

Abstract
The “Secretor” gene (FUT2) encodes an α(1,2)fucosyltransferase (E.C. 2.4.1.69) that elaborates α(1,2)fucose residues on mucosal epithelium and secreted mucins. While uterine α(1,2)fucosylated glycans have proposed to be involved in embryo adhesion, mice with a homozygous null mutation of Fut2 displayed normal fertility. To help develop alternative hypotheses for function, the cell type and regulation of Fut2 expression during the estrous cycle, hormone replacement, and pregnancy was examined in Fut2-LacZ reporter mice containing targeted replacement of Fut2 with bacterial lacZ. LacZ expression in the reproductive tract of Fut2-LacZ mice is most prominent in the glandular epithelium of the endocervix during estrus and pregnancy. Nuclear LacZ expression identifies cell specific expression of Fut2 in mucus secreting cells of the endocervix, uterine glands, foveolar pit and chief cells of the stomach, and goblet cells of the colon. In ovariectomized Fut2-LacZ mice, estradiol treatment stimulates X-gal staining in endocervix and uterus, but does not affect expression in stomach and colon. Northern blot analysis in wild type mice shows 15-fold elevations of Fut2 steady-state mRNA with estradiol treatment, while Fut1 varies little. Fut2 levels in the glandular stomach and distal colon remain constant while uterine Fut2 levels vary 8-fold during the estrous cycle. These data represent the first demonstration of a glycosyltransferase gene under tissue-specific hormonal regulation in a LacZ reporter mouse model. Endocervical expression of Fut2 in estrus and pregnancy may modify cervical mucus barrier properties from microbial infection analogous to the potential role of mucosal glycans in humans.
Keywords: endocervical glands, fucosyltransferase, glandular stomach, mucus
Abbreviations: FUT2, α(1,2)fucosyltransferase "Secretor" gene; FUT1, α(1,2)fucosyltransferase "H" gene; ER-α, estrogen receptor-α
Introduction
Post-translational modification of cellular glycans with fucose is essential to embryonic growth and development, fertility, and immune function (Smith et al., 2002). The enzymes responsible for synthesis of fucosylated glycans, called fucosyltransferases, are expressed in tissue-specific patterns. Alterations in the expression of fucosylated glycans, and their cognate fucosyltransferases, have been observed in multiple pathologic processes involving inflammation, cancer, and numerous ontogenic events involving signaling events by the Notch receptor family (Becker and Lowe, 2003). Mouse embryos lacking the protein O-fucosyltransferase 1 of the Notch pathway die at mid gestation (Shi and Stanley, 2003).
Classes of fucosyltransferase are divided by the glycosidic linkage catalyzed (Breton et al., 1998). FUT2, the "Secretor" gene, encodes an α(1,2)fucosyltransferase (E.C. 2.4.1.69) found in mucosal epithelium of buccal, gastrointestinal, respiratory, breast, and genitourinary tracts of humans and other mammals. Polymorphism of FUT2 is well recognized and is associated with a number of inactivating mutations (Oriol et al., 2000). Approximately 20% of various populations are homozygous for enzyme-inactivating mutations, called "non-secretors," lack α(1,2)fucose residues on secreted mucins. The function(s) of FUT2 remains controversial, but polymorphism of this gene has been proposed to be of medical importance since absence of a functional α(1,2)fucosyltransferase in mucosal tissues is associated with differences in susceptibility to microbial infections, inflammation, and malignancy (D'Adamo and Kelly, 2001; Ishitoya et al., 2002; Lindesmith et al., 2003; Marionneau et al., 2002; Ronchetti et al., 2001).
In the reproductive tract, expression of specific uterine glycans vary during the ovulatory cycle (Carson, 2002; Kimber et al., 2001). Correlated with alterations in the hydrodynamic properties of cervical mucus, cervical mucins show changes in oligosaccharide structures during the ovulatory cycle (Argueso et al., 2002; Yurewicz et al., 1987). In women, non-secretor status is associated with recurrent vulvovaginal candidiasis (Chaim et al., 1997), while absence of a functional FUT2 gene was associated with reduced risk of human immunodeficiency virus type 1 infection (Ali et al., 2000) suggesting that alterations in vaginal mucosal immunity are associated with expression of FUT2, either increased or decreased susceptibility depending on the pathogen.
Despite the large number of fucosylated glycans and proposed biological functions, little is know about the regulation of their expression. Since FUT2 is expressed in both gastrointestinal and reproductive tracts, we investigated the hormonal regulation and cell type of expression of Fut2 in a mutant lacZ reporter mouse expressing the bacterial reporter gene lacZ under the endogenous promoter for Fut2.
Results
Fut2-lacz reporter gene in mutant mice identifies uterine and endocervical glandular cell expression under estrogen regulation
The cell type and regulation of expression of Fut2 was investigated in mice with a targeted mutation of Fut2 that replaced 98% of the Fut2 open reading frame with a nuclear-localizing bacterial reporter gene lacZ (Domino et al., 2001). In mice homozygous for the Fut2-LacZ fusion gene, X-gal staining reveals specific nuclear staining with blue precipitate in glandular and lumenal uterine epithelial cells (Figure 1A). Lumenal cell expression varies in intensity within the uterus during estrus and is almost completely absent in diestrus (Figure 1C). In contrast, glandular epithelium staining is consistently strong between replicate experiments, but also shows decreased expression from estrus to diestrus. Uterine tissues from wild type mice are negative for X-gal staining in estrus and diestrus (Figure 1B and D).
Fig. 1.
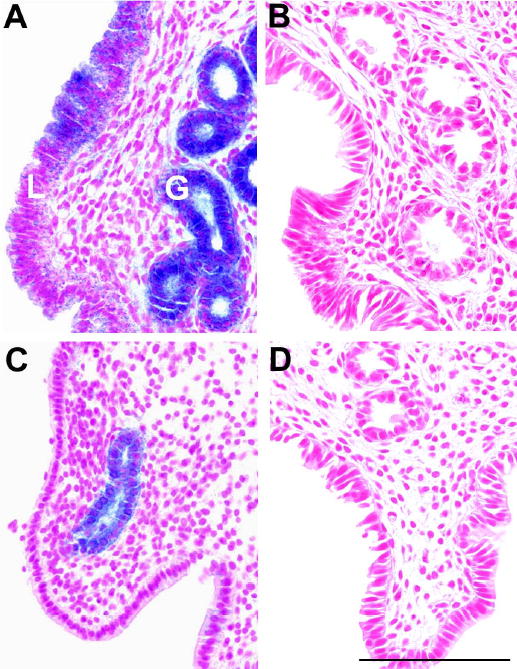
X-gal staining of uterus from Fut2-LacZ mutant mice and wild type controls during the estrous cycle. Top panels show typical microscopic sections of uteri from Fut2-LacZ (A) and wild type (B) mice in estrus. Lumenal epithelium (L) and glandular epithelium (G) are marked in panel A. Bottom panels show microscopic sections of uteri from Fut2-LacZ (C) and wild type (D) mice in diestrus. All panels have similar longitudinal orientations. Three mice were examined at each stage. X-gal staining of glandular cells is consistent between replicates while lumenal staining is weaker and varied across sections. Bar = 100 μm.
The effects of estradiol and progesterone treatments on LacZ expression were examined in ovariectomized Fut2-LacZ mice. Since previous Northern blot data demonstrated Fut2 expression in stomach and colon, X-gal staining was examined in these tissues as well as uterus. X-gal staining of stomach from Fut2-LacZ mice is positive irrespective of hormone replacement (Figure 2A, C, and E). In uterus, X-gal staining is negative with corn oil vehicle (Fig 2B) and estrogen-primed progesterone treatment (Figure 2D). However, with estradiol treatment, X-gal staining is grossly observed from cervix to uterine horns (Figure 2F). Tissues from ovariectomized wild type mice treated with estradiol are negative for X-gal staining (Figure 2G and H). Colon shows a pattern of X-gal staining unresponsive to hormone treatments, identical to stomach (not shown). Vagina, ovary, and oviduct do not show specific X-gal staining (not shown).
Fig. 2.

Whole organ X-gal staining of stomach and uterus of ovariectomized Fut2-LacZ and wild type mice under various hormone treatments. Tissues from four treatment groups were compared: Corn oil vehicle-treated Fut2-LacZ mouse stomach (A) and uterus (B); Estrogen-primed progesterone-treated Fut2-LacZ mouse stomach (C) and uterus (D); Estradiol-treated Fut2-LacZ mouse stomach (E) and uterus (F); Estradiol-treated wild type mouse stomach (G) and uterus (H). Magnification approximately 2X. The experiment was repeated two additional times with similar results.
To investigate the cell type of expression, microscopic examination of uterine and gastrointestinal tissues were performed following whole organ X-gal staining. In uterus, specific X-gal staining of both lumenal and glandular epithelial cells is seen with estradiol treatment (Figure 3A) while estrogen treated wild type mice shows no staining (Figure 3B). Fut2-LacZ mice treated with estrogen-primed progesterone (Figure 3C) or corn oil (Figure 3D) remain negative for X-gal expression.
Fig. 3.
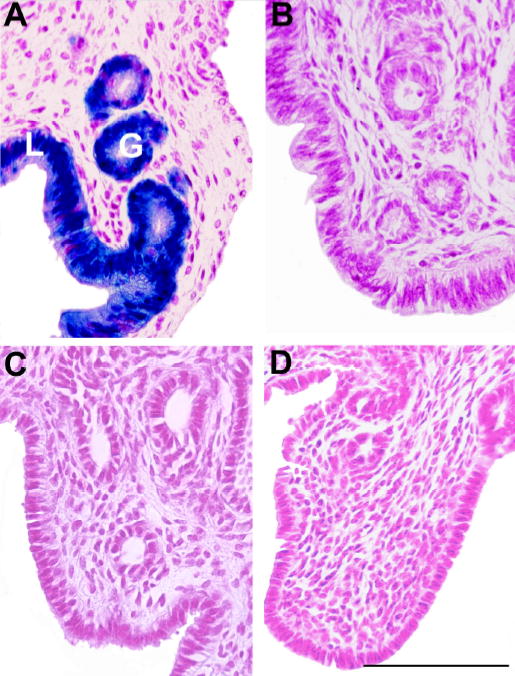
X-gal staining of uterus from Fut2-LacZ mutant mice and wild type controls under various hormone treatments. Microscopic sections of uteri from: A) Estradiol-treated Fut2-LacZ mouse staining glandular (G) and lumenal (L) epithelial cells; B) Progesterone-treated Fut2-LacZ mouse; C) Corn oil vehicle-treated Fut2-LacZ mouse; D) Estradiol-treated wild type mouse. All panels have similar longitudinal orientations. Bar = 100 μm.
In gastrointestinal tissues, microscopic examination of glandular stomach of Fut2-LacZ mice reveals specific lacZ expression in mucous-secreting foveolar pit and chief cells, while parietal cells are negative (Figure 4A). In colon, goblet cells stain densely with X-gal (Figure 4C). Corresponding sections from wild type mouse stomach and colon are negative for X-gal staining (Figure 4B and D).
Fig. 4.
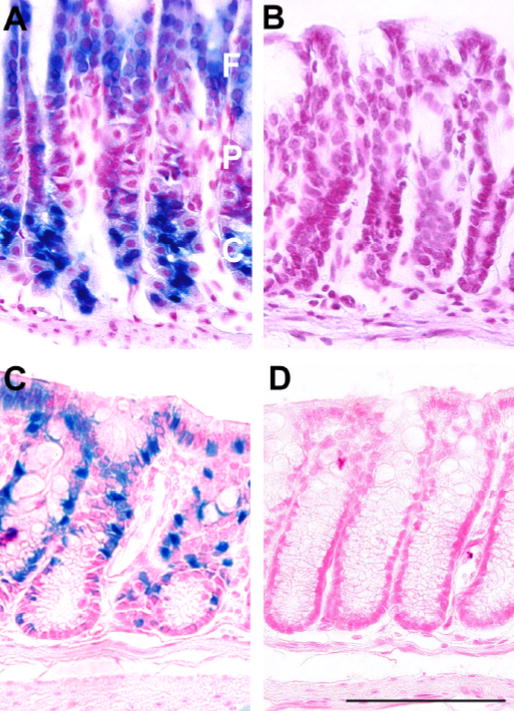
X-gal staining of gastrointestinal tissues from Fut2-LacZ mutant mice and wild type controls. Microscopic sections from: A) Fut2-LacZ glandular stomach showing X-gal staining of mucous-secreting foveolar (F) pit and chief (C) cells, while parietal (P) cells are negative; B) Wild type glandular stomach negative control; C) Fut2-LacZ distal colon showing X-gal staining of goblet cells; D) Wild type distal colon negative control. Stomach samples required cutting into strips to improve penetration of X-gal substrate sufficiently deep to stain chief cells. All panels have similar orientations. Similar results were obtained on three separate occasions. Bar = 100 μm.
Fut2-LacZ expression during pregnancy is restricted to endocervical glands
X-gal staining of uteri from mid-gestation (day 9.5) pregnant Fut2-LacZ mice reveals Fut2-LacZ expression restricted to the endocervix (Figure 5A). Microscopic sections of the uterus at a level between cervix and the most proximal embryo implantation site display minimal expression in glandular epithelium and no expression in lumenal epithelium (Figure 5C). No lacZ expression is seen in implantation sites or placental tissues (not shown), or in tissues from wild type pregnant mice (Figure 5B and D). Identical results are seen in late-gestation mice (day 16.5; not shown).
Fig. 5.
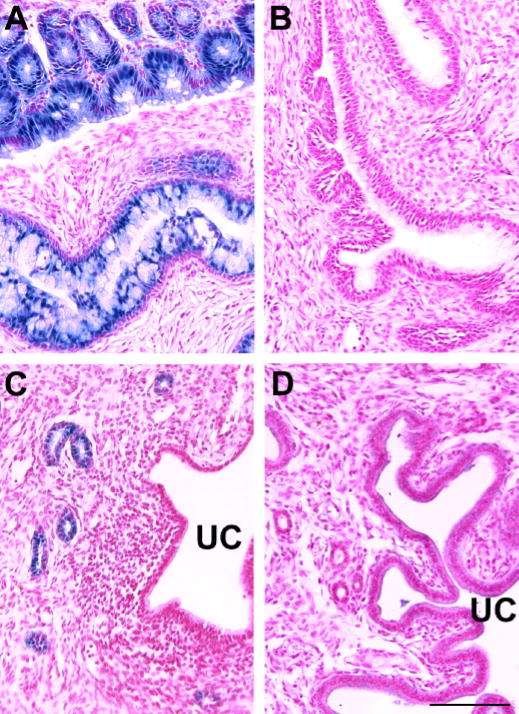
X-gal staining of uterus and endocervix in pregnant Fut2-LacZ mutant mice and pregnant control mice at mid-gestation (Day 9.5). Histologic sections are from: A) Fut2-LacZ endocervix revealing staining of mucus secreting glandular cells; B) X-gal stained wild type endocervix sectioned at a level similar to panel A; C) Fut2-LacZ pregnant uterus showing staining of glandular epithelium, but not lumenal epithelium (uterine cavity marked with UC). Uteri were sectioned at a level between cervix and the most proximal embryo implantation site. Placental implantation sites are negative for X-gal staining; D) X-gal stained wild type pregnant uterus sectioned at a level similar to panel C. The experiment was repeated at 16.5 days gestation, as well as 9.5 days, with identical results in two mice. Bar = 100 μm.
Wild type mice confirm hormonal regulation of Fut2, but not Fut1, steady-state mRNA levels
To confirm the hormonal regulation of the Fut2 locus in wild type mice, Northern blot analyses were performed using probes specific for Fut2 and a second α(1,2)fucosyltransferase expressed in uterus (Fut1) (Domino et al., 2001). For mRNA loading controls, ribosomal protein L32 and cyclophillin-A were tested for suitability since more common standardization genes, such as β-actin and GAPDH, are much more abundant than Fut2 mRNA. Both L32 and cyclophillin-A mRNA display constitutive expression in uterus, stomach and colon (not shown). For further analyses, L32 mRNA was chosen to normalize variations in mRNA loading.
The effects of estradiol and estrogen-primed progesterone treatments on Fut2 and Fut1 steady-state mRNA levels were compared in ovariectomized mice. Analogous to X-gal staining in Fut2-LacZ mice, estradiol treatment of ovariectomized wild type mice increases Fut2 expression 15-fold in uterus with no significant change by estrogen-primed progesterone treatment or estrogen plus progesterone (Figure 6A). Levels of Fut1 were tested since Fut1 was previously reported to be stimulated by estrogen in mouse uterus (Sidhu and Kimber, 1999) (those studies were published prior to the cloning of Fut2 and likely used a probe non-selective between Fut1 and Fut2). Using DNA probes selective for Fut1 vs. Fut2, Fut1 mRNA levels do not significantly vary with hormone replacement (Figure 6B).
Fig. 6.
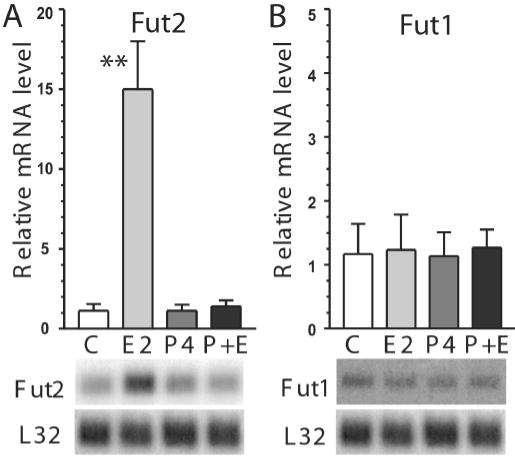
Effects of hormone replacement on steady-state uterine mRNA levels of Fut2 and Fut1 in ovariectomized mice. Using gene-specific probes, Fut2 (A) and Fut1 (B) steady-state mRNA levels are shown in ovariectomized mice treated with three injection regimens: vehicle-only control injections (C), β-estradiol (E2), estrogen-primed progesterone (P4), or estrogen and progesterone (E+P). Results are expressed relative to L32 mRNA and are plotted as ± SD (n=4 mice in each group). Note the lower relative scale for Fut1 compared to Fut2. Statistical comparisons are between vehicle-only control injections vs. other treatments (**p<0.01). PhosphorImager autoradiographs of representative Northern blots are shown below each bar from one mouse in the corresponding treatment group. Each lane corresponds with the bar labeled above. Blots were sequentially hybridized with probes specific for Fut1, followed by Fut2, and then L32, with stripping between hybridizations. The L32 autoradiograph shown under panel B is the same as under panel A since the same blot was probed for both Fut1 and Fut2.
In naturally cycling wild type mice, Fut2 uterine mRNA steady-state levels are highest in proestrus and estrus; elevated 6- to 8-fold compared to diestrus (Figure 7A). Metestrus levels are intermediate between estrus and diestrus. Relative to uterus, Fut2 mRNA levels are more highly expressed in stomach and colon, but do not vary significantly during the estrous cycle (Figure 7B and C).
Fig. 7.
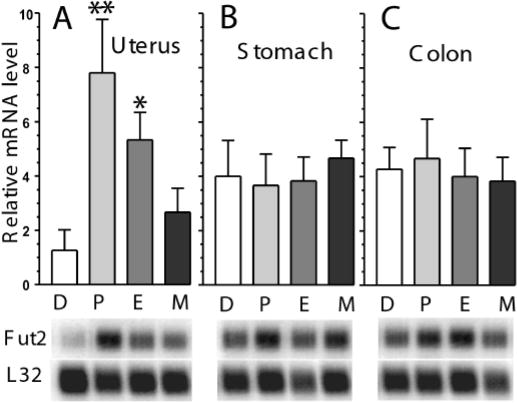
Fut2 mRNA levels during stages of the estrous cycle in reproductive and gastrointestinal tissues. Fut2 steady-state mRNA levels from: A) Uterus; B) Stomach; and C) Colon. Stages of the estrous cycle are diestrus (D), proestrus (P), estrus (E), metestrus 1 (M). Results are expressed relative to L32 mRNA and are plotted as ± SD (n=4 mice in each group). The y-axis in all three panels have the same relative scale. Statistical comparisons are between diestrus vs. other stages (*p<0.05, **p<0.01). PhosphorImager autoradiographs of representative Northern blots are shown below each bar from one mouse in the corresponding stage of the estrous cycle. Each lane corresponds with the bar labeled above. Blots were sequentially hybridized with probes specific for Fut2, followed by L32, with stripping between hybridizations.
Discussion
Estrogens markedly enhance the rate of N-linked glycoprotein synthesis in the uterus by stimulating the glycosylation apparatus including the mannosylphorphoryldolichol synthase pathway (Carson et al., 1990). Among the terminal glycosyltransferases, we demonstrate differential regulation by estradiol of Fut2, but not Fut1, between hormonally responsive and non-responsive tissues. X-gal staining in Fut2-LacZ mice recapitulates the hormonal control of the Fut2 locus in wild type mice in a tissue-specific, cell type-restricted manner. These results represent the first direct demonstration of a glycosyltransferase gene under tissue-specific hormonal regulation in a LacZ reporter mouse model. The genomic elements required for this hormonal regulation are not disrupted by the presence of targeting vector sequences and do not require the Fut2 open reading frame or 3′ region.
Localization of X-gal staining to glandular mucosa of the endocervix throughout pregnancy is an unexpected finding. In studies focusing on a potential role of fucosyltransferase genes in implantation, decreased fucosyltransferase expression was noted in early pregnancy (Sidhu and Kimber, 1999; White and Kimber, 1994). In this study, minimal levels of X-gal staining are seen in the gravid uterus. Prominent X-gal expression, however, is present in the endocervix. Since endocervical glands represent only a small portion of the total uterus upon homogenization, previous Northern blot analyses showing an overall decrease in expression during pregnancy are consistent with the data presented here.
While Fut2-null mice were noted to display normal fertility eliminating an essential role for this gene in implantation, the endocervix was not studied (Domino et al., 2001). Endocervical expression that varies during the estrous cycle and prominent expression during pregnancy suggests a potential protective role against ascending infection analogous to the hypothesized role of glycosyltransferases genes in the gastrointestinal tract (Henry, 2001). In humans, expression of fucosylated blood group antigens in cervical mucus and on the cell surface of vaginal cells vary cyclically in both secretor and non-secretor women (Schaeffer et al., 1994). Since host-pathogen interactions frequently involve cell surface glycans as adhesion molecules, polymorphism of the secretor gene have been proposed to be associated with susceptibility to microbial infections (Varki, 1993). A role for FUT2 in modifying cervical mucus barrier properties is supported by epidemiological studies of woman showing variation with the non-secretor polymorphism in the prevalence of recurrent vulvovaginal candidiasis with C. albicans, and urinary tract infection by some strains of E. coli (Chaim et al., 1997; Stapleton et al., 1995).
A possible mechanism for affect of FUT2 on microbe adhesion is alteration of cervical mucin glycosylation. Multiple mucin forms are expressed in endocervix and MUC5B expression varies with the ovulatory cycle (Gipson et al., 2001). Microbe binding could be altered by shifting the balance of mucin glycosylation between fucosylation and sulfation (Scharfman et al., 1996) similar to cystic fibrosis mice which were found to have elevated Fut2 expression in intestine (Thomsson et al., 2002). Expression of the Fut2 cannot always be considered protective for mucosal immunity, however, since the risk of human immunodeficiency virus type 1 infection was increased in Secretor women relative to Non-secretors (Ali et al., 2000). Other data suggest a positive effect on host-microbe interaction from the loss of FUT2, such as a lower incidence of Helicobacter pylori exposure in Secretor-null individuals (Ikehara et al., 2001). An effect of hormonally regulated genes on sexually transmitted infections, whether protective or deleterious, appears to depend on the specific infecting organism (Brabin, 2002).
This study did not address whether regulation of Fut2 expression by estrogen is direct or indirect. Estrogen receptor-α (ER-α) has been reported to be present in mouse uterus throughout gestation, but with decreasing levels as gestation progressed (Spong et al., 2000). Both epithelial and stromal ER-α were found to be necessary for the production of uterine secretory proteins (Buchanan et al., 1999). Estrogen receptor-β subtype ligands also exerted an effect on some markers of estrogen action (Frasor et al., 2003). To consider potential transcriptional regulation, the 5prime; genomic regions of mouse and human Fut2 were searched for estrogen response consensus sequences. Although annotation of the human and mouse Fut2 genes are currently provisional, mouse Fut2 mRNA (NCBI RefSeq NM_018876) and human FUT2 mRNA (NCBI RefSeq NM_000511) were aligned to the respective genome sequence from the Mouse and Human Genome Consortiums using the UCSC Genome Bioinformatics Human Genome Browser Gateway (Karolchik et al., 2003). The corresponding genomic regions (6000 bp) 5prime; to exon 1 from mouse (chr7:34307353-34313352 minus strand, February 2003 freeze) and human (chr19:53853281 to 53859280 plus strand, April 2003 freeze) were scanned with the MacInspector program V2.2 using TRANFAC 4.0 matrixes (Wingender et al., 2000). No consensus estrogen response elements were found, although multiple weak partial matches to the half palindrome site GGTCA are present.
Estrogen receptor-mediated transcriptional regulation may, however, proceed through non-classical ER-α/Sp1 binding where estrogen binds its receptor followed by binding to Sp1 transcription factors which then directs the complex to a binding motif in the 5prime; region (Khan et al., 2003). Multiple potential Sp1 binding motifs were present in the Fut2 5prime; region along with dozens of potential transcription factor sites. Alternatively to transcriptional control, the 3prime; untranslated region of human Fut2 mRNA may form a large stem-and-loop structure (1.2 kb) potentially regulating the stability and level of the Fut2 transcript (Koda et al., 1997). Study of cells from Fut2-LacZ tissues would benefit from the relative easy of assay for lacZ and may help elucidate molecular mechanism of Fut2 hormonal regulation. The Fut2-LacZ mouse may also serve as a model of the human non-secretor for testing the importance of cell surface α(1,2)fucosylated glycans in host-microbe interactions.
Materials and methods
Experimental animals and hormone treatments
Animal protocols were approved by the University of Michigan Committee on Use and Care of Animals. Groups of twelve mice, wild type strain 129X1/SvJ (The Jackson Laboratory, Bar Harbor, ME; stock no. 000691) or Fut2-LacZ mutant mice (Domino et al., 2001) (MGI accession ID 2183220, allele symbol Fut2tm1Sdo), age 8-9 weeks old, were followed by examination of vaginal cytology to determine the stage of estrous cycle (Nelson et al., 1982). For hormone treatments, groups of eight mice were ovariectomized as described (Nagy et al., 2002). Two weeks later, all mice were injected i.p. daily for 3 days with a priming dose of 0.1 μg β-estradiol (Sigma, St. Louis, MO; catalog no. E-8515). After a 2 day wait, mice were injected daily for 4 days with either 0.1 μg β-estradiol, 500 μg progesterone (Sigma, catalog no. P-0130), estrogen and progesterone, or vehicle-only corn oil (White and Kimber, 1994). For experiments requiring pregnant mice, females were housed overnight with male studs and vaginal plugs checked to determine day 0.5 of pregnancy. Pregnancy was confirmed by the presence of multiple gestational sacs when mice were euthanized. The targeting vector and generation of Fut2-LacZ mutant mice was previously described (Domino et al., 2001). In this study, the Fut2-LacZ homozygous mutant mice were on a hybrid 129X1/SvJ and C57BL/6J background generation F3-F5.
X-gal staining
Control experiments for X-gal staining using wild type strain 129X1/SvJ and strain C57BL/6J (stock no. 000664) showed no background staining in the tissues tested. The experimental mutant mice, Fut2-LacZ, were on a hybrid 129X1/SvJ and C57BL/6J background. To prepare tissues for X-gal staining, 8–9 week old Fut2-LacZ and wild type mice were terminally perfused with ice cold PBS followed by freshly thawed 4% paraformaldehyde (Turkay et al., 1999). Tissues were excised, rinsed in PBS, then partially fixed in ice cold 4% paraformaldyde for 20 min on ice with occasional shaking. Tissues were washed with 0.1 M sodium phosphate pH 7.3 three times and stained with X-gal overnight at 37 C as described (Saunders, 2002). To preserve the blue X-gal precipitant, tissues were post-fixed in 10% formalin and 0.2% glutaraldehyde overnight, then processed for paraffin sectioning and nuclear fast red staining avoiding all organic solvents (Histoserv, Inc., Germantown, MD). Whole organ pictures were photographed with a 35mm macrolens.
Northern blot analysis
Total RNA was isolated by the guanidine isothiocyanate/phenol method (TRIzol; GibcoBRL, Grand Island, NY) followed by isolation of polyadenylated RNA by Oligotex midi mRNA columns (Qiagen, Valencia, CA). Colon samples were taken from the mid portion of the large intestine of each animal. Independently isolated samples from individual mice containing 5 μg of polyadenylated RNA were run on formaldehyde agarose gels, transferred to nylon, and hybridized with 32P-dCTP random primed DNA probes specific for Fut1 and Fut2 exactly as previously described (Domino et al., 2001). Blots were stripped and hybridized with probes for either ribosomal protein L32 (113-bp probe purified from a Hind III digestion of mouse L32 template; BD Biosciences Pharmingen, San Diego, CA; no.45181P) or cyclophillin-A (Ambion, Austin, TX; mouse DECAprobe template no.7375). A PhosphorImager Model SP was used to visualize and quantify Northern blot radioactivity (Amersham Biosciences, Piscataway, NJ). Northern blot data were normalized to the level of L32 in each lane. One-way ANOVA with Dunnett’s Multiple Comparison Test was performed using GraphPad Prism version 3.00 for Windows (GraphPad Software, San Diego, CA).
Acknowledgments
The authors thank personnel of University of Michigan IPOX histology core and Histoserv, Inc., for histology processing. We thank the University of Michigan Transgenic Animal Model core for assistance with ovariectomies and management of the Fut2-LacZ mice. This work was supported in part by NIH grants HD01195, CA93856, and a University of Michigan Rackham Grant (to S.E.D.). E.A.H is a recipient of a University of Michigan Biomedical Research Council postdoctoral fellowship award.
References
- Ali S, Niang MA, N'Doye I, Critchlow CW, Hawes SE, Hill AV, Kiviat NB. Secretor polymorphism and human immunodeficiency virus infection in Senegalese women. J Infect Dis. 2000;181:737–739. doi: 10.1086/315234. [DOI] [PubMed] [Google Scholar]
- Argueso P, Spurr-Michaud S, Tisdale A, Gipson IK. Variation in the amount of T antigen and N-acetyllactosamine oligosaccharides in human cervical mucus secretions with the menstrual cycle. J Clin Endocrinol Metab. 2002;87:5641–5648. doi: 10.1210/jc.2002-020766. [DOI] [PubMed] [Google Scholar]
- Becker DJ, Lowe JB. Fucose: biosynthesis and biological function in mammals. Glycobiology. 2003;13:41R–53R. doi: 10.1093/glycob/cwg054. [DOI] [PubMed] [Google Scholar]
- Brabin L. Interactions of the female hormonal environment, susceptibility to viral infections, and disease progression. AIDS Patient Care & Stds. 2002;16:211–221. doi: 10.1089/10872910252972267. [DOI] [PubMed] [Google Scholar]
- Breton C, Oriol R, Imberty A. Conserved structural features in eukaryotic and prokaryotic fucosyltransferases. Glycobiology. 1998;8:87–94. doi: 10.1093/glycob/8.1.87. [DOI] [PubMed] [Google Scholar]
- Buchanan DL, Setiawan T, Lubahn DB, Taylor JA, Kurita T, Cunha GR, Cooke PS. Tissue compartment-specific estrogen receptor-alpha participation in the mouse uterine epithelial secretory response. Endocrinology. 1999;140:484–491. doi: 10.1210/endo.140.1.6448. [DOI] [PubMed] [Google Scholar]
- Carson DD. The glycobiology of implantation. Front Biosci. 2002;7:d1535–1544. doi: 10.2741/A858. [DOI] [PubMed] [Google Scholar]
- Carson DD, Farrar JD, Laidlaw J, Wright DA. Selective activation of the N-glycosylation apparatus in uteri by estrogen. J Biol Chem. 1990;265:2947–2955. [PubMed] [Google Scholar]
- Chaim W, Foxman B, Sobel JD. Association of recurrent vaginal candidiasis and secretory ABO and Lewis phenotype. J Infect Dis. 1997;176:828–830. doi: 10.1086/517314. [DOI] [PubMed] [Google Scholar]
- D'Adamo PJ, Kelly GS. Metabolic and immunologic consequences of ABH secretor and Lewis subtype status. Alternative Med Rev. 2001;6:390–405. [PubMed] [Google Scholar]
- Domino SE, Zhang L, Gillespie PJ, Saunders TL, Lowe JB. Deficiency of reproductive tract alpha(1,2)fucosylated glycans and normal fertility in mice with targeted deletions of the FUT1 or FUT2 alpha(1,2)fucosyltransferase locus. Mol Cell Biol. 2001;21:8336–8345. doi: 10.1128/MCB.21.24.8336-8345.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domino SE, Zhang L, Lowe JB. Molecular cloning, genomic mapping, and expression of two secretor blood group alpha (1,2)fucosyltransferase genes differentially regulated in mouse uterine epithelium and gastrointestinal tract. J Biol Chem. 2001;276:23748–23756. doi: 10.1074/jbc.M100735200. [DOI] [PubMed] [Google Scholar]
- Frasor J, Barnett DH, Danes JM, Hess R, Parlow AF, Katzenellenbogen BS. Response-specific and ligand dose-dependent modulation of estrogen receptor (ER) alpha activity by ERbeta in the uterus. Endocrinology. 2003;144:3159–3166. doi: 10.1210/en.2002-0143. [DOI] [PubMed] [Google Scholar]
- Gipson IK, Moccia R, Spurr-Michaud S, Argueso P, Gargiulo AR, Hill JA, 3rd, Offner GD, Keutmann HT. The Amount of MUC5B mucin in cervical mucus peaks at midcycle. J Clin Endocrinol Metab. 2001;86:594–600. doi: 10.1210/jcem.86.2.7174. [DOI] [PubMed] [Google Scholar]
- Henry SM. Molecular diversity in the biosynthesis of GI tract glycoconjugates. A blood-group-related chart of microorganism receptors. Transfus Clin Biol. 2001;8:226–230. doi: 10.1016/s1246-7820(01)00112-4. [DOI] [PubMed] [Google Scholar]
- Ikehara Y, Nishihara S, Yasutomi H, Kitamura T, Matsuo K, Shimizu N, Inada K, Kodera Y, Yamamura Y, Narimatsu H, Hamajima N, Tatematsu M. Polymorphisms of two fucosyltransferase genes (Lewis and Secretor genes) involving type I Lewis antigens are associated with the presence of anti-Helicobacter pylori IgG antibody. Cancer Epidemiol Biomarkers Prev. 2001;10:971–977. [PubMed] [Google Scholar]
- Ishitoya S, Yamamoto S, Mitsumori K, Ogawa O, Terai A. Non-secretor status is associated with female acute uncomplicated pyelonephritis. BJU Int. 2002;89:851–854. doi: 10.1046/j.1464-410x.2002.02782.x. [DOI] [PubMed] [Google Scholar]
- Karolchik D, Baertsch R, Diekhans M, Furey TS, Hinrichs A, Lu YT, Roskin KM, Schwartz M, Sugnet CW, Thomas DJ, Weber RJ, Haussler D, Kent WJ, University of California Santa C. The UCSC Genome Browser Database. Nucleic Acids Res. 2003;31:51–54. doi: 10.1093/nar/gkg129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan S, Abdelrahim M, Samudio I, Safe S. Estrogen receptor/Sp1 complexes are required for induction of cad gene expression by 17beta-estradiol in breast cancer cells. Endocrinology. 2003;144:2325–2335. doi: 10.1210/en.2002-0149. [DOI] [PubMed] [Google Scholar]
- Kimber SJ, Stones RE, Sidhu SS. Glycosylation changes during differentiation of the murine uterine epithelium. Biochem Soc Trans. 2001;29:156–162. doi: 10.1042/0300-5127:0290156. [DOI] [PubMed] [Google Scholar]
- Koda Y, Soejima M, Wang B, Kimura H. Structure and expression of the gene encoding secretor-type galactoside 2-alpha-L-fucosyltransferase (FUT2) Eur J Biochem. 1997;246:750–755. doi: 10.1111/j.1432-1033.1997.t01-1-00750.x. [DOI] [PubMed] [Google Scholar]
- Lindesmith L, Moe C, Marionneau S, Ruvoen N, Jiang X, Lindblad L, Stewart P, LePendu J, Baric R. Human susceptibility and resistance to Norwalk virus infection. Nat Med. 2003;9:548–553. doi: 10.1038/nm860. [DOI] [PubMed] [Google Scholar]
- Marionneau S, Ruvoen N, Le Moullac-Vaidye B, Clement M, Cailleau-Thomas A, Ruiz-Palacois G, Huang P, Jiang X, Le Pendu J. Norwalk virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology. 2002;122:1967–1977. doi: 10.1053/gast.2002.33661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy, A., Behringer, R., Gertsenstein, M. and Vintersten, K. (2002) Manipulating the mouse embryo: a laboratory manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
- Nelson JF, Felicio LS, Randall PK, Sims C, Finch CE. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 1982;27:327–339. doi: 10.1095/biolreprod27.2.327. [DOI] [PubMed] [Google Scholar]
- Oriol R, Candelier JJ, Mollicone R. Molecular genetics of H. Vox Sang. 2000;78:105–108. [PubMed] [Google Scholar]
- Ronchetti F, Villa MP, Ronchetti R, Bonci E, Latini L, Pascone R, Bottini N, Gloria-Bottini F. ABO/Secretor genetic complex and susceptibility to asthma in childhood. Eur Respir J. 2001;17:1236–1238. doi: 10.1183/09031936.01.99109101. [DOI] [PubMed] [Google Scholar]
- Saunders, T.L. (2002) Reporter molecules in genetically engineered mice. In Hofker, M.H. and van Deursen, J. (eds), Methods in Molecular Biology: Transgenic Mouse Methods and Protocols Humana Press, Totowa, New Jersey, pp. 125–143. [DOI] [PubMed]
- Schaeffer AJ, Navas EL, Venegas MF, Anderson BE, Kanerva C, Chmiel JS, Duncan JL. Variation of blood group antigen expression on vaginal cells and mucus in secretor and nonsecretor women. J Urol. 1994;152:859–864. doi: 10.1016/s0022-5347(17)32591-0. [DOI] [PubMed] [Google Scholar]
- Scharfman A, Van Brussel E, Houdret N, Lamblin G, Roussel P. Interactions between glycoconjugates from human respiratory airways and Pseudomonas aeruginosa. Am J Respir Crit Care Med. 1996;154:S163–169. doi: 10.1164/ajrccm/154.4_Pt_2.S163. [DOI] [PubMed] [Google Scholar]
- Shi S, Stanley P. Protein O-fucosyltransferase 1 is an essential component of Notch signaling pathways. Proc Natl Acad Sci USA. 2003;100:5234–5239. doi: 10.1073/pnas.0831126100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidhu SS, Kimber SJ. Hormonal control of H-type alpha(1-2)fucosyltransferase messenger ribonucleic acid in the mouse uterus. Biol Reprod. 1999;60:147–157. doi: 10.1095/biolreprod60.1.147. [DOI] [PubMed] [Google Scholar]
- Smith PL, Myers JT, Rogers CE, Zhou L, Petryniak B, Becker DJ, Homeister JW, Lowe JB. Conditional control of selectin ligand expression and global fucosylation events in mice with a targeted mutation at the FX locus. J Cell Biol. 2002;158:801–815. doi: 10.1083/jcb.200203125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spong CY, McCune SK, Sternberg EM, Gustafsson JA. Maternal estrogen receptor-beta expression during mouse gestation. Am J Reprod Immunol. 2000;44:249–252. doi: 10.1111/j.8755-8920.2000.440410.x. [DOI] [PubMed] [Google Scholar]
- Stapleton A, Hooton TM, Fennell C, Roberts PL, Stamm WE. Effect of secretor status on vaginal and rectal colonization with fimbriated Escherichia coli in women with and without recurrent urinary tract infection. J Infect Dis. 1995;171:717–720. doi: 10.1093/infdis/171.3.717. [DOI] [PubMed] [Google Scholar]
- Thomsson KA, Hinojosa-Kurtzberg M, Axelsson KA, Domino SE, Lowe JB, Gendler SJ, Hansson GC. Intestinal mucins from cystic fibrosis mice show increased fucosylation due to an induced Fucalpha1-2 glycosyltransferase. Biochem J. 2002;367:609–616. doi: 10.1042/BJ20020371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turkay A, Saunders T, Kurachi K. Intrauterine gene transfer: gestational stage-specific gene delivery in mice. Gene Ther. 1999;6:1685–1694. doi: 10.1038/sj.gt.3301007. [DOI] [PubMed] [Google Scholar]
- Varki A. Biological roles of oligosaccharides: all of the theories are correct. Glycobiology. 1993;3:97–130. doi: 10.1093/glycob/3.2.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White S, Kimber SJ. Changes in alpha (1–2)-fucosyltransferase activity in the murine endometrial epithelium during the estrous cycle, early pregnancy, and after ovariectomy and hormone replacement. Biol Reprod. 1994;50:73–81. doi: 10.1095/biolreprod50.1.73. [DOI] [PubMed] [Google Scholar]
- Wingender E, Chen X, Hehl R, Karas H, Liebich I, Matys V, Meinhardt T, Pruss M, Reuter I, Schacherer F. TRANSFAC: an integrated system for gene expression regulation. Nucleic Acids Res. 2000;28:316–319. doi: 10.1093/nar/28.1.316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yurewicz EC, Matsuura F, Moghissi KS. Structural studies of sialylated oligosaccharides of human midcycle cervical mucin. J Biol Chem. 1987;262:4733–4739. [PubMed] [Google Scholar]