

Abstract
Salmonella enterica serovar Typhimurium (S. Typhimurium) elicits the starvation-stress response (SSR) due to starvation for an essential nutrient, e.g. a carbon/energy source (C-source). As part of the SSR, the alternative sigma factor σE is activated and induced. The authors suspect that this activation is, in part, triggered by changes in the S. Typhimurium cell envelope occurring during the adaptation from growth to carbon/energy starvation (C-starvation), and resulting in an increased need for σE-regulated factors involved in the proper folding and assembly of newly synthesized proteins destined for this extracytoplasmic compartment. This led to the hypothesis that a σE activation signal might arise during C-source shifts that cause the induction of proteins localized to the extracytoplasmic compartment, i.e. the outer membrane or periplasm, of the cell. To test this hypothesis, cultures were grown in minimal medium containing enough glucose to reach mid-exponential-phase, plus a non-limiting amount of a secondary ‘less-preferred’ but utilizable carbon/energy source. The σE activity was then monitored using plasmids carrying rpoEP1– and rpoEP2–lacZ transcriptional fusions, which exhibit σE-independent and -dependent lacZ expression, respectively. The secondary C-sources maltose, succinate and citrate, which have extracytoplasmic components involved in their utilization (e.g. LamB), resulted in a discernible diauxic lag period and a sustained increase in σE activity. Growth transition from glucose to other utilizable phosphotransferase (PTS) and non-PTS C-sources, such as trehalose, mannose, mannitol, fructose, glycerol, d-galactose or l-arabinose, did not cause a discernible diauxic lag period or a sustained increase in σE activity. Interestingly, a shift from glucose to melibiose, which does not use an extracytoplasmic-localized protein for uptake, did cause an observable diauxic lag period but did not result in a sustained increase in σE activity. In addition, overexpression of LamB from an arabinose-inducible promoter leads to a significant increase in σE activity in the absence of a glucose to maltose shift or C-starvation. Furthermore, a ΔlamB : : Ω-Kmr mutant, lacking the LamB maltoporin, exhibited an approximately twofold reduction in the sustained σE activity observed during a glucose to maltose shift, again supporting the hypothesis. Interestingly, the LamB protein lacks the typical Y-X-F terminal tripeptide of the OmpC-like peptides that activate DegS protease activity leading to σE activation. It does, however, possess a terminal pentapeptide (Q-M-E-I-W-W) that may function as a ligand for a putative class II PDZ-binding site. The authors therefore propose that the σE regulon of S. Typhimurium not only is induced in response to deleterious environmental conditions, but also plays a role in the adaptation of cells to new growth conditions that necessitate changes in the extracytoplasmic compartment of the cell, which may involve alternative signal recognition and activation pathways that are independent of DegS.
Abbreviations: C-source, carbon/energy source; C-starvation, carbon/energy starvation; IM, inner membrane; MS hiC, MOPS-buffered salts medium non-limiting in glucose; MS loC, MOPS-buffered salts medium limiting for glucose; OM, outer membrane; OMP, outer-membrane protein; PEP, phosphoenolpyruvate; PP, periplasmic; PTS, phosphotransferase; SSR, starvation-stress response
INTRODUCTION
Salmonella enterica serovar Typhimurium (Salmonella Typhimurium) is an aetiological agent of water/food-borne gastrointestinal-based disease in both animals and humans (Spector, 1998). An important feature of S. Typhimurium epidemiology and pathogenesis in both warm- and cold-blooded hosts is its ability to sense, adapt to and survive in a number of environments that expose the bacteria to stresses, such as nutrient starvation, extremes in osmolarity, temperatures or pH, which can threaten viability and/or limit growth (Spector, 1998; Humphreys et al., 1999, 2003). Thus, bacterial responses to these stresses can affect the spread and pathogenesis of this important pathogen.
One of the most common stresses encountered by S. Typhimurium, and many other bacteria, outside the confines of the laboratory is starvation for a carbon/energy source (C-source). S. Typhimurium, as well as other non-spore-forming bacteria, responds to carbon/energy starvation (C-starvation) with global alterations in gene expression, morphology and physiology. This response is known as the starvation-stress response (SSR) (Spector & Cubitt, 1992; Seymour et al., 1996; Spector, 1998; Spector et al., 1999b). The SSR serves two functions: (1) to combat the long-term effects of C-starvation, and (2) to generate cross-resistance to other environmental stresses. The SSR includes aspects of: (a) avoidance, involving the production of new or higher-affinity C-source-utilization systems in an attempt to scavenge any utilizable C-sources from the environment; and (b) starvation/stress survival if an alternative C-source remains unavailable, ultimately generating a more efficient and hardy cell (Spector, 1998).
The SSR genes identified thus far are under the control of one or more of three sigma factors, σ70, σS and σE (O’Neal et al., 1994; Spector et al., 1999a; Kenyon et al., 2002). The alternative sigma factor σS is encoded by the rpoS gene (Tanaka et al., 1993). During the SSR, σS is induced early in the response to C-starvation in both Salmonella and Escherichia coli (W. J. Kenyon & M. P. Spector, unpublished data; Fang et al., 1992; Tanaka et al., 1993), and during the shift from glucose to lactose utilization in E. coli (Fischer et al., 1998). Glucose is the carbon source of choice for Salmonella and many other bacteria. While Salmonella is growing on glucose, the uptake and utilization of other carbon sources is inhibited, in part as a result of catabolite repression and/or inducer exclusion (Notley & Ferenci, 1995; Moat et al., 2002). Glucose exhaustion results in a shift to utilize alternative C-sources present in the growth medium. In E. coli, a shift from glucose to lactose utilization causes growth to transiently halt, producing what is called a diauxic lag period. During diauxie, functions needed for lactose utilization are induced, allowing the cell to resume growth. In addition, σS and σS-dependent genes are induced during this diauxic lag period (Fischer et al., 1998).
The alternative sigma factor σE (encoded by the rpoE gene) is activated by certain stresses to the cell envelope that lead to the accumulation of denatured/unfolded proteins (Mecsas et al., 1993, 1995; Missiakas et al., 1996; Jones et al., 1997; Alba et al., 2002). Previous studies indicate that, once activated, σE initiates the transcription of several genes with extracytoplasmic functions required for the cell envelope to combat the effects of the stress. In the absence of stress, σE is normally bound to the inner-membrane (IM) protein RseA, an anti-sigma factor or inhibitor of σE, preventing its interaction with RNA polymerase, and thus the transcription of σE-dependent genes (De Las Penas et al., 1997; Missiakas et al., 1997; Alba et al., 2002, Miticka et al., 2003). When a specific stress occurs to the cell envelope, RseA is degraded by DegS and YaeL, σE is then released and is able to bind to core RNA polymerase (E). EσE then directs the transcription of its dependent genes, many of which possess extracytoplasmic functions. A number of σE-regulated genes identified in E. coli encode cell-envelope-associated proteins that act on misfolded proteins, leading to their degradation, proper refolding or correct insertion into the outer membrane (OM), e.g. periplasmic (PP) proteases, peptidyl-prolyl isomerases and molecular chaperones (Erickson & Gross, 1989; Danese & Silhavy, 1997; Dartigalongue et al., 2001; Alba et al., 2002).
Previous findings in our laboratory (Kenyon et al., 2002) showed that σE activity and protein levels increase during the initial 4–5 h of C-starvation, and reach peak levels at around 48–72 h of C-starvation. This suggests that C-starvation produces signals that lead to σE activation. Furthermore, rpoE mutants are defective in long-term C-starvation and C-starvation-inducible cross-resistance (Kenyon et al., 2002), indicating that σE is an additional key regulator of the SSR. What is more, σE is critical for the virulence of S. Typhimurium in the mouse virulence model (Humphreys et al., 1999).
In an effort to discern the role of the σE regulon in the SSR, we examined its activity in the avoidance component of the SSR by measuring σE-dependent transcription during shifts from glucose to less-preferred but utilizable C-sources. Here we report that shifts to some but not all secondary C-sources tested resulted in a sustained increase in σE activity. The common feature of C-sources resulting in increased σE activity appears to be the induction of an OM-associated and/or PP-binding-protein component involved in its utilization. Based on our findings and apparent common features of these utilization systems, we hypothesized that one or more members of the σE regulon is needed for the adaptation of the cell envelope to growth on less-preferred but utilizable C-sources, specifically those C-sources with transport systems possessing extracytoplasmic solute receptors and/or OM proteins. Therefore, σE-regulated functions appear to play a role not only in the responses to certain environmental stresses, but also in the adaptation of Salmonella to new environmental conditions not typically associated with cell envelope stress.
METHODS
Bacterial strains, plasmids and oligonucleotide primers
The bacterial strains, plasmids and oligonucleotide primers (Invitrogen) used are listed in Table 1. Transductions were carried out using the high-transducing derivative of bacteriophage P22, P22 HT int-105 (Chan et al., 1972). All strains were determined to be non-lysogens for P22 by demonstrating sensitivity to the H5 derivative of bacteriophage P22 (Maloy, 1990).
Table 1.
Bacterial strains, plasmids and primers
| Strain, plasmid or primer | Relevant characteristic* | Source/reference |
|---|---|---|
| Strains | ||
| SL1344 | hisG46 (mouse- virulent S. Typhimurium) | Hoseith & Stocker (1981) |
| ST55 | SL1344 lamB : : MudJ (lac Kmr) | This study |
| ST213 | SL1344 rpoE : : Ω-Kmr | Kenyon et al. (2002) |
| ST276 | Wild-type S. Typhimurium NCTC12023/pKD46 (Apr) | Mark Roberts† |
| SMS758 | SL1344/pRS1274 (Apr) | Kenyon et al. (2002) |
| SMS759 | SL1344/pTF-p1 (Apr) | Kenyon et al. (2002) |
| SMS760 | SL1344/pTF-p2 (Apr) | Kenyon et al. (2002) |
| SMS865 | ST276 ΔlamB : : Ω-Kmr | This study |
| SMS871 | SL1344 ΔlamB : : Ω-Kmr | This study |
| SMS872 | SMS871/pRS1274 (Apr) | This study |
| SMS873 | SMS871/pTF-p1 (Apr) | This study |
| SMS874 | SMS871/pTF-p2 (Apr) | This study |
| SMS907 | SMS758/pKS34 (Cmr) | This study |
| SMS908 | SMS760/pKS34 (Cmr) | This study |
| SMS909 | ST213/pTF-p2+pKS34 (Apr Cmr) | This study |
| Plasmids | ||
| pRS1274 | Vector for making transcriptional lacZ fusions (Apr) | Simons et al. (1987) |
| pTF-p1 | rpoEp1 : : lacZ in pRS1274 (rpoE-independent promoter; Apr) | Testerman et al. (2002) |
| pTF-p2 | rpoEp2 : : lacZ in pRS1274 (rpoE-dependent promoter; Apr) | Testerman et al. (2002) |
| pKD46 | Carries λ-Red γ, β and Exo genes under the control of an arabinose-inducible araBAD promoter, ParaBAD (Apr) | Datsenko & Wanner (2000) |
| pKD4 | Carries FRT-Kmr-FRT Ω-cassette (Apr) | Datsenko & Wanner (2000) |
| pBAD33 | Vector for cloning genes under the control of ParaBAD (Cmr) | Guzman et al. (1995) |
| pKS33 | 1·4 kb fragment carrying lamB in pGEM-TEasy (Apr) | This study |
| pKS34 | SacI–SphI fragment from pKS33 with lamB+ in pBAD33 (Cmr) | This study |
| Primers | Sequence (5′ to 3′): | |
| PR18 | CCATCAGATCCCGAATAATCCAATGTCCTCCC | |
| PR29 | GTTTTCCCAGTCACGAC | |
| PR48 | AGGTGGCAGCGTTTAAAGAA | |
| PR49 | GCCCTCGTTACGTCAGAT | |
| PR122 | AGGTGGCAGCGTTTAAAGAAAAGCAATGATCTCAGGAGATGTGTAGGCTGGAGCTGCTTC | |
| PR123 | GCCCCTCGTTACGTCAGATGACCATCGTATTACCACCAGACATATGAATATCCTCCTTA | |
Kmr, Kanamycin resistance; Apr, ampicillin resistance; Cmr, chloramphenicol resistance.
Dr Mark Roberts, School of Veterinary Medicine, University of Glasgow, Glasgow, UK.
Media and supplements
Cultures of the desired strains were grown in a modified MOPS-buffered salts (MS) minimal medium (Neidhardt et al., 1974) containing either 0·4 % (w/v) glucose (MS hiC) or 0·03 % (w/v) glucose (MS loC), and are described in detail elsewhere (Spector & Cubitt, 1992; O’Neal et al., 1994). Histidine was added to minimal medium to a final concentration of 20 μM. The appropriate antibiotics were added as needed: ampicillin (Ap) (30 μg ml−1), kanamycin (Km) (50 μg ml−1) and chloramphenicol (Cm) (50 μg ml−1). The rich medium used was Luria–Bertani (LB) broth and agar (Difco).
Identification of a lamB : : MudJ (lac Kmr) -carrying strain from a library of C-starvation-inducible MudJ : : lac fusion (insertion) strains
A lamB : : MudJ (lac Kmr) fusion-carrying strain was identified from our library of strains carrying MudJ (lac Kmr) insertions demonstrating C-starvation induction (Spector et al., 1988, 1999a), using single-primer PCR, DNA sequence analysis and blastp search analysis as previously described (Spector et al., 1999a, b). The insertion of the MudJ : : lac in lamB was confirmed by the presence of the correct size PCR product using chromosomal DNA from ST55 as a template, a forward primer specific for the 3′-end of MudJ (PR18) and a reverse primer specific for the 3′-end of lamB (PR49).
Growth and σE activation during C-source shifts
For growth and σE-activation analysis during glucose to alternative C-source shifts, overnight cultures of the desired strains were diluted 1 : 100 into fresh MS loC medium supplemented with one of the following: maltose, succinate, citrate, melibiose, d-trehalose, d-mannose, d-mannitol, d-fructose, glycerol, l-arabinose or d-galactose, all at a final concentration of 0·4 % (w/v), or no additional C-source (all C-sources were purchased from Sigma). The appropriate antibiotics were added as needed. The concentration of glucose in MS loC is the concentration used to produce C-starved cells (Spector & Cubitt, 1992; O’Neal et al., 1994; Kenyon et al., 2002) and provides sufficient glucose to support growth for 3–4 generations before being exhausted. Therefore, growth can only continue if an alternative utilizable C-source is available in the growth medium. Growth was monitored spectrophotometrically by determining OD600 at pre-determined time intervals. Activation of σE was determined by measuring β-galactosidase activity as a reporter of promoter activity from plasmid pTF-P2; in addition, β-galactosidase activity driven by a rpoE-independent promoter (pTF-P1) was measured as a control (Kenyon et al., 2002). β-Galactosidase activity was assayed by the method of Miller (1992) and is expressed in Miller units. The results presented represent the mean±sem from at least three separate experiments.
Generation of the lamB deletion mutant
The λ-Red mutagenesis system provides a more efficient way of disrupting and deleting chromosomal genes, allowing the characterization of a gene’s function (Datsenko & Wanner, 2000), using the precise insertion of a selected antibiotic-resistance cassette into the gene of interest accompanied by a selected deletion. Using a slightly modified protocol and S. Typhimurium strain ST276 (carrying pKD46, a plasmid with the λ-Red recombination enzymes under the control of the l-arabinose-inducible ParaBAD promoter; Table 1), we constructed a defined deletion of the lamB gene from the S. Typhimurium chromosome, replacing it with an Ω-Kmr cassette.
Following the protocol of Datsenko & Wanner (2000), a PCR product was generated containing the Kmr cassette from pKD4 and sequence from the flanking region of the lamB gene of S. Typhimurium. PCR primers PR122 and PR123 (Table 1) were designed to contain a 40 bp sequence homologous to lamB DNA, and a sequence corresponding to the P1 and P2 sites on pKD4. PCR reactions using these primers, PCR SuperMix High Fidelity (Invitrogen) and pKD4 as a template were run in an Ericomp Thermal Cycler. The size of the PCR product (approximately 1·6 kb) was confirmed by agarose gel electrophoresis, and the DNA was cleaned using the Promega Wizard PCR Prep Kit. The DNA was then digested with DpnI (Promega) for 3 h at 37 °C, and cleaned using the Promega Wizard DNA Clean-up Kit.
Electrocompetent cells of strain ST276 were prepared by growing an overnight culture of ST276 in 1 ml LB-Ap with shaking at 37 °C. The following morning, the overnight culture was diluted 1 : 100 into 25 ml of LB containing 1 mM l-arabinose, and grown to an OD600 of between 0·6 and 0·8. Freshly grown cells were pelleted, washed twice with 10 ml ice-cold sterile distilled water, and once with 10 ml ice-cold sterile 10 % (v/v) glycerol.
DpnI-digested DNA was then electroporated into the freshly prepared electrocompetent cells using an E. coli Pulsar electroporation unit and the protocol provided by the manufacturer (Bio-Rad). The cells were plated onto LB-Km agar and incubated overnight at 37 °C. Km-resistant colonies were then streaked for isolation onto LB-Km agar, and DNA was isolated from LB-Km overnight cultures using the Promega Genomic DNA Purification Kit. The presence of the desired mutation was confirmed by PCR analysis using primers PR48 and PR49 (Table 1) specific for the lamB sequence outside of the deletion site.
Construction of plasmid pKS34 carrying lamB under the control of an arabinose-inducible promoter, ParaBAD : : lamB
Chromosomal DNA isolated from S. Typhimurium SL1344 (Wizard Genomic DNA Purification Kit; Promega) served as template for high-fidelity PCR amplification (Platinum PCR SuperMix High Fidelity; Invitrogen) of the lamB gene using the forward primer PR48 and the reverse primer PR49 (Table 1). The 1·4 kb PCR product was purified and ligated into pGEM-T Easy (Promega), and E. coli JM109 competent cells (Promega) were transformed with this, following the manufacturer’s protocols. Plasmid minipreps (Wizard Plus Minipreps; Promega) from the ampicillin-resistant transformants were screened by PCR using primers PR48 and PR29 (Table 1) to determine that the insert was oriented in the same direction as the lacZ gene in order to facilitate cloning into pBAD33. The subsequent plasmid was designated pKS33.
Digestion of pKS33 with SacI and SphI yielded a 1·5 kb fragment carrying the lamB gene. This fragment was isolated using agarose gel electrophoresis, extracted (Freeze ′N Squeeze columns; Bio-Rad) and cleaned (Wizard DNA Clean-Up System; Promega). It was then ligated with SacI/SphI-digested pBAD33 (a generous gift from Dr Jon Beckwith, Harvard University, Boston, MA, USA), and E. coli JM109 cells (Promega) were transformed with it and plated onto LB-Cm plates. Plasmid minipreps (Wizard Plus Minipreps; Promega) from transformant colonies were screened for the lamB insert by digestion with SacI and SphI, and agarose gel electrophoresis. The resultant plasmid was designated pKS34.
Overexpression of LamB and σE activation
To measure the effect of LamB overproduction on σE activation as measured from a σE-dependent promoter in pTF-p2 we transformed SMS760 (SL1344/pTF-p2) with pKS34 to produce strain SMS908 (Table 1). The desired strains were grown overnight in LB plus the appropriate antibiotic. Overnight cultures were then diluted 1 : 100 into fresh LB broth with the appropriate antibiotic(s), and grown to mid-exponential phase before being divided into six separate cultures to which 0, 25, 50, 100, 200 or 400 μM l-arabinose was added. Cultures were then incubated for an additional 1 h at 37 °C with aeration, at which point β-galactosidase activity was assayed using the method of Miller (1992).
RESULTS AND DISCCUSION
Shifts from glucose to maltose, citrate or succinate result in a sustained increase in σE activity
In S. Typhimurium, a number of C-source utilization enzymes and systems are induced during the transition from growth to C-starvation in an effort to scavenge any and all available C-sources, and avoid entry into the physiologically more drastic ‘survival’ mode (Spector, 1998). Many of the utilization systems involve the induction of OM, PP and/or IM components in addition to cytoplasmic proteins. An example of an OM component of an alternative C-source utilization system is LamB, which functions as a porin-like protein involved in the uptake of maltose and maltodextrins (Schulein & Benz, 1990; Hall et al., 1997). Induction of a lamB : : lacZ fusion during C-starvation, as well as during shifts from glucose to maltose, showed that lamB is induced approximately 17-fold by C-starvation (1·4±0·4 Miller units in exponential-phase cells compared to 23·9±2·4 Miller units in 5 h C-starved cells), and approximately 46-fold during shifts from glucose to maltose (1·5±0·7 Miller units in cells prior to the shift to maltose compared to 70·1±1·7 Miller units 4 h after the shift to maltose). These results, along with previous findings that σE is a key regulator of the SSR and is activated within the first few hours of C-starvation (Kenyon et al., 2002), led us to examine the role of σE early in the SSR during the ‘avoidance’ phase. For this, we assessed σE activity during the transition from growth on glucose to less-preferred but utilizable C-sources in S. Typhimurium.
Fig. 1(a) illustrates the typical increase in σE activity (as determined by increased β-galactosidase expression driven by an σE-dependent promoter) observed during C-starvation (Kenyon et al., 2002), and is included for comparison purposes. Shifts from glucose to maltose, citrate or succinate all produced discernible, yet transient, diauxic lag periods, and more importantly sustained increases in σE activity (Fig. 1b–d) comparable to C-starved cells (Fig. 1a) over the same time period. In contrast, shifts from glucose to maltose, citrate, succinate or any other alternative C-source tested, as well as C-starvation, produced relatively constant β-galactosidase activity [2478±69 (sem) Miller units] over the same time period from the σE-independent (σ70-dependent) promoter in pTF-p1.
Fig. 1.

Growth and σE activation during shifts from glucose to (a) no secondary C-source (early C-starvation), (b) maltose, (c) citrate and (d) succinate. Graphs represent growth (OD600) (•) and σE activation (▪) as reported by measuring lacZ expression (β-galactosidase activity) driven by a σE-dependent promoter. β-Galactosidase activity is monitored using the method of Miller (1992) and is given in Miller units. Data for β-galactosidase activities represent means±sem (n≥3).
A common feature of the utilization systems for these C-sources appears to be the presence of an OM- and/or PP-protein component in addition to IM transporters. In S. Typhimurium, this includes the LamB OM-maltoporin complex and the MalE extracytoplasmic solute receptor (binding protein) for maltose utilization (Schulein & Benz, 1990; Hall et al., 1997; Lawhon et al., 2003). Utilization of citrate (a C6-tricarboxylate) involves an extracytoplasmic (PP) solute-receptor (binding) protein without a specific OM component. TctC is the tricarboxylate-binding-receptor component of the TctABC tripartite-tricarboxylate transporter (TTT) of S. Typhimurium involved in citrate utilization (Winnen et al., 2003).
The scenario for succinate utilization in S. Typhimurium is more complicated. The homologues of the dctPQM genes encoding the aerobic dicarboxylate-transporter system DctPQM in Rhodobacter capsulatus (Rabus et al., 1999; Janausch et al., 2002) are apparently absent from the S. Typhimurium genome (McClelland et al., 2001). However, there are at least three hypothetical genes encoding putative extracytoplasmic solute receptors for C4-dicarboxylates that may play a role in succinate utilization (STM3169, GenBank accession number AE008845,17756–18746; yiaO, accession number AE008870, 9637–10623; STM4054, accession number AE008889, complement 10233–11216). Of these three, YiaO along with YiaM/YiaN, and STM4054 along with STM4052/STM4053 may constitute tripartite ATP-independent periplasmic- (TRAP) transporter systems involved in succinate utilization (Rabus et al., 1999; Janausch et al., 2002). In addition, S. Typhimurium possesses the genes for two other C4-dicarboxylate-transport systems: (1) the anaerobic Dcu dicarboxylate-transporter system, which appears to be involved in fumarate respiration rather than C4-dicarboxylate utilization and as part of this function actually results in succinate efflux from the cytoplasm (Janausch et al., 2002), and (2) a homologue of the DctA H+/Na+: C4-dicarboxylate/dicarboxylic amino acid symporter, which was found to be a major C4-dicarboxylate transporter during aerobic growth in E. coli. Mutants of dctA in E. coli grow very poorly or not at all on succinate and other C4-dicarboxylates (Davies et al., 1999). However, its cognate extracytoplasmic solute receptor (i.e. succinate-binding protein) has not been definitively identified. In E. coli, the cbt gene was proposed as the putative C4-dicarboxylate-binding protein possibly as part of an OM porin (Lo, 1977; Lo & Bewick, 1981). Whether this complex is expressed under the conditions tested in this study has not been demonstrated, but recent evidence suggests that cbt may be an allele of the ferric-enterobactin-transport component gene fepA, which is iron regulated (Braun & Braun, 2002) in both E. coli and Salmonella. Thus, the involvement of PP and/or OM component(s) in succinate utilization in S. Typhimurium, although likely, is ambiguous and clearly complicated.
Shifts from glucose to the PEP : PTS-dependent sugars trehalose, mannose, mannitol or fructose do not result in a sustained increase in σE activity
Many sugars are translocated into the cell by the phosphoenolpyruvate (PEP) : sugar phosphotransferase system (PTS) (Postma et al., 1993). The utilization of secondary PTS sugars, and some other non-PTS sugars, is catabolite repressed and/or subject to inducer exclusion during growth on glucose. This is in part a result of low intracellular levels of the signal nucleotide cyclic AMP (cAMP) caused by greatly reduced adenylate cyclase activity, resulting from greatly reduced levels of the phosphorylated Enzyme IIA component of the glucose-specific PTS (P~EnzIIAgluc). The exhaustion of glucose leads to accumulation of P~Enz IIAgluc and the activation of adenylate cyclase. The resulting increased cAMP levels, and in some cases the presence of the C-source (or a derivative), induces the expression of the utilization enzymes for the secondary C-source. For some of these C-sources, this involves the production of specific Enzyme IIA and IIB components for its PTS. The IIB component for these PTS sugars is an IM protein, while their IIA component is associated with the cytoplasmic side of the IM; however, there are no PP or OM components involved in the PTS. We therefore wanted to determine if shifts from glucose to known PTS sugars might also generate signals leading to sustained σE activation.
In contrast to maltose, citrate and succinate, a shift from glucose to the PTS-dependent sugars trehalose (Fig. 2a), fructose (Fig. 2b), mannose or mannitol (data not shown) did not lead to a discernible diauxic lag period or sustained σE activation. Results for mannose and mannitol were very similar to that of fructose. These results indicate that a shift from glucose to a secondary C-source, in general, is insufficient to cause sustained σE activation. It also demonstrated more specifically that shifts from glucose to PTS sugars do not generate signals leading to sustained σE activation, thus supporting our hypothesis, given that their utilization does not involve PP and/or OM components.
Fig. 2.

Growth and σE activation during shifts from glucose to (a) trehalose and (b) fructose. Graphs represent growth (OD600) (•) and σE activation (▪) as reported by measuring lacZ expression (β-galactosidase activity) driven by an σE-dependent promoter. Promoter activity is reported by measuring lacZ expression (β-galactosidase activity) driven by the σE-dependent rpoE promoter. β-Galactosidase activity is monitored using the method of Miller (1992) and is given in Miller units. OD600 values above 2·0 are not plotted. Data for β-galactosidase activities represent means±sem (n≥3).
Shifts from glucose to the non-PTS C-sources melibiose, l-arabinose, d-galactose or glycerol do not result in a sustained increase in σE activity
Some secondary C-sources do not require the PTS for their utilization, but instead rely on a specific IM-associated transporter, permease or channel to bring them into the cytoplasm for utilization; although they may exhibit catabolite repression and/or inducer exclusion (as described above). Moreover, in some cases, their expression in S. Typhimurium is uncertain. Examples of non-PTS C-sources that exhibit these characteristics include glycerol, l-arabinose, d-galactose and melibiose in S. Typhimurium (Postma, 1977; Muller et al., 1985; Benner-Luger & Boos, 1988; Horazdovsky & Hogg, 1989; Kehres & Hogg, 1992; Moat et al., 2002). Thus, we examined whether shifts from glucose to one of these C-sources would result in a sustained increase in σE activity.
Neither shifts from glucose to melibiose (Fig. 3a), l-arabinose (Fig. 3b), d-galactose, nor glycerol (data not shown) resulted in a sustained increase in σE activity; although there was a transient increase in some cases. The results for d-galactose and glycerol were very similar to the result for l-arabinose. Interestingly, the shift from glucose to melibiose produced an observable transitory diauxic lag period, but again this was not accompanied by a sustained elevation in σE activity (Fig. 3a). These latter findings suggest that it is not the occurrence of a diauxic lag period that results in signals leading to a sustained rise in σE activity.
Fig. 3.

Growth and σE activation during shifts from glucose to (a) melibiose and (b) l-arabinose. Graphs represent growth (OD600) (•) and σE activation (▪) as reported by measuring lacZ expression (β-galactosidase activity) driven by a σE-dependent promoter. β-Galactosidase activity is monitored using the method of Miller (1992) and is given in Miller units. OD600 values above 2·0 are not plotted. Data for β-galactosidase activities represent means±sem (n≥3).
In E. coli, there appears to be a high-affinity arabinose-transport system encoded by the araFGH operon, with AraF being an arabinose-binding protein and AraGH composing an IM-transport system (Horazdovsky & Hogg, 1989; Kehres & Hogg, 1992). However, the araFG genes are not present in the S. Typhimurium genome (McClelland et al., 2001; GenBank accession number AE006468 or NC_003197). This supports our hypothesis since the PP-binding protein for arabinose AraF would not be produced. Database analysis of the S. Typhimurium LT-2 genome sequence did not indicate known or putative AraF homologues.
In both E. coli and S. Typhimurium, d-galactose can be transported into the cell through the galactose-specific galactose permease GalP or the binding-protein-dependent methyl-galactoside Mgl system (Postma, 1977; Muller et al., 1985; Benner-Luger & Boos, 1988). The mglB-encoded protein is a PP-binding protein that can bind to galactose as well as other sugars, e.g. fucose. Although this might appear contradictory to our hypothesis, galactose only slightly induces the mgl genes in a galP+ strain background (Postma, 1977; Muller et al., 1985; Benner-Luger & Boos, 1988). Thus, the MglB protein is not induced significantly under our experimental conditions. So, the lack of a sustained increase in σE activity during a glucose to d-galactose shift can still be explained in the context of our hypothesis. The pertinent characteristics for all the secondary C-sources tested are summarized in Table 2.
Table 2.
Summary of the pertinent characteristics of the secondary C-sources
| C-source shifted to | PTS sugar* | PP-BP† | OMP‡ | Diauxic lag | Sustained σE activation |
|---|---|---|---|---|---|
| Maltose | − | + | + | + | + |
| Citrate | − | + | − | + | + |
| Succinate | − | + | − | + | + |
| Trehalose | + | − | − | − | − |
| Mannose | + | − | − | − | − |
| Mannitol | + | − | − | − | − |
| Fructose | + | − | − | − | − |
| Melibiose | − | − | − | + | − |
| Glycerol | − | − | − | − | − |
| l-Arabinose | − | NK | − | − | − |
| Galactose | − | NK | − | − | − |
PTS sugar, utilization of C-source requires the PEP-dependent : PTS system.
PP-BP, utilization system has a PP-binding protein component; nk, not known to be present in S. Typhimurium genome (accession no. AE006468 or NC_003197, Salmonella typhimurium LT-2 complete genome sequence) or not known to be expressed under the conditions being tested.
OMP, utilization system has an OMP component.
Inability to produce the LamB protein reduces the activation of σE during a shift from glucose to maltose
As stated previously, a common feature of maltose, citrate and succinate utilization appears to be the production of an OM and/or extracytoplasmic (PP) solute-receptor component(s) involved in their utilization, e.g. the LamB OM-porin-like protein complex (Schulein & Benz, 1990), or the extracytoplasmic solute receptors involved in C4-dicarboxylate and C6-tricarboxylate transport (Widenhorn et al., 1988; Rabus et al., 1999; Janausch et al., 2002; Winnen et al., 2003). Another possible factor involved in σE activation under these conditions might be a slower growth rate produced by the secondary C-source, e.g. shifts to citrate or succinate (Fig. 1b–c). To investigate this, we created a targeted deletion of the lamB gene to knock-out LamB production.
We targeted the lamB gene for several reasons. First, it is not essential for the utilization of maltose, yet it is induced during shifts from glucose to maltose (discussed above). Second, a lamB : : Ω-Km mutant strain (SMS871) exhibits virtually identical growth, compared to the wild-type parent strain, during glucose to maltose shifts (Fig. 4a). Furthermore, the situation for citrate or succinate utilization is problematic for this study. Mutants of tctC, lacking the extracytoplasmic tricarboxylate-binding-protein/receptor component, are defective in citrate utilization (Winnen et al., 2003), and thus may mimic conditions of C-starvation. What’s more, succinate utilization in Salmonella is not very well characterized. There are at least three putative C4-dicarboxylate-binding proteins in the S. Typhimurium genome (McClelland et al., 2001), and the presence of C4-dicarboxylate PP- and/or OM-utilization components in S. Typhimurium as well as E. coli still requires clarification. Deletion of one or more of the C4-dicarboxyate PP-binding proteins is likely to make the cells defective in succinate utilization similar to the scenario for citrate utilization.
Fig. 4.

Effect of a lamB knock-out mutation on σE activation during shifts from glucose to maltose utilization. (a) Growth (inset) and (b) σE activation, reported as fold-induction from a σE-dependent promoter, during shifts from glucose to maltose in the wild-type lamB+ strain (SMS760) (•) and ΔlamB mutant derivative (SMS874) (○). Time zero represents the initiation of the diauxic lag period. Fold-induction was calculated by dividing the value at each time point by the value at time zero. Promoter activity is reported by measuring lacZ expression (β-galactosidase activity) driven by the σE-dependent rpoE promoter. β-Galactosidase activity was determined using the method of Miller (1992). Data presented represent mean fold-inductions from at least three independent experiments. The sem for the data points is ≥24 % of the mean.
As would be predicted from our hypothesis, the fold-induction of the σE-dependent promoter is reduced in a lamB knock-out mutant to ~50 % of the fold-induction levels seen in the wild-type parent: a twofold-induction compared to a fourfold-induction, respectively (Fig. 4b). Residual activation of σE may be the result of the production of MalE, the extracytoplasmic maltose-binding protein/receptor required for maltose utilization, as well as other maltodextrin-utilization proteins that are also expressed under these conditions (Hall et al., 1997). A previous study in E. coli found that at lower levels of expression a folding-defective MalE protein is degraded by DegP or Protease III (Betton et al., 1998), supporting this idea. Unfortunately, knocking-out a required utilization component, such as MalE, would prevent growth on the C-source, and would again mimic C-starvation. But the fact that the loss of LamB production stifles the increase in σE activity normally seen during glucose to maltose shifts, yet the lamB mutant exhibits virtually indistinguishable growth rates under these conditions (Fig. 4a), supports the hypothesis that one or more σE-regulated functions are needed for efficient LamB incorporation into the OM during this adaptation process.
Overproduction of LamB protein in the absence of glucose starvation or a glucose to maltose shift increases activation of σE
To further demonstrate that up-regulated expression of LamB directly results in increased σE activity, we overproduced LamB from the arabinose-inducible araBAD promoter in pBAD33 (Guzman et al., 1995). As shown in Fig. 5, increased σE activity was observed when we artificially overproduced LamB from an arabinose-inducible promoter (SMS908) in the absence of glucose starvation or during a glucose to maltose shift (Fig. 5b). An increase in σE activity was not observed under these conditions in a strain lacking the pKS34 plasmid (SMS760), indicating that it is the overproduction of LamB and not the addition of arabinose itself that results in increased σE activity. Furthermore, the increased levels of β-galactosidase activity observed with rising concentrations of arabinose in SMS908 (Fig. 5b) required a functional σE protein, since in similar experiments with the rpoE : : Ω-Kmr mutant SMS909 low but constant β-galactosidase activities (approximately 10 Miller units), equivalent to a strain carrying the pRS1274 vector alone, were detected.
Fig. 5.
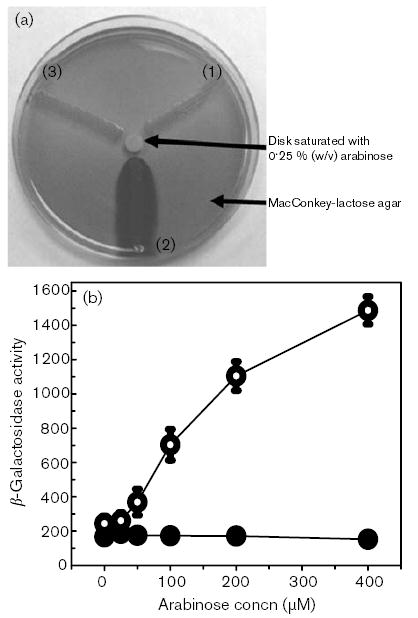
Effect of overproduction of LamB in LB medium. (a) Photograph of strains rpoE+(pKS34, pRS1274) (1), rpoE+(pKS34, pTF-p2) (2) and rpoE : : Ω-Kmr(pKS34, pTF-p2) (3). Strains were streaked onto MacConkey-lactose agar, and a disk saturated with 0·25 % (w/v) l-arabinose was placed in the centre of the plate. Production of β-galactosidase activity is indicated by the appearance of a red colour (seen here as dark grey) in the streak and the agar surrounding the streak. (b) β-Galactosidase activity from pTF-p2 alone (SMS760, •), and from pTF-p2 in response to increasing amounts of arabinose-induced expression of LamB from pKS34 (SMS908, ○). β-Galactosidase activity is monitored using the method of Miller (1992) and is given in Miller units. Data for β-galactosidase activities represent means±sem (n≥3).
These results demonstrate that the inappropriate overproduction of LamB can also generate signals that result in a sustained increase in σE activity. This is not that surprising since the artificial or inappropriate overproduction of several other outer-membrane proteins (OMPs), including OmpC or OMP-like C-terminal peptides, can up-regulate σE activity through increased DegS-mediated proteolysis (Ades et al., 1999; Walsh et al., 2003). An interesting difference in LamB is the absence of the Y-X-F [Tyr-X-Phe; where X can be several different amino acids such as Tyr (Y) or Gln (Q)] C-terminus that OmpC and several other OMPs possess. This is proposed to be the peptide signal/ligand that binds to the PDZ-domain of DegS, activating its protease activity (Walsh et al., 2003). Interestingly, the LamB protein (accession no. NP_463096) possesses a Q-M-E-I-W-W (Gln-Met-Glu-Ile-Trp-Trp) C-terminal pentapeptide that exhibits some similarity to the Φ-X-Φ [where Φ is a hydrophobic residue such as Val (V), Leu (L), Ile (I), Met (M) or Pro (P)] ligand for the proposed class II PDZ-binding site (Ponting, 1997; Songyang et al., 1997); although the closest match is the M-E-I sequence two residues distal to the C-terminus of the protein. This suggests that LamB generated-signals may differ, or work through different pathways from the typical C-terminal peptide signal or DegS-dependent pathways reported to result in σE activation. Intriguingly, the C-terminal pentapeptide of the tricarboxylate-binding receptor TctC (F-G-L-A-K, Phe-Glu-Leu-Ala-Lys; accession no. AAG31205) and the putative dicarboxylate-binding receptors STM3169 (Y-I-E-N-M, Tyr-Ile-Glu-Asn-Met; accession no. NP_462084), YiaO (V-D-E-V-Q, Val-Asp-Glu-Val-Gln; accession no. NP_462573) and STM4054 (Q-T-A-K-N, Gln-Thr-Ala-Lys-Asn; accession no. NP_462935) are also lacking the typical OMP-like C-terminal peptide sequence, again suggesting that alternative signals or DegS-independent pathways may be involved.
A question arises as to what σE-regulated functions might be involved in this adaptation process. We are currently searching for these functions. We do not believe them to be either DegP, FkpA or SurA since we have found that in S. Typhimurium they are not regulated by σE (or C-starvation) under the conditions tested here (W. J. Kenyon, A. Stevenson, S. Humphreys, M. Roberts & M. P. Spector, unpublished data). Thus, some alternative protease(s), chaperone(s), peptidyl-prolyl isomerase(s) or other function(s) as-yet-unidentified, would appear to be involved in the adaptation of the cell envelope to C-starvation conditions.
Studies in E. coli (Ades et al., 2003) identified trends in σE activity occurring before, during and after the inducing stress of high temperature. During the initiation of the stress response, σE activity is elevated. The activity remains elevated during the adaptation phase while the cell adapts to the stress. When the stress is removed, σE activity drops below that seen in normal conditions, and then slowly rises to pre-stress levels. A similar scenario can be postulated for shifts from glucose to maltose, for example, which even though not typically considered a stress do require the cells to adapt to the change in growth conditions. Interestingly, during C-starvation σE levels are seen to continue to rise over a 48–72 h period before levelling off (Kenyon et al., 2002).
Conclusions
We report here that shifts from glucose to maltose, citrate or succinate, but not trehalose, mannose, mannitol, fructose, glycerol, l-arabinose, galactose or melibiose, result in a sustained elevation of σE activity. The common feature of maltose, citrate and succinate utilization is the production of extracytoplasmic (PP) solute-receptors/binding proteins (e.g. TctC) and/or OM transport proteins or porins (e.g. LamB). The other secondary C-sources tested required either PEP : PTS-dependent-transport systems (e.g. trehalose, mannose, mannitol and fructose) or non-PTS IM-transport systems (e.g. glycerol, l-arabinose, galactose and melibiose). However, an extracytoplasmic component in their utilization systems is either absent, unknown or not expressed under the conditions tested in S. Typhimurium. The findings presented here support the hypothesis that σE-regulated functions are needed for the adaptation of cells to new environmental conditions that necessitate changes in the extracytoplasmic compartment of the cell, i.e. OM or periplasm. This implies that members of the σE regulon have functions beyond the repair or elimination of damaged extracytoplasmic proteins resulting from an environmental stress.
Acknowledgments
We thank Drs J. W. Foster, Z. Aliabadi, D. McGee and T. Testerman for helpful discussions and advice. Portions of this research were supported by Public Health Service-NIAID grant AI060759-01 and National Science Foundation (NSF) grant MCB-9985981 (to M. P. S.). E. J. was also a participant of a Summer Research Program funded through a grant from the NSF-Research Experiences for Undergraduates (REU) Program.
References
- Ades SE, Connelly LE, Alba BM, Gross CA. The Escherichia coli σE-dependent extracytoplasmic stress response is controlled by the regulated proteolysis of an anti-σ factor. Genes Dev. 1999;13:2449–2461. doi: 10.1101/gad.13.18.2449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ades SE, Grigorova IL, Gross CA. Regulation of the alternative sigma factor σE during initiation, adaptation, and shutoff of the extracytoplasmic heat shock response in Escherichia coli. J Bacteriol. 2003;185:2512–2519. doi: 10.1128/JB.185.8.2512-2519.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alba BM, Leeds JA, Onufryk C, Lu CZ, Gross CA. DegS and YaeL participate sequentially in the cleavage of RseA to activate the σE-dependent extracytoplasmic stress response. Genes Dev. 2002;16:2156–2168. doi: 10.1101/gad.1008902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benner-Luger D, Boos W. The mglB sequence of Salmonella typhimurium LT-2; promoter analysis by gene fusions and evidence for a divergently oriented gene coding for the mgl repressor. Mol Gen Genet. 1988;214:579–587. doi: 10.1007/BF00330498. [DOI] [PubMed] [Google Scholar]
- Betton JM, Sassoon N, Hofnung M, Laurent M. Degradation versus aggregation of misfolded maltose-binding protein in the periplasm of Escherichia coli. J Biol Chem. 1998;273:8897–8902. doi: 10.1074/jbc.273.15.8897. [DOI] [PubMed] [Google Scholar]
- Braun V, Braun M. Iron transport and signaling in Escherichia coli. FEBS Lett. 2002;529:78–85. doi: 10.1016/s0014-5793(02)03185-x. [DOI] [PubMed] [Google Scholar]
- Chan RK, Botstein D, Watanabe T, Ogata Y. Specialized transduction of tetracycline resistance by phage P22 in Salmonella typhimurium. II. Properties of a high-frequency-transducing lysate. Virology. 1972;50:883–898. doi: 10.1016/0042-6822(72)90442-4. [DOI] [PubMed] [Google Scholar]
- Danese PN, Silhavy TJ. The σE and the Cpx signal transduction systems control the synthesis of periplasmic protein-folding enzymes in Escherichia coli. Genes Dev. 1997;11:1183–1193. doi: 10.1101/gad.11.9.1183. [DOI] [PubMed] [Google Scholar]
- Dartigalongue C, Missiakas D, Raina S. Characterization of the Escherichia coli σE regulon. J Biol Chem. 2001;276:20866–20875. doi: 10.1074/jbc.M100464200. [DOI] [PubMed] [Google Scholar]
- Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A. 2000;97:6640–6645. doi: 10.1073/pnas.120163297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies SJ, Golby P, Omrani D, Broad SA, Harrington VL, Guest JR, Kelly DJ, Andrews SC. Inactivation and regulation of the aerobic C4-dicarboxylate transport (dctA) gene of Escherichia coli. J Bacteriol. 1999;181:5624–5635. doi: 10.1128/jb.181.18.5624-5635.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Las Penas A, Connelly L, Gross CA. The σE-mediated response to extracytoplasmic stress in Escherichia coli is transduced by RseA and RseB, two negative regulators of σE. Mol Microbiol. 1997;24:373–385. doi: 10.1046/j.1365-2958.1997.3611718.x. [DOI] [PubMed] [Google Scholar]
- Erickson JW, Gross CA. Identification of the σE subunit of Escherichia coli RNA polymerase: a second alternate sigma factor involved in high-temperature gene expression. Genes Dev. 1989;3:1462–1471. doi: 10.1101/gad.3.9.1462. [DOI] [PubMed] [Google Scholar]
- Fang FC, Libby SJ, Buchmeier NA, Loewen PC, Switala J, Harwood J, Guiney DG. The alternative σ factor KatF (RpoS) regulates Salmonella virulence. Proc Natl Acad Sci USA. 1992;89:11978–11982. doi: 10.1073/pnas.89.24.11978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer D, Teich A, Neubauer P, Hengge-Aronis R. The general stress sigma factor σS of Escherichia coli is induced during diauxic shift from glucose to lactose. J Bacteriol. 1998;180:3203–3206. doi: 10.1128/jb.180.23.6203-6206.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guzman LM, Belin D, Carson MJ, Beckwith J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol. 1995;177:4121–4130. doi: 10.1128/jb.177.14.4121-4130.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall JA, Gehring K, Nikaido H. Two modes of ligand binding in maltose-binding protein of Escherichia coli. Correlation with the structure of ligands and the structure of binding protein. J Biol Chem. 1997;272:17605–17609. doi: 10.1074/jbc.272.28.17605. [DOI] [PubMed] [Google Scholar]
- Horazdovsky BF, Hogg RW. Genetic reconstitution of the high-affinity l-arabinose transport system. J Bacteriol. 1989;171:3053–3059. doi: 10.1128/jb.171.6.3053-3059.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoseith SK, Stocker BAD. Aromatic-dependent Salmonella typhimurium are non-virulent and effective as live vaccines. Nature. 1981;291:238–239. doi: 10.1038/291238a0. [DOI] [PubMed] [Google Scholar]
- Humphreys S, Stevenson A, Bacon A, Weinhardt AB, Roberts M. The alternative sigma factor, σE, is critically important for the virulence of Salmonella Typhimurium. Infect Immun. 1999;67:1560–1568. doi: 10.1128/iai.67.4.1560-1568.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphreys S, Rowley G, Stevenson A, Kenyon WJ, Spector MP, Roberts M. The role of periplasmic peptidyl-prolyl-isomerases in Salmonella virulence. Infect Immun. 2003;71:5386–5388. doi: 10.1128/IAI.71.9.5386-5388.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janausch IG, Zientz E, Tran QH, Kröger A, Unden G. C4-dicarboxylate carriers and sensors in bacteria. Biochim Biophys Acta. 2002;1553:39–56. doi: 10.1016/s0005-2728(01)00233-x. [DOI] [PubMed] [Google Scholar]
- Jones CH, Danese PN, Pinkner JS, Silhavy TJ, Hultgren SJ. The chaperone-assisted membrane release and folding pathway is sensed by two signal transduction systems. EMBO J. 1997;16:6394–6406. doi: 10.1093/emboj/16.21.6394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehres DG, Hogg RW. Escherichia coli K-12 arabinose-binding protein mutants with altered transport properties. Protein Sci. 1992;1:1652–1660. doi: 10.1002/pro.5560011213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenyon WJ, Sayers DG, Humphreys S, Roberts M, Spector MP. The starvation-stress response of Salmonella enterica serovar Typhimurium requires σE, but not CpxR-regulated extra-cytoplasmic functions. Microbiology. 2002;148:113–122. doi: 10.1099/00221287-148-1-113. [DOI] [PubMed] [Google Scholar]
- Lawhon SD, Frye JG, Suyemoto M, Porwollik S, McClelland M, Altier C. Global regulation by CsrA in Salmonella typhimurium. Mol Microbiol. 2003;48:1633–1645. doi: 10.1046/j.1365-2958.2003.03535.x. [DOI] [PubMed] [Google Scholar]
- Lo TC. The molecular mechanism of dicarboxylic acid transport in Escherichia coli K-12. J Supramol Struct. 1977;7:463–480. doi: 10.1002/jss.400070316. [DOI] [PubMed] [Google Scholar]
- Lo TC, Bewick MA. Use of a nonpenetrating substrate analogue to study the molecular mechanism of the outer membrane dicarboxylate transport system in Escherichia coli K-12. J Biol Chem. 1981;256:5511–5517. [PubMed] [Google Scholar]
- Maloy, S. R. (1990). Experimental Techniques in Bacterial Genetics. Boston, MA: Jones & Bartlett.
- McClelland M, Sanderson KE, Spieth J 23 other authors. Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature. 2001;413:852–856. doi: 10.1038/35101614. [DOI] [PubMed] [Google Scholar]
- Mecsas J, Rouviere PE, Erickson JW, Donohue TJ, Gross CA. The activity of σE, an Escherichia coli heat-inducible σ-factor, is modulated by expression of outer membrane proteins. Genes Dev. 1993;7:2618–2628. doi: 10.1101/gad.7.12b.2618. [DOI] [PubMed] [Google Scholar]
- Mecsas J, Welch R, Erickson JW, Gross CA. Identification and characterization of an outer membrane protein, OmpX, in Escherichia coli that is homologous to a family of outer membrane proteins including Ail of Yersinia enterocolitica. J Bacteriol. 1995;177:799–804. doi: 10.1128/jb.177.3.799-804.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, J. H. (1992). A Short Course in Bacterial Genetics: a Laboratory Manual and Handbook for Escherichia coli and Related Bacteria. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory.
- Missiakas D, Betton JM, Raina S. New components of protein folding in extracytoplasmic compartments of Escherichia coli SurA, FkpA and Skp/OmpH. Mol Microbiol. 1996;21:871–884. doi: 10.1046/j.1365-2958.1996.561412.x. [DOI] [PubMed] [Google Scholar]
- Missiakas D, Mayer MP, Lemaire M, Georgopoulos C, Raina S. Modulation of the Escherichia coli σE (RpoE) heat-shock transcription-factor activity by the RseA, RseB and RseC proteins. Mol Microbiol. 1997;24:355–371. doi: 10.1046/j.1365-2958.1997.3601713.x. [DOI] [PubMed] [Google Scholar]
- Miticka H, Rowley G, Rezuchova B, Homerova D, Humphreys S, Farn J, Roberts M, Kormanec J. Transcriptional analysis of the rpoE gene encoding extracytoplasmic stress response sigma factor sigma E in Salmonella enterica serovar Typhimurium. FEMS Microbiol Lett. 2003;226:307–314. doi: 10.1016/S0378-1097(03)00600-1. [DOI] [PubMed] [Google Scholar]
- Moat, A. G., Foster, J. W. & Spector, M. P. (2002). Microbial Physiology, 4th edn. New York: Wiley-Liss.
- Muller N, Heine HG, Boos W. Characterization of the Salmonella typhimurium mgl operon and its gene products. J Bacteriol. 1985;163:37–45. doi: 10.1128/jb.163.1.37-45.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neidhardt FC, Bloch PL, Smith DF. Culture medium for enterobacteria. J Bacteriol. 1974;119:736–747. doi: 10.1128/jb.119.3.736-747.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Notley L, Ferenci T. Differential expression of mal genes under cAMP and endogenous inducer control in nutrient-stressed Escherichia coli. Mol Microbiol. 1995;16:121–129. doi: 10.1111/j.1365-2958.1995.tb02397.x. [DOI] [PubMed] [Google Scholar]
- O’Neal CR, Gabriel WM, Turk AK, Libby SJ, Fang FC, Spector MP. RpoS is necessary for both positive and negative regulation of starvation-survival genes during phosphate, carbon, and nitrogen starvation in Salmonella typhimurium. J Bacteriol. 1994;176:4610–4616. doi: 10.1128/jb.176.15.4610-4616.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ponting CP. Evidence for PDZ domains in bacteria, yeast, and plants. Protein Sci. 1997;6:464–468. doi: 10.1002/pro.5560060225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Postma PW. Galactose transport in Salmonella typhimur-ium. J Bacteriol. 1977;129:630–639. doi: 10.1128/jb.129.2.630-639.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Postma PW, Lengeler JW, Jacobson GR. Phosphoenolpyruvate:carbohydrate phosphotransferase systems of bacteria. Microbiol Rev. 1993;57:543–594. doi: 10.1128/mr.57.3.543-594.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rabus R, Jack DL, Kelly DJ, Saier MH., Jr TRAP transporters: an ancient family of extracytoplasmic solute-receptor-dependent secondary active transporters. Microbiology. 1999;145:3431–3445. doi: 10.1099/00221287-145-12-3431. [DOI] [PubMed] [Google Scholar]
- Schulein K, Benz R. LamB (maltoporin) of Salmonella typhimurium: isolation, purification and comparison of sugar binding with LamB of Escherichia coli. Mol Microbiol. 1990;4:625–632. doi: 10.1111/j.1365-2958.1990.tb00631.x. [DOI] [PubMed] [Google Scholar]
- Seymour RL, Mishra PV, Khan MA, Spector MP. Essential roles of core starvation-stress response loci in carbon-starvation-inducible cross-resistance and hydrogen peroxide-inducible adaptive resistance to oxidative challenge in Salmonella typhimurium. Mol Microbiol. 1996;20:497–505. doi: 10.1046/j.1365-2958.1996.5451068.x. [DOI] [PubMed] [Google Scholar]
- Simons RW, Houman F, Kleckner N. Improved single and multi-copy lac-based cloning vectors for protein and operon fusions. Gene. 1987;53:85–96. doi: 10.1016/0378-1119(87)90095-3. [DOI] [PubMed] [Google Scholar]
- Songyang Z, Fanningm AS, Fu C 7 other authors. Recognition of unique carboxyl-terminal motifs by distinct PDZ domains. Science. 1997;275:73–77. doi: 10.1126/science.275.5296.73. [DOI] [PubMed] [Google Scholar]
- Spector MP. The starvation-stress response (SSR) of Salmonella. Adv Microb Physiol. 1998;40:235–279. doi: 10.1016/s0065-2911(08)60133-2. [DOI] [PubMed] [Google Scholar]
- Spector MP, Cubitt CL. Starvation-inducible loci of Salmonella typhimurium, regulation and roles in starvation survival. Mol Microbiol. 1992;6:1467–1476. doi: 10.1111/j.1365-2958.1992.tb00867.x. [DOI] [PubMed] [Google Scholar]
- Spector MP, Park YK, Tirgari S, Gonzalez T, Foster JW. Identification and characterization of starvation-regulated genetic loci in Salmonella typhimurium by using Mu d-directed lacZ operon fusion. J Bacteriol. 1988;170:345–351. doi: 10.1128/jb.170.1.345-351.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spector MP, DiRusso CC, Pallen MJ, Garcia del Portillo F, Dougan G, Finlay BB. The medium-/long-chain fatty acyl-CoA dehydrogenase (fadF) gene of Salmonella typhimurium is a phase 1 starvation-stress response (SSR) locus. Microbiology. 1999a;145:15–31. doi: 10.1099/13500872-145-1-15. [DOI] [PubMed] [Google Scholar]
- Spector MP, Garcia del Portillo F, Bearson SM, Mahmud A, Magut M, Finlay BB, Dougan G, Foster JW, Pallen MJ. The rpoS-dependent starvation-stress response locus stiA encodes a nitrate reductase (narZYWV) required for carbon-starvation-inducible thermotolerance and acid tolerance in Salmonella typhimurium. Microbiology. 1999b;145:3035–3045. doi: 10.1099/00221287-145-11-3035. [DOI] [PubMed] [Google Scholar]
- Tanaka K, Takayanagi Y, Fujita N, Ishihama A, Takahashi H. Heterogeneity of the principal σ factor in Escherichia coli: the rpoS gene product, σ38, is a second principal factor of RNA polymerase in stationary-phase Escherichia coli. Proc Natl Acad Sci U S A. 1993;90:3511–3515. doi: 10.1073/pnas.90.8.3511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Testerman TL, Vasquez-Torres A, Xu Y, Jones-Carson J, Libby SJ, Fang FC. The alternative sigma factor σE controls antioxidant defences required for Salmonella virulence and stationary-phase survival. Mol Microbiol. 2002;43:771–782. doi: 10.1046/j.1365-2958.2002.02787.x. [DOI] [PubMed] [Google Scholar]
- Walsh NP, Alba BM, Bose B, Gross CA, Sauer RT. OMP peptide signals initiate the envelope-stress response by activating DegS protease via relief of inhibition mediated by its PDZ domain. Cell. 2003;113:61–71. doi: 10.1016/s0092-8674(03)00203-4. [DOI] [PubMed] [Google Scholar]
- Winnen B, Hvorup RN, Saier MH., Jr The tripartite tricarboxylate transporter (TTT) family. Res Microbiol. 2003;154:457–465. doi: 10.1016/S0923-2508(03)00126-8. [DOI] [PubMed] [Google Scholar]
- Widenhorn KA, Somers JM, Kay WW. Expression of the divergent tricarboxylate transport operon (tctI) of Salmonella typhimurium. J Bacteriol. 1988;170:3223–3227. doi: 10.1128/jb.170.7.3223-3227.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]