

Abstract
The intestinal microflora is a positive health asset that crucially influences the normal structural and functional development of the mucosal immune system. Mucosal immune responses to resident intestinal microflora require precise control and an immunosensory capacity for distinguishing commensal from pathogenic bacteria. In genetically susceptible individuals, some components of the flora can become a liability and contribute to the pathogenesis of various intestinal disorders, including inflammatory bowel diseases. It follows that manipulation of the flora to enhance the beneficial components represents a promising therapeutic strategy. The flora has a collective metabolic activity equal to a virtual organ within an organ, and the mechanisms underlying the conditioning influence of the bacteria on mucosal homeostasis and immune responses are beginning to be unravelled. An improved understanding of this hidden organ will reveal secrets that are relevant to human health and to several infectious, inflammatory and neoplastic disease processes.
Keywords: commensal bacteria, homeostasis, intestinal epithelium
Introduction
Host–microbe interactions occur primarily along mucosal surfaces, and one of the largest interfaces is the human intestinal mucosa. The intestine is adapted to bi-directional host–flora exchange and harbours a diverse bacterial community that is separated from the internal milieu by only a single layer of epithelial cells. Resident bacteria outnumber human somatic and germ cells tenfold and represent a combined microbial genome well in excess of the human genome (Shanahan, 2002). Collectively, the flora has a metabolic activity equal to a virtual organ within an organ (Bocci, 1992).
Most bacterial species cannot be cultured, but modern molecular methods, such as broad-range sequencing of 16S ribosomal RNA from amplified bacterial nucleic acid extracted from faeces or biopsies, indicate evolutionary divergence that can be used to identify and classify bacteria. The availability of bacterial sequence data has facilitated the development of molecular probes for fluorescence in situ hybridization, DNA microarrays and gene chips that can identify and enumerate specific species. These molecular approaches have been used to examine the individuality and stability of the flora over time and to detect shifts in its composition after weaning, exposure to antibiotics or dietary changes. Although the intestinal flora of an adult alters with lifestyle, diet and age (Hopkins et al, 2001), the prevailing influence of the host genotype over environmental factors on an individual's microbial diversity has been shown in a comparative study of the flora of adults with varying degrees of genetic relatedness (Zoetendal et al, 2001).
The structure and composition of the gut flora reflects natural selection at both the microbial and host levels, which promotes mutual cooperation within and functional stability of this complex ecosystem. Although bacteria predominate, archaea and eukarya are also represented. Acid, bile and pancreatic secretions hinder the colonization of the stomach and proximal small intestine by most bacteria. However, bacterial density increases in the distal small intestine, and in the large intestine rises to an estimated 1011–1012 bacteria per gram of colonic content, which contributes to 60% of faecal mass (Fig 1A). In addition to variations in the composition of the flora along the axis of the gastrointestinal tract, surface-adherent and luminal microbial populations also differ (Eckburg et al, 2005), and the ratio of anaerobes to aerobes is lower at the mucosal surfaces than in the lumen.
Figure 1.
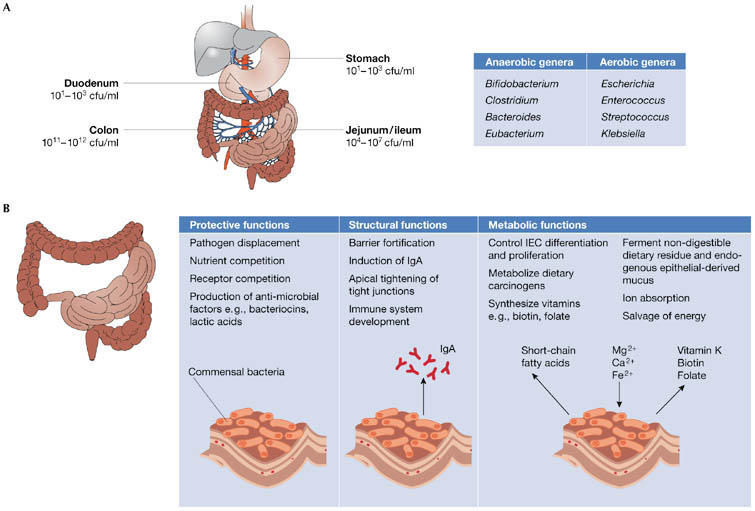
Functions of the intestinal flora. (A) Bacteria density increases in the jejunum/ileum from the stomach and duodenum, and in the large intestine, colon-residing bacteria achieve the highest cell densities recorded for any ecosystem. The most common anaerobic and aerobic genera are listed. (B) Commensal bacteria exert a miscellany of protective, structural and metabolic effects on the intestinal mucosa.
The fetal gut is sterile but colonization begins immediately after birth and is influenced by the mode of delivery, the infant diet, hygiene levels and medication (Gronlund et al, 1999). Enterobacteria and bifidobacteria represent early colonizers, although differences in gut microflora composition and the incidence of infection occur between breast- and formula-fed infants (Mountzouris et al, 2002). It seems that these pioneering bacteria can modulate gene expression in the host to create a suitable environment for themselves and can prevent growth of other bacteria introduced later to the ecosystem (Xu & Gordon, 2003).
Lessons from a life without intestinal microflora
Enteric bacteria form a natural defence barrier and exert numerous protective, structural and metabolic effects on the epithelium (Fig 1B). Their influence on intestinal physiology has been shown in comparative studies of germ-free and colonized animals. Germ-free animals are more susceptible to infection and have reduced vascularity, digestive enzyme activity, muscle wall thickness, cytokine production ands serum immunoglobulin levels, smaller Peyer's patches and fewer intraepithelial lymphocytes, but increased enterochromaffin cell area (Shanahan, 2002). However, reconstitution of germ-free mice with an intestinal microflora is sufficient to restore the mucosal immune system (Umesaki et al, 1995). Indeed, colonization of germ-free mice with a single species, Bacteroides thetaiotaomicron, affects the expression of various host genes that influence nutrient uptake, metabolism, angiogenesis, mucosal barrier function and the development of the enteric nervous system (Xu & Gordon, 2003). Moreover, ligands from commensal bacteria and commensal-derived symbiosis factors influence the normal development and function of the mucosal immune system (Mazmanian et al, 2005; Rakoff-Nahoum et al, 2004). Commensal bacteria profoundly influence the development of humoral components of the gut mucosal immune system (Weinstein & Cebra, 1991) and also modulate the fine-tuning of T-cell repertoires and T-helper (Th)-cell type 1 or type 2 cytokine profiles (Cebra, 1999; Shanahan, 2002). Thus, it is possible that the composition of the colonizing flora influences individual variations in immunity.
The intestinal microbiome has a metabolic activity that is both adaptable and renewable (Bocci, 1992). Through the production of short-chain fatty acids, resident bacteria positively influence intestinal epithelial cell differentiation and proliferation, and mediate other metabolic effects (Fig 1B; Shanahan, 2002). Together, this complex metabolic activity recovers valuable energy and absorbable substrates for the host, and provides energy and nutrients for bacterial growth and proliferation. Colonization increases the uptake of glucose in the intestine and, compared with colonized mice, germ-free mice require a greater caloric intake to sustain a normal body weight (Backhed et al, 2004). This implicates gut bacteria as modulators of fat deposition in the host.
Host–flora communication at the mucosal surface
Host defence requires an accurate interpretation of the microenvironment to distinguish commensal organisms from episodic pathogens and a precise regulation of subsequent responses. The epithelium provides the first sensory line of defence and active sampling of resident bacteria, pathogens and other antigens is mediated by three main types of immunosensory cell (Fig 2). First, surface enterocytes serve as afferent sensors of danger within the luminal microenvironment by secreting chemokines and cytokines that alert and direct innate and adaptive immune responses to the infected site (Shanahan, 2005). Second, M cells that overlie lymphoid follicles sample the environment and transport luminal antigens to subadjacent dendritic cells and other antigen-presenting cells. Third, intestinal dendritic cells themselves have a pivotal immunosensory role and can directly sample gut contents by either entering or extending dendrites between surface enterocytes without disrupting tight junctions (Rescigno et al, 2001). Dendritic cells can ingest and retain live commensal bacteria and travel to the mesenteric lymph node where immune responses to commensal bacteria are induced locally (Macpherson & Uhr, 2004). Thus, acting as a gatekeeper, the mesenteric lymph node prevents access of commensal bacteria to the internal milieu.
Figure 2.
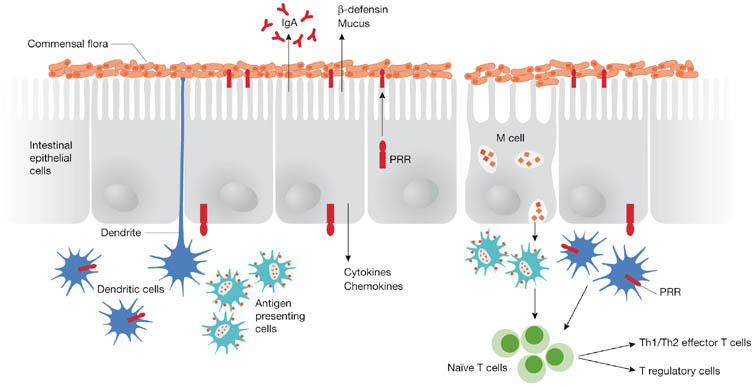
Immunosensory detection of intestinal bacteria. Surface enterocytes secrete many immune mediators in response to antigens, including antibacterial peptides, immunoglobulin A (IgA) and chemokines. Specialized epithelial cells, termed M cells, transport and deliver antigens to antigen-presenting cells, which subsequently process antigens and present them to naïve T cells. Antigen-presenting dendritic cells (DCs) also survey and sample the mucosal microenvironment. Pattern recognition receptors (PRRs) expressed by DCs and enterocytes mediate the detection of bacterial antigens, and DCs modulate immune responsiveness or tolerance by promoting either effector or regulatory T cells.
The ability of immunosensory cells to discriminate pathogenic from commensal bacteria is mediated, in part, by two major host pattern recognition receptor (PRR) systems—the family of Toll-like receptors (TLRs) and the nucleotide-binding oligomerization domain/caspase recruitment domain isoforms (NOD/CARD; Cario, 2005). These PRRs have a fundamental role in immune-cell activation in response to specific microbial-associated molecular patterns. For example, TLR2 is activated by peptidoglycan and lipotechoic acids, TLR4 by lipopolysaccharide, TLR5 by flagellin, and NOD1/CARD4 and NOD2/CARD15 function as intracellular receptors of peptidoglycan subunits.
TLRs and NOD proteins are expressed by surface enterocytes and dendritic cells (Abreu et al, 2005), and in the gut PRRs seem to be crucial for bacterial–host communication. Decreased enterocyte proliferation and levels of cytoprotective factors have been observed in TLR-defective mice, and TLR signals mediated by commensal bacteria or their ligands are essential for intestinal barrier function and repair of the gut (Fukata et al, 2005; Rakoff-Nahoum et al, 2004). Many PRR ligands are expressed by commensal bacteria, yet the healthy gut does not evoke inflammatory responses to these bacteria. Conversely, some commensal bacteria exert protective effects by attenuating pro-inflammatory responses induced by various enteropathogenic bacteria (Kelly et al, 2004; Ma et al, 2004; O'Hara et al, 2006). The host and bacterial mechanisms that underpin these effects are being explored.
Host systems that contribute to homeostasis. In the healthy gut, TLR expression profiles contribute to homeostasis. Normal enterocytes express low levels of TLR2, TLR4 and the co-receptor MD-2, and lack membrane-bound CD14, which is a co-receptor for lipopolysaccharide (Abreu et al, 2001; Otte et al, 2004). There have been conflicting reports relating to the restricted expression of certain TLRs in the gut (Kelly et al, 2005). In particular, expression of TLR5 in vitro seems to be basolateral (Gewirtz et al, 2001), but TLR5 is expressed on both the apical and basolateral poles in vivo. This illustrates that intestinal epithelial cell lines might express different levels, or have a different cellular distribution, of TLRs compared with normal gut mucosa. In gastric epithelial cells, TLR5 can redistribute to a predominantly basolateral localization in response to Helicobacter pylori infection (Cario & Podolsky, 2000; Schmausser et al, 2004), suggesting that under certain conditions TLR5 redistributes to the basolateral membrane where it is ideally positioned to detect translocated flagellin. In addition, intestinal enterocytes constitutively or inducibly express high levels of the TLR inhibitor Toll-interacting protein (Tollip; Otte et al, 2004). Tollip expression correlates directly with the luminal bacterial load in vivo and is highest in healthy colonic mucosa. Surface enterocytes also express high levels of a Toll/interleukin-1 receptor-containing inhibitory molecule referred to as single immunoglobulin IL-1R-related molecule (SIGIRR). SIGIRR-deficient animals are more susceptible to experimentally induced colitis, suggesting a role for SIGIRR in tuning mucosal tolerance towards commensal flora (Garlanda et al, 2004). Additional TLR suppressors, such as A20 and TRIAD3A, have been identified in other systems (Boone et al, 2004; Chuang & Ulevitch, 2004), but whether these mediators regulate TLR signalling in the gut is unknown.
Recent evidence suggests that, similar to Tollip, NOD2 might suppress inflammatory cascades, and mutations of NOD2 are associated with Crohn's disease (Bairead et al, 2003). NOD2 can modulate signals transmitted through TLR3, TLR4 and TLR9 (Netea et al, 2005; van Heel et al, 2005). Although wild-type NOD2 activates pro-inflammatory signals, stimulation of NOD2 with peptidoglycan has been shown to inhibit TLR2-driven Th1 cytokine responses, and in the absence of NOD2 peptidoglycan triggers imbalanced TLR2-mediated cytokine production (Watanabe et al, 2004). Conversely, peptidoglycan induces a pro-inflammatory phenotype in mutant mice expressing dysfunctional NOD2 (Maeda et al, 2005), suggesting that in some situations NOD2 mutations might lead to a gain-of-function and elevated pro-inflammatory cytokine production. In the healthy gut, excessive Th1 responses to the resident flora are also inhibited by the controlling influence of regulatory T cells and tolerance-inducing dendritic cells (Rook & Brunet, 2005).
Effector mechanisms of the commensal flora. Host inflammatory responses to pathogenic bacteria and other stress signals are pivotally controlled by the transcription factor nuclear factor (NF)-κB. Most commensal bacteria do not activate NF-κB, but rather certain species can restrain inflammatory signals in response to Salmonella typhimurium and its flagellin through pathways that seem to involve NF-κB (O'Hara et al, 2006). Several distinct mechanisms by which commensal bacteria limit NF-κB signalling have been elucidated (Fig 3). These include inhibition of epithelial proteasome function, degradation of the NF-κB counter-regulatory factor IκBα or nuclear export of the NF-κB subunit, p65, through a peroxisome proliferator-activated receptor (PPAR)γ-dependent pathway (Kelly et al, 2004; Neish et al, 2000; Petrof et al, 2004). Some commensal bacteria might inhibit specific signalling via TLR4 by elevating PPARγ expression and uncoupling NF-κB-dependent target genes in a negative-feedback loop (Dubuquoy et al, 2003). Induction of transforming growth factor-β and nerve growth factor, and mitogen-activated protein kinase and protein kinase B pathways have also been implicated in the anti-inflammatory effects mediated by various commensal bacteria (Ma et al, 2004; Yan & Polk, 2002). Moreover, probiotic DNA has been shown to restrain cytokine-induced pro-inflammatory responses in intestinal enterocytes (Jijon et al, 2004), but the precise immunomodulatory impact of bio-available commensal DNA in vivo requires investigation.
Figure 3.
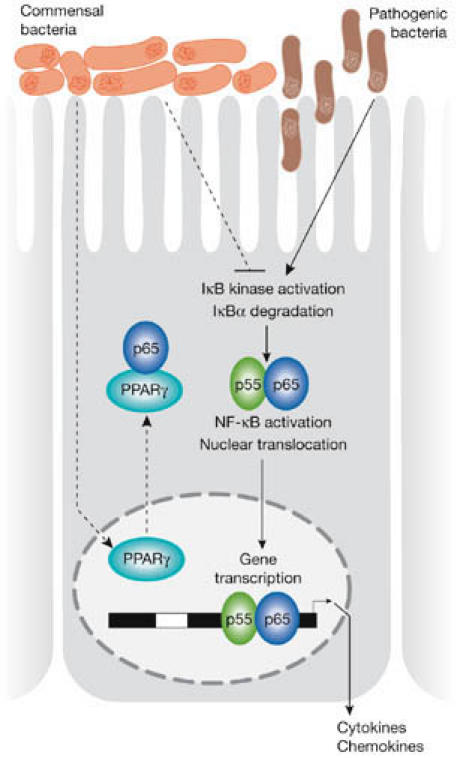
Schematic illustration of the mechanisms by which some commensal bacteria limit pathogen-induced nuclear factor (NF)-κB signalling. Pathogenic bacteria such as Salmonella typhimurium trigger IκB kinase activation, IκBα degradation and nuclear translocation of p50/p65 NF-κB subunits. Some commensal bacteria offset these affects by promoting the nuclear export of activated p65 through associations with peroxisome proliferator-activated receptor (PPAR)γ, thereby terminating promoter activation. Other commensal bacteria inhibit IκBα degradation.
Molecular mimicry of host molecules, whereby bacteria display surface molecules resembling those of the surface of the host, could contribute to immune hyporesponsiveness to some resident bacteria. It has been shown that the synthesis of fucosylated surface molecules by the commensal strain Bacteroides fragilis confers a competitive survival advantage, which allows these bacteria to reside in close contact with intestinal enterocytes that are covered with similar fucosylated glycoproteins and glycolipids (Coyne et al, 2005). Commensal-derived metabolites and the composition of bacterial surface structures can also have various impacts on the host immune system (Grangette et al, 2005; Menard et al, 2004). Recent comparative genomic analysis and bioinformatic data mining have revealed the presence of type III and type IV secretion systems in commensal bacteria (Nagai & Roy, 2003; Tampakaki et al, 2004). Hitherto such secretion systems had only been described for pathogenic bacteria (Backert & Selbach, 2005), and now warrant studies to determine whether commensal bacteria use analogous secretion systems to deliver effector molecules that subsequently mediate evasion of host immune surveillance mechanisms in the gut.
The silent partner in disease manifestation
The gut flora is normally a health asset, but in genetically susceptible individuals some components of the flora can become a liability. Aberrant immune responses to luminal antigens underlie the manifestations of inflammatory bowel disease (IBD), which comprises Crohn's disease and ulcerative colitis, and in susceptible individuals, tissue damage results from immunological misperception of danger within the indigenous flora or from failure of normal tolerance to enteric bacteria (Shanahan, 2002). It remains unclear whether the inflammatory responses in IBD are elicited in response to a specific subset of intestinal microbes or whether tolerance to commensal bacteria in general is affected.
Innate immune responses to the commensal flora educate the immune system and influence adaptive responses to exogenous antigens. With increased sanitation and hygiene in developed nations, genes that were once survival factors in an earlier era could become risk factors for immune-hypersensitivity disorders in a modern sanitized environment (Rook & Brunet, 2005). Under certain circumstances, commensal bacteria have been shown to fuel the progression towards colorectal malignancy by generating reactive metabolites, converting pro-carcinogens to carcinogens and altering host carbohydrate expression (Hope et al, 2005). Recent reports suggest that the gut microbiota of an individual has a specific metabolic efficiency, and differences in gut flora composition between individuals might regulate energy storage and even a predisposition to obesity (Backhed et al, 2004). Moreover, the microflora itself is a substantial consumer of energy, and increasing adiposity has been shown to mediate changes in the diversity of the gut flora (Ley et al, 2005).
Conventional therapies for IBD and atopy primarily target mucosal inflammatory responses, but the contribution of the indigenous flora to certain clinical manifestations underpins the rationale for therapeutic manipulation of enteric bacteria using pharmabiotics. Pharmabiotic is a generic term to encompass any form of therapeutic exploitation of the commensal flora, including the use of live probiotic bacteria, probiotic-derived biologically active metabolites, prebiotics, synbiotics or genetically modified commensal bacteria. Indeed, there is evidence supporting a therapeutic role for probiotic strategies for treating mucosal inflammatory disorders such as IBD, atopy, infection, diarrhoea, cancer and arthritis (Shanahan, 2005; Yan & Polk, 2004). The concept that gut microbial diversity is linked to obesity deserves extensive exploration, and the assessment of energy-efficient species in lean compared with obese individuals might yield new treatment strategies for chronic obesity.
Conclusions
The 2005 Nobel prize in Physiology and Medicine awarded to Robin Warren and Barry Marshall is a reminder that the solution to some human diseases does not reside solely within the host but rather might be found at the interface with the microbial environment. Manipulation of the flora is becoming a realistic therapeutic and prophylactic strategy for many infectious, inflammatory and even neoplastic diseases within the gut. However, the promise of pharmabiotics is unlikely to be completely fulfilled without greater attention to the secrets held within the forgotten inner organ represented by the enteric microflora. The flora might be a rich repository of metabolites that can be exploited for therapeutic benefit. Elucidating the molecular details of host–flora interactions is, therefore, a pre-requisite for a ‘bugs to drugs' programme of discovery.

A.M.O'Hara & F. Shanahan
Acknowledgments
The authors are supported in part by Science Foundation Ireland. F. S. has been affiliated with a campus-based company (Alimentary Health Ltd). The content of this article was neither influenced nor constrained by this fact.
References
- Abreu MT, Vora P, Faure E, Thomas LS, Arnold ET, Arditi M (2001) Decreased expression of Toll-like receptor-4 and MD-2 correlates with intestinal epithelial cell protection against dysregulated proinflammatory gene expression in response to bacterial lipopolysaccharide. J Immunol 167: 1609–1616 [DOI] [PubMed] [Google Scholar]
- Abreu MT, Fukata M, Arditi M (2005) TLR signaling in the gut in health and disease. J Immunol 174: 4453–4460 [DOI] [PubMed] [Google Scholar]
- Backert S, Selbach M (2005) Tyrosine-phosphorylated bacterial effector proteins: the enemies within. Trends Microbiol 13: 476–484 [DOI] [PubMed] [Google Scholar]
- Backhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, Semenkovich CF, Gordon JI (2004) The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA 101: 15718–15723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bairead E et al. (2003) Association of NOD2 with Crohn's disease in a homogenous Irish population. Eur J Hum Genet 11: 237–244 [DOI] [PubMed] [Google Scholar]
- Bocci V (1992) The neglected organ: bacterial flora has a crucial immunostimulatory role. Perspect Biol Med 35: 251–260 [DOI] [PubMed] [Google Scholar]
- Boone DL et al. (2004) The ubiquitin-modifying enzyme A20 is required for termination of Toll-like receptor responses. Nat Immunol 5: 1052–1060 [DOI] [PubMed] [Google Scholar]
- Cario E (2005) Bacterial interactions with cells of the intestinal mucosa: Toll-like receptors and NOD2. Gut 54: 1182–1193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cario E, Podolsky DK (2000) Differential alteration in intestinal epithelial cell expression of Toll-like receptor-3 (TLR3) and TLR4 in inflammatory bowel disease. Infect Immun 68: 7010–7017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cebra JJ (1999) Influences of microbiota on intestinal immune system development. Am J Clin Nutr 69: 1046S–1051S [DOI] [PubMed] [Google Scholar]
- Chuang TH, Ulevitch RJ (2004) Triad3A, an E3 ubiquitin-protein ligase regulating Toll-like receptors. Nat Immunol 5: 495–502 [DOI] [PubMed] [Google Scholar]
- Coyne MJ, Reinap B, Lee MM, Comstock LE (2005) Human symbionts use a host-like pathway for surface fucosylation. Science 307: 1778–1781 [DOI] [PubMed] [Google Scholar]
- Dubuquoy L, Jansson EA, Deeb S, Rakotobe S, Karoui M, Colombel JF, Auwerx J, Pettersson S, Desreumaux P (2003) Impaired expression of peroxisome proliferator-activated receptor γ in ulcerative colitis. Gastroenterology 124: 1265–1276 [DOI] [PubMed] [Google Scholar]
- Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA (2005) Diversity of the human intestinal microbial flora. Science 308: 1635–1638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukata M et al. (2005) Toll-like receptor-4 is required for intestinal response to epithelial injury and limiting bacterial translocation in a murine model of acute colitis. Am J Physiol Gastrointest Liver Physiol 288: G1055–G1065 [DOI] [PubMed] [Google Scholar]
- Garlanda C et al. (2004) Intestinal inflammation in mice deficient in Tir8, an inhibitory member of the IL-1 receptor family. Proc Natl Acad Sci USA 101: 3522–3526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gewirtz AT, Navas TA, Lyons S, Godowski PJ, Madara JL (2001) Cutting edge: bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J Immunol 167: 1882–1885 [DOI] [PubMed] [Google Scholar]
- Grangette C et al. (2005) Enhanced antiinflammatory capacity of a Lactobacillus plantarum mutant synthesizing modified teichoic acids. Proc Natl Acad Sci USA 102: 10321–10326 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gronlund MM, Lehtonen OP, Eerola E, Kero P (1999) Fecal microflora in healthy infants born by different methods of delivery: permanent changes in intestinal flora after cesarean delivery. J Pediatr Gastroenterol Nutr 28: 19–25 [DOI] [PubMed] [Google Scholar]
- Hope ME, Hold GL, Kain R, El-Omar EM (2005) Sporadic colorectal cancer—role of the commensal microbiota. FEMS Microbiol Lett 244: 1–7 [DOI] [PubMed] [Google Scholar]
- Hopkins MJ, Sharp R, Macfarlane GT (2001) Age and disease related changes in intestinal bacterial populations assessed by cell culture, 16S rRNA abundance, and community cellular fatty acid profiles. Gut 48: 198–205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jijon H, Backer J, Diaz H, Yeung H, Thiel D, McKaigney C, De Simone C, Madsen K (2004) DNA from probiotic bacteria modulates murine and human epithelial and immune function. Gastroenterology 126: 1358–1373 [DOI] [PubMed] [Google Scholar]
- Kelly D, Campbell JI, King TP, Grant G, Jansson EA, Coutts AG, Pettersson S, Conway S (2004) Commensal anaerobic gut bacteria attenuate inflammation by regulating nuclear-cytoplasmic shuttling of PPAR-γ and RelA. Nat Immunol 5: 104–112 [DOI] [PubMed] [Google Scholar]
- Kelly D, Conway S, Aminov R (2005) Commensal gut bacteria: mechanisms of immune modulation. Trends Immunol 26: 326–333 [DOI] [PubMed] [Google Scholar]
- Ley RE, Backhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI (2005) Obesity alters gut microbial ecology. Proc Natl Acad Sci USA 102: 11070–11075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma D, Forsythe P, Bienenstock J (2004) Live Lactobacillus reuteri is essential for the inhibitory effect on tumor necrosis factor α-induced interleukin-8 expression. Infect Immun 72: 5308–5314 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macpherson AJ, Uhr T (2004) Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science 303: 1662–1665 [DOI] [PubMed] [Google Scholar]
- Maeda S, Hsu LC, Liu H, Bankston LA, Iimura M, Kagnoff MF, Eckmann L, Karin M (2005) Nod2 mutation in Crohn's disease potentiates NF-κB activity and IL-1β processing. Science 307: 734–738 [DOI] [PubMed] [Google Scholar]
- Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL (2005) An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 122: 107–118 [DOI] [PubMed] [Google Scholar]
- Menard S, Candalh C, Bambou JC, Terpend K, Cerf-Bensussan N, Heyman M (2004) Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after intestinal transport. Gut 53: 821–828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mountzouris KC, McCartney AL, Gibson GR (2002) Intestinal microflora of human infants and current trends for its nutritional modulation. Br J Nutr 87: 405–420 [DOI] [PubMed] [Google Scholar]
- Nagai H, Roy CR (2003) Show me the substrates: modulation of host cell function by type IV secretion systems. Cell Microbiol 5: 373–383 [DOI] [PubMed] [Google Scholar]
- Neish AS, Gewirtz AT, Zeng H, Young AN, Hobert ME, Karmali V, Rao AS, Madara JL (2000) Prokaryotic regulation of epithelial responses by inhibition of IκB-α ubiquitination. Science 289: 1560–1563 [DOI] [PubMed] [Google Scholar]
- Netea MG et al. (2005) Nucleotide-binding oligomerization domain-2 modulates specific TLR pathways for the induction of cytokine release. J Immunol 174: 6518–6523 [DOI] [PubMed] [Google Scholar]
- O'Hara AM, O'Regan P, Fanning A, O'Mahony C, MacSharry J, Lyons A, Bienenstock J, O'Mahony L, Shanahan F (2006) Functional modulation of human intestinal epithelial cell responses by Bifidobacterium infantis and Lactobacillus salivarius. Immunology 118: 202–215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otte JM, Cario E, Podolsky DK (2004) Mechanisms of cross hyporesponsiveness to Toll-like receptor bacterial ligands in intestinal epithelial cells. Gastroenterology 126: 1054–1070 [DOI] [PubMed] [Google Scholar]
- Petrof EO, Kojima K, Ropeleski MJ, Musch MW, Tao Y, De Simone C, Chang EB (2004) Probiotics inhibit nuclear factor-κB and induce heat shock proteins in colonic epithelial cells through proteasome inhibition. Gastroenterology 127: 1474–1487 [DOI] [PubMed] [Google Scholar]
- Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, Edberg S, Medzhitov R (2004) Recognition of commensal microflora by Toll-like receptors is required for intestinal homeostasis. Cell 118: 229–241 [DOI] [PubMed] [Google Scholar]
- Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, Granucci F, Kraehenbuhl JP, Ricciardi-Castagnoli P (2001) Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat Immunol 2: 361–367 [DOI] [PubMed] [Google Scholar]
- Rook GA, Brunet LR (2005) Microbes, immunoregulation and the gut. Gut 54: 317–320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmausser B, Andrulis M, Endrich S, Lee SK, Josenhans C, Muller-Hermelink HK, Eck M (2004) Expression and subcellular distribution of Toll-like receptors TLR4, TLR5 and TLR9 on the gastric epithelium in Helicobacter pylori infection. Clin Exp Immunol 136: 521–526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shanahan F (2002) The host–microbe interface within the gut. Best Pract Res Clin Gastroenterol 16: 915–931 [DOI] [PubMed] [Google Scholar]
- Shanahan F (2005) Physiological basis for novel drug therapies used to treat the inflammatory bowel diseases. I. Pathophysiological basis and prospects for probiotic therapy in inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol 288: G417–G421 [DOI] [PubMed] [Google Scholar]
- Tampakaki AP, Fadouloglou VE, Gazi AD, Panopoulos NJ, Kokkinidis M (2004) Conserved features of type III secretion. Cell Microbiol 6: 805–816 [DOI] [PubMed] [Google Scholar]
- Umesaki Y, Okada Y, Matsumoto S, Imaoka A, Setoyama H (1995) Segmented filamentous bacteria are indigenous intestinal bacteria that activate intraepithelial lymphocytes and induce MHC class II molecules and fucosyl asialo GM1 glycolipids on the small intestinal epithelial cells in the ex-germ-free mouse. Microbiol Immunol 39: 555–562 [DOI] [PubMed] [Google Scholar]
- van Heel DA, Ghosh S, Hunt KA, Mathew CG, Forbes A, Jewell DP, Playford RJ (2005) Synergy between TLR9 and NOD2 innate immune responses is lost in genetic Crohn's disease. Gut 54: 1553–1557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe T, Kitani A, Murray PJ, Strober W (2004) NOD2 is a negative regulator of Toll-like receptor-2-mediated T helper type 1 responses. Nat Immunol 5: 800–808 [DOI] [PubMed] [Google Scholar]
- Weinstein PD, Cebra JJ (1991) The preference for switching to IgA expression by Peyer's patch germinal center B cells is likely due to the intrinsic influence of their microenvironment. J Immunol 147: 4126–4135 [PubMed] [Google Scholar]
- Xu J, Gordon JI (2003) Inaugural Article: Honor thy symbionts. Proc Natl Acad Sci USA 100: 10452–10459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan F, Polk DB (2002) Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells. J Biol Chem 277: 50959–50965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan F, Polk DB (2004) Commensal bacteria in the gut: learning who our friends are. Curr Opin Gastroenterol 20: 565–571 [DOI] [PubMed] [Google Scholar]
- Zoetendal EG, Akkermans ADL, Akkermans-van Vilet WM, DeVisser JAGM, DeVos WM (2001) The host genotype affects the bacterial community in the human gastrointestinal tract. Microb Ecol Health Dis 13: 129–134 [Google Scholar]