

Abstract
Sjögren's syndrome is a multisystem inflammatory rheumatic disease that is classified into primary and secondary forms, with cardinal features in the eye (keratoconjunctivitis sicca) and mouth (xerostomia). The aetiology behind this autoimmune exocrinopathy is probably multifactorial and influenced by genetic as well as by environmental factors that are as yet unknown. A genetic predisposition to Sjögren's syndrome has been suggested on the basis of familial aggregation, animal models and candidate gene association studies. Recent advances in molecular and genetic methodologies should further our understanding of this complex disease. The present review synthesizes the current state of genetics in Sjögren's syndrome.
Keywords: apoptosis, autoimmune disease, candidate genes, cytokines, HLA
Introduction
Sjögren's syndrome is an autoimmune exocrinopathy of unknown aetiology. It is a member of the group of inflammatory rheumatic disorders classified as connective tissue diseases. Clinical experience indicates not only an overlap among these disorders but also a close relationship of, for example, autoantibody profiles [1]. The genetic implications of this overlap has not been extensively explored, and the genetics behind Sjögren's syndrome per se are not well characterized.
There is no single disease-specific diagnostic criterion for Sjögren's syndrome. For diagnosis, the most functional criteria are the recently modified European classification criteria, which include a list of exclusions [2]. In addition to the subjective symptoms of dry eyes and dry mouth, the following objective signs should be present: ocular signs by Schirmer's I test and/or Rose Bengal score; focal sialadenitis by histopathology; salivary gland involvement by either salivary scintigraphy, parotid sialography or unstimulated salivary flow; and autoantibodies of Ro/ Sjögren syndrome antigen A (SSA) and/or La/Sjögren syndrome antigen B (SSB) specificity.
Sjögren's syndrome occurs worldwide and in all ages. However, the peak incidence is in the fourth and fifth decades of life, with a female:male ratio of 9:1. A number of studies have shown great variation in the frequency of Sjögren's syndrome (for review [3]). Prevalence studies have demonstrated that sicca symptoms and primary Sjögren's syndrome affects a considerable percentage of the population, with precise numbers dependent on the age group studied and on the criteria used [4]. A cautious but realistic estimate from the studies presented thus far is that primary Sjögren's syndrome is a disease with a prevalence not exceeding 0.6% of the general population (6/1000).
Although generally considered a T-cell-mediated disease, potential mechanisms underlying Sjögren's syndrome range from disturbances in apoptosis [5,6] to circulating autoantibodies against the ribonucleoproteins Ro and La [7,8] or cholinergic muscarinic receptors [9-11] in salivary and lacrimal glands in genetically predisposed individuals. Others relate reduced salivary and tear flow to aberrant glandular aquaporin-5 water channel proteins [12-14], although this is not unambiguous [15]. Possibly of greater importance is the recently described selective downregulation of aquaporin-1 expression in myoepithelial cells in salivary glands in primary Sjögren's syndrome [16].
Genetic predisposition to Sjögren's syndrome
A genetic predisposition to Sjögren's syndrome appears to exist, and several families involving two or more cases of Sjögren's syndrome have been described [17-23]. However, the level of genetic contribution is not known. Because large twin studies in Sjögren's syndrome are lacking, the twin concordance rate cannot be estimated. Only a few case reports describing twins with primary Sjögren's syndrome are available [24-27]. Twins exhibited a very similar phenotype with almost identical clinical presentation, including dry eyes and dry mouth; similar serological data (IgG, IgM, IgA, C3, C4, antinuclear antibody, anti-Ro/SSA and anti-La/SSB, rheumatoid factor), with identical fine specificity in their immune responses to 60kDa Ro/SSA; and identical labial salivary gland focus scores [24,27].
Familial clustering of different autoimmune diseases and co-association of multiple autoimmune diseases in individuals have frequently been reported [28]. Interestingly, it is common for a Sjögren's syndrome proband to have relatives with other autoimmune diseases (approximately 30–35%) [17,29,30]. Furthermore, Sjögren's syndrome exists in two forms – primary and secondary; the form that is present depends on whether it occurs alone or together with other connective diseases, such as systemic lupus erythematosus or rheumatoid arthritis [31]. Clustering of non-major histocompatibility complex (MHC) susceptibility candidate loci in human autoimmune diseases supports a hypothesis that, in some cases, clinically distinct autoimmune diseases may be controlled by a common set of susceptibility genes [32].
Sjögren's syndrome is considered a complex disorder. Susceptibility to the disease can be ascribed to an interplay between genetic factors and the environment. In complex diseases, one specific gene is neither necessary nor sufficient for disease expression. This makes the genetics behind these diseases more complicated than those of diseases with a simple Mendelian character.
Sjögren's syndrome is major histocompatility complex associated
The polymorphic MHC genes are the best documented genetic risk factors for the development of autoimmune diseases overall [33-35]. With regard to Sjögren's syndrome, the most relevant MHC complex genes are the class II genes, specifically the human leukocyte antigen (HLA)-DR and HLA-DQ alleles [36]. Patients of different ethnic origins exhibit different HLA gene associations [37]. In Caucasians of northern and western European backgrounds, including North American Caucasians, Sjögren's syndrome is among several autoimmune diseases that are associated with the haplotypes B8, DRw52 and DR3. The increased frequency of HLA-B8 was presumably due to an association with the HLA class II allele HLA-DRB1*03. However, a novel association of HLA class I alleles (i.e. HLA-A24) to susceptibility to primary Sjögren's syndrome was recently reported [38]. Beyond that, an association with DR2 has been found in Scandinavians [39] and with DR5 in Greeks [40]. All of the haplotypes are in strong linkage disequilibrium, resulting in certain difficulties in establishing which of the genes contains the locus that confers the risk. DQCAR is a very polymorphic CA repeat microsatellite located between the HLA DQA1 and DQB1 gene and specific DQCAR alleles have been found to be in tight linkage disequilibrium with known HLA DR-DQ haplotypes. HLA-DQB1 CAR1/CAR2 allele frequencies were found to be significantly different in patients with Sjögren's syndrome as compared with healthy control individuals in a study in which the Kaplan criteria were used to classify Sjögren's syndrome [41].
Apparently, the HLA haplotype may influence the severity of autoimmune disease. It has been claimed that Sjögren's syndrome patients with DQ1/DQ2 alleles have a more severe autoimmune disease than do patients with any other allelic combination at HLA-DQ [42], and the DR3-DQ2 haplotype has been indicated as a possible marker for a more active immune response in Finnish patients with Sjögren's disease [43].
HLA is associated with the presence of Ro and La autoantibodies in Sjögren's syndrome
Distinct HLA haplotypes have been associated with certain degrees of autoantibody diversification in Sjögren's syndrome [44]. Autoantibodies to Ro/SSA and La/SSB have been found to be associated with DR3, DQA and DQB alleles [45-47]. A dose-dependent contribution of DQα-34Q and DQβ-26L, in addition to the DRB1*03-DQB1*02-DQA1*0501 haplotype encompassing the transethnically associated DQβ-DI motif, represented the strongest contributors to the formation of an anti-Ro/La response in Norwegian patients with Sjögren's syndrome [45]. A stronger correlation has been found between anti-Ro/SSA autoantibodies and DR3/DR2 than that with the disease itself [45,48-50]. In Japanese persons, HLA class II allele association has been reported to differ among anti-Ro/SSA-positive individuals according to the presence or absence of coexisting anti-La/SSB [51].
Cytokine polymorphisms in Sjögren's syndrome
Cytokines serve to mediate and regulate immune and inflammatory responses, and have been implicated in the pathogenesis of a variety of autoimmune diseases, including Sjögren's syndrome. Numerous investigators have attempted to analyze the association of primary Sjögren's syndrome with cytokine polymorphisms, but at present no convincing relationship has been identified (for review [52]).
Both human and animal studies indicate the involvement of IL-10 in the pathogenesis of primary Sjögren's syndrome [53] and mice transgenic for IL-10 develop a Fas-ligand-mediated exocrinopathy that resembles Sjögren's syndrome [54]. A recent study described an association between primary Sjögren's syndrome and IL-10 promoter polymorphisms in a cohort of Finnish individuals, and a specific haplotype was found to correlate with high plasma levels of IL-10 [55]. Conversely, no association was found for IL-10 promoter polymorphism and primary Sjögren's syndrome or the presence of Ro autoantibodies in an Australian cohort of primary Sjögren's syndrome patients [56].
The IL-1 receptor antagonist regulates IL-1 activity in inflammatory disorders by binding to IL-1 receptors and thus inhibiting the activity of IL-1. The human IL-1 receptor antagonist gene (i.e. IL1RN) has a variable allelic polymorphism within intron 2 as a result of variation in number of an 86-base-pair sequence repeat [52]. An increased frequency and carriage rate of the IL1RN*2 allele has been found in primary Sjögren's syndrome [57]. No statistically significant association can be ascribed to tumour necrosis factor-α and primary Sjögren's syndrome [58].
Additional candidate gene studies
Because there is no disease-specific criterion for Sjögren's syndrome, the candidate genes studied may be related to other autoimmune phenotypes also. Mutations in the apoptosis genes have been identified as a possible cause or a contributing factor to human diseases [59], and the role of apoptosis has also been a major topic in autoimmune diseases, including primary Sjögren's syndrome (for review [5,6]).
An increased frequency of apoptosis in ductal epithelial cells of the salivary glands leading to reduced salivary flow has been proposed as a possible disease mechanism [60,61]. Other investigators have suggested that inflammatory mononuclear cells are able to escape apoptosis because of defects in the death signalling pathway, which lead to accumulation of lymphocytes to displacement of functioning acinar cells [6,62]. In the complex cascade of apoptotic signal molecules, Fas and Fas ligand are central actors. An insert of a retrotransposon in the Fas gene was discovered in the murine model MRL/lpr-lpr, which exhibits progressive focal sialadenitis, and has as such been forwarded as a possible explanation for aberrant apoptosis in that experimental model [63-66]. This finding led to speculation over whether a similar phenomenon may be present in the human Fas gene. Now, more than 20 distinct Fas mutations are known in humans, and mutations in this gene have been identified as cause of or factor contributing to human diseases, such as autoimmune lymphoproliferative syndrome type I (for review [59]). Polymorphisms in the Fas and FasL genes have also been found in primary Sjögren's syndrome [67]. However, none of the polymorphisms in Fas or FasL entailed amino acid changes in patients with primary Sjögren's syndrome, and at present there exists no clear-cut mutation or defect in these genes that is clearly associated with primary Sjögren's syndrome and as such can be regarded as a disease-determining factor. Notably, the apoptosis cascade is built up of a huge number of signal molecules, and the possibility that there should be polymorphisms or mutations of vital importance for development of the disease among these factors still exists. Thus far, however, a definite role for apoptosis in primary Sjögren's syndrome cannot be confirmed.
The contribution of Ro/SSA and La/SSB in Sjögren's syndrome is not fully understood. It is not known how tolerance breakdown and autoantibody response to Ro/SSA and La/SSB is generated. The ribonucleoproteins are endogenous proteins that are normally hidden from the immune system, and should subsequently not give rise to abnormal B-cell responses. However, stress such as ultraviolet radiation, viral infections and apoptosis have been suggested to lead to undesirable cell surface exposure of autoantigens to the immune system [7]. Ro/SSA and La/SSB have been demonstrated in surface blebs of apoptotic ultraviolet-irradiated keratinocytes, implying a role in systemic lupus erythematosus [68]. Not much is known from a genetic point of view, but an association study has been performed in Ro52[69]. A single nucleotide polymorphism in intron 3 of the Ro52 gene was found to be strongly associated with the presence of anti-Ro52 autoantibodies in primary Sjögren's syndrome [69]. This is interesting because alternative mRNA is made by deleting exon 4, which encodes a putative leucine zipper domain, to generate a shorter version of the Ro52 protein [70].
Genes that encode transporters associated with antigen processing (i.e. TAP genes) have also been associated with susceptibility to Sjögren's syndrome [71]. Others have indicated a putative role for the cysteine-rich secretory protein 3 (CRISP-3) gene as an early response gene that may participate in the pathophysiology of the autoimmune lesions of Sjögren's syndrome [72].
A 44-fold increased risk for the development of B-cell lymphoma has been documented in Sjögren's syndrome, and a role for activated B cells has been implicated [73]. Notably, it is not known whether B-cell activation is a primary cause or a secondary manifestation in Sjögren's syndrome. Patients are known to have increased levels of serum IgG [3]. Although the cellular basis of this hypergammaglobulinaemia and the strong associations of certain autoantibodies with particular MHC class II molecules have been intensively examined, little is known about the usage of IgV (immunoglobulin variable) region genes, and especially by autoantibodies in autoimmune diseases. Therefore, a study of IgVλ light chain gene usage in primary Sjögren's syndrome patients was undertaken by Kaschner et al. [74]. Those investigators identified molecular differences from controls in V-J recombination and concluded that disturbed regulation of B-cell maturation with abnormal selection, defects in editing immunoglobulin receptors and abnormal mutational targeting may contribute to the emergence of autoimmunity in Sjögren's syndrome.
Rheumatoid factors are autoantibodies against antigenic determinants that are present on the Fc portion of human IgG, and are found in sera and saliva of 60–80% of patients with primary Sjögren's syndrome [3]. We found rheumatoid factor to be present in sera of 91% of Norwegian anti-Ro-positive patients with primary Sjögren's syndrome [45]. The genetic origin and the mechanisms underlying its generation have been investigated in primary Sjögren's syndrome [75]. In such patients, rheumatoid factor used diverse VH region genes, the majority of which show no evidence of somatic hypermutation, whereas light chain variable (VL) sequences exhibited a moderate contribution of somatic hypermutation [76].
Understanding primary Sjögren's syndrome in view of animal models
An appropriate animal model of Sjögren's syndrome could greatly advance our ability to identify the target antigens, characterize the immune mechanisms and define the genetic background. Several animal models, both experimentally induced and spontaneous inflammatory reactions with features of human Sjögren's syndrome, have been reported and previously reviewed [77].
The nonobese diabetic (NOD) mouse develops a disease that mimicks human type 1 diabetes mellitus and has been intensively studied for this phenotype. It also spontaneously develops sialadenitis and several other features of Sjögren's syndrome, including autoantibodies against Ro/SSA [77]. The NOD mouse carries the MHC H2g7 haplotype. In order to study the importance of NOD non-MHC genes, an H2q congenic NOD mouse, namely NOD.Q, was established [78,79]. Recently, a gene segregation experiment was conducted in a (NOD.Q × B10.Q)F2 cross, and genetic mapping revealed one locus (Nss1) associated with sialadenitis on chromosome 4 (LOD score 4.7; Fig. 1) [78]. The H2g7 haplotype was not critical for sialadenti-tis development in the NOD background because the NOD.Q mouse also developed sialadenitis. The genetic control of sialadenitis appeared to be unique in comparison with diabetes and arthritis, because no loci associated with these diseases have been identified at the same location [79]. This supports earlier findings that the sicca syndrome occurs independently of autoimmune diabetes, and NOD MHC I-Ag7 was not essential for exocrine tissue autoimmunity [80].
Figure 1.
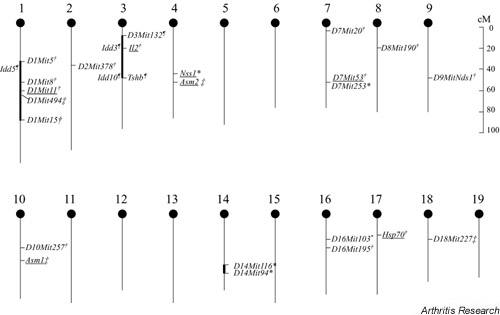
Chromosomal map illustrating the location of identified quantitative trait loci associated with sialadenitis development in various murine models. Chromosomal positions are based on the map from the Jackson Laboratory http://informatics.jax.org/. Sialadenitis susceptibility loci are drawn from *[78], ¶[82], †[83] and ‡[84]; markers with LOD score >3.3 are underscored, however, for the markers Il2, Asm2 and Hsp70 a LOD score >3.3 was only obtained in females and for the marker D1Mit153 only in males.
More recently, alleles from chromosomes 1 and 3 of NOD mice have been found to combine to influence Sjögren's syndrome-like autoimmune exocrinopathy [81], and two intervals contribute synergistically to the development of Sjögren's syndrome on a healthy murine background; this has also been demonstrated in the NOD mouse after crossing (Fig. 1) [82]. Very recently, chromosome 1 was reported to be a major susceptibility region for development of autoimmune sialadenitis [83]. In different matings of NOD mice, including a (NOD × C57BL/6 [B6])F2 cross, a (NOD × NZW)F2 cross, and a ([NOD × B6] × NOD) backcross, an association with the middle region of chromosome 1 was detected in all crosses.
The NZB, MRL/lpr, NOD and NFS/sld strains are all experimental murine models that spontaneously develop salivary gland inflammation, of which the MRL/lpr and the NOD strains present with serum anti-Ro/SSA antibodies. An insertion of an ET-transposon in the Fas gene has been found to be responsible for the lpr genotype in the MRL/lpr mouse [65]. Similar Fas gene insertions could not be traced in Sjögren's syndrome patients [67]. A genome-wide scan of MRL/lpr mice revealed four susceptible loci, mapped on chrosome 10, 18, 4 and 1, which were recessively associated with sialadenitis [84]. The sialadenitis in MRL/lpr mice is probably under the control of polygenic inheritance, because the loci manifested an additive effect in a hierarchical manner. The different susceptibility loci reported for sialadenitis are outlined in Fig. 1.
Transgenic mice have frequently been used as models to study the role of viruses in the pathogenesis of a variety of diseases and to determine the importance of cytokines such as IL-10 [54]. Transgenic expression of IL-10 induced apoptosis of glandular tissue and promoted infiltration of lymphocytes. Transgenic mice containing the human T-cell lymphotropic virus type-1 tax gene under the control of the viral long terminal repeat (LTR) develop an exocrinopathy that involves the salivary and lacrimal glands, resembling the pathology of Sjögren's syndrome [85]. It was suggested that human T-cell lymphotropic virus type-1 may represent a primary event in the development of exocrinopathy by virally induced proliferation and perturbation of the function of ductal epithelium. Sialadenitis and inflammation in lachrymal glands histologically resembling Sjögren's syndrome have also been found in mice transgenic for hepatitis C virus envelope genes [86].
Conclusion
Very little is known about the genetics of Sjögren's syndrome. Although not conclusive, however, recent findings in animal breeding studies are promising with respect to resolving issues in Sjögren's syndrome. Of special interest were the major susceptibility loci for autoimmune sialadenitis demonstrated on chromosomes 1, 4 and 10 in murine models. For instance, the chromosomal regions around Nss1 on chromosome 4 harbour a set of genes that are probably of importance for different kinds of autoimmune syndromes, because several loci associated with autoimmune disease models for systemic lupus erythematosus and autoimmune haemolytic anaemia are clustered around Nss1 (for review [78]). Interestingly, no association between sialadenitis in the NOD.Q and collagen-induced arthritis was observed [78].
Human linkage studies of Sjögren's syndrome families, in addition to analyses of gene expression signatures on microarrays, will probably be an important source of information in the future. Identification of new genetic markers may lead to development of better diagnostic and prognostic tests in Sjögren's syndrome, including its systemic complications. However, as with the other rheumatic diseases, it is anticipated that both overlap and discrepancies will be detected during genome screens. Given the likely heterogeneity of Sjögren's syndrome, advances will probably not be made without future global collaboration.
Abbreviations
HLA = human leukocyte antigen; IL = interleukin; LTR = long terminal repeat; MHC = major histocompatibility complex; NOD = nonobese diabetic; SSA = Sjögren syndrome antigen A; SSB = Sjögren syndrome antigen B.
Acknowledgments
Acknowledgements
Studies by the authors were financed with the aid of EXTRA funds from the Norwegian Foundation for Health and Rehabilitation, the European BIOMED program (BMH4-CT98-3489) and the Broegelmann Foundation.
References
- Jonsson R, Brokstad KA. Sjögren's syndrome. In: Austen KF, Frank MM, Atkinson JP, Cantor H, editor. In Samter's Immunologic Diseases. 6. Philadelphia: Lippincott, Williams & Wilkins; 2001. pp. 495–504. [Google Scholar]
- Vitali C, Bombardieri S, Jonsson R, Moutsopoulos HM, Alexander EL, Carsons SE, Daniels TE, Fox PC, Fox RI, Kassan SS, Pillemer SR, Talal N, Weisman MH. Classification criteria for Sjögren's syndrome: a revised version of the European criteria proposed by the American-European Consensus Group. Ann Rheum Dis. 2002;61:554–558. doi: 10.1136/ard.61.6.554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jonsson R, Haga H-J, Gordon T. Sjögren's syndrome. In: Koopman WJ, editor. Arthritis and Allied Contitions: a Textbook of Rheumatology. 14. Philadelphia: Lippincott, Williams & Wilkins,; 2001. pp. 1736–1759. [Google Scholar]
- Silman AJ, Rooney BK. Epidemiology of Sjögren's syndrome. In: Eriksson E, Jonsson R, editor. In The 100-year Anniversary of Henrik Sjögren. Jönköping: Hygiea,; 1999. pp. 53–57. [Google Scholar]
- Bolstad AI, Jonsson R. The role of apoptosis in Sjögren's syndrome. Ann Med Interne. 1998;149:25–29. [PubMed] [Google Scholar]
- Ohlsson M, Skarstein K, Bolstad AI, Johannessen AC, Jonsson R. Fas-induced apoptosis is a rare event in Sjögren's syndrome. Lab Invest. 2001;81:95–105. doi: 10.1038/labinvest.3780215. [DOI] [PubMed] [Google Scholar]
- Ohlsson M, Jonsson R, Brokstad K. Subcellular redistribution and surface exposure of the Ro52, Ro60 and La48 autoantigen during apoptosis in human ductal epithelial cells: a possible mechanism in the pathogenesis of Sjögren's syndrome. Scand J Immunol. 2002. [DOI] [PubMed]
- Gordon TP, Bolstad AI, Rischmueller M, Jonsson R, Waterman SA. Autoantibodies in primary Sjögren's syndrome: new insights into mechanisms of autoantibody diversification and disease pathogenesis. Autoimmunity. 2001;34:123–132. doi: 10.3109/08916930109001960. [DOI] [PubMed] [Google Scholar]
- Beroukas D, Goodfellow R, Hiscock J, Jonsson R, Gordon TP, Waterman SA. Up-regulation of M3-muscarinic receptors in labial salivary gland acini in primary Sjögren's syndrome. Lab Invest. 2002;82:203–210. doi: 10.1038/labinvest.3780412. [DOI] [PubMed] [Google Scholar]
- Bacman S, Sterin-Borda L, Camusso JJ, Arana R, Hubscher O, Borda E. Circulating antibodies against rat parotid gland M3 muscarinic receptors in primary Sjögren's syndrome. Clin Exp Immunol. 1996;104:454–459. doi: 10.1046/j.1365-2249.1996.42748.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphreys-Beher MG, Brayer J, Yamachika S, Peck AB, Jonsson R. An alternative perspective to the immune response in autoimmune exocrinopathy: induction of functional quiescence rather than destructive autoaggression. Scand J Immunol. 1999;49:7–10. doi: 10.1046/j.1365-3083.1999.00490.x. [DOI] [PubMed] [Google Scholar]
- Ma T, Song Y, Gillespie A, Carlson EJ, Epstein CJ, Verkman AS. Defective secretion of saliva in transgenic mice lacking aquaporin-5 water channels. J Biol Chem. 1999;274:20071–20074. doi: 10.1074/jbc.274.29.20071. [DOI] [PubMed] [Google Scholar]
- Steinfeld S, Cogan E, King LS, Agre P, Kiss R, Delporte C. Abnormal distribution of aquaporin-5 water channel protein in salivary glands from Sjögren's syndrome patients. Lab Invest. 2001;81:143–148. doi: 10.1038/labinvest.3780221. [DOI] [PubMed] [Google Scholar]
- Tsubota K, Hirai S, King LS, Agre P, Ishida N. Defective cellular trafficking of lacrimal gland aquaporin-5 in Sjögren's syndrome. Lancet. 2001;357:688–689. doi: 10.1016/S0140-6736(00)04140-4. [DOI] [PubMed] [Google Scholar]
- Beroukas D, Hiscock J, Jonsson R, Waterman SA, Gordon TP. Subcellular distribution of aquaporin 5 in salivary glands in primary Sjögren's syndrome. Lancet. 2001;358:1875–1876. doi: 10.1016/S0140-6736(01)06900-8. [DOI] [PubMed] [Google Scholar]
- Beroukas D, Hiscock J, Gannon BJ, Jonsson R, Gordon TP, Waterman SA. Selective downregulation of aquaporin-1 in salivary glands in primary Sjögren's syndrome. Lab Invest. 2002. [DOI] [PubMed]
- Reveille JD, Wilson RW, Provost TT, Bias WB, Arnett FC. Primary Sjögren's syndrome and other autoimmune diseases in families. Prevalence and immunogenetic studies in six kindreds. Ann Intern Med. 1984;101:748–756. doi: 10.7326/0003-4819-101-6-748. [DOI] [PubMed] [Google Scholar]
- Lichtenfeld JL, Kirschner RH, Wiernik PH. Familial Sjögren's syndrome with associated primary salivary gland lymphoma. Am J Med. 1976;60:286–292. doi: 10.1016/0002-9343(76)90439-3. [DOI] [PubMed] [Google Scholar]
- Doni A, Brancato R, Bartoletti L, Berni G. Familiar characteristics of Sjögren's disease. (Clinical contribution and considerations) [in Italian]. Riv Crit Clin Med. 1965;65:750–759. [PubMed] [Google Scholar]
- Koivukangas T, Simila S, Heikkinen E, Rasanen O, Wasz-Hockert O. Sjögren's syndrome and achalasia of the cardia in two siblings. Pediatrics. 1973;51:943–945. [PubMed] [Google Scholar]
- Mason AM, Golding PL. Multiple immunological abnormalities in a family. J Clin Pathol. 1971;24:732–735. doi: 10.1136/jcp.24.8.732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boling EP, Wen J, Reveille JD, Bias WB, Chused TM, Arnett FC. Primary Sjögren's syndrome and autoimmune hemolytic anemia in sisters. A family study. Am J Med. 1983;74:1066–1071. doi: 10.1016/0002-9343(83)90820-3. [DOI] [PubMed] [Google Scholar]
- Sabio JM, Milla E, Jimenez-Alonso J. A multicase family with primary Sjögren's syndrome. J Rheumatol. 2001;28:1932–1934. [PubMed] [Google Scholar]
- Scofield RH, Kurien BT, Reichlin M. Immunologically restricted and inhibitory anti-Ro/SSA in monozygotic twins. Lupus. 1997;6:395–398. doi: 10.1177/096120339700600409. [DOI] [PubMed] [Google Scholar]
- Besana C, Salmaggi C, Pellegrino C, Pierro L, Vergani S, Faravelli A, Rugarli C. Chronic bilateral dacryo-adenitis in identical twins: a possible incomplete form of Sjögren syndrome. Eur J Pediatr. 1991;150:652–655. doi: 10.1007/BF02072627. [DOI] [PubMed] [Google Scholar]
- Koga T, Ebata H, Tanigawa K, Yamagata Y. Identical twins of Sjorgren's syndrome with renal tubular acidosis (author's transl) [in Japanese]. Nippon Naika Gakkai Zasshi. 1980;69:1458–1462. [PubMed] [Google Scholar]
- Bolstad AI, Haga HJ, Wassmuth R, Jonsson R. Monozygotic twins with primary Sjögren's syndrome. J Rheumatol. 2000;27:2264–2266. [PubMed] [Google Scholar]
- Becker KG, Simon RM, Bailey-Wilson JE, Freidlin B, Biddison WE, McFarland HF, Trent JM. Clustering of non-major histocompatibility complex susceptibility candidate loci in human autoimmune diseases. Proc Natl Acad Sci USA. 1998;95:9979–9984. doi: 10.1073/pnas.95.17.9979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka A, Igarashi M, Kakinuma M, Oh-i T, Koga M, Okuda T. The occurrence of various collagen diseases in one family: a sister with ISSc, PBC, APS, and SS and a brother with systemic lupus erythematosus. J Dermatol. 2001;28:547–553. doi: 10.1111/j.1346-8138.2001.tb00028.x. [DOI] [PubMed] [Google Scholar]
- Reveille JD, Arnett FC. The immunogenetics of Sjögren's syndrome. Rheum Dis Clin North Am. 1992;18:539–550. [PubMed] [Google Scholar]
- Jonsson R, Haga HJ, Gordon TP. Current concepts on diagnosis, autoantibodies and therapy in Sjögren's syndrome. Scand J Rheumatol. 2000;29:341–348. doi: 10.1080/030097400447525. [DOI] [PubMed] [Google Scholar]
- Heward J, Gough SC. Genetic susceptibility to the development of autoimmune disease. Clin Sci (Lond) 1997;93:479–491. doi: 10.1042/cs0930479. [DOI] [PubMed] [Google Scholar]
- Nepom GT. MHC and autoimmune diseases. Immunol Ser. 1993;59:143–164. [PubMed] [Google Scholar]
- Merriman TR, Todd JA. Genetics of autoimmune disease. Curr Opin Immunol. 1995;7:786–792. doi: 10.1016/0952-7915(95)80049-2. [DOI] [PubMed] [Google Scholar]
- Tomlinson IP, Bodmer WF. The HLA system and the analysis of multifactorial genetic disease. Trends Genet. 1995;11:493–498. doi: 10.1016/s0168-9525(00)89159-3. [DOI] [PubMed] [Google Scholar]
- Reveille JD. The molecular genetics of systemic lupus erythematosus and Sjögren's syndrome. Curr Opin Rheumatol. 1992;4:644–656. [PubMed] [Google Scholar]
- Kang HI, Fei HM, Saito I, Sawada S, Chen SL, Yi D, Chan E, Peebles C, Bugawan TL, Erlich HA, et al. Comparison of HLA class II genes in Caucasoid, Chinese, and Japanese patients with primary Sjögren's syndrome. J Immunol. 1993;150:3615–3623. [PubMed] [Google Scholar]
- Loiseau P, Lepage V, Djelal F, Busson M, Tamouza R, Raffoux C, Menkes CJ, Meyer O, Charron D, Goldberg D. HLA class I and class II are both associated with the genetic predisposition to primary Sjögren syndrome. Hum Immunol. 2001;62:725–731. doi: 10.1016/s0198-8859(01)00253-1. [DOI] [PubMed] [Google Scholar]
- Manthorpe R, Morling N, Platz P, Ryder LP, Svejgaard A, Thomsen M. HLA-D antigen frequencies in Sjögren's syndrome. Differences between the primary and secondary form. Scand J Rheumatol. 1981;10:124–128. doi: 10.3109/03009748109095284. [DOI] [PubMed] [Google Scholar]
- Papasteriades CA, Skopouli FN, Drosos AA, Andonopoulos AP, Moutsopoulos HM. HLA-alloantigen associations in Greek patients with Sjögren's syndrome. J Autoimmun. 1988;1:85–90. doi: 10.1016/0896-8411(88)90079-0. [DOI] [PubMed] [Google Scholar]
- Harley JB, Reichlin M, Arnett FC, Alexander EL, Bias WB, Provost TT. Gene interaction at HLA-DQ enhances autoantibody production in primary Sjögren's syndrome. Science. 1986;232:1145–1147. doi: 10.1126/science.3458307. [DOI] [PubMed] [Google Scholar]
- Kerttula TO, Collin P, Polvi A, Korpela M, Partanen J, Maki M. Distinct immunologic features of Finnish Sjögren's syndrome patients with HLA alleles DRB1* DQA1*0501, and DQB1*0201. Alterations in circulating T cell receptor gamma/delta subsets. Arthritis Rheum. 0301;39:1733–1739. doi: 10.1002/art.1780391017. [DOI] [PubMed] [Google Scholar]
- Kacem HH, Kaddour N, Adyel FZ, Bahloul Z, Ayadi H. HLA-DQB1 CAR1/CAR2, TNFa IR2/IR4 and CTLA-4 polymorphisms in Tunisian patients with rheumatoid arthritis and Sjögren's syndrome. Rheumatology (Oxford) 2001;40:1370–1374. doi: 10.1093/rheumatology/40.12.1370. [DOI] [PubMed] [Google Scholar]
- Rischmueller M, Lester S, Chen Z, Champion G, Van Den Berg R, Beer R, Coates T, McCluskey J, Gordon T. HLA class II phenotype controls diversification of the autoantibody response in primary Sjögren's syndrome (pSS). Clin Exp Immunol. 1998;111:365–371. doi: 10.1046/j.1365-2249.1998.00504.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolstad AI, Wassmuth R, Haga HJ, Jonsson R. HLA markers and clinical characteristics in Caucasians with primary Sjögren's syndrome. J Rheumatol. 2001;28:1554–1562. [PubMed] [Google Scholar]
- Nakken B, Jonsson R, Brokstad KA, Omholt K, Nerland AH, Haga HJ, Halse AK. Associations of MHC class II alleles in Norwegian primary Sjögren's syndrome patients: implications for development of autoantibodies to the Ro52 autoantigen. Scand J Immunol. 2001;54:428–433. doi: 10.1046/j.1365-3083.2001.00993.x. [DOI] [PubMed] [Google Scholar]
- Fei HM, Kang H, Scharf S, Erlich H, Peebles C, Fox R. Specific HLA-DQA and HLA-DRB1 alleles confer susceptibility to Sjögren's syndrome and autoantibody production. J Clin Lab Anal. 1991;5:382–391. doi: 10.1002/jcla.1860050604. [DOI] [PubMed] [Google Scholar]
- Arnett FC, Hamilton RG, Reveille JD, Bias WB, Harley JB, Reichlin M. Genetic studies of Ro (SS-A) and La (SS-B) autoantibodies in families with systemic lupus erythematosus and primary Sjögren's syndrome. Arthritis Rheum. 1989;32:413–419. doi: 10.1002/anr.1780320410. [DOI] [PubMed] [Google Scholar]
- Hamilton RG, Harley JB, Bias WB, Roebber M, Reichlin M, Hochberg MC, Arnett FC. Two Ro (SS-A) autoantibody responses in systemic lupus erythematosus. Correlation of HLA-DR/DQ specificities with quantitative expression of Ro (SS-A) autoantibody. Arthritis Rheum. 1988;31:496–505. doi: 10.1002/art.1780310406. [DOI] [PubMed] [Google Scholar]
- Wilson WA, Scopelitis E, Michalski JP. Association of HLA-DR7 with both antibody to SSA(Ro) and disease susceptibility in blacks with systemic lupus erythematosus. J Rheumatol. 1984;11:653–657. [PubMed] [Google Scholar]
- Miyagawa S, Shinohara K, Nakajima M, Kidoguchi K, Fujita T, Fukumoto T, Yoshioka A, Dohi K, Shirai T. Polymorphisms of HLA class II genes and autoimmune responses to Ro/SS-ALa/SS-B among Japanese subjects. Arthritis Rheum. 1998;41:927–934. doi: 10.1002/1529-0131(199805)41:5<927::AID-ART21>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- Magnusson V, Nakken B, Bolstad AI, Alarcon-Riquelme ME. Cytokine polymorphisms in systemic lupus erythematosus and Sjögren's syndrome. Scand J Immunol. 2001;54:55–61. doi: 10.1046/j.1365-3083.2001.00965.x. [DOI] [PubMed] [Google Scholar]
- Halse AK, Tengner P, Wahren-Herlenius M, Haga H-J, Jonsson R. Increased frequency of cells secreting interleukin-6 and interleukin-10 in peripheral blood of patients with primary Sjögren's syndrome. Scand J Immunol. 1999;49:533–538. doi: 10.1046/j.1365-3083.1999.00533.x. [DOI] [PubMed] [Google Scholar]
- Saito I, Haruta K, Shimuta M, Inoue H, Sakurai H, Yamada K, Ishi-maru N, Higashiyama H, Sumida T, Ishida H, Suda T, Noda T, Hayashi Y, Tsubota K. Fas ligand-mediated exocrinopathy resembling Sjögren's syndrome in mice transgenic for IL-10. J Immunol. 1999;162:2488–2494. [PubMed] [Google Scholar]
- Hulkkonen J, Pertovaara M, Antonen J, Lahdenpohja N, Pasternack A, Hurme M. Genetic association between interleukin-10 promoter region polymorphisms and primary Sjögren's syndrome. Arthritis Rheum. 2001;44:176–179. doi: 10.1002/1529-0131(200101)44:1<176::AID-ANR23>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- Rischmueller M, Limaye V, Lester S, Downie-Doyle S, Pile K, Bardy P, Gordon TP. Polymorphisms of the interleukin 10 gene promoter are not associated with anti-Ro autoantibodies in primary Sjögren's syndrome. J Rheumatol. 2000;27:2945–2946. [PubMed] [Google Scholar]
- Perrier S, Coussediere C, Dubost JJ, Albuisson E, Sauvezie B. IL-1 receptor antagonist (IL-1RA) gene polymorphism in Sjögren's syndrome and rheumatoid arthritis. Clin Immunol Immunopathol. 1998;87:309–313. doi: 10.1006/clin.1998.4520. [DOI] [PubMed] [Google Scholar]
- Jean S, Quelvennec E, Alizadeh M, Guggenbuhl P, Birebent B, Perdriger A, Grosbois B, Pawlotsky PY, Semana G. DRB1*15 and DRB1*03 extended haplotype interaction in primary Sjögren's syndrome genetic susceptibility. Clin Exp Rheumatol. 1998;16:725–728. [PubMed] [Google Scholar]
- Mullauer L, Gruber P, Sebinger D, Buch J, Wohlfart S, Chott A. Mutations in apoptosis genes: a pathogenetic factor for human disease. Mutat Res. 2001;488:211–231. doi: 10.1016/s1383-5742(01)00057-6. [DOI] [PubMed] [Google Scholar]
- Manganelli P, Quaini F, Andreoli AM, Lagrasta C, Pilato FP, Zuc-carelli A, Monteverdi R, D'Aversa C, Olivetti G. Quantitative analysis of apoptosis and bcl-2 in Sjögren's syndrome. J Rheumatol. 1997;24:1552–1557. [PubMed] [Google Scholar]
- Matsumura R, Umemiya K, Kagami M, Tomioka H, Tanabe E, Sugiyama T, Sueishi M, Nakajima A, Azuma M, Okumura K, Sumida T. Glandular and extraglandular expression of the Fas-Fas ligand and apoptosis in patients with Sjögren's syndrome. Clin Exp Rheumatol. 1998;16:561–568. [PubMed] [Google Scholar]
- Skarstein K, Nerland AH, Eidsheim M, Mountz JD, Jonsson R. Lymphoid cell accumulation in salivary glands of autoimmune MRL mice can be due to impaired apoptosis. Scand J Immunol. 1997;46:373–378. doi: 10.1046/j.1365-3083.1997.d01-142.x. [DOI] [PubMed] [Google Scholar]
- Mountz JD, Zhou T, Su X, Wu J, Cheng J. The role of programmed cell death as an emerging new concept for the pathogenesis of autoimmune diseases. Clin Immunol Immunopathol. 1996;80:S2–S14. doi: 10.1006/clin.1996.0136. [DOI] [PubMed] [Google Scholar]
- Chu JL, Drappa J, Parnassa A, Elkon KB. The defect in Fas mRNA expression in MRL/lpr mice is associated with insertion of the retrotransposon, ETn. J Exp Med. 1993;178:723–730. doi: 10.1084/jem.178.2.723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J, Zhou T, He J, Mountz JD. Autoimmune disease in mice due to integration of an endogenous retrovirus in an apoptosis gene. J Exp Med. 1993;178:461–468. doi: 10.1084/jem.178.2.461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adachi M, Watanabe-Fukunaga R, Nagata S. Aberrant transcription caused by the insertion of an early transposable element in an intron of the Fas antigen gene of lpr mice. Proc Natl Acad Sci USA. 1993;90:1756–1760. doi: 10.1073/pnas.90.5.1756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolstad AI, Wargelius A, Nakken B, Haga HJ, Jonsson R. Fas and Fas ligand gene polymorphisms in primary Sjögren's syndrome. J Rheumatol. 2000;27:2397–2405. [PubMed] [Google Scholar]
- Casciola-Rosen LA, Anhalt G, Rosen A. Autoantigens targeted in systemic lupus erythematosus are clustered in two populations of surface structures on apoptotic keratinocytes. J Exp Med. 1994;179:1317–1330. doi: 10.1084/jem.179.4.1317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakken B, Jonsson R, Bolstad AI. Polymorphisms of the Ro52 gene associated with anti-Ro 52-kd autoantibodies in patients with primary Sjögren's syndrome. Arthritis Rheum. 2001;44:638–646. doi: 10.1002/1529-0131(200103)44:3<638::AID-ANR112>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- Chan EK, Di Donato F, Hamel JC, Tseng CE, Buyon JP. 52-kD SS-A/Ro: genomic structure and identification of an alternatively spliced transcript encoding a novel leucine zipperminus autoantigen expressed in fetal and adult heart. J Exp Med. 1995;182:983–992. doi: 10.1084/jem.182.4.983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumagai S, Kanagawa S, Morinobu A, Takada M, Nakamura K, Sugai S, Maruya E, Saji H. Association of a new allele of the TAP2 gene, TAP2*Bky2 (Val577), with susceptibility to Sjögren's syndrome. Arthritis Rheum. 1997;40:1685–1692. doi: 10.1002/art.1780400919. [DOI] [PubMed] [Google Scholar]
- Tapinos NI, Polihronis M, Thyphronitis G, Moutsopoulos HM. Characterization of the cysteine-rich secretory protein 3 gene as an early-transcribed gene with a putative role in the pathophysiology of Sjögren's syndrome. Arthritis Rheum. 2002;46:215–222. doi: 10.1002/1529-0131(200201)46:1<215::AID-ART10024>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- Kassan SS, Thomas TL, Moutsopoulos HM, Hoover R, Kimberly RP, Budman DR, Costa J, Decker JL, Chused TM. Increased risk of lymphoma in sicca syndrome. Ann Intern Med. 1978;89:888–892. doi: 10.7326/0003-4819-89-6-888. [DOI] [PubMed] [Google Scholar]
- Kaschner S, Hansen A, Jacobi A, Reiter K, Monson NL, Odendahl M, Burmester GR, Lipsky PE, Dorner T. Immunoglobulin Vlambda light chain gene usage in patients with Sjögren's syndrome. Arthritis Rheum. 2001;44:2620–2632. doi: 10.1002/1529-0131(200111)44:11<2620::aid-art442>3.0.co;2-m. [DOI] [PubMed] [Google Scholar]
- Elagib KE, Borretzen M, Jonsson R, Haga HJ, Thoen J, Thompson KM, Natvig JB. Rheumatoid factors in primary Sjögren's syndrome (pSS) use diverse VH region genes, the majority of which show no evidence of somatic hypermutation. Clin Exp Immunol. 1999;117:388–394. doi: 10.1046/j.1365-2249.1999.00963.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elagib KE, Borretzen M, Thompson KM, Natvig JB. Light chain variable (VL) sequences of rheumatoid factors (RF) in patients with primary Sjögren's syndrome (pSS): moderate contribution of somatic hypermutation. Scand J Immunol. 1999;50:492–498. doi: 10.1046/j.1365-3083.1999.00624.x. [DOI] [PubMed] [Google Scholar]
- Jonsson R, Skarstein K. Experimental models of Sjögren's Syndrome. In: Theofilopoulos AN, Bona CA, editor. The Molecular Pathology of Autoimmune Diseases. 2. New York: Taylor & Francis;; 2002. pp. 437–452. [Google Scholar]
- Johansson ACM, Nakken B, Sundler M, Lindqvist AK, Johannesson M, Alarcon-Riquelme M, Bolstad AI, Humphreys-Beher MG, Jonsson R, Skarstein K, Holmdahl R. The genetic control of sialadenitis versus arthritis in a NOD.QxB10.Q F2 cross. Eur J Immunol. 2002;32:243–250. doi: 10.1002/1521-4141(200201)32:1<243::AID-IMMU243>3.0.CO;2-X. [DOI] [PubMed] [Google Scholar]
- Johansson AC, Sundler M, Kjellen P, Johannesson M, Cook A, Lindqvist AK, Nakken B, Bolstad AI, Jonsson R, Alarcon-Riquelme M, Holmdahl R. Genetic control of collagen-induced arthritis in a cross with NOD and C57BL/10 mice is dependent on gene regions encoding complement factor 5 and FcgammaRIIb and is not associated with loci controlling diabetes. Eur J Immunol. 2001;31:1847–1856. doi: 10.1002/1521-4141(200106)31:6<1847::aid-immu1847>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Robinson CP, Yamachika S, Bounous DI, Brayer J, Jonsson R, Holmdahl R, Peck AB, Humphreys-Beher MG. A novel NOD-derived murine model of primary Sjögren's syndrome. Arthritis Rheum. 1998;41:150–156. doi: 10.1002/1529-0131(199801)41:1<150::AID-ART18>3.0.CO;2-T. [DOI] [PubMed] [Google Scholar]
- Brayer J, Lowry J, Cha S, Robinson CP, Yamachika S, Peck AB, Humphreys-Beher MG. Alleles from chromosomes 1 and 3 of NOD mice combine to influence Sjögren's syndrome-like autoimmune exocrinopathy. J Rheumatol. 2000;27:1896–1904. [PubMed] [Google Scholar]
- Cha S, Nagashima H, Brown VB, Peck A, Humphreys-Beher M. Two NOD Idd-associated intervals contribute synergistically to the development of autoimmune exocrinopathy (Sjögren's syndrome) on a healthy murine backgrund. Arthritis Rheum. 2002;46:1390–1398. doi: 10.1002/art.10258. [DOI] [PubMed] [Google Scholar]
- Boulard O, Fluteau G, Eloy L, Damotte D, Bedossa P, Garchon HJ. Genetic analysis of autoimmune sialadenitis in nonobese diabetic mice: a major susceptibility region on chromosome 1. J Immunol. 2002;168:4192–4201. doi: 10.4049/jimmunol.168.8.4192. [DOI] [PubMed] [Google Scholar]
- Nishihara M, Terada M, Kamogawa J, Ohashi Y, Mori S, Nakatsuru S, Nakamura Y, Nose M. Genetic basis of autoimmune sialadenitis in MRL/lpr lupus-prone mice: additive and hierarchical properties of polygenic inheritance. Arthritis Rheum. 1999;42:2616–2623. doi: 10.1002/1529-0131(199912)42:12<2616::AID-ANR16>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- Green JE, Hinrichs SH, Vogel J, Jay G. Exocrinopathy resembling Sjögren's syndrome in HTLV-1 tax transgenic mice. Nature. 1989;341:72–74. doi: 10.1038/341072a0. [DOI] [PubMed] [Google Scholar]
- Koike K, Moriya K, Ishibashi K, Yotsuyanagi H, Shintani Y, Fujie H, Kurokawa K, Matsuura Y, Miyamura T. Sialadenitis histologically resembling Sjögren syndrome in mice transgenic for hepatitis C virus envelope genes. Proc Natl Acad Sci USA. 1997;94:233–236. doi: 10.1073/pnas.94.1.233. [DOI] [PMC free article] [PubMed] [Google Scholar]