

Abstract
Embryonic stem (ES) cells have been investigated in repair of the CNS following neuronal injury and disease; however, the efficacy of these cells in treatment of postinjury pain is far from clear. In this study, we evaluated the therapeutic potential of predifferentiated mouse ES cells to restore sensory deficits following spinal cord injury (SCI) in mice. The pain model used unilateral intraspinal injection of quisqualic acid (QUIS) into the dorsal horn between vertebral levels T13 and L1. Seven days later, 60,000 predifferentiated ES cells or media were transplanted into the site of the lesion. Histological analysis at 7, 14, and 60 days post-transplantation revealed that animals receiving ES cell transplants suffered significantly less tissue damage than animals receiving media alone. Transplanted cells provided immediate effects on both spontaneous and evoked pain behaviors. Treatment with ES cells resulted in 0% (n = 28) excessive grooming behavior versus 60% (18 of 30) in media-treated animals. In the acetone test (to assess thermal allodynia), mice recovered to preinjury levels by 12 days after ES cell transplant, whereas control animals injected with media after SCI did not show any improvement up to 60 days. Similarly, the von Frey test (to assess mechanical allodynia) and the formalin test (to assess nociceptive hyperalgesia) showed that transplantation of predifferentiated ES cells significantly reduced these pain behaviors following injury. Here we show that predifferentiated ES cells act in a neuroprotective manner and provide antinociceptive and therapeutic effects following excitotoxic SCI.
INTRODUCTION
Insult to the spinal cord initiates a devastating cascade of concomitant events that include anatomical, physiological, and neurochemical changes often leading to neuronal cell death and syrinx formation (1–4). Loss of motor function, altered sensory function, and/or development of chronic or neuropathic pain may develop depending on the location and severity of injury (5–7). Chronic central pain following spinal cord injury (SCI) occurs in 60% to 80% of SCI patients and is severely compromising, to both adjustment after injury and patient quality of life (8). Numerous neurosurgical, pharmacological, psychological, and physiological approaches have been explored, with limited success, to alleviate central pain following injury. Narcotic analgesics remain the most common approach in the management of moderate to severe pain (9); however, adverse side effects and the potential for addiction or tolerance to pharmacological modalities have led to guarded use of these agents in pain management involving continuous long-term treatment (9,10). Furthermore, these agents appear to be relatively ineffective in controlling more severe chronic pain syndromes, particularly central pain (11). Thus, a clear need exists for more efficacious and safer approaches in long-term chronic pain therapies.
In recent years, a rodent model of chronic pain following SCI has been developed and thoroughly characterized. The excitotoxic SCI model, developed by Yezierski and colleagues (4), has been used for the study of chronic pain syndromes following SCI. In this model, an intraspinal microinjection of the mixed AMPA/metabotropic glutamate receptor agonist quisqualic acid (QUIS) produces progressive pathological changes (i.e., neuronal loss, astrocytic scarring, spinal cavitations, and prominent inflammatory response) that closely resemble those observed following traumatic or ischemic SCI. Importantly, this model results in well-characterized pain behaviors that allow for a broad analysis of sensory dysfunction following injury (12).
It has been suggested that embryonic stem (ES) cells possess great potential for repair of the damaged central nervous system (CNS). Several studies have shown ES cells to survive, propagate, and even provide some beneficial effects in the rodent CNS after experimental injury (13–22). For treatment of central pain syndromes, there have also been reports using transplantation of GABAergic and serotonergic precursor cells of different origin (23–25). In a recent report, transplantation of naive neural stem cells derived from adult rat spinal cords significantly improved motor function after spinal cord injury (26). Unfortunately, it also caused aberrant host fiber sprouting associated with allodynia-like hypersensitivity of nonaffected forepaws. Therefore, given the highly reactive environment that exists following injury as well as the suggestion that glial cues predominate at the site of injury, directed predifferentiation of ES cells into neuronal progenitors may greatly improve survivability and effect following transplantation. In a recent collaborative study, we demonstrated that predifferentiated ES cells could provide sensorimotor improvement following traumatic brain injury without any side effects (27). Using a similar differentiation protocol, we attempted to investigate the potential of these cells to address pain syndromes that occur following SCI.
The focus of our study was to utilize mouse ES cells predifferentiated into neuronal and glial progenitors for treatment of pain syndromes following SCI. In the first stage of research, we used a modified retinoic acid protocol (28) to induce differentiation of mouse embryonic stem cells into neuronal and glial precursors suitable for transplantation. In the second stage of this project, we transplanted these cells into injured spinal cord and conducted a battery of sensory behavioral tests to evaluate associated pain syndromes including examination of both spontaneous and evoked pain behaviors. The results of this study demonstrate for the first time that neural progenitors generated from mouse ES cells can limit secondary tissue damage following SCI, alleviate induced pain behaviors, and prevent the development of chronic central pain.
MATERIALS AND METHODS
Embryonic Stem Cell Culture
Undifferentiated, pluripotent mouse embryonic stem (ES) cells (D3 cell line; American Type Culture Collection, Manassas, VA) were utilized in this study. Murine ES cells were grown, propagated, and differentiated into neural precursors using retinoic acid.
Murine ES cells were transfected with the pEGFP-N3 construct (Clontech, Palo Alto, CA, USA) in the presence of FuGENE 6 (Roche, Indianapolis, IN, USA) as described (27). Transfected colonies were then selected in the presence of 200 mg/mL G418 (Sigma) and FACS sorted on a FACSVantage SE flow cytometry system (Becton Dickinson) to achieve 90% of GFP-labeled cells. Transgenic D3 cells were further used for induction of neural cells.
Cells were cultured in gelatinized 25-cm flasks in a medium consisting of Dulbecco’s modified Eagle’s medium (DMEM), 15% fetal bovine serum (FBS) (ES grade), 100 units/mL penicillin, 100 μg/mL streptomycin, 2 mM l-glutamine, 0.1 mM nonessential amino acids, 10 ng/mL murine leukemia inhibitory factor (all from Stem Cell Technologies, Vancouver, Canada), and 100 μM β-mercaptoethanol (Sigma Chemical, St. Louis, MO, USA). Cells were passaged at approximately 80% to 90% confluence, usually every other day. Our published 4–/4+ protocol was used for differentiation of ES cells (28,29). Briefly, during differentiation induction maintenance medium was exchanged for differentiation medium (Iscove’s modified Dulbecco’s medium, 15% FBS, l-glutamine [2 mM], nonessential amino acids [0.1 mM], 100 units/mL penicillin, and 100 μg/mL streptomycin, but no LIF or β-mercaptoethanol). Passaged cells (2.5 × 106) in differentiation medium were placed in 100-mm, low-adhesion Petri dishes where they began to form embryoid bodies (EBs) after 1 to 2 days. The medium was replaced with fresh medium every 2 days. After 4 days, EBs were gently transferred to a 15-mL tube and allowed to stand until the EBs settled to the bottom (~10 min). The medium was replaced and EBs were transferred back to the original dish by pipetting. At day 5, all-trans retinoic acid (Sigma) (1 × 10−6 M) was added to the culture. Medium was changed and RA treatment was repeated on day 7. On day 9, EBs were collected and seeded on poly-l-ornithine/fibronectin-coated flasks in differentiation medium. The next day, medium was replaced with differentiation/Neurobasal plus B27 supplement 1:1. The cells were cultured for another 2 to 3 days and then harvested for the injections. The medium was aspirated and discarded, and the cells were washed with PBS, trypsinized, and centrifuged in DMEM/FBS, after which they were re-suspended in Neurobasal medium plus B27. At this point, the cells could be transplanted or plated for immunocytochemical analysis. We routinely produce ~60% neurons and less than 40% of glial cells using this protocol (28–30).
Animal Surgery
Mature 8-week-old male mice (outbred ICR strain; Charles River Laboratories, Wilmington, MA, USA) were housed with unlimited access to food and water and a 12-h light/dark schedule. All procedures, behavioral tests and tissue collection methods were approved by the East Carolina University Animal Care and Use Committee.
The technique of intraspinal injection was similar to that described in previous reports (4,45). Mice were prepared for surgery by shaving the back and washing the surgical area with betadine solution (Purdue Frederick, Stamford, CT, USA). Surgery was performed under isoflurane anesthesia (1% to 4% inhalation to effect). After a midline incision, the spinal muscles were dissected away from the spinal column. The spinous process and vertebral lamina of one spinal segment were removed, exposing the spinal cord. Unilateral microinjections were made in a single segment at spinal levels ranging from T12 to L2.
Glass micropipettes (tip diameter 10–15 μm) attached to a 10-μL Hamilton syringe were used for microinjections. The syringe was mounted on a micro-injector (Kopf 5000; David Kopf Instruments, Tujunga, CA, USA) attached to a micromanipulator. Injections were made between the dorsal vein and dorsal root entry zone at a depth of 250 to 350 μm below the surface of the cord. Each animal received 0.6 μL of 125 mM QUIS (Sigma) in PBS injected over 3 min (three tracks of 0.2 μL separated by 0.3 mm, parallel to the long axis of the cord). This amount of QUIS produces overgrooming behavior in ~60% of animals according to previous studies (31,32). After microinjections, muscles were sutured in layers and the overlying skin was closed with staples. A subcutaneous injection of 0.9% saline was given to prevent dehydration, and a topical antibiotic was applied to the surgical wound. Animals were returned to their home cages where they were allowed to recover for 7 days before further surgical procedures.
After a 1-week period of recovery, mice were anesthetized with isoflurane and washed with betadine. Skin staples and sutures were removed, and scar tissue at the site of previous injury was gently excised revealing sites of prior QUIS microinjections. Similar to the preceding microinjections, predifferentiated mouse ES cells were delivered into the injury (three tracks of 0.2 μL separated by 0.3 mm, parallel to the long axis of the cord) over a 3- to 5-min time. ES cells had to be delivered more gradually than QUIS due to the viscous nature of the concentration (~100,000 cells/μL). Throughout the procedure and before injections, cells were kept at 37 °C and re-suspended gently to prevent clumping. Media-treated animals received similar microinjections of QUIS and, 7 days later, microinjections of cell differentiation media equal in volume and location. Control animals did not receive any injections. After the injections, muscles were sutured and the skin was closed using staples. Topical antibiotic was applied to the surgical wound.
Behavioral Tests
Excessive grooming behavior
Animals were assessed daily for the presence of excessive grooming behavior (self-directed biting and scratching resulting in damage to superficial and deeper layers of the skin) following excitotoxic SCI. Assessment of grooming behavior was based on the following classification scheme: class I: hair removal over contiguous portions of a dermatome; class II: extensive hair removal combined with signs of damage to the superficial layers of skin; class III: hair removal and damage to dermal layers of skin; and class IV: subcutaneous tissue damage (experiment terminated) (4). Based on these criteria, all animals were divided into overgroomers (classes I–IV) and nonovergroomers. Because over-grooming usually developed from 8 to 12 days postinjury, the postlesion time point for rating severity of overgrooming behavior was 12 to 14 days. All experiments were performed in a blinded manner.
Thermal allodynia (acetone test)
To examine cold allodynia, each animal underwent behavioral testing to determine sensitivity to thermal stimuli (33). Before testing, a 15-min quiescent period was allowed for the animal to acclimate to the observation chamber. A small amount of acetone solution (50–100 μL) was applied to the glabrous skin of the ipsilateral paw while the animal stood on an elevated screen. During each test session, acetone was applied 5 times (at 5-min intervals). The number of responses was counted between each application. Responses included hindpaw lifting, licking, and withdrawal of the paw. Three preinjury testing sessions were used to determine baseline values. After injury, animals were tested every third day until day 21, then once every 7 days until day 60. The acetone and von Frey tests were conducted on separate days to ensure that they did not interfere with one another. All tests were performed in a blinded manner.
Mechanical allodynia (von Frey filament test)
The von Frey filament test has been widely used in experiments testing mechanical sensitivity following neurological insult (34–37). In our experiments, we used the von Frey test to assess preinjury mechanical sensitivity, post-QUIS mechanical sensitivity, and mechanical sensitivity after ES cell transplantation or media injection. Before testing, a 15-min quiescent period was allowed for the animal to acclimate to the observation chamber. Filaments with stimulus intensities ranging from 0.008 to 300 g were applied to the glabrous skin of each hind paw 8 times while the animal stood on an elevated screen. Filaments were applied to the point of bending, at which time substantiation of response or non-response was determined. Responses included hindpaw withdrawal and orientation toward the stimulus. During each test session, the filament that produced a threshold response (response to stimuli 50% of the number of times applied) in each animal was determined for both the left and right hindpaws. Three preinjury testing sessions were used to determine baseline values. After injury, animals were tested every third day until day 21, then once every 7 days until day 60. This test was used to evaluate injury-induced changes in mechanical threshold. All tests were performed in a blinded manner.
Nociceptive hyperalgesia (formalin test)
The formalin test is a well-characterized study of nociception often used to evaluate postinjury pain (38–43). Attention was paid to the ethical guidelines for investigations of experimental pain in conscious animals. The formalin test was conducted according to a previously established protocol (42). Before testing, a 15-min quiescent period was allowed for the animal to acclimate to the observation chamber. A 25-μL volume of formalin (5%) in sterile saline was injected subcutaneously into the dorsal surface of the ipsilateral hindpaw using a 30-G needle. Directly after the formalin injection, each animal was placed in a glass cylinder on a flat, reflective surface to allow clear observation of the injected paw. Responses included licking or flicking of the injected paw. Responses were recorded immediately after each formalin injection for a period of 50 min. The total number of responses recorded during periods of 0–5 min (first phase) and 15–50 min (second phase) after formalin injection were measured as indicators of pain. The formalin test was performed once on the day of humane killing in each time point (7, 14, or 60 days). All tests were performed in a blinded manner.
Tissue Collection and Processing
At 7, 14, or 60 days after ES cell or media microinjection (10 animals per survival period), animals were deeply anesthetized with sodium pentobarbital (100 mg/kg intraperitoneally). Each animal was intracardially perfused first with cold PBS (20 mL) and then cold 4% paraformaldehyde (20 mL) in PBS (pH 7.4); thoracic and lumbar spinal cord was removed and postfixed in 4% paraformaldehyde for 12 to 16 h. Fixed tissue was stored in 30% sucrose solution at 4 °C until sectioning.
For immunohistochemistry, tissue was embedded in tissue-freezing medium (Triangle Biomedical Science, Durham, NC, USA), and sectioned into 30-μm coronal slices using a cryostat microtome before being processed as free-floating sections.
Lesion Analysis
Serial reconstruction of the lesion was done as described previously (32). For lesion analysis, serial cross-sections of 50 μm were collected in 0.1 M PBS and mounted on superfrost microscope slides (Fisher Scientific, Pittsburgh, PA, USA). All sections were stained with toluidine blue to identify the area of neuronal loss and injury. Quantification of the rostral to caudal extent of QUIS-induced tissue damage was performed to compare the hypothesized neuroprotective effects of transplanted ES cells. Pathophysiology of transverse 50-μm sections were examined using a Nikon SMZ 800 dissecting microscope. The criteria used to distinguish intact from damaged tissue included the absence of neurons and the presence of degenerating neuron profiles and inflammatory cells (44,45). Cross-sections of the spinal cord were examined with light microscopy, and images of the sections were taken using Spot digital camera and Spot Software v. 3.5.8 (Diagnostic Instruments, Sterling Heights, MI, USA). Reconstructions of the area of tissue damage was made with ImageJ software (NIH). Areas of the gray matter and the area of the damage at the lesion epicenter as well as 20 sections rostral and 20 caudal (2200 μm total length) were measured using ImageJ. The area of normal gray matter for each 50-μm section was compared with the corresponding segmental level of uninjured control cords to determine the percentage of gray matter damage in the injured cord for the area of gray matter in segments T12 to L3. Mean percentages of gray matter damage at 50-μm intervals were calculated to create a longitudinal profile of the distribution of damage in each group. In addition, to obtain an overall descriptor of the amount of tissue damage, the total lesion volume was calculated by subtracting the volume of gray matter remaining in injured animals from the normal volume as determined from the control. Student t tests were used to determine differences in mean lesion volume between each QUIS-alone group and its corresponding stem cell treatment group. Approximately 4 mm of tissue was examined from each animal. If the full extent of damage could not be examined, the cord was excluded. The total length of tissue damage was compared for ES cell–transplanted and media-injected mice. Lesion analysis was performed only for 60-day groups of animals. The experiments were performed in a blinded manner.
Immunohistochemistry
Immunostaining of free-floating thoracic and lumbar spinal cord sections (30 μm) was performed as previously described (46,47) using the Elite ABC Kit (Vector Laboratories, Burlingame, CA, USA). Sections were first washed for 10 min in PBS with 0.3% Triton X-100 (PBST) and then incubated for 30 min in ice-cold methanol containing 1% H2O2 to abolish endogenous peroxidase activity. Sections were then washed twice in PBST for 10 min, followed by 1-h incubation with blocking solution, 10% normal goat serum in PBST. Sections were incubated with primary antibodies for 1 h to overnight at 4 °C. They were subsequently washed 3 times for 15 min with PBST and incubated with secondary bi-otinylated antibodies for 1 h. The sections were washed again 3 times in PBST for 15 min and then stained for 2 h using Elite ABC Kit (avidinbiotinylated horse-radish peroxidase complex) (Vector Laboratories). After staining, tissue antigens were visualized with a DAB Substrate Kit for Peroxidase (Vector Laboratories). Stained free-floating sections were placed on superfrost microscope slides (Fisher Scientific) and air-dried overnight. Slides were soaked in Citrisolve (Fisher Scientific) for two 15-min washes and permanently mounted with Permount (Fisher Scientific). Controls included tissue samples processed without primary antibody, without secondary antibody, and without ABC complex. All experiments were repeated 3 times.
Immunocytochemistry
Immunostaining of ES cells was performed as previously described (29). ES cells were grown on gelatin-coated coverslips. After differentiation induction with RA, ES cells were fixed with 4% paraformaldehyde in PBS and processed using the Elite ABC Kit. Cells were washed for 10 min in PBST and then incubated for 10 min in 80% methanol containing 1% hydrogen peroxide. The cells were then washed twice in PBST for 5 min before blocking with avidin and biotin for 15 min each, followed by 1-h incubation with blocking solution (10% normal goat serum in PBST). The cells were then incubated with primary antibodies 1 h to overnight at 4 °C. After incubation, primary antibodies were removed and cells were washed 3 times in PBST for 10 min each. The cells were subsequently incubated with secondary antibodies from the Elite ABC kit for 1 h. Secondary antibodies were removed and the cells were washed 3 times in PBST for 10 min each. The cells were then incubated for 1 h in ABC reagent. ABC reagent was removed and the cells were washed 3 times in PBS for 5 min before being stained with DAB. After DAB staining, the cells were washed in distilled water. Slides were allowed to air-dry overnight. Dry slides were soaked in Citrisolve (Fisher Scientific) for 15 min, and a coverslip was mounted using DPX (Fluka Biochemika, Switzerland). Immunostaining controls included cell samples without primary antibodies, without secondary antibodies, and without ABC complex.
Immunofluorescence
To study coexpression of antigens, cultured ES cells and tissue from spinal cords were stained using immunofluorescent, dual-staining techniques. Secondary antibodies conjugated with fluorescent dyes FITC (green fluorescence) and Texas red (red fluorescence) were used to show intracellular localization of diverse antigens of interest. Counterstaining with DAPI, a nuclear dye (blue fluorescence), was also performed.
Antibodies
The following antibodies were used for immunostaining procedures. Anti- GFP (chicken; polyclonal; 1:1000) (Chemicon), neuron-specific anti–MAP-2 rabbit polyclonal antibodies (immunoreactivity with neuronal MAP-2A and MAP-2B; 1:2000) (Chemicon), neuron-specific anti-NeuN (mouse; polyclonal; 1:500) (Chemicon), glial specific anti-GFAP (rabbit; polyclonal; 1:1000) (Chemicon), and mouse monoclonal antibodies against Lim2 (1:100) (Developmental Studies Hybridoma Bank) to detect precursors of dorsal interneurons, mouse monoclonal MNR2/HB9 (1:100) antibodies to detect motoneuron progenitors (Developmental Studies Hybridoma Bank), and anti-synapsin rabbit polyclonal antibodies 1:1000 (Sigma); antisynaptophysin mouse monoclonal 1:400 (Sigma), anti-PSD95 monoclonal 1:500 (Upstate Biotechnology, Waltham, MA, USA), and anti-phosphohistone H3 (Ser10) mouse monoclonal 1:100 (Cell Signaling Technology, Beverly, MA, USA) as a marker of mitotic cells. For immunofluorescence all primary antibodies were applied in 1:100 dilution.
Microscopy
All slides were examined under an Olympus IMT-2 inverted fluorescent microscope. Images were photographed using a Spot digital camera and Spot Software v. 3.5.8 (Diagnostic Instruments, Sterling Height, MI, USA).
Statistical Analysis
Statistical analysis was performed with 1-way ANOVA, and a posttest Newman-Keuls multiple comparison test was performed using GraphPad Prism (v. 3.00 for Windows; GraphPad Software, San Diego, CA, USA).
RESULTS
Behavioral Tests
Excessive grooming behavior
Animals were assessed daily for the presence of excessive grooming behavior (self-directed biting and scratching resulting in damage to superficial and deeper layers of the skin) following excitotoxic SCI. Because overgrooming usually developed from 8 to 12 days postinjury, the postlesion time point for rating severity of overgrooming behavior was 12 to 14 days. Of the 28 animals receiving SCI and stem cell treatment, none developed excessive grooming behavior, whereas 18 of the 30 animals receiving media injections (60.0%) developed excessive grooming behavior with latency of onset ranging from 8 to 12 days postinjury (P < 0.0001) (Table 1). The excessive grooming behavior targeted the abdomen, hip, and thigh areas (Figure 1). These areas corresponded to dermatomes associated with spinal segments at or caudal to the site of injury (T13 to L2). All grooming observed in media-injected mice was rated in severity as class II or III. Excessive grooming behavior is associated with the secondary injury cascade, and its onset strongly correlates to the amount of tissue damage at associated spinal segments. Excessive grooming behavior subsided approximately 7 weeks after injury in media-transplanted mice. The lack of overgrooming in ES cell–transplanted animals strongly suggests that stem cells transplanted at the site of injury greatly reduced the damage following excitotoxic SCI.
Table 1.
Presence of excessive grooming behavior in ES cell–transplanted and media-injected mice.
| Grooming Behavior Observed |
||||||
|---|---|---|---|---|---|---|
| Excessive Grooming (class I–IV), n (%) |
||||||
| Treatment | Total No. Mice | Normal, n (%) | I | II | III | IV |
| Predifferentiated ES cells | 28 | 28 (100) | 0 | 0 | 0 | 0 |
| Media | 30 | 12 (40) | 0 | 7 (23.3) | 11 (36.7) | 0 |
Figure 1.
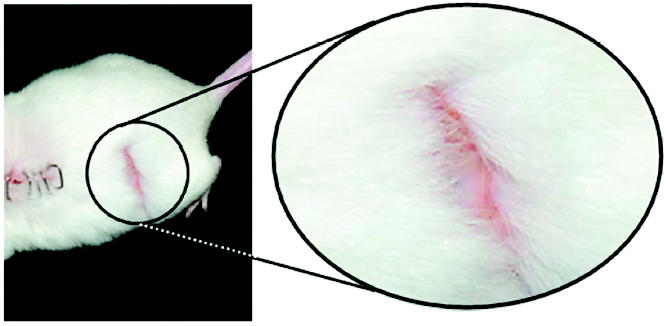
An example of excessive grooming behavior displayed by 60% of the media-injected mice. Excessive grooming behavior was not observed in ES cell–transplanted mice.
Thermal allodynia (acetone test)
Intact animals did not respond to stimulus at any time point. Before injury, application of room temperature acetone did not evoke response in any animals. After SCI, all groups of animals initially exhibited a highly significant (P < 0.001) number of responses compared with baseline values (Figure 2A-C). There were no statistical differences (P > 0.05) between experimental groups before treatment with stem cells or media. After ES cell transplantation (day 0, gray arrow), the number of responses immediately decreased, indicating a reduction of thermal allodynia. The numbers of responses recorded were consistent in all ES cell groups tested and returned to preinjury baseline levels at approximately 12 days after ES cell transplantation (P > 0.05), where they remained throughout the remainder of the experiment. Groups treated with media after SCI continued to show elevated sensitivity to thermal stimuli, indicated by amplified numbers of responses. The number of responses in media-treated animals was significantly higher (P < 0.001) than in ES cell–treated animals in every testing session through the remainder of the experiment (56 days). Media-injected animals did not return to baseline values at any point following injury (Figure 2B, C).
Figure 2.

Total number of responses to room temperature (23 °C) acetone. (A) Seven days after ES cell or media treatment. (B) Fourteen days after ES cell or media treatment. (C) Sixty days after ES cell or media treatment. Black arrow, QUIS injury; gray arrow, ES cell transplantation or media injection; •, ES cell–transplanted mice; ▪, media-injected mice. ** P < 0.001.
Mechanical allodynia (von Frey filament test)
Following QUIS injury, all mice demonstrated hypersensitivities to mechanical stimuli. The filament intensities required to elicit a response from each animal were much lower for the ipsilateral hindpaw in both ES cell–transplanted and media-injected mice. By 12 days after treatment with stem cells, the ipsilateral values returned to normal preinjury levels (Figure 3B, C). The sensory threshold of the ipsilateral hindpaw of media-treated animals did not return to baseline values during the observation/testing period (56 days). The effect of QUIS injury was less apparent in contralateral hindpaws (Figure 4A-C). Media-injected mice significantly differed from ES cell–transplanted mice (P < 0.01) at 9, 21, and 28 days. Significance (P < 0.001) was observed for the contralateral hindpaw between ES cell–and media-treated mice at 6, 12, 15, 18, 35, 42, 49, and 56 days.
Figure 3.

Allodynia to mechanical stimuli applied to the ipsilateral hindpaw for ES cell–transplanted and media-injected mice. (A) Seven days after ES cell (n = 7) or media (n = 10) treatment. (B) Fourteen days after ES cell (n = 10) or media (n = 10) treatment. (C) Sixty days after ES cell (n = 11) or media (n = 10) treatment. y-axis, percent of preinjury baseline values;•, ES cell–transplanted mice; ▪, media-injected mice; black arrows, QUIS injection; gray arrows, ES cell transplantation or media injection. *P < 0.01, **P < 0.001.
Figure 4.

Allodynia to mechanical stimuli applied to the contralateral hind paw in ES cell transplanted and media injected mice. (A) Seven days after ES cell (n = 7) or media (n = 10) treatment. (B) Fourteen days after ES cell (n = 10) or media (n = 10) treatment. (C) Sixty days after ES cell (n = 11) or media (n = 10) treatment. y-axis, percent of preinjury baseline values; •, ES cell–transplanted mice; ▪, media-injected mice; black arrows, QUIS injection; gray arrows, ES cell transplantation or media injection. *P < 0.01, **P < 0.001.
Nociceptive hyperalgesia (formalin test)
The injection of formalin resulted in a progressive biphasic response of the injected paw. The early phase began immediately after formalin injection and peaked during the first 5 min. The late phase followed, lasted the remainder of the observation period (50 min), and peaked between 20 and 40 min.
Figure 5A shows the results of the formalin test 7 days after transplantation of ES cells or media injection compared with uninjured, intact mice. ES cell–treated mice responded similarly to intact mice. Mice that received media injections showed significantly (P < 0.001) more responses than intact or stem cell–transplanted mice at both early and late phases.
Figure 5.
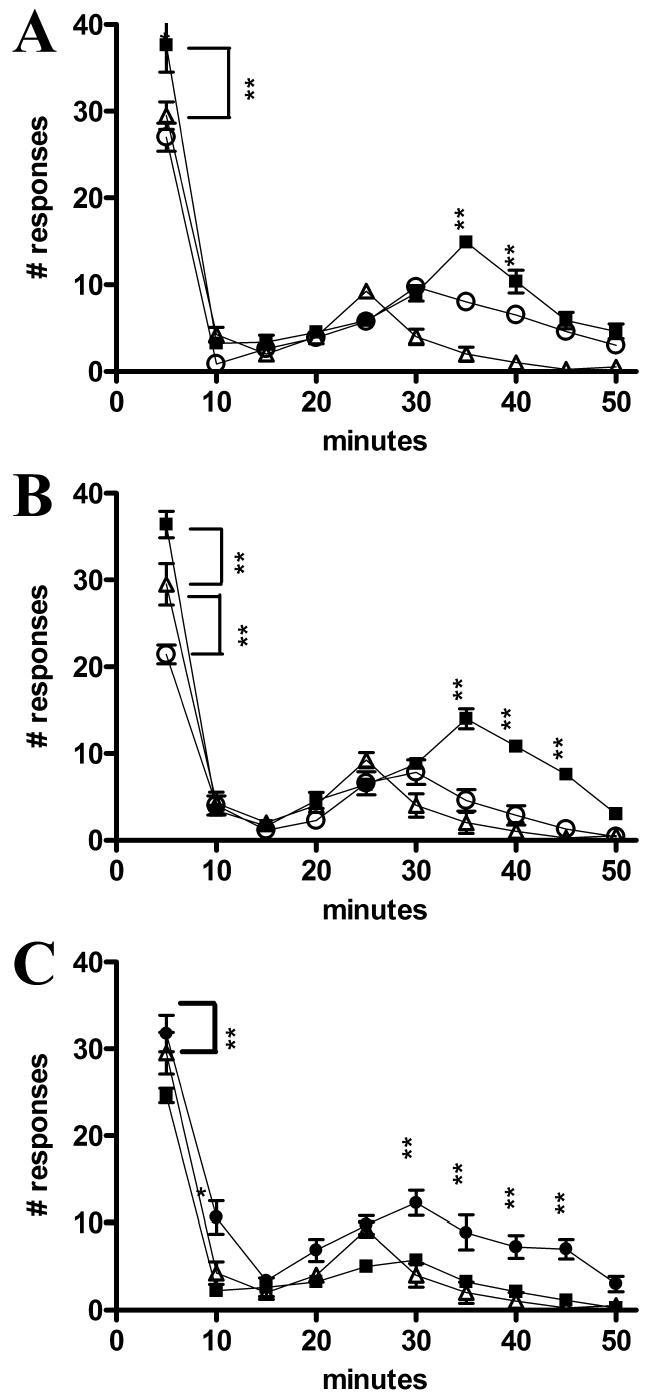
Number of responses to formalin injection over a 50-min observation period. (A) Seven days after ES cell (n = 7) and media (n = 10) treatment and intact mice (n = 5). (B) Fourteen days after ES cell (n = 10) and media (n = 10) treatment and intact mice (n = 5). (C) Sixty days after ES cell (n = 11) and media (n = 10) treatment and intact mice (n = 5). •, ES cell–transplanted mice;▪, media-injected mice;▵, intact mice. *P < 0.01, ** P < 0.001.
Figure 5B demonstrates the results of the formalin test 14 days after transplantation of ES cells or media injection compared with uninjured, intact mice. Media-injected animals demonstrated a significantly (P < 0.001) elevated response to the formalin injection in both early and late phases; however, ES cell–transplanted mice had significantly (P < 0.001) fewer responses than intact mice in the initial nociceptive phase.
Figure 5C illustrates the results of the formalin test 60 days after transplantation of ES cells or media injection compared with uninjured, intact mice. As in the 7- and 14-day groups, the number of responses recorded for the 60-day group of media-injected animals was significantly (P < 0.001) higher than intact or stem cell–transplanted animals at the later (inflammatory) phase of testing. The number of early-phase responses, although somewhat elevated in media-injected mice, showed no statistical significance (P > 0.05) compared with intact animals. ES cell–transplanted mice displayed significantly (P < 0.001) fewer early-phase responses than intact mice.
Lesion analysis
Previous research using the excitotoxic SCI model suggests a correlation between the onset of pain-related behaviors and the extent of tissue damage (48–50). A critical threshold of tissue damage appeared vital for initiation of spontaneous pain behaviors. Lesion analysis showed that at 60 days after ES cell transplantation or media injection, there was a dramatic difference in the longitudinal extent of tissue damage. The tissue damage observed in ES cell–treated mice, 60 days posttransplantation, extended 1118 ± 83.7 μm compared with 2176 ± 142.3 μm in media-injected mice (P < 0.0001). Given that none of the ES cell–transplanted mice displayed evidence of excessive grooming, it is hypothesized that the critical threshold of tissue damage required for the onset of overgrooming behavior was never reached. Previous studies have also suggested that the amount of tissue damage at the epicenter of injury is critical to the onset of spontaneous overgrooming behaviors (48–50). We also assessed the volume of the remaining gray matter in an unpaired t test. The analysis of spared tissue at 60 days revealed that mice transplanted with ES cells suffered significantly less tissue damage than mice that received media injections. Transplantation of ES cells did reduce the overall size of the lesion cavity, as demonstrated by a larger remaining volume of tissue. Statistical comparisons at 60 days revealed that whereas the average volume of gray matter in intact mice was 91.82 ± 2.250 μm3, the volume of spared gray matter in QUIS-injured mice was significantly decreased at 33.26 ± 1.538 μm3 (P < 0.0001). Mice treated with stem cells had a markedly higher volume of spared gray matter (58.92 ± 5.095 μm3; P < 0.005) then nontreated mice (Figure 6). The results indicate that transplantation of ES cells significantly reduced the extent of gray matter loss following injury.
Figure 6.
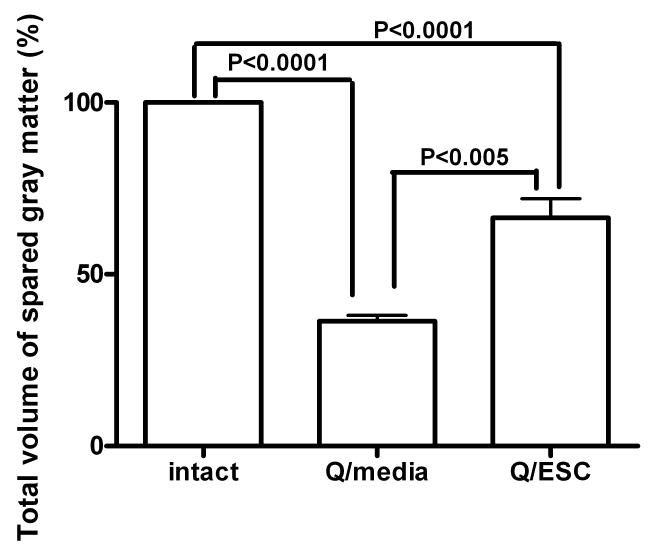
Volume of spared gray matter in ES cell–transplanted and media-injected mice at 60 days.
Histological Analysis
Analysis of the spinal cords showed a difference between ES cell–transplanted and media-injected animals. Fluorescent microscopy showed that the lesion cavity created by QUIS was filled with GFP-positive cells in stem cell–treated animals (Figure 7A). The transplanted cells appeared to fill the lesion area and to survive. Using high magnification at day 7, potential neuronal processes were observed branching from GFP-positive cells (Figure 7C). Immunohistochemical staining of spinal cord showed little specific staining against NeuN or GFAP antibodies at the site of the injury in media-treated animals at 7 days. Most media-injected animals had a fluid-filled area at the site of injection (resembling a syrinx) or collapse of the dorsal horn at 14 days (Figure 8A, C). In stem cell–treated animals, however, the collapse of the dorsal horn was less pronounced and NeuN-positive cells were visible in the dorsal horn (Figure 8B). In media-injected mice at 60 days, damage was limited to a severe narrowing of the neck of the ipsilateral dorsal horn (Figures 9A, C), whereas in ES cell–transplanted mice (Figure 9B), NeuN staining revealed somewhat shrunken dorsal horn ipsilateral to injury. These histological differences between media-and stem cell–injected animals correspond to the previous lesion analysis (Figure 6). Immunostaining against GFP and immunofluorescence against NeuN and GFAP demonstrated that the transplanted cells acquired neuronal phenotype and survived in the dorsal horn up to 60 days posttransplantation (Figure 10, upper panel). Conversely, some cells with acquired neuronal phenotype migrated into ipsilateral ventral horn (Figure 10, lower panel), which was visualized by MAP-2 neuron-specific staining. The histology of the spinal cord revealed evidence that supports our behavior data. It was apparent that less tissue damage was sustained in ES cell–transplanted mice, indicating a neuroprotective effect.
Figure 7.

Fluorescence microscopy of GFP-expressing cells 14 days after ES cell transplantation (A–C). (A) GFP cells fill epicenter of injury. Arrow indicates transplanted cells. (B) GFP cells migrating into surrounding tissue. Arrow indicates origin of migrating GFP-positive cells. (C) GFP cell with processes. Arrows indicate cell body and processes.
Figure 8.

Immunohistochemical staining 14 days after media injection (A, C) and ES cell transplantation (B, D). (A) Visualization of NeuN-positive cells in media-injected spinal cord. Arrows indicate lesion and the absence of neuronal staining ipsilateral to injury. (B) Visualization of NeuN-positive cells in ES cell–transplanted spinal cord. Arrows indicate side ipsilateral to injury and cell transplant. (C) Visualization of GFAP-positive cells in media-injected spinal cord. Arrows indicate lesion ipsilateral to injury and increased intensity of GFAP immunostaining ipsilateral to injury. (D) Visualization of GFAP-positive cells in ES cell–transplanted spinal cord. Arrows indicate dorsal horn preservation on side ipsilateral to injury and cell transplant.
Figure 9.

Immunohistochemical staining 60 days after media injection (A, C) and ES cell transplantation (B, D). (A) Visualization of NeuN-positive cells in media-injected spinal cord. Arrows indicate neuronal loss in all laminae ipsilateral to injury. (B) Visualization of NeuN-positive cells in ES cell–transplanted spinal cord. Arrows indicate side ipsilateral to injury. Note increased number of immunostained neuronal cells. (C) Visualization of GFAP-positive cells in media-injected spinal cord. Arrows indicate areas of high staining intensity in side ipsilateral to injury. (D) Visualization of GFAP-positive cells in ES cell–transplanted spinal cord. Arrows indicate increased intensity of GFAP staining in only a small area on side ipsilateral to injury and cell transplant.
Figure 10.

Survival of ES cells ipsilateral to the lesion at 60 days. Upper panel: Immunodetection of GFP (A, DAB staining), NeuN (B, immunofluorescence), and GFAP (C, immunofluorescence) in the ipsilateral dorsal horn. Small white or black arrows indicate neurons. Big white or black arrow points at collapsed dorsal horn. Lower panel: Double immunofluorescence for GFP and MAP-2 (a neuronal marker). (A) GFP fluorescent cells. (B) MAP-2–positive cells. (C) Merge of 2 images. Small white arrows indicate neurons.
Cytological Analysis
Daily observations were made using an inverted microscope during differentiation induction of ES cells. Digital images were taken of live cultures to record the progression of cells during the differentiation process. Immunocytochemical analysis after differentiation provided evidence that our ES cells displayed neural properties.
During differentiation induction, ES cells formed free-floating cell aggregates, or embryoid bodies (Figure 11A). We also examined mitotic figures during the treatments using immunostaining with anti-phosphohistone H3 (Ser10) antibodies. Phosphorylation at Ser10 of histone H3 is tightly correlated with chromosome condensation during mitosis and serves as a marker of dividing cells (Figure 11B). By the late stages of the 4–/4+ differentiation protocol, extensive neural clusters were observed in culture (Figure 11E).
Figure 11.

ES cells in vitro. Live culture pictures (A, C, E). (A) Free-floating embryoid bodies. (C) Differentiation induction, neural progenitors sprouting from embryoid body. (E) Neural network of differentiated cells. Immunocytochemical staining of cultured ES cells (B, D, F). (B) GFP-positive embryoid body viewed under fluorescence microscopy. Mitotic, histone H3–positive cells were visualized with Texas red. (D) Neural network. Staining against synapsin. (F) Double immunostaining of RA differentiated ES cells. Positive staining against Map-2 conjugated with FITC for green fluorescence indicates neuronal phenotype; LIM-2 conjugated with Texas red for red fluorescence allows further characterization of highly specialized dorsal interneurons.
The purpose of differentiating ES cells was to generate cell types suitable for transplantation into the spinal cord that might provide recovery of sensory function following injury. Therefore, immunophenotyping of differentiated cells prepared to be transplanted was performed on the same day as transplantation. The FACS analysis showed that the culture prepared for transplantation contained 8.430% ± 0.8514% of glial cell and 51.43% ± 2.944% of neurons, with remaining cells not yet expressing any specific neural markers. Of neurons there were 33.75% ± 2.903% LIM-2–positive cells and 32.74% ± 5.839% HB9-positive cells. These differentiated cells displayed positive immunostaining against synapsin (Figure 11D). In addition, immunostaining with antibodies against the specific marker of progenitors/young dorsal interneuron LIM-2 and against the specific marker of progenitors/young motor neurons HB9 confirmed that our protocol enabled differentiation of some cells into dorsal interneuron progenitors (Figure 11F), as well as into motor neurons (29).
DISCUSSION
The results of this study demonstrate that transplantation of predifferentiated ES cells can attenuate both evoked and spontaneous pain behaviors following excitotoxic SCI. After stem cell transplantation, both mechanical and thermal thresholds returned to preinjury values, and excessive grooming did not develop. The immediacy of neuroprotection and the sustained antinociception after ES cell transplantation demonstrate the potential therapeutic value ES cell transplantation may provide following SCI.
This study utilized an excitotoxic model of SCI to examine the effects of predifferentiated embryonic stem cells on post-SCI pain behaviors. The excitotoxic model is relevant based on findings indicating that QUIS produces pathological and behavioral changes comparable to those following ischemic and traumatic human SCI (12,44). The excitotoxic model of SCI has been widely used for the study of the secondary pathology of SCI. Moreover, this model of injury results in behavioral sequelae that include allodynia, hyperalgesia, and spontaneous pain behaviors. Using the excitotoxic SCI model, studies have examined modalities that may limit secondary damage and provide antinociception for chronic pain (31,32,51,52). However, there have been no studies showing alleviation of chronic pain following ES cell transplant.
One of the key features of secondary damage following SCI is inflammation (53,54). The presence of an inflammatory cascade following QUIS injury is believed to be a major contributor to the secondary neuronal injury and onset of pain-related behaviors. Moreover, the development of inflammation can lead to the induction of proinflammatory cytokines and may provide a link between inflammation and the development of chronic pain (32). The patterns observed in the inflammatory phase of the formalin test suggest that ES cells may be directly or indirectly inhibiting or interfering with peripheral inflammatory signaling. The secondary inflammatory reaction is thought to be due to several peripheral chemical mediators released from damaged cells, including histamine, kinin, serotonin, and prostaglandin (42). These mediators take part in the inflammatory response and are able to stimulate nociceptors. In the formalin test, ES cell–transplanted mice displayed response patterns similar to those of intact mice during this inflammatory phase, whereas media-injected mice showed significantly elevated frequency of responses. Moreover, ES cell–transplanted mice displayed fewer responses during the inflammatory phase of the formalin test in each subsequent time point examined (7 < 14 < 60 days). The decrease in frequency of responses in subsequent time points after transplantation suggests that our ES cells may be capable of inhibiting or disrupting peripheral inflammatory signals.
In the present experiments, the morphological characteristics following QUIS and media microinjections were similar to previous reports (4,12,32,44,49,55,56). Spinal cord segments at or near QUIS injection sites contained hypertrophied neuronal profiles and increased staining for GFAP, especially in areas of neuronal degeneration and border areas of spinal lesions. Morphological changes were directly related to the duration of survival following injury. ES cell–transplanted mice had less intense staining against GFAP. Additionally, NeuN immunostaining showed greater neuronal survival in ES cell–transplanted mice. These experimental data suggest that transplanted ES cells reduced neuronal cell death and glial scarring associated with secondary injury.
Given that none of the ES cell–transplanted mice displayed evidence of excessive grooming, it is hypothesized that the critical threshold of tissue damage required for the onset of over-grooming behavior was never reached. Previous studies have also suggested that the amount of tissue damage at the epicenter of injury is critical to the onset of spontaneous overgrooming behaviors (48–50). Correspondingly, the size of the lesion cavity at the epicenter of injury was reduced after stem cell transplantation in our experiments.
The immediacy of the behavioral improvements observed in ES cell–transplanted animals suggests that other factors are at work in providing recovery after injury. In a collaborative study, it was recently shown that murine ES cells, differentiated via the same modified 4–/4+ protocol, could significantly lessen the sensorimotor impairments and reduce the extent of cortical damage observed following traumatic brain injury (27). The rapid and early (day 2 to 3 after transplantation) sensorimotor improvements observed in that study may be attributed not to reestablishment of host circuitry, but rather to some neurotrophic influences. Similar improvements have been demonstrated by administration of neuroprotective agents (for example, MgCl2 and vitamin B3), suggesting that neuroprotective mechanisms may be involved (30,57–59). Several studies have shown that media from neural cell cultures provides beneficial effects by transfer of neurotrophic factors (17,60). Indeed, in a recent follow-up study (unpublished observations), we showed that transplanted ES cells are a source of BDNF and IL-6 and increased cAMP in the injured host cells in the spinal cord.
In this study, the observed improvements were not only rapid but also long lasting. All spontaneous and evoked pain behaviors were attenuated, the size of the lesion was greatly reduced, and immunostaining revealed survival of the transplanted and differentiated cells into neurons cells up to 60 days.
The results of this study have shown that transplantation of predifferentiated ES cells holds great therapeutic potential in preventing chronic pain following excitotoxic spinal cord injury and alleviating injury-related allodynia and hyperalgesia.
ACKNOWLEDGMENTS
We are grateful to Dr. Michael Hoane for advice on behavioral testing and lesion analysis. The monoclonal antibodies for Lim2 and MNR2/HB9 developed by Thomas M. Jessell were obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the Department of Biological Sciences, the University of Iowa, Iowa City, IA, USA.
This work was supported in parts by grant from North Carolina Biotechnology Center #2004-MRG-1104 (A.K.M.).
Footnotes
Online address: http://molmed.org
REFERENCES
- 1.France RD, Krishnan KRR. (1988) Chronic pain. American Psychiatric Press, Washington, DC, pp. xx, 561.
- 2.Guttmann L. (1973) Spinal cord injuries: comprehensive management and research. Blackwell Scientific, Oxford, pp. xiii, 694 p.
- 3.Windle WF. (1980) The spinal cord and its reaction to traumatic injury: anatomy-physiology-pharmacology-therapeutics. M. Dekker, New York, pp. xi, 384.
- 4.Yezierski RP, Liu S, Ruenes GL, Kajander KJ, Brewer KL. Excitotoxic spinal cord injury: behavioral and morphological characteristics of a central pain model. Pain. 1998;75:141–55. doi: 10.1016/S0304-3959(97)00216-9. [DOI] [PubMed] [Google Scholar]
- 5.Siddall PJ, Loeser JD. Pain following spinal cord injury. Spinal Cord. 2001;39:63–73. doi: 10.1038/sj.sc.3101116. [DOI] [PubMed] [Google Scholar]
- 6.Balazy TE. Clinical management of chronic pain in spinal cord injury. Clin J Pain. 1992;8:102–10. doi: 10.1097/00002508-199206000-00007. [DOI] [PubMed] [Google Scholar]
- 7.Demirel G, Yllmaz H, Gencosmanoglu B, Kesiktas N. Pain following spinal cord injury. Spinal Cord. 1998;36:25–8. doi: 10.1038/sj.sc.3100523. [DOI] [PubMed] [Google Scholar]
- 8.Henwood P, Ellis JA. Chronic neuropathic pain in spinal cord injury: the patient’s perspective. Pain Res Manage. 2004;9:39–45. doi: 10.1155/2004/863062. [DOI] [PubMed] [Google Scholar]
- 9.Schmidt WK. (1997) An overview of current and investigational drugs for the treatment of chronic pain. In Therapeutic Developments in Chronic Pain. National Managed Healthcare Conference, pp. 1–17.
- 10.Hulsebosch CE. Recent advances in pathophysiology and treatment of spinal cord injury. Adv Physiol Educ. 2002;26:238–55. doi: 10.1152/advan.00039.2002. [DOI] [PubMed] [Google Scholar]
- 11.Sagen J. (1998) Transplantation for pain treatment. In Cell Transplantation for neurological disorders. Freeman TB, Widner H, Eds. Humana Press, Totowa, NJ.
- 12.Brewer KL, Yezierski RP. Effects of adrenal medullary transplants on pain-related behaviors following excitotoxic spinal cord injury. Brain Res. 1998;798:83–92. doi: 10.1016/s0006-8993(98)00398-9. [DOI] [PubMed] [Google Scholar]
- 13.Cao QL, et al. Pluripotent stem cells engrafted into the normal or lesioned adult rat spinal cord are restricted to a glial lineage. Exp Neurol. 2001;167:48–8. doi: 10.1006/exnr.2000.7536. [DOI] [PubMed] [Google Scholar]
- 14.Dong H, et al. Enhanced oligodendrocyte survival after spinal cord injury in Bax-deficient mice and mice with delayed Wallerian degeneration. J Neurosci. 2003;23:8682–91. doi: 10.1523/JNEUROSCI.23-25-08682.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Han SS, Kang DY, Mujtaba T, Rao MS, Fischer I. Grafted lineage-restricted precursors differentiate exclusively into neurons in the adult spinal cord. Exp Neurol. 2002;177:360–75. doi: 10.1006/exnr.2002.7995. [DOI] [PubMed] [Google Scholar]
- 16.Herrera J, et al. Embryonic-derived glial-restricted precursor cells (GRP cells) can differentiate into astrocytes and oligodendrocytes in vivo. Exp Neurol. 2001;171:11–21. doi: 10.1006/exnr.2001.7729. [DOI] [PubMed] [Google Scholar]
- 17.Kerr DA, et al. Human embryonic germ cell derivatives facilitate motor recovery of rats with diffuse motor neuron injury. J Neurosci. 2003;23:5131–40. doi: 10.1523/JNEUROSCI.23-12-05131.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Liu S, et al. Embryonic stem cells differentiate into oligodendrocytes and myelinate in culture and after spinal cord transplantation. Proc Natl Acad Sci U S A. 2000;97:6126–31. doi: 10.1073/pnas.97.11.6126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.McDonald JW. Repairing the damaged spinal cord. Sci Am. 1999;281:64–73. doi: 10.1038/scientificamerican0999-64. [DOI] [PubMed] [Google Scholar]
- 20.McDonald JW, Howard MJ. Repairing the damaged spinal cord: a summary of our early success with embryonic stem cell transplantation and remyelination. Prog Brain Res. 2002;137:299–309. doi: 10.1016/s0079-6123(02)37023-7. [DOI] [PubMed] [Google Scholar]
- 21.Myckatyn TM, Mackinnon SE, McDonald JW. Stem cell transplantation and other novel techniques for promoting recovery from spinal cord injury. Transpl Immunol. 2004;12:343–58. doi: 10.1016/j.trim.2003.12.017. [DOI] [PubMed] [Google Scholar]
- 22.Riess P, et al. Transplanted neural stem cells survive, differentiate, and improve neurological motor function after experimental traumatic brain injury. Neurosurgery. 2002;51:1043–52. doi: 10.1097/00006123-200210000-00035. [DOI] [PubMed] [Google Scholar]
- 23.Eaton MJ, et al. Transplants of neuronal cells bioengineered to synthesize GABA alleviate chronic neuropathic pain. Cell Transplant. 1999;8:87–101. doi: 10.1177/096368979900800102. [DOI] [PubMed] [Google Scholar]
- 24.Hains BC, Johnson KM, McAdoo DJ, Eaton MJ, Hulsebosch CE. Engraftment of serotonergic precursors enhances locomotor function and attenuates chronic central pain behavior following spinal hemisection injury in the rat. Exp Neurol. 2001;171:361–78. doi: 10.1006/exnr.2001.7751. [DOI] [PubMed] [Google Scholar]
- 25.Lin CR, et al. Intrathecal spinal progenitor cell transplantation for the treatment of neuropathic pain. Cell Transplant. 2002;11:17–24. [PubMed] [Google Scholar]
- 26.Hofstetter CP, et al. Allodynia limits the usefulness of intraspinal neural stem cell grafts; directed differentiation improves outcome. Nat Neurosci. 2005;8:346–53. doi: 10.1038/nn1405. [DOI] [PubMed] [Google Scholar]
- 27.Hoane MR, et al. Transplantation of neuronal and glial precursors dramatically improves sensorimotor function but not cognitive function in the traumatically injured brain. J Neurotrauma. 2004;21:163–74. doi: 10.1089/089771504322778622. [DOI] [PubMed] [Google Scholar]
- 28.Murashov AK, Pak ES, Hendricks WA, Tatko LM. 17beta-Estradiol enhances neuronal differentiation of mouse embryonic stem cells. FEBS Lett. 2004;569:165–8. doi: 10.1016/j.febslet.2004.05.042. [DOI] [PubMed] [Google Scholar]
- 29.Murashov AK, et al. Directed differentiation of embryonic stem cells into dorsal interneurons. FASEB J. 2005;19:252–4. doi: 10.1096/fj.04-2251fje. [DOI] [PubMed] [Google Scholar]
- 30.Hoane MR, Knotts AA, Akstulewicz SL, Aquilano M, Means LW. The behavioral effects of magnesium therapy on recovery of function following bilateral anterior medial cortex lesions in the rat. Brain Res Bull. 2003;60:105–14. doi: 10.1016/s0361-9230(03)00022-4. [DOI] [PubMed] [Google Scholar]
- 31.Abraham KE, McMillen D, Brewer KL. The effects of endogenous interleukin-10 on gray matter damage and the development of pain behaviors following excitotoxic spinal cord injury in the mouse. Neuroscience. 2004;124:945–52. doi: 10.1016/j.neuroscience.2004.01.004. [DOI] [PubMed] [Google Scholar]
- 32.Brewer KL, Bethea JR, Yezierski RP. Neuroprotective effects of interleukin-10 following excitotoxic spinal cord injury. Exp Neurol. 1999;159:484–93. doi: 10.1006/exnr.1999.7173. [DOI] [PubMed] [Google Scholar]
- 33.Choi Y, Yoon YW, Na HS, Kim SH, Chung JM. Behavioral signs of ongoing pain and cold allodynia in a rat model of neuropathic pain. Pain. 1994;59:369–76. doi: 10.1016/0304-3959(94)90023-X. [DOI] [PubMed] [Google Scholar]
- 34.Kim SH, Chung JM. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain. 1992;50:355–63. doi: 10.1016/0304-3959(92)90041-9. [DOI] [PubMed] [Google Scholar]
- 35.Kupers RC, Nuytten D, De Castro-Costa M, Gybels JM. A time course analysis of the changes in spontaneous and evoked behavior in a rat model of neuropathic pain. Pain. 1992;50:101–11. doi: 10.1016/0304-3959(92)90117-T. [DOI] [PubMed] [Google Scholar]
- 36.Kim SH, Na HS, Sheen K, Chung JM. Effects of sympathectomy on a rat model of peripheral neuropathy. Pain. 1993;55:85–92. doi: 10.1016/0304-3959(93)90187-T. [DOI] [PubMed] [Google Scholar]
- 37.Na HS, Leem JW, Chung JM. Abnormalities of mechanoreceptors in a rat model of neuropathic pain: possible involvement in mediating mechanical allodynia. J Neurophysiol. 1993;70:522–8. doi: 10.1152/jn.1993.70.2.522. [DOI] [PubMed] [Google Scholar]
- 38.Hunskaar S, Fasmer OB, Hole K. Formalin test in mice, a useful technique for evaluating mild analgesics. J Neurosci Methods. 1985;14:69–76. doi: 10.1016/0165-0270(85)90116-5. [DOI] [PubMed] [Google Scholar]
- 39.Hunskaar S, Berge OG, Broch OJ, Hole K. Lesions of the ascending serotonergic pathways and antinociceptive effects after systemic administration of p-chloroamphetamine in mice. Pharmacol Biochem Behav. 1986;24:709–14. doi: 10.1016/0091-3057(86)90578-2. [DOI] [PubMed] [Google Scholar]
- 40.Hunskaar S, Berge OG, Hole K. Dissociation between antinociceptive and anti-inflammatory effects of acetylsalicylic acid and indomethacin in the formalin test. Pain. 1986;25:125–32. doi: 10.1016/0304-3959(86)90014-X. [DOI] [PubMed] [Google Scholar]
- 41.Hunskaar S, Hole K. The formalin test in mice: dissociation between inflammatory and non-inflammatory pain. Pain. 1987;30:103–14. doi: 10.1016/0304-3959(87)90088-1. [DOI] [PubMed] [Google Scholar]
- 42.Rosland JH, Hunskaar S, Hole K. Modification of the antinociceptive effect of morphine by acute and chronic administration of clomipramine in mice. Pain. 1988;33:349–55. doi: 10.1016/0304-3959(88)90295-3. [DOI] [PubMed] [Google Scholar]
- 43.Tjolsen A, Berge OG, Hunskaar S, Rosland JH, Hole K. The formalin test: an evaluation of the method. Pain. 1992;51:5–17. doi: 10.1016/0304-3959(92)90003-T. [DOI] [PubMed] [Google Scholar]
- 44.Yezierski RP, Park SH. The mechanosensitivity of spinal sensory neurons following intraspinal injections of quisqualic acid in the rat. Neurosci Lett. 1993;157:115–9. doi: 10.1016/0304-3940(93)90656-6. [DOI] [PubMed] [Google Scholar]
- 45.Liu XZ, et al. Neuronal and glial apoptosis after traumatic spinal cord injury. J Neurosci. 1997;17:5395–406. doi: 10.1523/JNEUROSCI.17-14-05395.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Murashov AK, Talebian S, Wolgemuth DJ. Role of heat shock protein Hsp25 in the response of the orofacial nuclei motor system to physiological stress. Brain Res Mol Brain Res. 1998;63:14–24. doi: 10.1016/s0169-328x(98)00258-7. [DOI] [PubMed] [Google Scholar]
- 47.Murashov AK, et al. Crosstalk between p38, Hsp25 and Akt in spinal motor neurons after sciatic nerve injury. Brain Res Mol Brain Res. 2001;93:199–208. doi: 10.1016/s0169-328x(01)00212-1. [DOI] [PubMed] [Google Scholar]
- 48.Gorman AL, Yu CG, Ruenes GR, Daniels L, Yezierski RP. Conditions affecting the onset, severity, and progression of a spontaneous pain-like behavior after excitotoxic spinal cord injury. J Pain. 2001;2:229–40. doi: 10.1054/jpai.2001.22788. [DOI] [PubMed] [Google Scholar]
- 49.Abraham KE, McGinty JF, Brewer KL. Spinal and supraspinal changes in opioid mRNA expression are related to the onset of pain behaviors following excitotoxic spinal cord injury. Pain. 2001;90:181–90. doi: 10.1016/s0304-3959(00)00402-4. [DOI] [PubMed] [Google Scholar]
- 50.Yezierski RP. Pain following spinal cord injury: pathophysiology and central mechanisms. Prog Brain Res. 2000;129:429–49. doi: 10.1016/S0079-6123(00)29033-X. [DOI] [PubMed] [Google Scholar]
- 51.Yu CG, Fairbanks CA, Wilcox GL, Yezierski RP. Effects of agmatine, interleukin-10, and cyclosporin on spontaneous pain behavior after excitotoxic spinal cord injury in rats. J Pain. 2003;4:129–40. doi: 10.1054/jpai.2003.11. [DOI] [PubMed] [Google Scholar]
- 52.Plunkett JA, Yu CG, Easton JM, Bethea JR, Yezierski RP. Effects of interleukin-10 (IL-10) on pain behavior and gene expression following excitotoxic spinal cord injury in the rat. Exp Neurol. 2001;168:144–54. doi: 10.1006/exnr.2000.7604. [DOI] [PubMed] [Google Scholar]
- 53.Bethea JR, et al. Systemically administered interleukin-10 reduces tumor necrosis factor-alpha production and significantly improves functional recovery following traumatic spinal cord injury in rats. J Neurotrauma. 1999;16:851–63. doi: 10.1089/neu.1999.16.851. [DOI] [PubMed] [Google Scholar]
- 54.Bethea JR. Spinal cord injury-induced inflammation: a dual-edged sword. Prog Brain Res. 2000;128:33–42. doi: 10.1016/S0079-6123(00)28005-9. [DOI] [PubMed] [Google Scholar]
- 55.Brewer KL, Hardin JS. Neuroprotective effects of nicotinamide after experimental spinal cord injury. Acad Emerg Med. 2004;11:125–30. doi: 10.1111/j.1553-2712.2004.tb01421.x. [DOI] [PubMed] [Google Scholar]
- 56.Brewer KL, McMillan D, Nolan T, Shum K. Cortical changes in cholecystokinin mRNA are related to spontaneous pain behaviors following excitotoxic spinal cord injury in the rat. Brain Res Mol Brain Res. 2003;118:171–4. doi: 10.1016/j.molbrainres.2003.08.006. [DOI] [PubMed] [Google Scholar]
- 57.Hoane MR, Akstulewicz SL, Toppen J. Treatment with vitamin B3 improves functional recovery and reduces GFAP expression following traumatic brain injury in rats. J Neurotrauma. 2003;20:1189–99. doi: 10.1089/089771503770802871. [DOI] [PubMed] [Google Scholar]
- 58.Hoane MR, Barth TM. The window of opportunity for administration of magnesium therapy following focal brain injury is 24 h but is task dependent in the rat. Physiol Behav. 2002;76:271–80. doi: 10.1016/s0031-9384(02)00714-x. [DOI] [PubMed] [Google Scholar]
- 59.Hoane MR, Barth TM. The behavioral and anatomical effects of MgCl2 therapy in an electrolytic lesion model of cortical injury in the rat. Magnes Res. 2001;14:51–63. [PubMed] [Google Scholar]
- 60.Wilkins A, Majed H, Layfield R, Compston A, Chandran S. Oligodendrocytes promote neuronal survival and axonal length by distinct intracellular mechanisms: a novel role for oligo-dendrocyte-derived glial cell line-derived neurotrophic factor. J Neurosci. 2003;23:4967–74. doi: 10.1523/JNEUROSCI.23-12-04967.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]