

Abstract
Dramatic increases in childhood obesity necessitate a more complete understanding of neural mechanisms of hunger and satiation in pediatric populations. In this study, normal weight children and adolescents underwent functional magnetic resonance imaging (fMRI) scanning before and after eating a meal. Participants showed increased activation to visual food stimuli in the amygdala, medial frontal/orbitofrontal cortex, and insula in the pre-meal condition; no regions of interest responded in the post-meal condition. These results closely parallel previous findings in adults. In addition, we found evidence for habituation to food stimuli in the amygdala within the pre-meal session. These findings provide evidence that normal patterns of neural activity related to food motivation begin in childhood. Results have implications for obese children and adults, who may have abnormal hunger and satiation mechanisms.
Keywords: Amygdala, Obesity, fMRI, Food motivation, Pediatric
Introduction
Nearly 20% of the US adult population is clinically obese (body mass index [BMI] ≥30 kg/m2; Mokdad et al., 2000), a condition associated with increased morbidity, mortality, and health care costs (Allison et al., 1999; Colditz, 1999). The roots of adult obesity are likely established in childhood and adolescence, and increases in obesity prevalence have been documented that are comparable to those found in adults (Strauss and Pollack, 2001). Recent estimates of childhood overweight (BMI greater than 95 percentile for age and sex) in the United States range from 12 to 22% of the population (Strauss and Pollack, 2001).
Although several factors contribute to weight gain, consistent eating in excess of daily energy requirements plays a primary etiological role across the lifespan (Nielsen et al., 2002). Neural mechanisms are central to the regulation of these motivationally mediated behaviors (Tataranni and DelParigi, 2003); thus, the increasing concern surrounding the dramatic rise of obesity (Mokdad et al., 1999; 2000) has led to research aimed at understanding the neural substrates of appetitive function in humans (Tataranni and DelParigi, 2003; Wang et al., 2004). The role of the hypothalamus in food motivation is well established in literature on non-human primates (Rolls et al., 1976) as well as humans (Rolls, 1981; Stellar, 1954). More recently, however, limbic–frontal connectivity has become increasingly implicated in normal food motivation (Zald et al., 1998), as well as abnormal food intake and obesity (Tataranni and DelParigi, 2003). Previous human and animal studies suggest coordinated involvement of these areas in processing of emotional stimuli and representation of reward (Baxter and Murray, 2002; Gottfried et al., 2003; Kringelbach and Rolls, 2004; Rolls, 2004; Whalen, 1998). Specifying these networks has become an aim of brain activation studies using PET and fMRI.
Previous functional neuroimaging studies of food motivation have utilized a broad array of paradigms, including passive viewing of food images, conditioned responses to olfactory and gustatory stimuli, and inclusion of pre- and post-meal scans for comparison of motivational state within subjects. Across these studies, the most consistent findings include activation of orbitofrontal cortex (OFC), medial frontal cortex (MFC), amygdala, hippocampal formation, and insula (Gordon et al., 2000; Hinton et al., 2004; Killgore et al., 2003; LaBar et al., 2001; Morris and Dolan, 2001; O’Doherty et al., 2002; Small et al., 2001; Tataranni et al., 1999). For example, LaBar et al. (2001) scanned normal weight adults while they passively viewed images of food both before and after eating. They found differential responses to food stimuli in the amygdala, hippocampal formation, and fusiform gyrus during hunger, which resolved after eating.
Thus, there is consistent evidence from studies of food motivation in healthy weight individuals that limbic–frontal neural networks underlie normal processes of food motivation and regulation of food intake. It has been hypothesized that these neural systems also play an important role in controlling hyperphagia in obese individuals (e.g., Tataranni and DelParigi, 2003). Though still in its infancy, neuroimaging research on obese and overweight adults has shown differential patterns of neural activity in obese compared to lean individuals, including abnormal functioning in the hypothalamus (Matsuda et al., 1999), insula (DelParigi et al., 2003; Gautier et al., 2000), hippocampus (DelParigi et al., 2003; Gautier et al., 2000), and OFC (Gautier et al., 2000). These findings offer evidence for discrepancies in neural circuits involved in food motivation between obese and lean adults.
Although neural mechanisms of hunger and satiation have been examined in neuroimaging studies of both normal and obese adults, to our knowledge, no studies have been published in children and adolescents. This is of note since neuroimaging findings in children and adolescents are not always consistent with those in adults (Davidson et al., 2003). In particular, the amygdala and OFC show continued development into adulthood (Durston et al., 2001; Happaney et al., 2004) and may respond differently to appetitive stimuli based on age. Thus, investigation of the neural mechanisms underlying hunger and satiation in children is necessary for a complete understanding of normal and abnormal human appetitive function across the lifespan. The current study explores changes in neural response to food stimuli before and after eating in healthy weight children and adolescents, with the goal of identifying the normal neural circuitry of these processes for future research on overweight children and adolescents. Based on findings in previous studies on adults, we hypothesized greater activation in response to food stimuli in the OFC, MFC, insula, amygdala, and hippocampal formation during a pre-meal state compared to a post-meal state.
Materials and methods
Subjects
Written informed consent was obtained for the 5 female and 4 male children and adolescents who participated in this study. Participants ranged in age from 10 to 17 [mean (±SD) = 13.4 (±2.8)]. Participants were within the healthy weight-for-height ratio (BMI) for age and sex as determined by growth curve charts from the Center’s for Disease Control [mean (±SD) = 18.5 (±2.4)]. All individuals were free from psychiatric diagnoses and neurological illnesses (based on parental interview) and had normal vision. Eight individuals were right-handed. This study was approved by the Human Subjects Committee (IRB) at the University of Kansas Medical Center.
fMRI acquisition
Scanning was performed on a 3T head-only Siemens Allegra scanner (Siemens, Erlangen, Germany) fitted with a quadrature head coil. Participants’ heads were immobilized with head cushions. T1-weighted anatomic images were acquired with a 3D SPGR sequence (TR/TE = 23/4 ms, flip angle = 8°, FOV = 256 mm, matrix = 256 × 192, slice thickness = 1 mm). Single shot gradient echo fMRI scans were acquired in 43 contiguous coronal slices (repetition time/echo time [TR/TE] = 3000/40 ms, flip angle = 90°, field of view [FOV] = 192 mm, matrix = 64 × 64, slice thickness = 3 mm (0.5 mm skip), in-plane resolution = 3 × 3 mm, 130 data points). One anatomical and two functional sequences were run in each scanning session (i.e., pre-meal and post-meal).
Experimental paradigm
The experimental paradigm was based closely on LaBar et al. (2001). Participants viewed pictures of food, animals, and Gaussian-blurred low-level baseline control images during two scanning sessions: one after fasting for 4 h (pre-meal) and one immediately after eating a small uniform meal (post-meal) that was standardized for total number of calories [Kcal = 500], as well as macro- and micronutrient content. Previous studies examining the effect of satiation on brain activity have included a longer fasting period, typically 8 h, and utilized meals designed to fully sate participants (LaBar et al., 2001; Morris and Dolan, 2001). The current study implemented a 4 h fast and a meal standardized to provide approximately 500 Kcal. Our goal was to design a paradigm that accurately reflected the normal hunger and eating cycles. The order of sessions (pre-meal, post-meal) was counterbalanced across subjects so that approximately half the group (n = 5) started with the pre-meal session and half (n = 4) started with the post-meal session.
Activation paradigm
Stimuli of two categories (food and blurred baseline control images) were obtained from LaBar et al. (2001). Due to the age of the participants in this study, the comparison stimuli group of animals (rather than tools, as used by LaBar et al.) was chosen to keep participants attentive to the task and to control for general familiarity. All images for the animal category were obtained from professional stock CD-ROMs and matched to food and blurred control images on brightness, resolution, and size. In addition, by applying a Gaussian kernel to a subset of the animal images (so that the objects were not identifiable), approximately 150 new blurred baseline control images were obtained. To the greatest degree possible, animals that were reminiscent of food (i.e., fish) were removed from the stimuli pool to prevent the possible confusion between animal/food categorizations. Blurred objects were included as a low-level baseline comparison. All images were presented one time only to each subject.
Each functional scan involved three repetitions of each block of each stimulus condition type (i.e., food, animal), alternated between blocks of blurred images. Visual stimuli were projected through 3D limited view goggles (Resonance Technology, Inc., Northridge, California) connected to the stimuli-generating computer program (NeuroSTIM, Neuroscan, El Paso, TX). Stimulus presentation time was 2.5 s, with an interstimulus interval (ISI) of 0.5 s. Within each of the two functional scans, there was a total of 13 blocks of stimuli presentation; within each block, 10 images were presented. The order of category presentation was counterbalanced across subjects.
To ensure that participants were attending to the stimuli being presented, they were instructed to remember images for a memory test following the scanning session. From each of the food and animal groups, approximately 50% of the images used in the scanning session (30 images) were chosen for recall (old) and interspersed with 15 novel distracter images from the same category (new). Participants completed a recognition memory task outside the scanner, immediately following each scanning session. Participants were instructed to press one key if they had seen the image in the scanner (old) and another button if they had not seen the image (new).
Behavioral data analysis
Due to our small sample size (n = 9), we used A′ statistics, a non-parametric equivalent of d′ (Pollack and Norman, 1964). Mean A′ values were computed for food and non-food items in the pre- and post-meal condition. Wilcoxon sign rank tests (a non-parametric statistic) were used to compare observed performance to chance (i.e., 50% correct). In addition, in order to assess the effect of motivational state on memory for food images, A′ statistics on food and non-food items were compared across conditions on median proportion of correctly recognized items, using Wilcoxon signed rank tests (food: pre- vs. post-meal; non-food: pre- vs. post-meal).
fMRI data analysis
fMRI data were analyzed using the BrainVoyager 2000 (version 4.9.6) statistical package (Brain Innovation, Maastricht, Nether-lands). Preprocessing steps included trilinear 3D motion correction, sinc-interpolated slice scan time correction, 3D spatial smoothing with 4-mm Gaussian filter, and high pass filter temporal smoothing. Functional images were realigned to the anatomic images obtained within each session and normalized to the BrainVoyager template image, which conforms to the space defined by the Talairach and Tournoux’s stereotaxic atlas (Talairach and Tournoux, 1988). Motion in any run of more than 4 mm along any axis (x, y, or z) resulted in the discard of that run. Only one run was discarded due to excessive motion. Although some have questioned the use of transformation to stereotaxic space for children’s brains in group studies of structure and function (Gaillard et al., 2001), recent findings indicate that differences between age groups are beneath the resolution provided by current imaging technology (Burgund et al., 2002). Furthermore, functional analysis maps projected onto averaged anatomical images appear not to produce false variance between age groups (Burgund et al., 2002).
Activation maps were analyzed using statistical parametric methods (Friston et al., 1995) as implemented within the BrainVoyager 2000 software. Statistical contrasts were conducted using multiple regression analysis with the general linear model (GLM), allowing for multiple predictors to be built into the model. Regressors representing the experimental conditions of interest were modeled with a hemodynamic response filter and entered into the multiple regression analysis using a fixed-effects model. Contrasts between conditions of interest were assessed with t statistics. Statistical parametric maps were overlaid on three-dimensional renderings of an averaged-group structural scans.
Analysis began with a contrast of blurred control (baseline) vs. food (F) + non-food (NF) conditions within each session to assess whether the low-level baseline produced significant activation. No significant regions of change were noted. Secondly, whole-brain analyses of contrasts between stimuli conditions (i.e., F vs. NF) were conducted within each state (pre-meal and post-meal). Based on previous research in adults, a priori regions of interest (ROIs) included OFC, MFC, insula, amygdala, and hippocampal formation (hippocampus and parahippocampal gyrus). We defined ‘‘orbitofrontal cortex’’ based on Talairach coordinates mapped onto the Mai et al. (2003) atlas and corresponding to one of the orbital gyri. Medial frontal cortex was defined as frontal activations superior to these gyri and in the midline (±8 mm). Specifically, we predicted greater activation to food pictures, compared to non-food pictures and blurred baseline image, in these regions in the pre-meal condition. In order to identify potential regions nonspecifically implicated in food image processing, we also examined the simple effect of stimulus type (F vs. NF) collapsed across motivational states. A priori ROIs were not specified for this set of exploratory analyses.
Furthermore, an interaction analysis (motivational state × condition) was performed to examine the response to food (vs. baseline) between the pre-meal and post-meal conditions. In addition, in order to determine the difference between hunger-modulation response to food stimuli versus non-food stimuli, a 2 (food vs. non-food) × 2 (pre- vs. post-meal) interaction analysis was also performed. A priori ROIs for both interaction analyses included the same regions as noted for the F vs. NF contrasts. For all analyses, voxels in regions of interest predicted in each contrast were considered significant if the activation survived a statistical threshold of P < 0.05 (corrected for multiple comparisons for whole brain) and had a minimum cluster size of 3 contiguous voxels. Other areas were considered significant if they exceeded a threshold of P < 0.01 (corrected for whole brain) and had a minimum cluster size of 6 contiguous voxels.
Finally, follow-up analyses of some a priori ROIs found to be activated in the primary analyses were conducted. For ROI analyses, mean percent signal change values (food compared to blurred baseline, averaged across each block) in the maximum pixel within each region for each individual were exported to SPSS 11.0.2 for Macintosh (Statistical Package for the Social Sciences, Chicago, IL) for comparison between sessions using an analysis of variance (ANOVA). For anatomical verification of a priori region activation, Talairach coordinates for each maximum pixel were determined and confirmed by examination of an additional anatomical brain atlas (Mai et al., 2003).
Results
Behavioral data
Memory performance scores for both the pre-meal and post-meal sessions were higher than chance (pre-meal: Z = −2.67, P < 0.01; post-meal: Z = −2.67, P < 0.01, Wilcoxon signed rank test). Memory for food items was higher in the pre-meal condition [mean percentage correct (±SD) = 77.78% (±7.36); A′ = 0.85)] than in the post-meal condition [mean percentage (±SD) = 65.92% (±10.55); A′ = 0.83; Z = −2.07, P < 0.05, Wilcoxon signed rank test)], while memory for non-food items did not differ across feeding states [pre-meal: mean percentage correct (±SD) = 81.73% (±10.04); A′ = 0.88; post-meal: mean percentage correct (±SD) = 79.51%, (±9.86); A′ = 0.87; Z = −0.25, n.s., Wilcoxon signed rank test]. fMRI data
Simple effect of stimulus type
Assessing effect of stimulus type across motivational states, food elicited greater activation compared to non-food in the insula (x, y, z = −35, 3, −7), inferior frontal cortex (−23, 24, −4; BA 11/47), ventrolateral prefrontal cortex (−38, 33, 20; BA 9/10), parahippocampal gyrus (28, −27, −19; BA 36), and bilateral fusiform gyrus (28, −48, −10; BA 37; −29, −51, −10; BA 37). Non-food images, alternatively, generated greater activation bilaterally in the cerebellum.
Food vs. non-food contrasts
Pre-meal
The (Food–Non-food) direct contrast yielded greater activations to food before eating in a priori regions including the bilateral medial OFC, left lateral OFC, and bilateral MFC (see Table 1 for t values and Talairach coordinates). Post hoc regions are also listed in Table 1.
Table 1.
Regions reaching significance for the contrasts between food, non-food, and baseline stimuli categories during pre-meal and post-meal states ( P < 0.01, corrected for whole brain)
|
Coordinates |
||||||
|---|---|---|---|---|---|---|
| Contrast | Region | Brodmann’s area | x | y | z | t |
| Food > non-food | ||||||
| Pre-meal | Medial OFC | 10 | 4 | 54 | −4 | 5.66 |
| 10 | −2 | 57 | −4 | 6.94 | ||
| LateralOFC | 11 | −29 | 33 | −4 | 8.16 | |
| MFC | 10 | −8 | 61 | 20 | 5.79 | |
| Superior parietal cortex | 7 | 22 | −66 | 56 | 6.49 | |
| Cerebellum/fusiform | 19/37 | 22 | −54 | −13 | 9.32 | |
| 19/37 | −23 | −51 | −13 | 8.38 | ||
| Post-meal | Superior temporal gyrus | 21/22 | 58 | −12 | 2 | 6.17 |
| Fusiform gyrus | 37 | 25 | −54 | −10 | 6.30 | |
| 37 | −29 | −54 | −7 | 7.66 | ||
| Stimulus × motivational state interaction | ||||||
| Amygdala | – | 14 | −2 | −18 | 6.62 | |
| – | 17 | −11 | −15 | 5.64 | ||
| Medial OFC | 10 | 2 | 49 | 3 | 7.58 | |
| Lateral OFC | 47 | −35 | 33 | −7 | 6.15 | |
| 11 | 25 | 33 | −4 | 5.67 | ||
| MFC | 9 | −8 | 57 | 26 | 9.07 | |
| Insula | – | −38 | 21 | −1 | 7.48 | |
| Basal operculum | 47 | 40 | 21 | −10 | 6.60 | |
| 47 | −38 | 21 | −10 | 6.61 | ||
| Parahippocampal gyrus | 28 | 16 | −12 | −16 | 5.64 | |
| Cingulate gyrus | 23 | 1 | −15 | 32 | 5.68 | |
| Fusiform gyrus | 37 | −41 | −60 | −13 | 9.25 | |
| Inferior frontal gyrus | 44 | 49 | 18 | 8 | 6.34 | |
| Superior frontal gyrus | 6 | 10 | 9 | 65 | 8.03 | |
| 8 | −20 | 45 | 38 | 7.12 | ||
| Inferior temporal gyrus | 37 | 52 | −60 | −7 | 6.28 | |
| Globus pallidus | – | 10 | 3 | −4 | 5.67 | |
| Postcentral gyrus | 4 | −14 | −33 | 56 | 7.15 | |
| Precentral gyrus | 6 | 52 | 0 | 41 | 7.16 | |
| 6 | −5 | −27 | 68 | 6.73 | ||
| Supramarginal gyrus | 40 | 52 | −36 | 50 | 5.55 | |
| 40 | −47 | −39 | 44 | 5.74 | ||
| Cerebellum/fusiform | 37 | 19 | −45 | −16 | 7.36 | |
| 36 | 34 | −36 | −25 | 8.59 | ||
| Non-food > food | ||||||
| Pre-meal | Lateral temporal gyrus | 37 | 40 | −60 | 5 | −7.40 |
| Fusiform | 37 | 37 | −42 | −16 | −7.13 | |
| Cerebellum/fusiform | – | −38 | −48 | −19 | −9.90 | |
| Post-meal | Superior frontal gyrus | 6 | 1 | 18 | 54 | −5.76 |
| 9 | −17 | 61 | 29 | −6.53 | ||
| Cerebellum/fusiform | 37 | 34 | −42 | −22 | −8.34 | |
| – | −38 | −48 | −16 | −10.68 | ||
Post-meal
After eating, food stimuli elicited greater responses relative to non-food in several post hoc regions (see Table 1).
Stimulus × motivational state interaction contrast
The interaction contrasting pre- and post-meal sessions revealed neural responses to food that differed significantly depending on motivational state. Specifically, regions were identified in which the Food vs. Baseline contrast was greater in the pre-meal condition than in the post-meal condition. A priori regions responding to food images to a greater extent before eating than after eating included the right amygdala, right medial OFC, bilateral lateral OFC, left MFC, bilateral basal operculum/insula, right parahippocampal gyrus, right cingulate gyrus, and left fusiform gyrus (see Table 1). Fig. 1 depicts areas of activation in the amygdala, MFC, and MOFC. Post hoc regions are listed in Table 1.
Fig. 1.
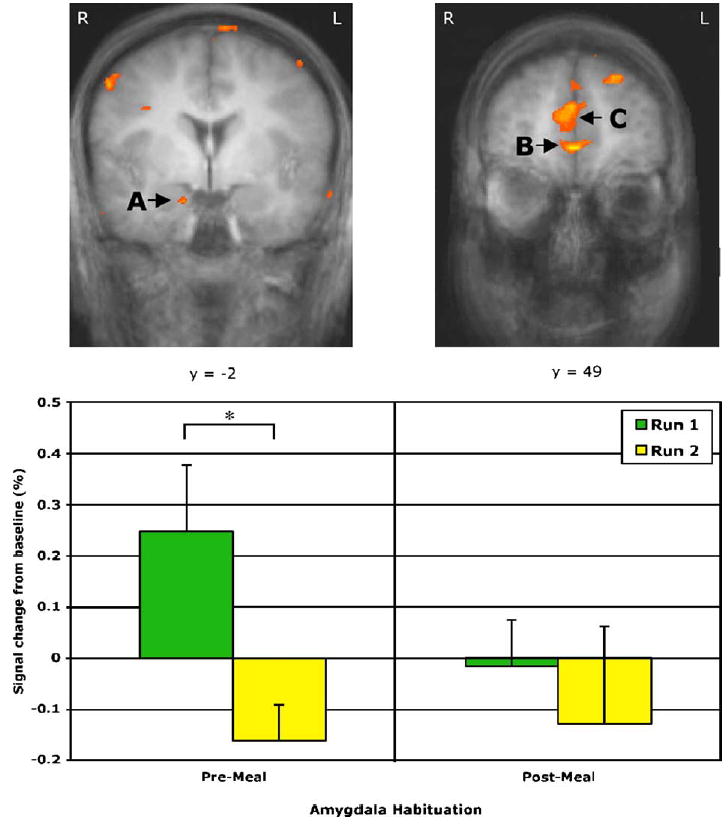
Statistical contrast maps highlighting results from the Stimulus × Motivational State Interaction contrast, coregistered with average structural MRI data from participants. Maps are presented in the coronal perspective. Regions of activation represent areas in which the Food vs. Baseline contrast varies with motivational state (greater pre-meal). Activations are noted in (A) amygdala, (B) medial orbitofrontal cortex (MOFC), and (C) medial frontal cortex (MFC). Below, the bar graph represents habituation to food stimuli within the amygdala at its maximum voxel [located at 14, −2, −18] separately for each run within the pre-meal and post-meal states. *P < 0.05.
Further analyses were conducted to examine the interaction between stimulus type (Food vs. Non-food) and motivational state (Pre-meal vs. Post-meal), which yielded activations in the superior frontal gyrus [x, y, z = −8, 24, 63, a region corresponding to Brodmann’s area (BA) 8] and in the fusiform gyrus (25, −45, −7; BA 37) at the P < 0.01 (corrected) level.
Region of interest analysis
In order to investigate further the nature of the activations seen in clusters within the amygdala and medial OFC (MOFC), detailed ROI analyses were conducted on these regions, as identified by the interaction analysis. A 2 (amygdala vs. MOFC) × 2 (pre-meal vs. post-meal) repeated-measures ANOVA on percent signal change revealed a main effect for motivational state (pre- vs. post-meal), with both regions exhibiting greater activation pre-meal compared to post-meal [ F(1,16) = 16.62, P < 0.01] but no main effect for region of interest [ F(1,16) = 0.21, n.s.].
Due to the likelihood of habituation within the amygdala over time (Wright et al., 2001), mean percent signal change values within this region were examined further. Within the amygdala, significant habituation (as defined by greater activation in run 1 vs. run 2) was exhibited during the pre-meal state [t(1,15) = 2.65, P < 0.05], but not during the post-meal session [t(1,16) = 0.52, n.s.] (see Fig. 1). Thus, significant activation in the amygdala occurred only in the first run of the pre-meal condition.
Discussion
Results indicate that patterns of neural activation to food stimuli are modulated by motivational state in children and adolescents of healthy weight. These findings closely replicate previous imaging studies in adult populations (e.g., LaBar et al., 2001), showing activation to food stimuli in limbic and paralimbic regions during pre-meal scans. Results from this study reveal specific neural patterns of activity during hunger, including increases to food in the OFC, amygdala, MFC, insula, parahippocampal gyrus, cingulate gyrus, and fusiform gyrus, and habituation to presentation of food stimuli explicitly in the amygdala. In addition, our stimulus type contrast suggests greater activation to food compared to non-food images in the OFC and MFC before eating, implying these areas contribute specifically to the processing of food stimuli in a state of high incentive. Finally, our data indicate that these functional connections are established by late childhood and adolescence.
Current and previous work specifically point to the OFC/MFC and amygdala in processes underlying food motivation. Both the amygdala and OFC have direct connections with the medial prefrontal cortex (Ongur et al., 2003; Rolls, 2004). The OFC is believed to play a central role in assessing the reward value of taste (Rolls, 2004). Due to the particular salience of food stimuli during a fasting state, it is not surprising that this region is also recruited in response to visual food stimuli. Increased activity in the OFC and
MFC in response to food stimuli is also consistent with recent findings related to food motivation in humans (Morris and Dolan, 2001; O’Doherty et al., 2002), which implicate these neural territories in evaluation of positive reinforcers (such as food). In addition, the MFC appears to mediate communication between the hypothalamus and the highly reward-based, affective OFC (Pucak et al., 1996). Ongur and colleagues theorize that some medial aspects of the MFC modulate visceral responses to affective stimuli in a highly motivational context (Ongur et al., 2003). Our results indicating increases to food stimuli in the MFC before eating support this theory.
The amygdala, which serves a complementary role in evaluating emotional valence of salient stimuli (Davidson, 2004), has been implicated in processing of food stimuli (Rolls, 2004) and in the search for and selection of potential food sources (Baxter and Murray, 2002). Although our results suggest that the patterns of response within the amygdala (pre-meal increase, post-meal normalization) parallel those seen in OFC/MFC, closer analysis indicates that increased activity is confined to the first run of the pre-meal condition. This finding is consistent with habituation effects to repeatedly presented stimuli previously reported in the amygdala (Wright et al., 2001) and likely reflects an initial response, with drop-off in activation after the salience of the stimuli has been assessed (Whalen, 1998). Thus, following an initial assessment of food stimuli salience, the amygdala may no longer be crucial to the overall process of monitoring the body’s motivational status.
Due to the well-established connection between binge-eating and emotional regulation, it is possible that the MFC/OFC, amygdala, and hippocampus may be involved in the development of abnormal neural signals leading to overeating during a heightened emotional state (Will et al., 2004), although it is difficult to rule out the possibility that the amygdala activation may also play a role in basic learning processes associated with food availability stimuli. In addition to emotional processing, these networks likely contribute to cognitive aspects of food stimuli processing. Hunger-modulated activation in the MFC/OFC, amyg-dala, and hippocampus in our young sample is consistent with research implicating these structures in encoding of food stimuli during the hungry state among adults (Morris and Dolan, 2001). In the context of food processing and hunger, interaction between memory and limbic functions of the hippocampus, in particular, may indicate recruitment of memory processes related to highly arousing stimuli (Morris and Dolan, 2001; Tataranni et al., 1999; Zald, 2003). Our behavioral results also tentatively support this evidence of enhanced memory for food during the hunger state, with increased memory for food stimuli pre-meal compared to post-meal. Cognitive processes such as memory may ultimately interplay with affective signals—hypersensitive memory for food, in the context of emotional dysregulation, may lead to stronger associations between food and pleasure and thereby contribute to overeating.
The insula contains the primary taste cortex (Zald, 2003) with direct connections with the amygdala and OFC. Our results suggest greater increases in insular activity in response to food stimuli before eating, which is consistent with previous findings in adults (Hinton et al., 2004; Morris and Dolan, 2001; Small et al., 2001); however, we also found a main effect for food presentation in another region of the insula, which is consistent with LaBar et al. (2001), who reported insula activation to food stimuli both before and after eating. Further studies are needed to elucidate the role of the insula in motivationally modulated response to food-related stimuli. Increased responses in the ACC have been documented in response to aversive gustatory stimuli (Zald et al., 1998), anticipatory taste during hunger (Gautier et al., 1999), and visual images of food during hunger (Tataranni et al., 1999). Our finding of increased activity in the anterior cingulate cortex in response to food stimuli during hunger is consistent with these reports and may be related to the monitoring of unpleasant internal sensations associated with hunger (Craig et al., 1996; Tataranni et al., 1999). Finally, food-related activation in the fusiform gyrus during hunger replicates previous findings (LaBar et al., 2001). Given the role of the fusiform in visual processing of both natural and manufactured objects (Joseph, 2001), this result presents evidence that the fusiform gyrus contributes to the processing of visual attributes specific to food.
Beyond basic replication of previous findings on food motivation in adults, our results extend the understanding of developmental aspects of neural mechanisms of appetitive function. This is the first study to examine food motivation in typically developing children and adolescents at a neural systems level. While adjustments in the body’s nutritional requirements occur across the lifespan (Troiano et al., 2000), these changes do not appear to be similarly reflected in changes in the neural circuitry of food motivation. Recent reports indicate similar establishment of normal functioning in other reward-related neural networks in children and adolescents (May et al., 2004). However, differences in neural functioning between childhood and adult processing have been noted in relation to high-order tasks such as word generation (Brown et al., 2005). It is possible that presence of normal developmental changes in brain functioning emerges in studies focused on processes that also show behavioral changes with age, while more basic processing of food (which could be considered a more innate or primitive function) is established early on and undergoes little variation in terms of either behavioral or neural functioning.
Furthermore, our results provide a basis for investigation of children and adolescents with obesity. Previous findings in adults with obesity indicate greater increases in the PFC and greater decreases in the OFC, amygdala, hippocampus, insula, and temporal lobe during satiation compared to lean individuals (DelParigi et al., 2003; Gautier et al., 2000). While these studies examining food motivation in overweight adults aid in the overall understanding of neural mechanisms of overeating and obesity, none have been performed in groups of children and adolescents. Through future studies on the comparison of adults and children with obesity, we might better understand the development of this condition and the relationship between early-onset of obesity and neural functioning.
Finally, our understanding of the typical neural response to food motivation may also inform our view of disorders in which food intake is abnormal. Individuals with eating disorders (i.e., anorexia nervosa and bulimia nervosa), who typically develop symptoms during adolescence (APA, 2000), maintain abnormal thought patterns involving food. Recent work demonstrates parallel dysfunction in neural mechanisms of food processing and motivation in this population. In general, neuroimaging studies suggest hyperactivation in regions involved in modulation of visceral responses to salient stimuli during motivation in chronically food-deprived individuals. This research provides evidence for reduction in activity during rest in prefrontal regions and the anterior cingulate cortex in eating disorders (Delvenne et al., 1999; Naruo et al., 2001; Takano et al., 2001; Wu et al., 1990), with a specific heightened response to presentation of food in the MPFC during hunger (Uher et al., 2004). Our results in normal food motivation also demonstrate a role for the MPFC, a region which showed large increases in response to food before eating in normal weight children and adolescents. Comparison of our results with those of Uher et al. may imply a role for the MPFC in the development of eating disorders. A complete understanding of MPFC dysfunction in eating disorders and its variance from typical function awaits further investigations comparing children and adolescents with and without eating disorders.
While the implications of the current study extend the understanding of normal appetitive functioning, some limitations must be acknowledged. First, our small sample size and use of a fixed-effects analysis preclude generalization to the population level of our age group. We included individuals within a wide age range (10–17) who may represent different developmental stages. Although we examined relationships between age and percent signal change using data in this study, we found no evidence for significant correlations in any of our ROIs. Future studies should be conducted in larger groups so that the effects of age can be more specifically addressed. Furthermore, this initial study utilized a blocked design, and we do not know if findings will extend to event-related designs. The blocked stimulus presentation may have also contributed to our observations of habituation in the amygdala across functional runs. Finally, given the possibility of susceptibility artifact in and around the MOFC (at a field strength of 3 T), we cannot conclude that other more ventral regions of MOFC were not also involved in food motivation.
In summary, normal weight children and adolescents showed increased activation to food images in the amygdala, medial frontal/orbitofrontal cortex, and insula in the pre-meal condition, which resolved following ingestion of a small meal. Our results indicate that neural responses to hunger and satiation in healthy weight children are similar to those previously documented in healthy weight adults. These findings provide evidence that normal patterns of neural activity underlying food motivation begin in childhood and closely parallel responses in adults. While obesity is a multifaceted condition, eating behavior is strongly influenced by neural processes, the roots of which begin in childhood. Current results, therefore, provide insights into the potential neural mechanisms contributing to the increasing problem of obesity.
Acknowledgments
This study was supported by grants from NICHD (HD41672) and the Hall Family Foundation. The authors are grateful to Allan Schmitt and Muriel Williams for technical assistance and Stacey Ward for help in project coordination. We would also like to thank Dr. Sandra Hall for statistical consultation and Joshua Powell for assistance in fMRI data analyses.
References
- Allison DB, Fontaine KR, Manson JE, Stevens J, VanItallie TB. Annual deaths attributable to obesity in the United States. JAMA. 1999;282:1530–1538. doi: 10.1001/jama.282.16.1530. [DOI] [PubMed] [Google Scholar]
- American Psychiatric Association, 2000. Diagnostic and Statistical Manual of Mental Disorders, 4th ed.-text revision. Author, Washington, DC (DSM-IV-TR).
- Baxter MG, Murray EA. The amygdala and reward. Nat Rev, Neurosci. 2002;3:563–573. doi: 10.1038/nrn875. [DOI] [PubMed] [Google Scholar]
- Brown TT, Lugar HM, Coalson RS, Meizin FM, Petersen SE, Schlaggar BL. Developmental changes in human cerebral functional organization for word generation. Cereb Cortex. 2005;15:275–290. doi: 10.1093/cercor/bhh129. [DOI] [PubMed] [Google Scholar]
- Burgund ED, Kang HC, Kelly JE, Buckner RL, Snyder AZ, Petersen SE, Schlaggar BL. Feasibility of a common stereo-tactic space for children and adults in fMRI studies of development. NeuroImage. 2002;17:184–200. doi: 10.1006/nimg.2002.1174. [DOI] [PubMed] [Google Scholar]
- Colditz G. Economic costs of obesity and inactivity. Med Sci Sports Exercise. 1999;31 (Suppl 11):S663–S667. doi: 10.1097/00005768-199911001-00026. [DOI] [PubMed] [Google Scholar]
- Craig AD, Reiman EM, Evans A, Bushnell MC. Functional imaging of an illusion of pain. Nature. 1996;384:258–260. doi: 10.1038/384258a0. [DOI] [PubMed] [Google Scholar]
- Davidson RJ. Well-being and affective style: neural substrates and biobehavioral correlates. Philos Trans R Soc Lond, Ser B Biol Sci. 2004;359:1395–1411. doi: 10.1098/rstb.2004.1510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidson MC, Thomas KM, Casey BJ. Imaging the developing brain with fMRI. Ment Retard Dev Disabil Res Rev. 2003;9:161–167. doi: 10.1002/mrdd.10076. [DOI] [PubMed] [Google Scholar]
- DelParigi A, Chen K, Salbe AD, Hill JO, Wing RR, Reiman EM, Tataranni PA. Persistence of abnormal neural responses to a meal in postobese individuals. Int J Obes. 2003;28:370–377. doi: 10.1038/sj.ijo.0802558. [DOI] [PubMed] [Google Scholar]
- Delvenne V, Goldman S, DeMaertelaer V, Lotstra F. Brain glucose metabolism in eating disorders assessed by positron emission tomography. Int J Eat Disord. 1999;25:29–37. doi: 10.1002/(sici)1098-108x(199901)25:1<29::aid-eat4>3.0.co;2-#. [DOI] [PubMed] [Google Scholar]
- Durston S, Hulshoff HE, Casey BJ, Giedd JN, Buitelaar JK, van Engeland H. Anatomical MRI of the developing human brain: what have we learned? J Am Acad Child Adolesc Psychiatry. 2001;40:1012–1020. doi: 10.1097/00004583-200109000-00009. [DOI] [PubMed] [Google Scholar]
- Friston KJ, Holmes AP, Worsley JJ. Statistical parametric maps in functional imaging: a general linear approach. Hum Brain Mapp. 1995;2:189–210. [Google Scholar]
- Gaillard WD, Grandin CB, Xu B. Developmental aspects of pediatric fMRI: considerations for image acquisition, analysis, and interpretation. NeuroImage. 2001;13:239–249. doi: 10.1006/nimg.2000.0681. [DOI] [PubMed] [Google Scholar]
- Gautier JF, Chen K, Uecker A, Bandy D, Frost J, Salbe AD, Pratley RE, Lawson M, Ravussin E, Reiman EM, Tataranni PA. Regions of the human brain affected during a liquid-meal taste perception in the fasting state: a positron emission tomography study. Am J Clin Nutr. 1999;70:806–810. doi: 10.1093/ajcn/70.5.806. [DOI] [PubMed] [Google Scholar]
- Gautier JF, Chen K, Salbe AD, Bandy D, Pratley RE, Heiman M, Ravussin E, Reiman EM, Tataranni PA. Differential brain responses to satiation in obese and lean men. Diabetes. 2000;49:838–846. doi: 10.2337/diabetes.49.5.838. [DOI] [PubMed] [Google Scholar]
- Gordon CM, Dougherty DD, Rauch SL, Emans SJ, Grace E, Lamm R, Alpert NM, Majzoub JA, Fischman AJ. Neuroanatomy of human appetitive function: a positron emission tomography investigation. Int J Eat Disord. 2000;27:163–171. doi: 10.1002/(sici)1098-108x(200003)27:2<163::aid-eat4>3.0.co;2-y. [DOI] [PubMed] [Google Scholar]
- Gottfried JA, O’Doherty J, Dolan RJ. Encoding predictive reward value in human amygdala and orbitofrontal cortex. Science. 2003;301:1104–1107. doi: 10.1126/science.1087919. [DOI] [PubMed] [Google Scholar]
- Happaney K, Zelazo PD, Stuss DT. Development of orbito-frontal function: current themes and future directions. Brain Cogn. 2004;55:1–10. doi: 10.1016/j.bandc.2004.01.001. [DOI] [PubMed] [Google Scholar]
- Hinton EC, Parkinson JA, Holland AJ, Arana FS, Roberts AC, Owen AM. Neural contributions to the motivational control of appetite in humans. Eur J Neurosci. 2004;20:1411–1418. doi: 10.1111/j.1460-9568.2004.03589.x. [DOI] [PubMed] [Google Scholar]
- Joseph JE. Functional neuroimaging studies of category specificity in object recognition: a critical review and meta-analysis. Cogn Affect Behav Neurosci. 2001;1:119–136. doi: 10.3758/cabn.1.2.119. [DOI] [PubMed] [Google Scholar]
- Killgore WDS, Young AD, Femia LA, Bogorodzki P, Rogowska J, Yurgelun-Todd DA. Cortical and limbic activation during viewing of high-versus low-calorie foods. NeuroImage. 2003;19:1381–1394. doi: 10.1016/s1053-8119(03)00191-5. [DOI] [PubMed] [Google Scholar]
- Kringelbach ML, Rolls ET. The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimaging and neurophysiology. Prog Neurobiol. 2004;72:341–372. doi: 10.1016/j.pneurobio.2004.03.006. [DOI] [PubMed] [Google Scholar]
- LaBar KS, Gitelman DR, Parrish TB, Kim Y, Nobre AC, Mesulam MM. Hunger selectively modulates corticolimbic activation to food stimuli in humans. Behav Neurosci. 2001;115:493–500. doi: 10.1037/0735-7044.115.2.493. [DOI] [PubMed] [Google Scholar]
- Mai, J.K., Assheuer, J., Paxinos, G., 2003. Atlas of the Human Brain, 2nd ed. Academic Press, San Diego.
- Matsuda M, Liu Y, Mahankali S, Pu Y, Mahankali A, Wang J, DeFronzo RA, Fox PT, Gao JH. Altered hypothalamic function in response to glucose ingestion in obese humans. Diabetes. 1999;48:1801–1806. doi: 10.2337/diabetes.48.9.1801. [DOI] [PubMed] [Google Scholar]
- May JC, Delgado MR, Dahl RE, Stenger VA, Ryan ND, Fiez JA, Carter CS. Event-related functional magnetic resonance imaging of reward-related brain circuitry in children and adolescents. Biol Psychiatry. 2004;55:359–366. doi: 10.1016/j.biopsych.2003.11.008. [DOI] [PubMed] [Google Scholar]
- Mokdad AH, Serdula MK, Dietz WH, Bowman BA, Marks JS, Koplan JP. The spread of the obesity epidemic in the United States, 1991–1998. JAMA. 1999;282:1519–1522. doi: 10.1001/jama.282.16.1519. [DOI] [PubMed] [Google Scholar]
- Mokdad AH, Serdula MK, Dietz WH, Bowman BA, Marks JS, Koplan JP. The continuing epidemic of obesity in the United States. JAMA. 2000;284:1650–1651. doi: 10.1001/jama.284.13.1650. [DOI] [PubMed] [Google Scholar]
- Morris JS, Dolan RJ. Involvement of human amygdala and orbitofrontal cortex in hunger-enhanced memory for food stimuli. J Neurosci. 2001;21:5304–5310. doi: 10.1523/JNEUROSCI.21-14-05304.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naruo T, Nakabeppu Y, Deguchi D, Nagai N, Tsutsui J, Nakajo M, Nozoe SS. Decreases in blood perfusion of the anterior cingulated gyri in anorexia nervosa restricters assessed by SPECT image analysis. BMC Psychiatry. 2001;1:2. doi: 10.1186/1471-244X-1-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen SJ, Siega-Riz AM, Popkin BM. Trends in energy intake in U.S. between 1977 and 1986: similar shifts seen across age groups. Obes Res. 2002;10:370–378. doi: 10.1038/oby.2002.51. [DOI] [PubMed] [Google Scholar]
- O’Doherty JP, Diechmann R, Critchley HD, Dolan RJ. Neural responses during anticipation of a primary taste reward. Neuron. 2002;33:815–826. doi: 10.1016/s0896-6273(02)00603-7. [DOI] [PubMed] [Google Scholar]
- Ongur D, Ferry AT, Price JL. Architectonic subdivision of the human orbital and medial prefrontal cortex. J Comp Neurol. 2003;460:425–449. doi: 10.1002/cne.10609. [DOI] [PubMed] [Google Scholar]
- Pollack I, Norman DA. A non-parametric analysis of recognition experiments. Psychon Sci. 1964;1:125–126. [Google Scholar]
- Pucak ML, Levitt JB, Lund JS, Lewis DA. Patterns of intrinsic and associational circuitry in monkey prefrontal cortex. J Comp Neurol. 1996;376:614–630. doi: 10.1002/(SICI)1096-9861(19961223)376:4<614::AID-CNE9>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- Rolls ET. Central nervous mechanisms related to feeding and appetite. Br Med Bull. 1981;37:131–134. doi: 10.1093/oxfordjournals.bmb.a071689. [DOI] [PubMed] [Google Scholar]
- Rolls ET. The functions of the orbitofrontal cortex. Brain Cogn. 2004;55:11–29. doi: 10.1016/S0278-2626(03)00277-X. [DOI] [PubMed] [Google Scholar]
- Rolls ET, Burton MJ, Mora F. Hypothalamic neuronal responses associated with the sight of food. Brain Res. 1976;111:53–66. doi: 10.1016/0006-8993(76)91048-9. [DOI] [PubMed] [Google Scholar]
- Small DM, Zatorre RJ, Dagher A, Evans AC, Jones-Gotman M. Changes in brain activity related to eating chocolate: from pleasure to aversion. Brain. 2001;124:1720–1733. doi: 10.1093/brain/124.9.1720. [DOI] [PubMed] [Google Scholar]
- Stellar E. The physiology of motivation. Psychol Rev. 1954;61:5–22. doi: 10.1037/h0060347. [DOI] [PubMed] [Google Scholar]
- Strauss RS, Pollack HA. Epidemic increase in childhood overweight, 1986–1998. JAMA. 2001;286:2845–2848. doi: 10.1001/jama.286.22.2845. [DOI] [PubMed] [Google Scholar]
- Takano A, Shiga T, Kitagawa N, Koyama T, Katoh C, Tsukamoto E, Tamaki N. Abnormal neuronal network in anorexia nervosa studied with I-123-IMP SPECT. Psychiatry Res. 2001;107:45–50. doi: 10.1016/s0925-4927(01)00093-2. [DOI] [PubMed] [Google Scholar]
- Talairach, J., Tournoux, P., 1988. Co-planar Stereotaxic Atlas of the Human Brain. Thieme Medical Publishers Inc., New York.
- Tataranni PA, DelParigi A. Functional neuroimaging: a new generation of human brain studies in obesity research. Obes Rev. 2003;4:229–238. doi: 10.1046/j.1467-789x.2003.00111.x. [DOI] [PubMed] [Google Scholar]
- Tataranni PA, Gautier JF, Chen K, Uecker A, Bandy D, Salbe AD, Pratley RE, Lawson M, Reiman EM, Ravussin E. Neuro-anatomical correlates of hunger and satiation in humans using positron emission tomography. Proc Natl Acad Sci U S A. 1999;96:459–4574. doi: 10.1073/pnas.96.8.4569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Troiano RP, Briefel RR, Carroll MD, Bialostosky K. Energy and fat intakes of children and adolescents in the United States: data from the national health and nutrition examination surveys. Am J Clin Nutr. 2000;72:1343S–1353S. doi: 10.1093/ajcn/72.5.1343s. Suppl. [DOI] [PubMed] [Google Scholar]
- Uher R, Murphy T, Brammer MJ, Dalgleish T, Phillips ML, Ng VW, Andrew CM, Williams SCR, Campbell IC, Treasure J. Medial prefrontal cortex activity associated with symptom provocation in eating disorders. Am J Psychiatry. 2004;161:1238–1246. doi: 10.1176/appi.ajp.161.7.1238. [DOI] [PubMed] [Google Scholar]
- Wang GJ, Volkow ND, Telang F, Jayne M, Ma J, Rao M, Zhu W, Wong CT, Pappas NR, Geliebter A, Fowler JS. Exposure to appetitive food stimuli markedly activates the human brain. Neuro-Image. 2004;21:1790–1797. doi: 10.1016/j.neuroimage.2003.11.026. [DOI] [PubMed] [Google Scholar]
- Whalen PJ. Fear, vigilance, and ambiguity: initial neuroimaging studies of the human amygdala. Curr Dir Psychol Sci. 1998;7:177–188. [Google Scholar]
- Will MJ, Franzblau EB, Kelley AE. The amygdala is critical for opioid-mediated binge eating of fat. NeuroReport. 2004;15:1857–1860. doi: 10.1097/00001756-200408260-00004. [DOI] [PubMed] [Google Scholar]
- Wright CI, Fischer H, Whalen PJ, McInerney SC, Shin LM, Rauch SL. Differential prefrontal cortex and amygdala habituation to repeatedly presented emotional stimuli. NeuroReport. 2001;12:379–383. doi: 10.1097/00001756-200102120-00039. [DOI] [PubMed] [Google Scholar]
- Wu JC, Hagman J, Buchsbaum MS, Blinder B, Derrfler M, Tai WY, Hazlett E, Sicotte N. Greater left cerebral hemispheric metabolism in bulimia assessed by positron emission tomography. Am J Psychiatry. 1990;147:309–312. doi: 10.1176/ajp.147.3.309. [DOI] [PubMed] [Google Scholar]
- Zald DH. The human amygdala and the emotional evaluation of sensory stimuli. Brain Res Rev. 2003;41:88–123. doi: 10.1016/s0165-0173(02)00248-5. [DOI] [PubMed] [Google Scholar]
- Zald DH, Lee JT, Fluegel KW, Pardo JV. Aversive gustatory stimulation activates limbic circuits in humans. Brain. 1998;121:1143–1154. doi: 10.1093/brain/121.6.1143. [DOI] [PubMed] [Google Scholar]