

Abstract
Background
Ov-CHI-1 is a chitinase specifically expressed in the infective stage larvae of the human filarial parasite Onchocerca volvulus. Evidence has show that it could be a vaccine candidate, however, there is no data available regarding the immunological status of people naturally exposed to infective stage larvae and thus provoked by this antigen.
Method
We analysed the Ov-CHI-1-specific immune response present in four endemic foci of human onchocerciasis (Ecuador, Nigeria, Togo and Cameroon) by enzyme-linked immunosorbent assays and T-cell proliferation assays.
Results
In these foci of infection, antibodies to Ov-CHI-1 were found to be present in only 22% of individuals from Ecuador, but were detected in 42–62% of infected individuals in the three foci from West Africa (Nigeria, Togo and Cameroon). There was found to be no relationship between antibody level and age, gender, or infection intensity as indicated by microfilarial density and numbers of skin nodules. The isotype response to Ov-CHI-1 was dominated by the presence of IgG3, IgG1 was present to a lesser extent. Our results show a positive correlation between N- and C-termini of Ov-CHI-1 in their ability to provoke humoral and cellular immune responses in the human. Peripheral blood mononuclear cell (PBMC) proliferative responses to Ov-CHI-1 when assayed, were found to be significantly higher in the individuals from endemic areas and there was a statistically elevated response to Ov-CHI-1 in the infected individuals when compared to putative immune individuals.
Conclusion
Ov-CHI-1 is an antigen that we have found strongly induces both humoral and cellular immune responses in humans.
Introduction
Onchocerciasis is caused by infection with the parasitic filarial nematode, Onchocerca volvulus. Typical pathology of onchocerciasis are the symptoms of dermal atrophy and blindness. The parasitic disease is present in 36 countries of Africa, the Arabian Peninsula and the Americas. 18 million people are infected and a further 120 million people worldwide are at risk of infection [1]. Pathogenesis of filarial disease is thought to be caused mainly by host inflammatory and immune responses to the parasite and to microfilarial and adult worm antigens [2-4]. Although a recent study has related pathology present in the eye with the presence of endosymbiotic Wolbachia bacteria contained within the onchocerca worm [5]. It is likely that both the nematode and Wolbachia antigens are involved in the disease pathogenesis.
Strategies to eliminate onchocerciasis have, in the past, been based on vector control and / or mass treatment with the microfilaricidal drug ivermectin [6]. A more efficient control method has been suggested which would use drug treatment combined with a prophylactic vaccine. Therefore, it is important to characterise antigens playing critical roles in parasitic development and transmission processes. Advances made by the filarial genome project may be useful to this end. Protective immunity generated against filarial parasites have been linked, in one study, with the developmental stages of late L3 and L4 [7]. Irradiated L3 larvae have been found to induce protective immunity in rodent models of filarial disease [8-11], and several recombinant L3 antigens have been proposed as potential vaccine candidates [12,13]. The post-infective stage and L3 to L4 moult are significant developmental stages and the molecules expressed at this time may be of special interest.
We have cloned and characterised a chitinase gene from infective stage larvae of O. volvulus, Ov-CHI-1 [14,15]. Ov-CHI-1 is a chitinase with several distinct features, namely, stage-specificity, (only expressed within infective stage larvae); organ specificity, (localised within the inclusion bodies of the glandular oesophagus); being secreted during late L3/L4 transition; and exhibiting high immunogenicity in experimental immunisation [14-16]. Recent studies have highlighted the importance of this secreted, stage-specific chitinase in experimental vaccinations to filarial parasites. For example, Canlas et al., have shown that the monoclonal antibody to Brugia malayi chitinase, MF1, can cause the clearance of circulating B. malayi microfilaria (mf) in passive transfer experiments [17]. Immunization of Mongolian jirds with live attenuated Samonella typhimurium expressing active A. viteae chitinase resulted in a reduction in adult worm burden of 51.4% [Lucis R, personal communication, 1999]. DNA vaccination with Ov-CHI-1 in our laboratory also resulted in a 53% reduction in parasite survival in a mouse chamber model of onchocercasis [18]. Immunization of jirds with recombinant B. malayi chitinase also induced partial protection against microfilaremia although it did not reduce adult worm burden [19]. There is no data available however, on the immunological status of people naturally exposed to, and provoked by, this antigen. Thus, we have studied Ov-CHI-1-specific immune responses in several endemic foci of human onchocerciasis. We believe that data generated will increase knowledge of this molecule and will aid further evaluation of the role of Ov-CHI-1 in the pathogenesis of onchocerciasis.
Materials and Methods
Filarial material
Adult worms and microfilariae of O. volvulus were recovered from nodules excised, after full informed consent and ethical review, from patients attending a clinic at the Medical Research Laboratories (MRL) in Kumba, Cameroon (as part of a clinical management programme). Microfilariae were purified on discontinuous Percoll gradients. Blackflies (Simulium damnosum s.l.) infected with O. volvulus L3 larvae were obtained from the MRL, Kumba, Cameroon. Cryopreserved infective larvae were supplied by Dr Lustigman (New York Blood Centre, New York, USA) on behalf of the Edna McConnell Clark Foundation Onchocerciasis Resources Project (New York, USA).
Expression of recombinant chitinase antigens
The details of expression and characterisation of recombinant Ov-CHI-1 have been described previously [14]. Briefly, all recombinant antigens used in this study were expressed in BL21 E. coli using the expression vector pJC40 [20]. The full-length protein consists of 497 amino acids; the 5' protein encoded by the first 1107 bp cDNA and the 3' protein consists of 138 amino acids which was encoded by the 3' 528 bp cDNA. A His-tag sequence was fused at the NH2 terminus to allow the recombinant antigens to be purified by affinity chromatography on a Nickel column (Probond Resin, Invitrogen, Paisley, UK).
Human sera
Sera were obtained from the Edna McConnell Clark Foundation (EMCF) onchocerciasis serum bank, maintained at the Swiss Tropical Institute, Basel, Switzerland. Samples from the EMCF serum bank originated from endemic regions within Cameroon (n = 182), Togo (n = 67), Nigeria (n = 39) and Ecuador (n = 55). In each of these locations, sera were derived from individuals showing parasitological signs of current infection (classified as 'infected' [INF]), or from individuals that had neither palpable nodules nor positive skin snips (classified as 'putatively immune' [PI]). EMCF criteria for defining putative immunity include a minimum of 20 years residency in the endemic area and with no history of recent treatment (past 3 years). Sera from unexposed individuals were obtained from volunteers from the United Kingdom. For IgG subclass measurement, 77 samples were selected based on total IgG optical densities (ODs) over 0.44 OD, including 21 from Ecuador (PI 13, INF 8), 37 from Cameroon (PI 18, INF 19) and 19 infected sera from Hohoe, Ghana (all INF).
Enzyme-linked immunosorbent assays (ELISA)
Human sera were obtained as described above, and analysed by ELISA for IgG antibodies against O. volvulus L3 chitinase antigens. Briefly, Maxisorp plates (Nalge Nunc International, USA) were coated overnight with purified Ov-CHI-1 at a concentration of 1 μg/ml in 0.05 M carbonate buffer (pH 9.6). Wells were blocked by overnight incubation with 20% (v/v) soya milk in TST (Tris-Sodium chloride-Tween buffer). Sera were diluted 1:200 in 20% soya / TST and applied to duplicate plates for 2 hours at room temperature. The wells were washed in TST, and goat anti-human IgG (H+L) horseradish peroxidase conjugate (Nordic Immunological Laboratories, Tilburg, The Netherlands) was added at 1:3000 dilution in 20% soya / TST for 1 hour at room temperature. Plates were washed and the assay was developed using 0.02% 2,2'-azinobis-(3-ethylbenzthiazoline-6-sulphonic acid) (ABTS) (Sigma, Dorset, UK). The absorbance at wavelength 405 nm was read on a Dynastic MR 5000 plate reader.
IgG subclass ELISA
Isotype-specific antibodies were measured by using an Ov-CHI-1 ELISA, employing isotype-specific conjugates. Briefly, Maxisorp plates were coated overnight with purified Ov-CHI-1 at a concentration of 1 μg/ml in 0.05 M carbonate buffer (pH 9.6). Wells were blocked by overnight incubation with 20% (v/v) soya milk in TST. Sera were diluted 1:200 in 20% soya / TST and applied to duplicate plates for 2 hours at room temperature. The plates then were probed with monoclonal antibodies to human IgG1 (1:200), IgG2 (1:100), IgG3 (1:200) and IgG4 (1:400) (Nordic: Set MAU/IgG1-4, Nordic Immunological Laboratories, Tilburg, The Netherlands) for 2 hours at room temperature, washed and probed again with goat anti-mouse antibody conjugated with horseradish peroxidase (Bio-Rad, Hemel Hempstead, UK) at the dilution 1:1000 for 1 hour at room temperature. Plates were then washed and the assay was developed as described above.
PBMC proliferation assays
T-cell proliferation assays were conducted as described previously, [21] using freshly isolated peripheral blood mononuclear cells (PBMC) from PI, O. volvulus-infected (INF), and "endemic controls" (i.e., ethnically identical individuals living in a nearby area where no O. volvulus transmission has occurred) individuals. Briefly, PBMC were isolated by centrifugation on lymphocyte separation medium (Organ on Technical, Durham, NC, USA). The cells were washed and plated onto 96-well flat-bottomed tissue culture plates (Corning, Cambridge, MA, USA) at a concentration of 1 × 106 cells/ml in a volume of 200 μl of RPMI 1640 (Bio Whittaker, Walkersville, MD, USA) supplemented with 10% human AB serum (Bio Whittaker Walkersville, MD, USA), 0.08 mg/ml gentamycin (Life Technology Gibco BRL, Gaithersburg, MD, USA), and 2 mM L-glutamine (Biofluids, Rockville, MD, USA). The PBMC were stimulated with purified recombinant O. voluvus full length chitinase protein and the C-terminal protein at 10 μg/ml. Appropriate positive and negative control cultures were also included. All stimulations were done in triplicate. The plates were incubated at 37°C and 5% CO2 for 5 days, pulsed with 1 μci per well with tritiated thymidine (DuPont, Boston, USA) for 16 h, and then harvested onto filter mats for subsequent scintillation counting. Results were expressed as a geometric mean of stimulation indexes.
Statistical analysis
Non-parametric statistical analysis was performed using a Mann Whitney U test, for a groups comparison, Spearman Rank analysis was used for testing for correlations.
Results and discussions
Serological responses in human onchocerciasis
Antibody responses to Ov-CHI-1 were measured in individuals from onchocerciasis endemic foci, alongside controls from the United Kingdom (Figure 1A). Optical densities for samples that exceeded the mean for UK sera plus 3 standard deviations (OD = 0.185) were taken to be antibody positive. Responses from the Ecuadorian individuals were generally weak, with only 22% showing levels of specific IgG above the threshold for positive with a mean OD of 0.155. Amongst the individuals from Africa, the proportion of those responding was higher at 62% for Cameroon, (mean OD = 0.260), 60% for Togo, (mean OD = 0.241) and 42% for Nigeria, (mean OD = 0.205). The geometric mean OD from each area showed a significant difference when compared with that from UK control group (P < 0.05). The differences in responses between endemic foci from West Africa and Ecuador also showed statistical significance (P < 0.05, Mann-Whitney U test). This difference in the levels of responsiveness measured in Latin America and Africa may reflect the difference in intensity of transmission found between the two continents. Indeed, this difference in antibody production between Ecuadorians and Africans has been previously observed with other O. volvulus antigens, such as in responses to the muscle isoform of tropomyosin, Ov-TMY-1 [22]. The significantly higher levels of antibodies against Ov-CHI-1 observed to be present in individuals from endemic areas than in the controls suggested that the immunity to this antigen is acquired by frequent exposure to infective-stage larvae. Ov-CHI-1 is a relatively abundant molecule with strong immunogenicity that could induce a lasting immunogenic response.
Figure 1.
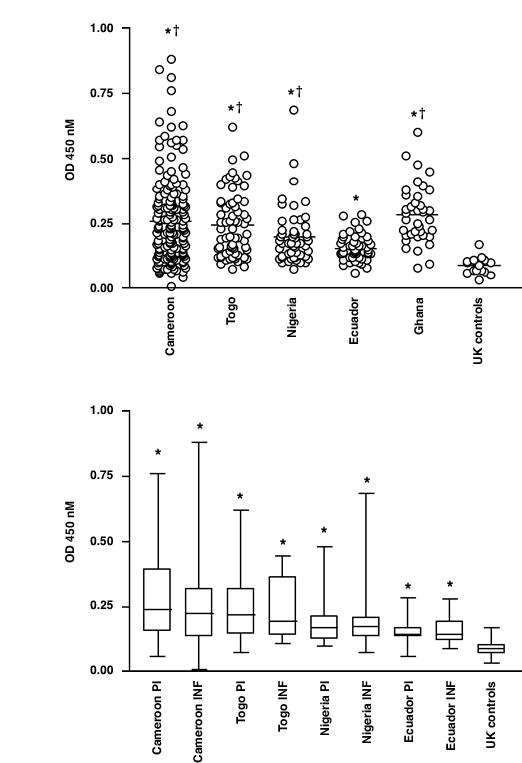
ELISA analysis of the IgG response to recombinant Ov-CHI-1 in human onchocerciasis. Figure 1A, Comparison of the response in sera of endemic individuals from 4 endemic foci with uninfected controls from the United Kingdom. Mean values are indicated by a horizontal line. Figure 1B, Comparison of the response in the sera of infected (INF) and putatively immune (PI) individuals from 4 endemic foci and from uninfected UK controls. Box and whisker plots illustrate the group mean values (horizontal bar), data points falling within the 25th to 75th percentile (boxed) and the range (vertical lines). * P = < 0.05 vs. UK controls † P = < 0.05 vs. Individuals from Eucador
Analysis of Ov-CHI-1 antibody levels found in infected (INF) and putatively immune (PI) individuals within the four endemic foci was carried out (Figure 1B). Data from the endemic foci was divided into two groups according to EMCF criteria. Antibody responses to this antigen appear to be remarkably persistent following infection, both INF and PI groups showed high levels although the antibody levels observed in the INF and PI groups were not significantly different from each other. Similar findings have also been reported in the investigation using recombinant Ov-DLC (cytoplasmic dynein light chain) and Ov-NIF-1 (A neutrophil inhibition factor) [23]. All these findings indicated that the acquired concomitant immunity seems to be present in the four endemic areas, which might have contributed to the deduction of the differences between PI and INF groups [24]. Furthermore, some Sowda individuals were included in the defined population who enrolled in our studies, (Sowda individuals have high levels of skin and lymphatic pathology but low or undetectable levels of microfilariae), which may affect the infectivity data if the grouping criteria was based on the numbers of microfilariae. Comparing antibody responses to Ov-CHI-1 with antibody responses to other molecules of O. volvulus [[25], Y.Wu et al., unpublished data] in the same populations, we found that Ov-CHI-1 is the only antigen, as yet, identified and measured in our laboratory that links elevated marginally antibody levels with putative immunity.
The human antibody response to Ov-CHI-1 is dominated by IgG3 isotype and to a lesser extent by the IgG1 subclass (Figure 2), the difference in levels between the two being significant (P < 0.01). IgG3 has been proposed to be involved in protective mechanisms against filarial parasites on the basis of its powerful ability to fix complement [26]. IgG3 has also been associated with mediating the targeting of antibody-dependent cell cytotoxicity (ADCC) by binding the Fcγ receptors of neutrophils, eosinophils and mononuclear cells [27]. A specific IgG3 to a 20 KDa antigen of O. volvulus has been found to predominate in amicrofilaraemic or putative immune individuals [28]. Boyer et al., suggested that IgG3 antibodies were involved in immunity to infection / re-infection and the control of microfilarial densities [29]. It is also notable that eosinophils have been implicated in the destruction of O. volvulus L3 larvae in a mouse model [30]. A contribution to elevated IgG3 levels could also be made by the carbohydrate epitopes in the induction of anti-Ov-CHI-1. In a study carried out by Mohanty et al., significantly elevated levels of IgG3 and IgG2 antibodies against filarial carbohydrate antigens were detected in PI individuals when compared with INF individuals. This suggests a potential role of carbohydrate antigens in the induction of protective immunity in human filariasis [31]. In our study, although high IgG3 levels coincided in some cases with the presence of microfilaremia, (Figure 2B), it is still notable that the highest levels of IgG3 were seen to be present in individuals free of microfilariae. In our data, IgG3 against full length Ov-CHI-1 shows neither age nor gender correlation (data not shown). Furthermore, there was no significant correlation of IgG1 and IgG3 levels (Figure 2C.) It is likely that induction of IgG3 is dependant on a property of the antigen itself. Further studies employing large numbers of individuals would help to ascertain this.
Figure 2.
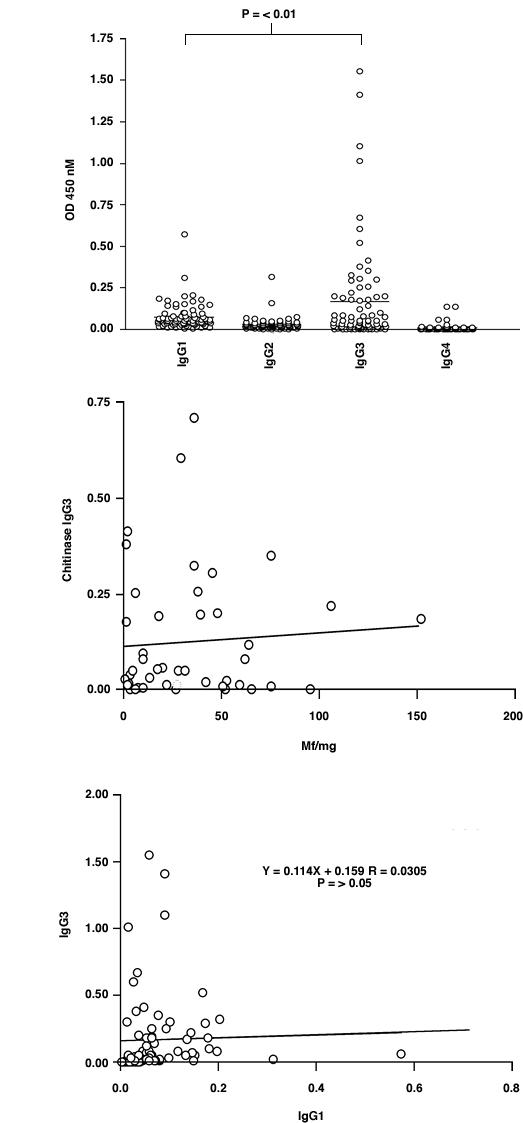
IgG subclass analysis by ELISA on Ov-CHI-1 full-length antigen using 77 selected sera (see text). The whole profile of IgG1-IgG4 responses is shown in Figure 2A. Figure 2B shows the relationship between microfilarial skin density and anti-Ov-CHI-1 IgG3 level in these sera. The relationship of IgG1 with IgG3 is shown in Figure 2C.
Studies of bacterial and yeast chitinase [32,33] indicate that the primary chitinase gene structure possesses four domains. A signal peptide; conserved catalytic domain present at the n-terminus; a serine/threonine-rich region located towards the C-terminal and a region which is considerably degenerated amongst filarial chitinases which adjoins the extreme C-terminal chitin binding domain. The location of our 5' and 3' Ov-CHI-1 protein is shown in Figure 3. Published mapping of the MF1 epitope of the microfilarial chitinase of B. malayi, a chitinase which shares 72% homology with Ov-CHI-1 at the amino acid level, has identified the most reactive region against monoclonal antibody MF1 to be located at the carboxy terminus of the gene [19]. The reactive region of the B. malayi microfilarial chitinase has a high PEST (proline, glutamic acid, serine, threonine) score in the serine/threonine-rich region that was suggested to be the calcium-binding region responsible for activating MF1 in B. malayi microfilariae, [34] although this epitope may not be shared with Ov-CHI-1. We measured the serological activity of patient sera against 5' and 3' chitinase separately, to ascertain where the epitopes recognised by human immune system against 5' and 3' chitinase are localized. This was done to ascertain the location of the epitopes recognised by human sera and to demonstrate which of these epitopes contributed to protection from infection as measured by microfilarial counts and nodule numbers present. To this end, we analysed the IgG level against 5' or 3' antigen separately, dividing them into PI and INF groups, as shown in Figure 4A. There was no difference found in levels of the IgG to 5' protein between PI and INF individuals, whilst IgG against the 3' Ov-CHI-1 was found to be significantly increased in the PI individuals compared to the INF individuals (P < 0.05). Panels of sera were also analysed by ELISA against the recombinant 5' and 3' proteins in parallel (Figure 4B). Levels of antibody to one antigen were plotted against levels to the other for all individual serum samples. Interestingly, a significant, positive correlation was observed in the sera tested, (Spearman Rank Correlation coefficient r = 0.5389, P < 0.001). This result indicates that individuals responded equally to 5' and 3' chitinase and the C-terminal and N-terminal proteins are closely related in terms of provoking humoral immune response.
Figure 3.
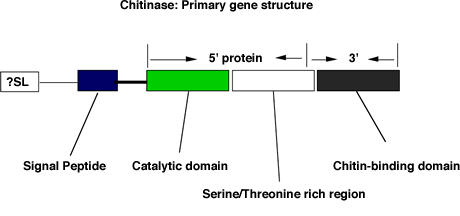
Schematic representation of filarial chitinase gene and the position of recombinant Ov-CHI-1 used in our studies
Figure 4.
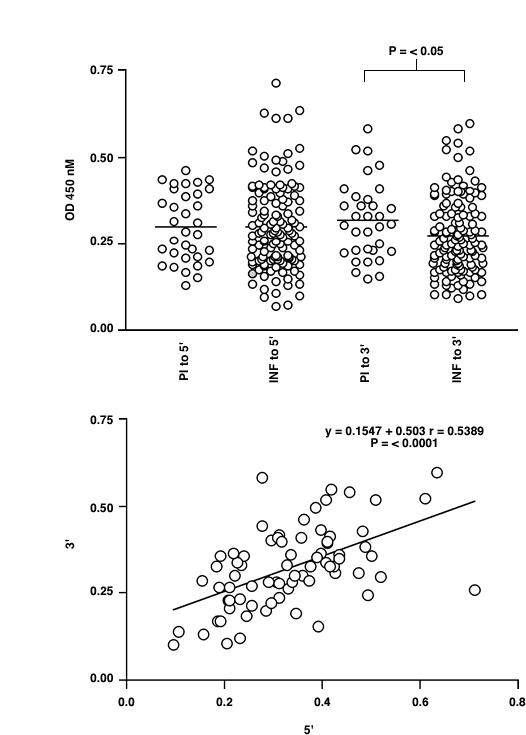
ELISA analysis of the IgG response to recombinant Ov-CHI-1 5' or 3' antigen in human onchocerciasis. Figure 4A: Comparison of the response in the sera of infected (INF) and putatively immune (PI) individuals. Figure 4B: The relationship of response to 5' antigen and 3' antigen for individual sera. The line of liner regression is shown for data sets in which there is a statistically significant correlation between responses to the two antigens (determined by Spearman's Rank Correlation Coefficient).
To investigate whether the T cell responses to Ov-CHI-1 were also involved in the processes of infection or immunity in human populations, we employed a T cell proliferation assay. Freshly isolated PBMCs from individuals whose immune status had been previously characterised [35,36] were used to measure the cellular immune responses either to the full length or to the C-terminal recombinant proteins ofOv-CHI-1, (Figure 5). Stimulation of PBMCs with the pokeweed mitogen as a control (data not shown) indicated that cells from all individuals in our experiments were in a valid condition. Results of the assays showed there was a statistically significant difference between controls (ethnically identical individuals living in a nearby area where no O. volvulus transmission has occurred) and INF and PI individuals (P = < 0.05). PBMCs from INF individuals had significantly greater proliferative responses to Ov-CHI-1 antigens than PBMCs from PI individuals (P = < 0.05). There seems to be a distinction between humoral and cellular responses in PI and INF individuals because PI individuals have higher antibody levels to Ov-CHI-1 than INF individuals, whilst T cell responsiveness of INF individuals was significantly higher than that of the PI individuals. The presence of segregation of humoral and cellular immune responses among PI and INF individuals indicates that it is possible that antibodies to Ov-CHI-1 could in some part mediate protection of the human from filarial infection. This phenomena could be caused by retention of memory B cells and high-affinity antibodies generated during the course on infection, the effectiveness of the antibodies produced improving steadily, so that it could provide long-term immunity to re-infection. It can also be noticed from Figure 5 that the cellular responses of human onchocerciasis exhibit similar pattern in PI and INF individuals to full-length antigen or 3' antigen. As the full-length protein has been characterized to be immunologically similar to 5' protein, we assume that the T-cell epitopes of Ov-CHI-1 are distributed on both termini of the protein. Ov-CHI-1 is an antigen capable of enhancing cell-mediated responses upon exposure to O. volvulus infection, whether cellular-mediated immunity is involved in protection against infection cannot be excluded. The results could also mean that the concomitant immunity of the infected individuals has to deal with these T cell responses.
Figure 5.
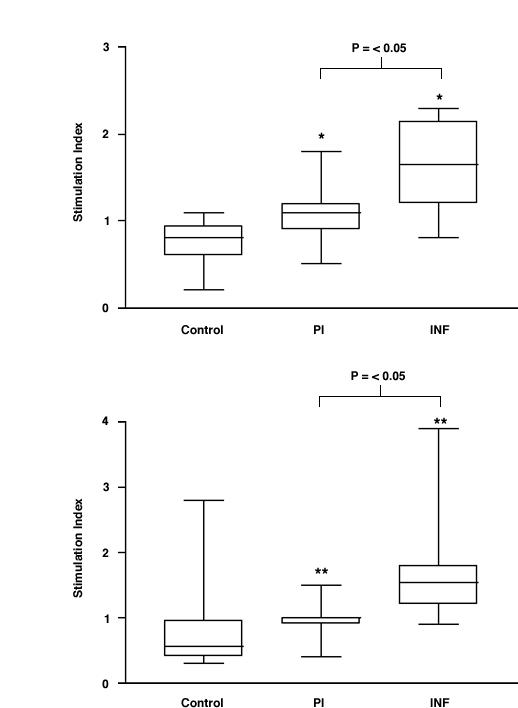
Human cellular immune responses to Ov-CHI-1. PBMC were isolated from individuals were stimulated with recombinant Ov-CHI-1 proteins. The results are expressed in stimulation indices. Figure 5A: the response to the full-length antigen. Figure 5B: the response to 3' antigen of Ov-CHI-1. * P = < 0.05 vs. controls ** P = < 0.01 vs. controls
When the data was analysed regarding measurement of total IgG versus full-length Ov-CHI-1, the results showed that there was no significant correlation between antibody levels when analysed with respect to age (Figure 6), sex (Figure 7), microfilarial densities (Figure 8) or nodule numbers (Figure 9) in any four endemic foci examined. The mean antibody titre in PI female individuals was found to be lower than that of INF female individuals, whilst in PI male individuals, it was relatively higher than that of INF male individuals (Figure 7). The results in Figure 8, demonstrate that the most intense IgG responses to Ov-CHI-1 were positively correlated with the microfilarial load of the skin in three of the four foci, excepting Togo. This could be interpreted as the antibody to Ov-CHI-1 having little effect on reduction of microfilarial density; it could also imply that it also has little effect on the adult worms. Ov-CHI-1 is a L3- stage specific antigen; it initiates synthesis in the late L2-stage of development, reaching its peak period of production when the third stage larvae are well developed. It is a secreted antigen, secretion being initiated on transmission from intermediate host to the human body. The expression of this larval specific chitinase is a transient process with a definite start and cessation. Considering these properties, we can assume that the immunity induced by Ov-CHI-1 would be limited to a short period following infection. In endemic areas where the transmission is very high, frequent exposure to Ov-CHI-1 is likely.
Figure 6.
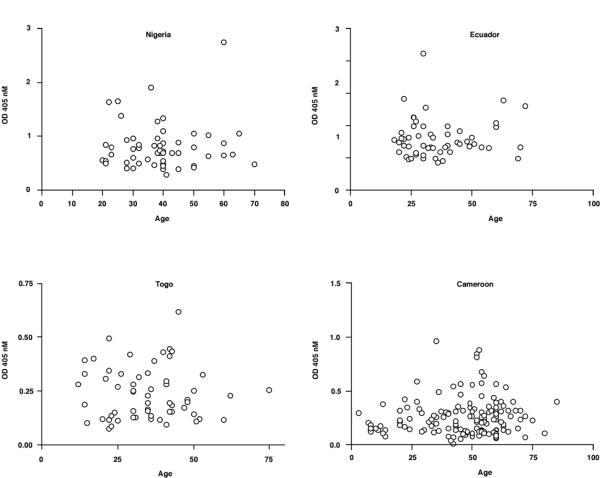
Analysis of IgG responses to Ov-CHI-1 categorized by age for individual individuals from four endemic regions. Figure 6A: Nigeria, Figure 6B: Ecuador, Figure 6C: Togo, Figure 6D: Cameroon.
Figure 7.
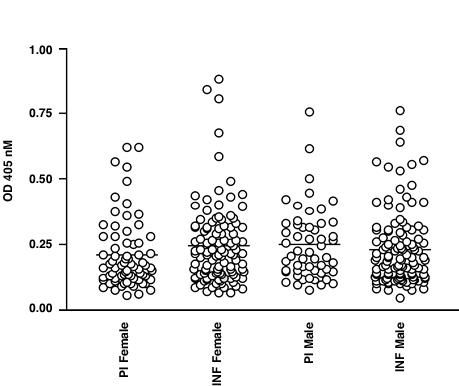
Analysis of IgG responses of human onchocerciasis to Ov-CHI-1 categorized by gender and comparison of responses in the sera of infected (INF) and putatively immune (PI) individuals from four endemic foci.
Figure 8.
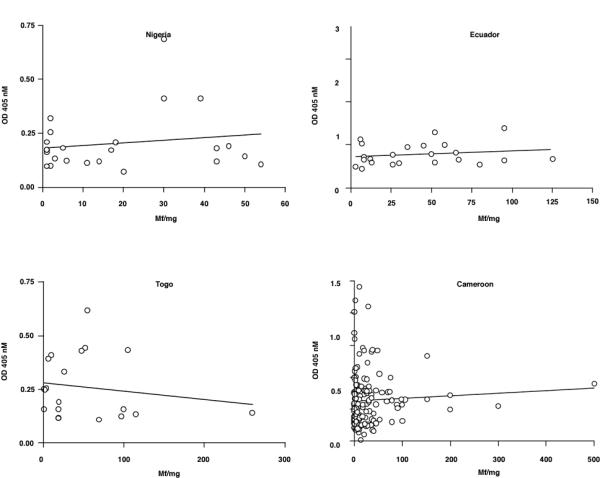
Analysis of IgG responses to Ov-CHI-1 plotted against microfilarial skin densities for individual individuals from four endemic regions. Figure 8A: Nigeria, Figure 8B Ecuador, Figure 8C Togo, Figure 8D. Cameroon. The line of linear regression is shown on each graph.
Figure 9.
Analysis of IgG responses to Ov-CHI-1 with respect to skin nodule number for individual subject from four endemic regions. Figure 9A: Cameroon, Figure 9B: Ecuador, Figure 9C: Nigeria.
This study reports the immune responses of human onchocerciasis patients to Ov-CHI-1 in endemic areas and has analysed the immunity generated against the larvae of O. volvulus. Although our results revealed that the natural infection of O. volvulus strongly provoked both humoral and cellular immune responses in individuals from endemic foci, there is insufficient evidence to support the use of OV-CHI-1 as a vaccine candidate against Onchocerciasis. Possibly, Ov-L3-CHI could be a molecule present in the developing infective stage larvae and employed as a target of prophylactic treatment.
Acknowledgments
Acknowledgements
We wish to thank Drs. Phil Hogarth and Nicki Weiss and the Edna McConnell Clark Foundation http://www.emcf.org/ for the provision of sera. This work was supported by a grant from the Wellcome Trust http://www.wellcome.ac.uk/.
Contributor Information
Yang Wu, Email: ywu@liv.ac.uk.
Gillian Egerton, Email: g.l.egerton@liv.ac.uk.
James S McCarthy, Email: J.McCarthy@sph.uq.edu.au.
Thomas B Nutman, Email: TNUTMAN@niaid.nih.gov.
Albert E Bianco, Email: t.bianco@wellcome.ac.uk.
References
- World Health Organisation Onchocerciasis (River Blindness): Fact Sheet No. 95, revised at February. 2000. http://www.who.int/inf-fs/en/fact095.html
- Philipp M, Davis TB, Storey N, Carlow CK. Immunity in filariasis: perspectives for vaccine development. Annu Rev Microbiol. 1988;42:685–716. doi: 10.1146/annurev.mi.42.100188.003345. [DOI] [PubMed] [Google Scholar]
- Ward DJ, Nutman TB, Zea-Flores G, et al. Onchocerciasis and immunity in humans: enhanced T cell responsiveness to parasite antigen in putatively immune individuals. J Infect Dis. 1988;157:536–543. doi: 10.1093/infdis/157.3.536. [DOI] [PubMed] [Google Scholar]
- Nelson G. Order Filariata: Filarial worms in Onchocerciasis. Advances in Parasitology. 1970;8:488–491. doi: 10.1016/s0065-308x(08)60254-7. [DOI] [PubMed] [Google Scholar]
- Saint Andre A, Blackwell NM, Hall LR, Hoerauf A, Brattig NW, Volkmann L, Taylor MJ, Ford L, Hise AG, Lass JH, Diaconu E, Pearlman E. The role of endosymbiotic Wolbachia bacteria in the pathogenesis of river blindness. Science. 2002;295:1892–1895. doi: 10.1126/science.1068732. [DOI] [PubMed] [Google Scholar]
- Molyneux DH. Onchocerciasis control in West Africa: current status and future of the Onchocerciasis Control Program. Parasitology Today. 1995;11:399–402. doi: 10.1016/0169-4758(95)80016-6. [DOI] [Google Scholar]
- Eisenbeises WF, Apfel H, Meyer TF. Protective immunity linked with a distinct developmental stage of a filarial parasite. J Immunol. 1994;152:735–742. [PubMed] [Google Scholar]
- Yates JA, Higashi GI, Brugia malayi. Vaccination of jirds with 60 cobalt-attenuated infective stage larvae. Am J Trop Med Hyg. 1985;34:1132–1137. doi: 10.4269/ajtmh.1985.34.1132. [DOI] [PubMed] [Google Scholar]
- Bancroft A, Devaney E. The analysis of the humoral response of the BALB/c mouse immunized with radiation attenuated third stage larvae of Brugia pahangi. 1993;15:153–162. doi: 10.1111/j.1365-3024.1993.tb00595.x. [DOI] [PubMed] [Google Scholar]
- Lucius R, Textor G, Kern A, Kirsten C. Acantholcheilonema viteae: vaccination of jirds with irradiation-attenuated stage-3 larvae and with exported larval antigens. Exp Parasitol. 1991;73:184–196. doi: 10.1016/0014-4894(91)90021-n. [DOI] [PubMed] [Google Scholar]
- Hayashi Y, Noda K, Shirasaka A, Nogami S, Nakamura M. Vaccination of BALB/c mice against Brugia malayi and B. pahangi with larvae attenuated by gamma irradiation. Jpn J Exp Med. 1984;54:177–181. [PubMed] [Google Scholar]
- Gregory WF, Atmadja AK, Allen JE, Maizels RM. The abundant larval transcript-1 and -2 genes of Brugia malayi encode stage-specific candidate vaccine antigens for filariasis. Infect Immun. 2000;68:4174–4179. doi: 10.1128/IAI.68.7.4174-4179.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham SP, Lustigman S, Trees AJ, Bianco AE. O. volvulus: comparative analysis of antibody responses to recombinant antigens in two animal models of onchocerciasis. Exp Parasitol. 2000;94:158–162. doi: 10.1006/expr.2000.4487. [DOI] [PubMed] [Google Scholar]
- Wu Y, Egerton G, Underwood AP, Sakuda S, Bianco AE. Expression and secretion of a larval-specific chitinase (family 18 glycosyl hydrolase) by the infective stages of the parasitic nematode, O. volvulus. J Biol Chem. 2001;276:42557–42564. doi: 10.1074/jbc.M103479200. [DOI] [PubMed] [Google Scholar]
- Wu Y, Adam R, Williams SA, Bianco AE. Chitinase genes expressed by infective larvae of the filarial nematodes, Acanthocheilonema viteae and O. volvulus. Mol Biochem Parasitol. 1996;75:207–209. doi: 10.1016/0166-6851(95)02529-4. [DOI] [PubMed] [Google Scholar]
- Adam R, Kaltmann B, Rudin W, Friedrich T, Marti T, Lucius R. Identification of chitinase as the immunodominant filarial antigen recognized by sera of vaccinated rodents. J Biol Chem. 1996;271:1441–1447. doi: 10.1074/jbc.271.35.21490. [DOI] [PubMed] [Google Scholar]
- Canlas M, Wadee A, Lamontagne L, Piessens WF. A monoclonal antibody to surface antigens on microfilariae of Brugia malayi reduces microfilaremia in infected jirds. Am J Trop Med Hyg. 1984;33:420–424. doi: 10.4269/ajtmh.1984.33.420. [DOI] [PubMed] [Google Scholar]
- Harrison R, Wu Y, Egerton G, Bianco AE. DNA immunization with O. volvulus chitinase induces protection to a challenge infection with L3 larvae in mice. Vaccine. 1999;18:647–655. doi: 10.1016/S0264-410X(99)00274-1. [DOI] [PubMed] [Google Scholar]
- Wang SH, Zheng HJ, Dissanayake S, Cheng WF, Tao ZH, Lin SZ, Piessens WF. Evaluation of recombinant chitinase and SXP1 antigens as antimicrofilarial vaccines. Am J Trop Med Hyg. 1997;56:474–481. doi: 10.4269/ajtmh.1997.56.474. [DOI] [PubMed] [Google Scholar]
- Clos J, Brandau S. PJC20 and pJC40 two high-copy-number vectors for T7 RNA polymerase-dependent expression of recombinant genes in Escherichia coli. Protein Expr Purif. 1994;5:133–137. doi: 10.1006/prep.1994.1020. [DOI] [PubMed] [Google Scholar]
- Cooper PJ, Espinel I, Paredes W, Guderian RH, Nutman TB. Impaired tetanus-specific cellular and humoral responses following tetanus vaccination in human onchocerciasis: a possible role for interleukin-10. J Infect Dis. 1998;178:1133–1138. doi: 10.1086/515661. [DOI] [PubMed] [Google Scholar]
- Jenkins RE, Taylor MJ, Gilvary NJ, Bianco AE. Tropomyosin implicated in host protective responses to microfilariae in onchocerciasis. Proc Natl Acad Sci U S A. 1998;95:7550–7555. doi: 10.1073/pnas.95.13.7550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lizotte-Waniewski M, Tawe W, Guiliano DB, Lu W, Liu J, Williams SA, Lustigman S. Identification of potential vaccine and drug target candidates by expressed sequence tag analysis and immunoscreening of O. volvulus larval cDNA libraries. Infect Immun. 2000;68:3491–3501. doi: 10.1128/IAI.68.6.3491-3501.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacDonald AJ, Turaga PS, Harmon-Brown C, Tierney TJ, Bennett KE, McCarthy MC, Simonek SC, Enyong PA, Moukatte DW, Lustigman S. Differential cytokine and antibody responses to adult and larval stages of O. volvulus consistent with the development of concomitant immunity. Infect Immun. 2002;70:2796–2804. doi: 10.1128/IAI.70.6.2796-2804.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Y, Egerton G, Ball A, Tanguay RM, Bianco AE. Characterization of the heat-shock protein 60 chaperonin from O. volvulus. Mol Biochem Parasitol. 2000;107:155–168. doi: 10.1016/S0166-6851(99)00227-3. [DOI] [PubMed] [Google Scholar]
- Stewart GR, Elson L, Araujo E, Guderian R, Nutman TB, Bradley JE. Isotype-specific characterization of antibody responses to O. volvulus in putatively immune individuals. Parasite Immunol. 1995;17:371–380. doi: 10.1111/j.1365-3024.1995.tb00904.x. [DOI] [PubMed] [Google Scholar]
- Khalife J, Dunne DW, Richardson BA, Mazza G, Thorne KJ, Capron A, Butterworth AE. Functional role of human IgG subclasses in eosinophil-mediated killing of schistosomula of Schistosoma mansoni. J Immunol. 1989;142:4422–4427. [PubMed] [Google Scholar]
- Bradley JE, Elson L, Tree TI, Stewart G, Guderian R, Calvopina M, Paredes W, Araujo E, Nutman TB. Resistance to O. volvulus: differential cellular and humoral responses to a recombinant antigen, OvMBP20/11. J Infect Dis. 1995;172:831–837. doi: 10.1093/infdis/172.3.831. [DOI] [PubMed] [Google Scholar]
- Boyer AE, Tsang VC, Eberhard ML, Zea-Flores G, Hightower A, Pilcher JB, Zea-Flores R, Zhou W, Reimer CB. Guatemalan human onchocerciasis. II. Evidence for IgG3 involvement in acquired immunity to O. volvulus and identification of possible immune-associated antigens. J Immunol. 1991;146:4001–4010. [PubMed] [Google Scholar]
- Lange AM, Yutanawiboonchai W, Lok JB, Trpis M, Abraham D. Induction of protective immunity against larval O. volvulus in a mouse model. Am J Trop Med Hyg. 1993;49:783–788. doi: 10.4269/ajtmh.1993.49.783. [DOI] [PubMed] [Google Scholar]
- Mohanty MC, Satapathy AK, Sahoo PK, Ravindran B. Human bancroftian filariasis – a role for antibodies to parasite carbohydrates. Clin Exp Immunol. 2001;124:54–61. doi: 10.1046/j.1365-2249.2001.01484.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuranda MJ, Robbins PW. Chitinase is required for cell separation during growth of Saccharomyces cerevisiae. J Biol Chem. 1991;266:19758–19767. [PubMed] [Google Scholar]
- Watanabe T, Suzuki K, Oyanagi W, Ohnishi K, Tanaka H. Gene cloning of chitinase A1 from Bacillus circulans WL-12 revealed its evolutionary relationship to Serratia chitinase and to the type III homology units of fibronectin. J Biol Chem. 1990;265:15659–65. [PubMed] [Google Scholar]
- Fuhrman JA, Lane WS, Smith RF, Piessens WF, Perler FB. Transmission-blocking antibodies recognize microfilarial chitinase in brugian lymphatic filariasis. Proc Natl Acad Sci U S A. 1992;89:1548–52. doi: 10.1073/pnas.89.5.1548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elson LH, Calvopina M, Paredes W, Araujo E, Bradley JE, Guderian RH, Nutman TB. Immunity to onchocerciasis: putative immune persons produce a Th1-like response to O. volvulus. J Infect Dis. 1995;171:652–658. doi: 10.1093/infdis/171.3.652. [DOI] [PubMed] [Google Scholar]
- Elson LH, Guderian RH, Araujo E, Bradley JE, Days A, Nutman TB. Immunity to onchocerciasis: identification of a putatively immune population in a hyperendemic area of Ecuador. J Infect Dis. 1994;169:588–594. doi: 10.1093/infdis/169.3.588. [DOI] [PubMed] [Google Scholar]