting might become fully internalized in the form of thoughts; thinking, similar to dreaming, might be conceived as abstract motor activities. Konrad Lorenz (1943) described this as “handling in the imaginary space” (“das Hantieren im Vorstellungsraum”). In general, all living entities are semantically closed (Rosen, 1991); only humans, thanks to artefaction and the evolution of culture, may be an exception, but much less so than one might assume. This is reminiscent of Plato, for whom all knowledge was just a ‘recollection' of the soul, and of Johann W. Goethe, who remarked that “we see only what we know.”
There is much less information processing than is assumed by the ‘life as information' or ‘life as computation' metaphors that have dominated biology for the past 50 years. All levels of complexity—from protein molecules, cells, tissues and individuals, to social institutions and culture—represent embodied knowledge that has accumulated over time and has been retained by natural selection. Triggering of pre-determined responses and, indeed, selecting from them, seems to be a more appropriate description than information processing.
All levels of complexity…represent embodied knowledge that has accumulated over time and has been retained by natural selection
Chemistry, more than any other science, abounds in emergences. When molecules of hydrogen and oxygen react to produce molecules of water, a substance with new properties emerges. Because life is a chemical system, emergent phenomena at various levels of hierarchy are as natural, inevitable and unequivocal, but also as unpredictable, as the emergence of water from hydrogen and oxygen. The brain itself is a chemical system: not a computer with hardware and software, but rather ‘wetware' (Kosslyn & Koenig, 1995). Perceptional and emotional qualia, and even consciousness and self-consciousness, lose much of their mystery if we think of them as emergences in complex chemical systems, in which myriads of teleonomic chemical interactions—molecular cognitions—are occurring all the time.
Cognitive biology is more a reinterpretation of the existing data than a research programme offering new experimental approaches to age-old problems. One can ask, as did Tom Wolfe (2000), “Why wrestle with Kant's God, freedom, and immortality when it is only a matter of time before neuroscience, probably through brain imaging, reveals the actual physical mechanism that fabricates these mental constructs, these illusions?” The answer is at least threefold.
Cognitive biology is more a reinterpretation of the existing data than a research programme offering new experimental approaches to age-old problems
First, our psychological constitution does not allow us to wait until science can provide all the answers to our vital questions; to achieve peace of mind, humankind has always strived for unified all-embracing understanding. In our time, the rapid progress of science allows us to upgrade quickly our interpretations, however transitory, to fit the state of current knowledge.
Second, the insights provided by cognitive biology might help us to cope with constraints imposed on our understanding of the human brain, which is also known as Kuhlenbeck's paradox (Gerlach, 1988). Our world of consciousness is a phenomenon of the brain, but our brain is also a phenomenon of the brain: a closure reminiscent of Kurt Gödel's central theorem, “This statement cannot be proven.” Similar to Gödel's explanation of this paradox, we would need some divine ‘meta brain' to fully comprehend the human brain. However, the analysis of cognition at lower levels of complexity might allow us to extrapolate this knowledge towards human cognition and, in this way, circumvent Kuhlenbeck's barrier.
The third answer brings us back to where this essay started. Human suffering might have been the driving force behind science, which appears to be a history of efforts to reduce human suffering. Why not make this mission explicit and central to science? Then, of course, a proper understanding of human cognition, with emotion as its main and inseparable constituent, is a prerequisite for achieving the goal. The impact of this understanding could be far-reaching. As the poet André Breton expressed, a mistake in the explication of man causes an error in the explication of the universe. The ‘theory of everything' might turn out not to be the final theory of the fundamental elements of the world, but rather the theory of the mind and its relationship to the universe.

References
- Bazarov IP (1983) Termodinamika. Moscow: Vysshaya Shkola [Google Scholar]
- Damasio AR (1994) Descartes Error: Emotion, Reason, and the Human Brain. New York, NY, USA: Putnam [Google Scholar]
- Feinberg TE (2001) Altered Egos. Oxford, UK: Oxford University Press [Google Scholar]
- Frauenfelder H, McMahon BH, Fenimore PW (2003) Myoglobin: the hydrogen atom of biology and a paradigm of complexity. Proc Natl Acad Sci USA 100: 8615–8617 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerlach J (1988) Gödel-theorem and Kuhlenbeck-paradox. Naturwissenschaften 75: 393–398 [Google Scholar]
- Gibson W, Sterling B (1991) The Difference Engine. New York, NY, USA: Bantam Books [Google Scholar]
- Goodwin BC (1978) A cognitive view of biological process. J Soc Biol Struct 1: 117–125 [Google Scholar]
- Gould SJ (2002) The Structure of Evolutionary Theory. Cambridge, MA, USA: Harvard University Press [Google Scholar]
- Jencks WP (1975) Binding energy, specificity, and enzymic catalysis: the Circe effect. Adv Enzymol Relat Areas Mol Biol 43: 219–410 [DOI] [PubMed] [Google Scholar]
- Kenet T, Bibitchkov D, Tsodyks M, Grinvald A, Arieli A (2003) Spontaneously emerging cortical representations of visual attributes. Nature 425: 954–956 [DOI] [PubMed] [Google Scholar]
- Kirschner MW, Gerhart JC (2005) The Plausibility of Life: Great Leaps of Evolution. New Haven, CT, USA: Yale University Press [Google Scholar]
- Kornberg A (1989) For the Love of Enzymes. Cambridge, MA, USA: Harvard University Press [Google Scholar]
- Kosslyn SM, Koenig O (1995) Wet Mind: The New Cognitive Neuroscience. New York, NY, USA: Free Press [Google Scholar]
- Kovác L (2000) Fundamental principles of cognitive biology. Evol Cogn 6: 51–69 [Google Scholar]
- Lewontin RC (1992) The dream of the human genome. New York Rev Books 39: 31–40 [PubMed] [Google Scholar]
- Lorenz K (1943) Die angeborenen Formen möglicher Erfahrung. Z Tierpsychol 5: 235–409 [Google Scholar]
- Miller GA (2003) The cognitive revolution: a historical perspective. Trends Cogn Sci 7: 141–144 [DOI] [PubMed] [Google Scholar]
- Mitchell P (1961) Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 191: 144–148 [DOI] [PubMed] [Google Scholar]
- Monod J (1970) Le Hasard et la Nécessité. Paris, France: Éditions du Seuil [Google Scholar]
- Popper KR (1979) Objective Knowledge: An Evolutionary Approach. Oxford, UK: Clarendon [Google Scholar]
- Radnitzky G, Bartley WW (1987) Evolutionary Epistemology, Rationality, and the Sociology of Knowledge. La Salle, IL, USA: Open Court [Google Scholar]
- Rose SP (2005) Human agency in the neurocentric age. EMBO Rep 6: 1001–1005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosen R (1991) Life Itself. New York, NY, USA: Columbia University Press [Google Scholar]
- Scaruffi P (2000) The nature of emotions. www.thymos.com/science/emotion.html
- Shannon CE (1948) A mathematical theory of communication. Bell System Technical J 27: 379–423; 623–656 [Google Scholar]
- Wilson TD (2002) Strangers to Ourselves: Discovering the Adaptive Unconscious. Cambridge, MA, USA: Harvard University Press [Google Scholar]
- Wolfe T (2000) Hooking Up. New York, NY, USA: Farrar, Strous & Giroux [Google Scholar]