

Abstract
Background
cAMP-induced Ca2+-influx in Dictyostelium is controlled by at least two non-mitochondrial Ca2+-stores: acidic stores and the endoplasmic reticulum (ER). The acidic stores may comprise the contractile vacuole network (CV), the endosomal compartment and acidocalcisomes. Here the role of CV in respect to function as a potential Ca2+-store was investigated.
Results
Dajumin-GFP labeled contractile vacuoles were purified 7-fold by anti-GFP-antibodies in a magnetic field. The purified CV were shown for the first time to accumulate and release Ca2+. Release of Ca2+ was elicited by arachidonic acid or the calmodulin antagonist W7, the latter due to inhibition of the pump. The characteristics of Ca2+-transport and Ca2+-release of CV were compared to similarly purified vesicles of the ER labeled by calnexin-GFP. Since the CV proved to be a highly efficient Ca2+-compartment we wanted to know whether or not it takes part in cAMP-induced Ca2+-influx. We made use of the LvsA--mutant expected to display reduced Ca2+-transport due to loss of calmodulin. We found a severe reduction of cAMP-induced Ca2+-influx into whole cells.
Conclusion
The contractile vacuoles in Dictyostelium represent a highly efficient acidic Ca2+-store that is required for cAMP-induced Ca2+-influx.
Background
The contractile vacuole (CV) network of Dictyostelium consists of tubes and bladders. It transiently fuses with the plasma membrane to expel water and ions and thereby serves as an efficient osmoregulatory organelle [1,2]. The CV-system is also assumed to be involved in Ca2+-transport since it contains a PMCA-type Ca2+-ATPase (PAT1), calmodulin [3] and a vacuolar proton pump that establishes a proton gradient for Ca2+-transport [4]. PAT1 is localized to the CV and the plasma membrane and is upregulated under conditions of Ca2+-stress [5]. Downregulation of PAT1 by antisense RNA reduced vesicular Ca2+-uptake.
We are interested in the characterization of Ca2+-stores that are involved in cAMP-induced Ca2+-influx. Previously, it has been shown that acidic Ca2+-stores and an IP3-sensitive store participate in this regulation [6-11]. Acidic means that the stores are equipped with a V-type H+-ATPase. Acidic vesicular Ca2+-stores in Dictyostelium comprise the CV-system, endosomes and acidocalcisomes [12,13]. In the present study we focus on the contribution of the CV-system to intracellular Ca2+ regulation.
It has been shown previously that GFP-tagged dajumin labels the entire CV whereas the endosomal compartments are devoid of the label [14]. By contrast, drainin, a peripheral membrane protein involved in discharge of the bladder, is found only on the bladder [15]. We used dajumin-GFP expressing cells to obtain a fraction enriched in CV membranes and used this fraction to measure Ca2+-transport. Ca2+-transport activity increased with enhanced purity of the CV. We also employed a LvsA minus strain which lacks the gene for the protein large volume sphereA (lvsA). Besides its involvement in cytokinesis [16] it is known that the LvsA-protein is localized to the CV. This association with the CV occurs only during the discharge phase of the vacuole. In the lvsA mutant calmodulin was lost from the CV-membranes and the CV became disorganized, unable to discharge its contents [17]. We found that vesicular Ca2+-transport in the lvsA-mutant was diminished and that cAMP-induced Ca2+-influx was drastically reduced, indicating that functional CV are absolutely required for the cAMP-dependent Ca2+-influx.
Results
Distribution of CV in vesicular fractions
We used differentiated cells 4 to 5 hrs after starvation for preparation of Ca2+-transporting vesicles because cAMP-induced Ca2+-influx is present at that time and endosomal content is low (see below). Cells labeled with dajumin-GFP as a marker for the CV-system or with calnexin-GFP cells as a marker for the endoplasmic reticulum (ER) are shown in Figure 1. Dajumin-GFP allows to visualize the dynamics of the bladder by formation of irregular ventricles and ducts (A). The ER is prominently labeled in the perinuclear region and close to the plasma membrane (B). The cells were lysed by passage through nuclepore filters. Vesicular fractions were separated by differential centrifugation and assayed for Ca2+-transporting activity. The dajumin-GFP label was detected in vesicular fractions with the majority being present in the fast sedimenting fraction P0 (Table 1). By contrast, most of the ER occurred in P1, whereas the lightest fraction P2 contained only a quarter of both organelles (Table 1). Plasma membranes, as shown previously, sedimented in P1 [18]. Ca2+-transport activity was strikingly concentrated in P0. The presence of endosomes was measured with RITC-dextran. In two independent experiments 38 ± 6% of the label was associated with P0, 62 ± 6% with P1 and none was present in P2. However, the amount of endosomes of 4 hour starved cells was low and barely detectable. This result is expected since the cells develop in the absence of nutrients. If endosomes accumulate Ca2+ their contribution to Ca2+-transport was therefore considered to be insignificant under the present experimental conditions.
Figure 1.
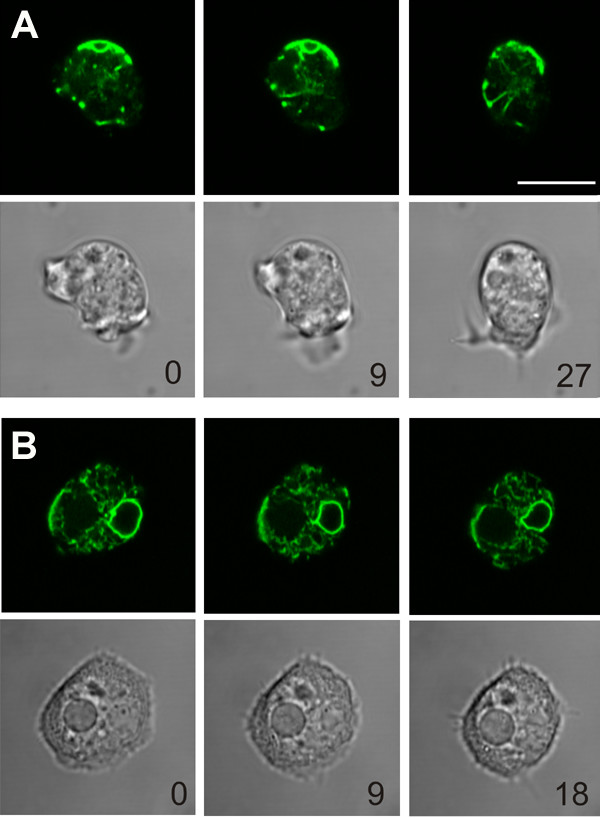
Dictyostelium wild-type Ax2 cells expressing dajumin-GFP or calnexin-GFP. (A): The contractile vacuole system of a Dictyostelium cell is visualized by expression of dajumin-GFP using live cell confocal microscopy (upper panels). Dynamics of the bladder is indicated by formation of irregular ventricles and ducts. Lower panels show the corresponding bright field images. (B): A Dictyostelium cell expressing calnexin-GFP to visualize the endoplasmic reticulum is shown in the upper panels. Note that the perinuclear region is intensively labeled as observed previously for calnexin-GFP and antibody-stained preparations, whereas a vacuole in the left part of the cell is devoid of any label. Time of depicted frames is indicated in seconds. Bar corresponds to 10 μm.
Table 1.
Characterization of crude vesicular fractions. Vesicles were obtained by differential centrifugation of lysed cells as described in Methods. The amount (percent) of CV (± S.D.) in each fraction was determined according to the dajumin-GFP label present, while that of the ER by the presence of calnexin-GFP label. Total Ca2+-transport activity obtained from 4 × 108 cells is shown for each fraction. Data are means ± SD. The number of independent experiments is given in brackets.
| Fraction | Sedimentation | CV | ER | Ca2+-uptake | |
| (g) | % | % | nmol | % | |
| P0 | 3.800 | 42.4 ± 4.0 (4) | 33.5 ± 3.7 (3) | 205 ± 42 (3) | 67 |
| P1 | 12.000 | 33.6 ± 7.2 (4) | 43.0 ± 2.8 (3) | 76 ± 24 (3) | 25 |
| P2 | 40.000 | 24.0 ± 10.6 (4) | 23.5 ± 1.3 (3) | 25 ± 6 (3) | 8 |
Purification of CV with magnetic antibody-beads
Since P0 was the richest source of CV we incubated P0 isolated from dajumin-GFP labeled cells with anti-GFP-magnetic beads and loaded the mixture onto a column. During loading and washing the beads were retained on the column by applying a magnetic field. Elution was performed in its absence. As shown in Table 2 almost all of the dajumin-label was present in the eluate (P0E) as well as most of Ca2+-uptake. The flow through (P0F) still contained some Ca2+-transport activity the percentage of which, however, exceeded only slightly the remaining dajumin-GFP-label.
Table 2.
Purification of CV and ER fractions by antibody. Dajumin-GFP labeled CV present in P0 were bound to anti-GFP-microbeads and separated from the unbound vesicles in a magnetic field as detailed in Methods. P0F represents the flow through, P0E is the eluate containing CV. Calnexin-GFP labeled ER present in P1E was purified in the same way. P1F: flow through, P1E: eluate containing ER. As described in Methods, the measured values of Ca2+-uptake were corrected for the same initial [Ca2+]ev. Data are means ± SD. The number of independent experiments is given in brackets.
| Fraction | GFP-label % | Ca2+-uptake % | Specific activity of Ca2+-ATPase (%) |
| P0F (Dajumin) | 12.4 ± 3.9 (3) | 15.6 ± 9.7 (3) | 18 ± 5.4 (3) |
| P0E (Dajumin) | 87.6 ± 3.9 (3) | 84.4 ± 9.7 (3) | 109 ± 9.3 (3) |
| P1F (Calnexin) | 13.8 ± 4.2 (4) | 13.4 ± 8.3 (4) | 8.1 ± 6.3 (4) |
| P1E (Calnexin) | 86.2 ± 4.2 (4) | 86.6 ± 8.3 (4) | 87 ± 66 (4) |
The selectivity of the GFP-antibody beads for GFP was tested by assaying P0 from unlabeled Ax2-cells, the parent strain of dajumin-GFP expressing cells, for binding to the anti-GFP-beads. In three independent experiments 54 ± 31 μg (n = 6) protein was associated with P0E in the controls whereas 192 ± 60 μg (n = 6) protein was present in P0E from dajumin-GFP labeled cells. This result shows that the GFP-antibody specifically targets the GFP-tagged protein and that 28% of unspecific binding does occur.
In order to avoid contamination of P0 and subsequent fractions with plasma membranes due to possible association of plasma membranes and CV by cortical actin filaments we preincubated the cells (all experiments of Table 2) with latrunculin B. Latrunculin B, a unique marine toxin, inhibits actin polymerization and disrupts microfilament organization [19]. In the presence of latrunculin B the cells rounded up and were completely immotile before they were pressed through the filters. The characteristics of Ca2+-transport, its inhibition by drugs as well as Ca2+-release in P0E were in the same range as in preparations performed without latrunculin B treatment. Latrunculin B treatment reduced the protein content of P0E by a factor of 1.26 but there was no statistically significant difference in the specific activity of the Ca2+-ATPase.
A specific marker for the CV as compared to the plasma membrane is the V-type H+-ATPase. Antibodies to the 37/42 kDa subunit revealed that the proton pump was markedly enriched in P0E (Fig. 2). A mitochondrial contribution to Ca2+-fluxes was prevented by inclusion of mitochondrial inhibitors in all vesicular Ca2+-transport measurements.
Figure 2.

Enrichment of V-type H+-ATPase in P0E. Western blot analysis with antibodies to V-type H+-ATPase was carried out as described in Methods using fractions of cells expressing dajumin-GFP. 7 μg of protein per lane of each fraction were applied.
Purification of ER with magnetic antibody-beads
Since P1 was the richest source of ER (Table 1) we incubated P1 isolated from calnexin-GFP labeled cells with anti-GFP-magnetic beads and separated ER-vesicles from other vesicles by a magnetic field. As shown in Table 2 almost all of the calnexin-GFP-label was found in the eluate (P1E) as well as most of Ca2+-uptake activity. The flow through (P1F) also contained some Ca2+-transport activity.
The results of Table 2 thus show that by using the anti-GFP-antibody beads a 6-7-fold enrichment of either CV or ER can be obtained. Furthermore, both fractions enriched either in CV or ER vesicles, displayed a similar enhancement of Ca2+-transport. This result demonstrates that vesicles of the CV of Dictyostelium are functional in Ca2+-homeostasis.
Properties of CV and ER fractions with respect to Ca2+-regulation
Since both purifications had given a strong enrichment of the GFP-label as well as in Ca2+-transport in the eluate (84–87%) and since in control experiments 28% unlabeled protein was associated with the eluate, we decided to test for the properties of CV and ER with respect to Ca2+-transport and Ca2+-release. As shown in Figure 3 P0E, the eluate of dajumin-GFP-labeled cells containing CV, transported Ca2+ efficiently. Note, that the slope is fairly steep and uptake of Ca2+ is settled in about 8 min. In three independent experiments we found that starting at an extravesicular Ca2+-concentration ([Ca]2+ev) of 0.53 ± 0.02 μM uptake ceased to 0.10 ± 0.05 μM [Ca]2+ev in the presence of 32 ± 12 μg protein. In agreement with previous results obtained for acidic Ca2+-stores the calmodulin antagonist W7 as well as arachidonic acid (AA) caused Ca2+-release in P0E. The response to W7 (30–100 μM) amounted to 21.4 ± 10.2 nmol Ca2+/mg protein (n = 4) and to AA (10–20 μM) 23.9 ± 7.2 nmol Ca2+/mg protein (n = 4).
Figure 3.

Ca2+-uptake and Ca2+-release in P0E. P0E was obtainedby GFP-antibody purification. Ca2+-uptake was induced by 0.4 mM ATP and measured with Fluo-3 as described in Methods. A Ca2+-calibration pulse was followed by further Ca2+-uptake. W-7 and AA induced Ca2+-release, 4-BrA23187 elicited further Ca2+-release. One out of three independent experiments is shown.
In Figure 4 Ca2+-uptake by P1E, the eluate of calnexin-GFP-labeled cells containing ER, is shown. In contrast to P0E P1E was less responsive to AA. In three out of four experiments no Ca2+-release was measured. The one that responded to AA had a lower purification factor with respect to GFP fluorescence compared with the other three, 3.8 versus 7.8 ± 0.96 (n = 3). IP3, most of the times, was inefficient to elicit Ca2+-release. In general, Ca2+-uptake was slower in P1E as compared to P0E starting at 0.56 ± 0.15 μM [Ca]2+ev and ceased to extravesicular Ca2+-concentrations of about 0.36 ± 0.08 μM in the presence of 27 ± 12 μg protein (n = 4), indicating that the CV is more relevant in maintaining a basal cytosolic Ca2+-concentration of 50 to 60 nM [10] than the ER.
Figure 4.

Ca2+-uptake and Ca2+-release in P1E. P1E was obtainedby GFP-antibody-purification. Ca2+-measurements were performed as described in the legend to Fig. 1. One out of three independent experiments is shown.
In addition, we tested for inhibition of Ca2+-uptake. Table 3 shows that concanamycin A (CMA), a powerful inhibitor of V-type H+-ATPases [20,21] inhibited P0E as expected for a compartment that uses a proton gradient for Ca2+-transport and a Ca2+-transport ATPase PAT1. Under our experimental conditions BHQ, an inhibitor of Ca2+-transport ATPases, inhibited Ca2+-uptake only weakly despite its reported property to inhibit cAMP-induced Ca2+-influx in Dictyostelium [22,23]. BHQ was more efficient in inhibiting Ca2+-uptake into the ER-fraction. The latter was also sensitive to thapsigargin (Tg) a blocker of SERCA-type Ca2+-ATPases. In both fractions, P0E and P1E, Ca2+-uptake was inhibited by low concentrations (30–60 μM) of Sr2+ by about 50%.
Table 3.
Inhibition of Ca2+-uptake activity. The purified CV-fraction P0E and the purified ER-fraction P1E were tested for sensitivity of Ca2+-uptake to Tg (22 μM), BHQ (200 μM), CMA (40 μM) and Sr2+ (30–60 μM) as indicated. nd: not determined. Data are means ± SD. The number of independent experiments is given in brackets.
| Fraction | Tg % | BHQ % | CMA % | Sr2+ % |
| P0E (CV) | nd | 25 ± 13 (3) | 57 ± 6 (4) | 52 ± 13 (3) |
| P1E (ER) | 45 ± 23 (4) | 72 ± 4 (3) | nd | 43 ± 6 (4) |
Is the CV involved in cAMP-induced Ca2+-influx?
Given our results that the CV is a Ca2+-store that transports Ca2+ most efficiently at low Ca2+-concentrations we wanted to know whether the store is also involved in receptor-mediated Ca2+-flux. Although it was previously shown that NBD-Cl and CMA inhibited the cAMP mediated Ca2+-influx [7,22] these results only proved that acidic stores as a whole were involved. We made use of the LvsA--mutant whose CV-function is abolished with respect to fusion of CV with the plasma membrane and is disturbed by a loss of calmodulin. Previously, we have shown that the calmodulin antagonist W7 inhibits Ca2+-transport of acidic Ca2+-stores [24]. A localized loss of calmodulin confined to the CV would have the same consequences. Therefore, it was interesting to know whether or not Ca2+-influx was also disturbed in the mutant. As shown in Table 4 there was no receptor-mediated Ca2+-influx in the LvsA minus strain at physiological cAMP-concentrations (0.1–10 μM) in contrast to the parent strain DH1. Only higher concentrations of cAMP yielded Ca2+-influx that was reduced as compared to DH1 (Table 4).
Table 4.
Stimulus-induced Ca2+-influx in LvsA-and DH1. Ca2+-influx was determined with a Ca2+-sensitive electrode before and after addition of cAMP or AA at an extracellular Ca2+-concentration of 2–3 μM. Data are means ± SD. The number of independent experiments is given in brackets.
| Ca2+-influx pmol/107 cells | ||
| Stimulus (μM) | LvsA- | DH1 |
| cAMP 0.1 | < 0.1 (7) | nd |
| 1 | < 0.1 (7) | 100 ± 59 (3) |
| 10 | 1 ± 5 (9) | 203 ± 107 (3) |
| 20 – 50 | 50 ± 1 (2) | nd |
| AA 10 | < 0.1 (7) | 70 ± 13 (3) |
Besides cAMP, arachidonic acid (AA) was shown to elicit Ca2+-influx. AA acts by eliciting Ca2+-release from Ca2+-stores which is thought to induce a capacitative Ca2+-influx [7,23]. In the LvsA minus mutant AA did not evoke Ca2+-influx in contrast to the parent strain (Table 4). Instead Ca2+-efflux occurred. In seven independent experiments 6–10 μM AA induced an efflux of Ca2+ of 276- ± 153 pmol/107cells.
Analysis of vesicular Ca2+-transport in the mutant revealed that transport activity was present in all three fractions of P0, P1 and P2 albeit at a somewhat lower extent than in Ax2 (Table 5). Therefore, the small amount of cAMP-induced Ca2+-influx observed in the LvsA minus strain cannot be due to the lack of vesicular Ca2+-transport in the mutant.
Table 5.
Vesicular Ca2+-transport of LvsA-. The specific Ca2+-transport activity of vesicular fractions of LvsA- and Ax2 is shown. Data are means ± SD. The number of independent experiments is given in brackets.
| Ca2+-uptake nmol/mg | ||
| LvsA- | Ax2 | |
| P0 | 42 ± 19 (5) | 68 ± 9.8 (3) |
| P1 | 29 ± 17 (3) | 36 ± 7.9 (3) |
| P2 | 15 ± 3 (3) | 23 ± 2 (3) |
Discussion
In this study we have shown that one of the acidic compartments of Dictyostelium, the CV-system, is required for cAMP-induced Ca2+-influx in intact cells and that isolated CV-vesicles mediate an essential part of Ca2+-transport as well as Ca2+-release in response to AA. Previously, it was only known that acidic Ca2+-stores were involved. The use of dajumin-GFP-labeled CV allowed a 7-fold enrichment of CV by a specific method utilizing antibodies directed against the GFP-tag. Ca2+-transport was enhanced 5- to 6-fold. This result is due to Ca2+-transport activity of another Ca2+-store that was retained in the flow through (P0F). Enriched CV displayed a potent and efficient Ca2+-transport that resulted in a low basal extravesicular Ca2+ concentration. Moreover, the Ca2+ taken up could be released by the calmodulin antagonist W-7 and by AA. Both compounds have been shown to be potent regulators of the acidic Ca2+-stores [23,24]. In addition, Ca2+-uptake was sensitive to CMA, an inhibitor of V-type H+-ATPases. Moreover, the V-type proton pump was concentrated in P0E. A similar enrichment was also obtained for the ER using calnexin-GFP-labeled cells. These Ca2+-stores differed from those of the CV by a greater sensitivity to BHQ, a lower responsiveness to AA and a higher basal extravesicular Ca2+-concentration following Ca2+-uptake.
In order to examine whether the Ca2+-transport occurring in CV is also involved in cAMP-mediated Ca2+-influx we made use of a mutant, LvsA minus. In this mutant the CV becomes disorganized during the discharge phase of the vacuole and calmodulin dissociates from the CV-membranes. The CV can swell but seems unable to discharge. The LvsA-protein that belongs to the family of BEACH-proteins transiently binds to the CV. We found that the mutant was strongly impaired in receptor-mediated Ca2+-influx when physiological cAMP-concentrations of 0.1–1 μM and even higher concentrations of 10 μM cAMP were applied. A significant Ca2+-influx was apparent only at elevated cAMP concentrations of 20 to 50 μM. Even so, influx was inhibited by about 75% as compared to the parent strain. Other compounds that induce Ca2+-influx in Dictyostelium are long chain fatty acids like AA. They act independently of the cAMP-receptor and of cell differentiation [7]. Similarly to cAMP, AA did not cause Ca2+-influx in the LvsA-minus-strain, instead Ca2+-efflux occurred. These results demonstrate that in order to activate capacitative Ca2+-influx an intact CV-system is absolutely necessary.
What is the role of the LvsA-protein in Ca2+-influx?
The function of the LvsA-protein in Dictyostelium is not yet known. BEACH-proteins belong to several classes and only a few proteins have been characterized. The FAN-protein mediates TNF-activation via the MAP-kinase pathway [25]. Neurobeachin conveys anchoring of the regulatory subunit of protein kinase A (PKA) [26]. In both cases, the BEACH-protein participates in signal transduction that could lead to altered Ca2+-regulation in Dictyostelium. Elevated cAMP-levels in Dictyostelium were shown to affect both, basal [Ca2+]i and stimulus-induced [Ca2+]i-transients [27].
Apart from this, it was shown that the loss of the LvsA-protein was accompanied by a loss of calmodulin from the CV-membranes. Since we have shown that the calmodulin antagonist W7 inhibits Ca2+-transport of acidic Ca2+-stores [24] we conclude that Ca2+-uptake by the CV is inhibited in the absence of calmodulin. This could be an explanation of the severe reduction of cAMP-mediated Ca2+-influx measured in the LvsA--mutant strain.
Conclusion
Among the acidic Ca2+-stores the contractile vacuole represents an efficient Ca2+ transport organelle that sequesters Ca2+ to low extravesicular Ca2+ concentrations and releases Ca2+ in response to arachidonic acid. Furthermore, it is essential for the occurrence of receptor-mediated Ca2+ influx to physiological concentrations of cAMP.
Methods
Chemicals
N-(6-aminohexyl)5-chloro-1-naphthalene sulfonamide (W-7) 2,5-di(tert-butyl)-1,4-hydroquinone (BHQ) and concanamycin A (CMA) were from Fluka (Buchs, Switzerland). Fluo-3 was obtained from MobiTec (Göttingen, Germany), inositol 1,4,5-triphosphate and thapsigargin from Alexis (Grünberg, Germany) and arachidonic acid from Sigma (München, Germany). The μMACS GFP-tagged protein isolation kit was purchased from Miltenyi Biotec (Bergisch Gladbach, Germany).
Cells and culture
The strains were grown in liquid culture as discribed by Sonnemann et al. [28]. The dajumin-GFP [14] and calnexin-GFP [29] strains were raised in the presence of 50 μg/ml G418. The LvsA-minus-strain [16] was grown with 100 μg/ml ampicillin and 60 μg/ml streptomycin and its parent strain DH1 additionally with 20 μg/ml uracil. Differentiation was induced by washing the cells twice in ice cold Sørensen phosphate buffer (17 mM KH2/Na2H, PO4, pH 6.0). Cells were shaken on a rotary shaker at 23°C, 150 rpm at 2 × 107 cells/ml until use.
Preparation of vesicles
20 ml 2 × 107cells/ml were shaken in Sørensen phosphate buffer containing 5 mM EGTA for 2–4 h, washed once in ice-cold 20 mM Hepes buffer, pH 7.2, resuspended at 2 × 108 cells/ml, and lysed by passage through nuclepore filters. Immediately, 3% sucrose, 50 mM KCl, 1 mM MgCl2, 10 μg/ml leupeptin, 1 μg/ml aprotinin and 1.25 mM dithiothreitol were added (final concentration). After centrifugation for 5 min at 150 × g in order to remove unbroken cells, the supernatant was further fractionated by centrifugation for 5 min at 3800 × g. The sediment (P0) was resuspended in 1 ml 10 mM Tris buffer, pH 7.8, 3% sucrose, 50 mM KCl, 1 mM MgCl2, 20 μg/ml leupeptin and 2 μg/ml aprotinin (TKS-buffer). The supernatant was centrifuged at 12.000 × g for 20 min. The sediment (P1) was resuspended in 0.9 ml TKS-buffer. The supernatant was further centrifugated at 40.000 × g for 30 min. The fluffy sediment was resuspended in about 0.6 ml TKS-buffer and designated P2.
Purification of vesicles with anti-GFP-magnetic microbeads
400 μl P0 suspended in 10 mM Tris-buffer, pH 7.8, 50 mM KCl, 20 μg/ml leupeptin and 2 μg/ml aprotinin (TK-buffer) was incubated for 30 min at 4°C with 100 μl anti-GFP-microbeads and then transferred to a Mini MACS-separation column exposed to a magnetic field. The column had been rinsed with 500 μl TK-buffer. The flow through was centrifuged for 5 min at 3.800 × g. The sediment was resuspended in 150 μl of TKS-buffer and designated P0F. In the meantime the column was washed with 500 μl TK-buffer, 500 μl TK-buffer containing 0.1 M NaCl and 500 μl TK-buffer containing 0.3 M NaCl. In case the column was running inefficiently the contents were transferred to a new column by pressing 500 μl TK-buffer through the old column in the absence of a magnetic field onto the new column positioned in the field. The final elution was performed in the same way after the column had been washed free of salt with TK-buffer. The washing solutions usually contained negligible amounts of the GFP-label. The purified eluate was centrifuged for 5 min at 3.800 × g, resuspended in about 350 μl TKS-buffer and designated P0E.
In most experiments the cells were treated with 8 μM latrunculin B for 20 min before lysis to disrupt F-actin filaments. Under this condition the cells rounded up and became immotile. Nonetheless, the dajumin-GFP-label remained associated with P0 and purification did not change significantly.
Ca2+-transport
Ca2+-transport was measured essentially as described [23]. In brief, about 60 μl fraction was added to 10 mM Hepes, pH 7.2, 50 mM KCl, 3% sucrose, 6 μg/ml antimycin A, 6 μg/ml oligomycin A, 100 μM NaN3, 2 mM MgCl2, and about 0.5 μM Fluo-3 in a total volume of 1 ml. Ca2+-uptake was initiated by addition of 400 μM ATP. Fluo-3 fluorescence was monitored at 505 nm excitation and 526 nm emission with a fluorimeter (Perkin Elmer 650-10S, Überlingen, Germany). Since [Ca2+]ev at the beginning of the experiment is not known and Ca2+-ATPase activity is increasing with increasing Ca2+-concentrations the results shown in Table 2 were corrected for the same initial [Ca2+]ev in order to relate the outcome of different experiments to each other. This was done by using a calibration curve at different [Ca2+]ev obtained with either P0 or P1, respectively, and adjusting the measured value to a [Ca2+]ev of 1 μM. The Ca2+-ATPase activities of both, P0 and P1, increased by a factor of about 3.3 from 0.5 μM to 1.5 μM [Ca2+]ev. Statistical data are expressed throughout as the mean ± S.D.
Immunoblotting
Seven microgram of protein per lane were resolved by SDS-PAGE in 12.5% gels, blotted and immunoblotted with a V-type H+-ATPase antibody (kindly provided by Margaret Clarke) preferentially reacting with the 37/42 kD subunits of the proton pump.
Other measurements
cAMP-induced Ca2+-influx was measured as described [7]. The centrifugation time of the cells (LvsA- and DH1) during every washing step was shortened without any loss of cells. Protein concentrations were determined with the Coomassie protein assay reagent (Pierce) using bovine serum albumin as standard. Endosomal content was determined by fluid phase uptake of RITC-dextran as described, except that labeling was performed in phosphate buffer pH 6.0 [18].
Abbreviations
AA: arachidonic acid; BHQ: 2,5-di(tert-butyl)-1,4-hydroquinone; CV: contractile vacuole; CMA: concanamycin A; ER: endoplasmic reticulum; Tg: thapsigargin
Authors' contributions
DM performed the organelle experiments and in collaboration with CS and DFL designed the experiments and wrote the manuscript. DFL measured receptor-mediated Ca2+-influx and responses of living cells to AA; CS cultured the strains and prepared the figures. ADL contributed the strains LvsA- and DH1 and participated in the preparation of the manuscript as did AMT, who also contributed the GFP-strains and Figure 1.
Acknowledgments
Acknowledgements
We thank Günther Gerisch for contributing the GFP-strains and helpful comments on the manuscript and Mirate Iraki for performing the Western blot. We are grateful to the Deutsche Forschungsgemeinschaft for support. We are grateful to Margaret Clarke who suggested that the CV mediate cAMP-induced Ca2+-influx and for the kind gift of the V-type H+-ATPase antibody.
Contributor Information
Dieter Malchow, Email: Renate.Gimmi@uni-konstanz.de.
Daniel F Lusche, Email: Daniel.Lusche@web.de.
Christina Schlatterer, Email: Christina.Schlatterer@uni-konstanz.de.
Arturo De Lozanne, Email: a.delozanne@mail.utexas.edu.
Annette Müller-Taubenberger, Email: amueller@lrz.uni-muenchen.de.
References
- Heuser J, Zhu Q, Clarke M. Proton pumps populate the contractile vacuoles of Dictyostelium amoebae. J Cell Biol. 1993;121:1311–1327. doi: 10.1083/jcb.121.6.1311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nolta KV, Steck TL. Isolation and initial characterization of the bipartite contractile vacuole complex from Dictyostelium discoideum. J Biol Chem. 1994;269:2225–2233. [PubMed] [Google Scholar]
- Zhu Q, Clarke M. Association of calmodulin and an unconventional myosin with the contractile vacuole complex of Dictyostelium discoideum. J Cell Biol. 1992;118:347–358. doi: 10.1083/jcb.118.2.347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke M, Köhler J, Arana Q, Liu T, Heuser J, Gerisch G. Dynamics of the vacuolar H(+)-ATPase in the contractile vacuole complex and the endosomal pathway of Dictyostelium cells. J Cell Sci. 2002;115:2893–2905. doi: 10.1242/jcs.115.14.2893. [DOI] [PubMed] [Google Scholar]
- Moniakis J, Coukell MB, Janiec A. Involvement of the Ca2+-ATPase PAT1 and the contractile vacuole in calcium regulation in Dictyostelium discoideum. J Cell Sci. 1999;112:405–414. doi: 10.1242/jcs.112.3.405. [DOI] [PubMed] [Google Scholar]
- Newell PC, Malchow D, Gross JD. The role of calcium in aggregation and development of Dictyostelium. Experientia. 1995;51:1155–1165. doi: 10.1007/BF01944733. [DOI] [PubMed] [Google Scholar]
- Schaloske R, Malchow D. Mechanism of cAMP-induced Ca2+-influx in Dictyostelium: the role of phospholipase A2. Biochem J. 1997;327:233–238. doi: 10.1042/bj3270233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaloske R, Schlatterer C, Malchow D. A Xestospongin C-sensitive Ca2+ store is required for cAMP-induced Ca2+-influx and cAMP-oscillations in Dictyostelium. J Biol Chem. 2000;275:8404–8408. doi: 10.1074/jbc.275.12.8404. [DOI] [PubMed] [Google Scholar]
- Traynor D, Milne JL, Insall RH, Kay RR. Ca2+ signalling is not required for chemotaxis in Dictyostelium. EMBO J. 2000;19:4846–4854. doi: 10.1093/emboj/19.17.4846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlatterer C, Happle K, Lusche DF, Sonnemann J. Cytosolic [Ca2+]-transients in Dictyostelium discoideum depend on the filling state of internal stores and on an active SERCA Ca2+-pump. J Biol Chem. 2004;279:18407–18414. doi: 10.1074/jbc.M307096200. [DOI] [PubMed] [Google Scholar]
- Wilczynska Z, Happle K, Müller-Taubenberger A, Schlatterer C, Malchow D, Fisher PR. Release of Ca2+ from the endoplasmic reticulum contributes to Ca2+ signaling in Dictyostelium discoideum. Eukaryot Cell. 2005;4:1513–1525. doi: 10.1128/EC.4.9.1513-1525.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlatterer C, Walther P, Müller M, Mendgen K, Zierold K, Knoll G. Calcium stores in differentiated Dictyostelium discoideum: prespore cells sequester calcium more efficiently than prestalk cells. Cell Calcium. 2001;29:171–182. doi: 10.1054/ceca.2000.0181. [DOI] [PubMed] [Google Scholar]
- Marchesini N, Ruiz FA, Vieira M, Docampo R. Acidocalcisomes are functionally linked to the contractile vacuole of Dictyostelium discoideum. J Biol Chem. 2002;277:8146–8153. doi: 10.1074/jbc.M111130200. [DOI] [PubMed] [Google Scholar]
- Gabriel D, Hacker U, Köhler J, Müller-Taubenberger A, Schwartz JM, Westphal M, Gerisch G. The contractile vacuole network of Dictyostelium as a distinct organelle: its dynamics visualized by a GFP marker protein. J Cell Sci. 1999;112:3995–4005. doi: 10.1242/jcs.112.22.3995. [DOI] [PubMed] [Google Scholar]
- Becker M, Matzner M, Gerisch G. Drainin required for membrane fusion of the contractile vacuole in Dictyostelium is the prototype of a protein family also represented in man. EMBO J. 1999;18:3305–3316. doi: 10.1093/emboj/18.12.3305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwak E, Gerald N, Larochelle DA, Vithalani KK, Niswonger ML, Maready M, De Lozanne A. LvsA, a protein related to the mouse beige protein, is required for cytokinesis in Dictyostelium. Mol Biol Cell. 1999;10:4429–4439. doi: 10.1091/mbc.10.12.4429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerald NJ, Siano M, De Lozanne A. The Dictyostelium LvsA protein is localized on the contractile vacuole and is required for osmoregulation. Traffic. 2002;3:50–60. doi: 10.1034/j.1600-0854.2002.30107.x. [DOI] [PubMed] [Google Scholar]
- Flaadt H, Schaloske R, Malchow D. Mechanism of cAMP-induced H+-efflux of Dictyostelium cells: a role for fatty acids. J Biosci. 2000;25:243–252. doi: 10.1007/BF02703931. [DOI] [PubMed] [Google Scholar]
- Spector I, Shochet NR, Blasberger D, Kashman Y. Latrunculins--novel marine macrolides that disrupt microfilament organization and affect cell growth: I. Comparison with cytochalasin D. Cell Motil Cytoskeleton. 1989;13:127–144. doi: 10.1002/cm.970130302. [DOI] [PubMed] [Google Scholar]
- Rooney EK, Gross JD, Satre M. Characterisation of an intracellular Ca2+ pump in Dictyostelium. Cell Calcium. 1994;16:509–522. doi: 10.1016/0143-4160(94)90081-7. [DOI] [PubMed] [Google Scholar]
- Temesvari LA, Rodriguezparis JM, Bush JM, Zhang LY, Cardelli JA. Involvement of the vacuolar proton-translocating ATPase in multiple steps of the endo-lysosomal system and in the contractile vacuole system of Dictyostelium discoideum. J Cell Sci. 1996;109:1479–1495. doi: 10.1242/jcs.109.6.1479. [DOI] [PubMed] [Google Scholar]
- Flaadt H, Jaworski E, Malchow D. Evidence for two intracellular calcium pools in Dictyostelium - the cAMP-induced calcium influx is directed into a NBD-Cl- and 2,5-di-(tert-butyl)-1,4-hydroquinone-sensitive pool. J Cell Sci. 1993;105:1131–1135. doi: 10.1242/jcs.105.4.1131. [DOI] [PubMed] [Google Scholar]
- Schaloske R, Sonnemann J, Malchow D, Schlatterer C. Fatty acids induce release of Ca2+ from acidosomal stores and activate capacitative Ca2+-entry in Dictyostelium discoideum. Biochem J. 1998;332:541–548. doi: 10.1042/bj3320541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malchow D, Lusche DF, Schlatterer C. A link of Ca2+ to cAMP oscillations in Dictyostelium: the calmodulin antagonist W-7 potentiates cAMP relay and transiently inhibits the acidic Ca2+-store. BMC Dev Biol. 2004;4:7. doi: 10.1186/1471-213X-4-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adam-Klages S, Adam D, Wiegmann K, Struve S, Kolanus W, Schneider-Mergener J, Kronke M. FAN, a novel WD-repeat protein, couples the p55 TNF-receptor to neutral sphingomyelinase. Cell. 1996;86:937–947. doi: 10.1016/S0092-8674(00)80169-5. [DOI] [PubMed] [Google Scholar]
- Wang X, Herberg FW, Laue MM, Wullner C, Hu B, Petrasch-Parwez E, Kilimann MW. Neurobeachin: A protein kinase A-anchoring, beige/Chediak-higashi protein homolog implicated in neuronal membrane traffic. J Neurosci. 2000;20:8551–8565. doi: 10.1523/JNEUROSCI.20-23-08551.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lusche DF, Bezares-Roder K, Happle K, Schlatterer C. cAMP controls cytosolic Ca2+ levels in Dictyostelium discoideum. BMC Cell Biology. 2005;6:12. doi: 10.1186/1471-2121-6-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnemann J, Aichem A, Schlatterer C. Dissection of the cAMP induced cytosolic calcium response in Dictyostelium discoideum: the role of cAMP receptor subtypes and G protein subunits. FEBS Lett. 1998;436:271–276. doi: 10.1016/S0014-5793(98)01139-9. [DOI] [PubMed] [Google Scholar]
- Müller-Taubenberger A, Lupas AN, Li H, Ecke M, Simmeth E, Gerisch G. Calreticulin and calnexin in the endoplasmic reticulum are important for phagocytosis. Embo J. 2001;20:6772–6782. doi: 10.1093/emboj/20.23.6772. [DOI] [PMC free article] [PubMed] [Google Scholar]