

Abstract
Hypericum perforatum (Hp) is known for possessing antidepressant and antiviral activities. Despite its use as an alternative to conventional antidepressants, the identification of the cytotoxic chemicals derived from this herb is incomplete. In this study, the cytotoxicity of Hp extracts prepared in solvents ranging in polarity, fractions of one extract, and purified compounds were examined in three cell lines. All extracts exhibited significant cytotoxicity; those prepared in ethanol (no hyperforin, 3.6 μM hypericin, and 134.6 μM flavonoids) showed between 7.7 and 77.4% cell survival (p < 0.0001 and 0.01), whereas the chloroform and hexane extracts (hyperforin, hypericin, and flavonoids not detected) showed approximately 9.0 (p < 0.0001) and 4.0% (p < 0.0001) survival. Light-sensitive toxicity was observed primarily with the ethanol extracts sequentially extracted following removal of material extracted in either chloroform or hexane. The absence of light-sensitive toxicity with the Hp extracts suggests that the hypericins were not playing a prominent role in the toxicity of the extracts.
Keywords: Hypericum perforatum, St. John’s wort, phototoxicity, cytotoxicity, hypericin, pseudohypericin, rutin, quercetin, quercitrin, isoquercitrin, hyperoside, chlorogenic acid, 8-isoprostane
ABBREVIATIONS USED : Hp, Hypericum perforatum; SJW, St. John’s wort; NCRPIS, North Central Regional Plant Introduction Station
INTRODUCTION
Hypericum perforatum (Hp) is a perennial herbaceous plant, also known as St. John’s wort (SJW) or Klamath weed, and its preparations have been used externally for wound healing and internally to relieve the symptoms of neurological disorders, namely, mild to moderate depression, since the early 1800s (1–4). Early use was concentrated primarily in Europe and Asia, where the plant originates, but the recent incursion of herb-based alternative medicines and the misconception that natural equals safe have broadly increased the market for Hp in Europe and the United States (1, 4). The efficacy of SJW as an antidepressant has been extensively studied along with its safety (1–6). The clinical and animal trials conducted thus far demonstrate Hp extracts to be just as effective as tricyclic antidepressants with fewer short-term side effects, but recent evidence of drug–herb interactions involving the hepatic cytochrome P450 enzyme system may be one of several adverse reactions to this herb that need to be elucidated before it can be deemed a safe form of alternative medicine (1–5, 7).
Because of the increasing public interest in this herb and its many intriguing biological activities, extensive analysis has been applied toward identifying the chemical composition of Hp in order to determine which compounds may be responsible for its antidepressant (1–5), antibacterial (1, 2), cytotoxic (8, 9), and antiviral (10–13) activities. However, most studies conducted have only evaluated the biological activities exhibited by ethanol and methanol extractions prepared from Hp plant material and the individual compounds identified within these fractions. Compounds common to many plant species have also been found in Hp, including an array of flavonoids, phenolic acids, proanthocyanidins, xanthones, and essential oils, but several compounds unique to this plant species were also discovered (1, 2, 4). A majority of the herb’s antidepressant activity have recently been attributed to hyperforin and adhyperforin, and these compounds also possess antibacterial and cell growth inhibitory activity (1–3, 14). The naphthodianthrone compounds, hypericin and its major metabolite pseudohypericin, have been shown to possess potent antiviral and cytotoxic properties upon light exposure (10–12, 15).
Despite the abundance of information collected thus far on the biological activities possessed by Hp, additional toxicological assessment is needed to ensure its safe use by the public. Little is known about the cytotoxicity of Hp extracts prepared in solvents other than water, ethanol, or methanol. Although most manufacturers currently prepare their Hp products via aqueous alcoholic extraction in various concentrations of either ethanol or methanol with water, new products and procedures are being developed every day to improve product effectiveness or reduce expense. Products obtained via plant material extraction contain a complex mixture of chemicals whose individual biological activities may be considerably different when present in a mixture (1).
The overarching objective of our research is to determine constituents of Hp that account for health-promoting and toxic activities of this dietary supplement with the goal of improving Hp as a dietary supplement. The goal of this initial study was to identify the role of constituents of Hp in the light- and dose-dependent cytotoxicity of Hp extracts prepared via either Soxhlet or room temperature extraction in solvents with a range of polarities from two different sources of dried plant material. The cytotoxicities of fractions obtained from an ethanol extract were also assessed along with several purified chemicals for the purpose of determining whether classes of compounds in the extracts or known reference chemicals may be contributing to the cytotoxicity exhibited by this herb.
MATERIALS AND METHODS
Safety
Organic solvents, such as chloroform and hexane, are toxic chemicals and should be properly handled using a fume hood.
Plant Extraction and Fractionation
Six grams of dried aerial portions of Hp plant material [Frontier Herb (Norway, IA) or North Central Regional Plant Introduction Station (NCRPIS) (Ames, IA)] was extracted by either the Soxhlet extraction method for 6 h or room temperature shaking for 24 h. The dried plant material obtained from Frontier Herb was harvested at the budding stage in Bulgaria in 1999, air-dried, and received at the Norway, IA, warehouse in 2001. The plant material provided by NCRPIS was harvested in October 2003, 139 days after transplantation into the fields, and the aerial portions consisted of the top of the plant harvested (above 12 in. off the ground). Once this plant material was harvested, it was dried at 38 °C in a forced air dryer for 8 days at a constant humidity and ground with a 40 mesh screen. Plant material was stored at −20 °C until extraction. The extracts were prepared in solvents that range in hydrophobicity, including ethanol, water, chloroform, hexane, or sequential extractions that involved extraction of the plant material first with either chloroform or hexane, which was removed, and the resulting residue was dried and re-extracted in ethanol. To elaborate on the sequentially extracted plant material, the extracts obtained from the first extraction with either chloroform or hexane were removed and the remaining residue was evaporated to dryness and re-extracted in ethanol. The extracts that were prepared with sequential extraction were then denoted either ethanol(–chloroform) or ethanol(–hexane) to represent ethanol extracts without the material removed by either chloroform or hexane, respectively. The ethanol extracts prepared from the Frontier Herb plant material were prepared in 100% ethanol via Soxhlet extraction and 70% ethanol via room temperature shaking to explore optimal extraction procedures. The NCRPIS material was extracted with 95% ethanol using the Soxhlet method because this approach was found to yield optimal extraction. These different ethanol extracts were not directly compared in this series of investigations. Once all extractions were complete, each was evaporated to dryness and dissolved in the minimum amount of dimethyl sulfoxide (DMSO) (Sigma-Aldrich, St. Louis, MO) necessary. All extracts were stored at −30 °C in the dark and used as stock solutions for treatment preparations. Each extract stock solution was added to media at 1% for the initial screening, which was conducted to allow comparison of their relative importance in the dried plant. The Soxhlet ethanol extract prepared from the Frontier Herb material was fractionated using a C18-affinity cartridge and eluded with increasing increments of acetonitrile (ACN) in water. Three fractions were chosen for cytotoxicity assessment, the 20% ACN fraction, a fraction containing the 25, 27, 30, and 40% ACN elutions combined, and a fraction containing the 50, 60, and 70% ACN elutions combined. All solvents used for extraction were high-performance liquid chromatography (HPLC) grade from Fisher Scientific Company, except the 100% ethanol.
Reference Compounds
The chemicals used for the identification and quantification of compounds within the Hp extracts include the following: hyperforin, hypericin (Molecular Probes, Eugene, OR); pseudohypericin (Calbiochem-Novabiochem, La Jolla, CA); chlorogenic acid, quercetin, and rutin (Fisher Scientific, Hanover Park, IL); and quercitrin, isoquercitrin, and hyperoside (ChromaDex, Santa Anna, CA).
The HPLC system was composed of Beckman System Gold 126 solvent module, model 508 autosampler, model 168 detector (Beckman Coulter, Inc., Fullerton, CA), and a RP-C18, 5 μm, 250 mm × 10 mm i.d. YMC-ODC-AM-303 column (YMC, Inc., Wilmington, NC). All Hypericum extracts were filtered through 0.45 μm poly(tetrafluoro-ethylene) filters (Alltech Associates Inc., Deerfield, IL) before injecting into the HPLC. Two HPLC methods were employed to identify and quantify the individual constituents in the Hypericum extracts, one for the flavonoid compounds and one for the hypericin compounds.
The percent repeatability, reproducibility, and minimum detection levels (μM) for the HPLC quantification of these compounds are provided in Table 1. The concentration of each reference compound (μM) within each of the Hp extracts that were assessed for cytotoxicity is provided in Tables 2 and 3.
Table 1.
Repeatability and Reproducibility of HPLC Analysis and Extraction of Hpa
| quantified standard compounds | hypericin | pseudohypericin | rutin | quercetin | hyperoside | quercitrin | isoquercitrin | chlorogenic acid |
|---|---|---|---|---|---|---|---|---|
| minimum detection levels (μM) | 9.3 | 6.1 | 2.2 | 3.0 | 1.8 | 1.5 | 2.8 | 13.2 |
| repeatability (%; ± SDcv) | ||||||||
| HPLC (no. of observations) | 3.94 ± 1.61 (12) | 3.96 ± 2.46 (12) | 1.52 ± 0.99 (12) | 1.19 ± 0.43 (12) | 7.21 ± 10.26 (12) | 2.17 ± 1.51 (12) | ||
| reproducibility (%; ± SDcv) | ||||||||
| Soxhlet (no. of observations) | 5.78 ± 2.69 (8) | 9.92 ± 4.72 (8) | 8.00 ± 5.52 (8) | 8.07 ± 3.94 (7) | 9.98 ± 2.61 (8) | 5.42 ± 2.40 (8) | 4.62 ± 2.86 (2) | |
| Shaking (no. of observations) | 6.15 ± 2.25 (4) | 6.40 ± 1.55 (4) | 7.35 ± 3.86 (4) | 7.62 ± 8.18 (4) | 19.49 ± 29.92 (4) | 10.72 ± 2.48 (2) | ||
The minimum detection levels listed for each reference chemical are the lowest concentration (μM) capable of detection by the HPLC quantification method. The repeatability and reproducibility data represent the percent coefficient of variation (CV) ± the standard deviation for each CV.
Table 2.
Compounds Identified and Quantified in Hp Extracts Prepared from Dried Plant Materiala
| Frontier Herb plant material | ||||||||
|---|---|---|---|---|---|---|---|---|
| μ M
|
||||||||
| quantified standard compounds | hypericin | pseudohypericin | rutin | quercetin | hyperoside | quercitrin | isoquercitrin | chlorogenic acid |
| Soxhlet extraction | ||||||||
| 100% EtOH | 364.7 ± 6.2 | 515.4 ± 16.4 | 6579.8 ± 50.7 | detectable | 3534.2 ± 17.9 | 894.9 ± 313.5 | 2446.0 ± 32.8 | |
| chloroform | undetectable amounts of all standards tested | |||||||
| hexane | undetectable amounts of all standards tested | |||||||
| 100% ethanol (–chloroform) | 335.0 ± 9.1 | 493.6 ± 7.9 | 6101.1 ± 5.1 | detectable | 3374.6 ± 35.7 | 674.9 ± 0.0 | 2244.8 ± 47.6 | |
| 100% ethanol (–hexane) | 339.9 ± 8.6 | 484.6 ± 14.9 | 4956.4 ± 110.0 | detectable | 2791.9 ± 25.4 | 588.7 ± 51.5 | 1868.2 ± 119.8 | |
| room temperature shaking | ||||||||
| 70% EtOH | 214.1 ± 7.9 | 194.3 ± 16.4 | 3106.0 ± 45.5 | detectable | 1951.4 ± 22.3 | 638.3 ± 30.1 | 988.1 ± 12.0 | |
| chloroform | undetectable amounts of all standards tested | |||||||
| hexane | undetectable amounts of all standards tested | |||||||
| 100% ethanol (–chloroform) | 217.0 ± 5.1 | 188.3 ± 17.9 | 3174.4 ± 25.3 | detectable | 2096.1 ± 19.5 | 573.8 ± 23.2 | 1167.5 ± 9.1 | |
| 100% ethanol (–hexane) | 202.2 ± 5.1 | 155.6 ± 9.6 | 2120.9 ± 20.2 | detectable | 1698.7 ± 17.4 | 395.4 ± 79.9 | 795.8 ± 17.4 | |
| H2O | undetectable amounts of all standards tested | |||||||
Standard compounds identified and quantified by HPLC analysis within the extract stock solutions prepared from 6 g of dried Frontier Herb Hp plant material (mean ± standard deviation). Detectable refers to a standard that was identified, but the amount present was too low to be quantified (n = 3).
Table 3.
Compounds Identified and Quantified in Hp Extracts Prepared via Soxhlet Extraction from Dried Plant Materiala
| NCRPIS plant material | ||||||||
|---|---|---|---|---|---|---|---|---|
| μM
|
||||||||
| quantified standard compounds | hypericin | pseudohypericin | rutin | quercetin | hyperoside | quercitrin | isoquercitrin | chlorogenic acid |
| 95% EtOH | ||||||||
| common | 240.8 ± 10.7 | 325.1 ± 3.4 | 852.3 ± 32.1 | detectable | 2331.0 ± 24.3 | 384.5 ± 9.6 | 1692.8 ± 16.4 | 2549.1 ± 4.5 |
| Helos | 277.5 ± 16.4 | 406.3 ± 13.4 | 1964.3 ± 37.6 | detectable | 2627.4 ± 45.5 | 482.7 ± 10.4 | 2256.7 ± 56.7 | |
| PI 325351 | 179.4 ± 3.4 | 199.2 ± 10.7 | 9602.6 ± 50.0 | detectable | 1939.5 ± 21.5 | 233.9 ± 9.6 | 1749.3 ± 11.3 | |
| PI 371528 | 179.4 ± 6.2 | 56.5 ± 3.0 | 8120.9 ± 70.5 | detectable | 376.6 ± 16.4 | 91.2 ± 8.6 | 1894.0 ± 139.0 | |
| chloroform | ||||||||
| common | undetectable amounts of all standards tested | |||||||
| Helos | undetectable amounts of all standards tested | |||||||
| PI 325351 | undetectable amounts of all standards tested | |||||||
| PI 371528 | undetectable amounts of all standards tested | |||||||
Standard compounds identified and quantified by HPLC analysis within the extract stock solutions prepared from 6 g of dried NCRPIS Hp plant material (mean ± standard deviation). Detectable refers to a standard that was identified, but the amount present was too low to be quantified (n = 3).
Cell Lines and Growth Conditions
The cytotoxicity of the Hp extracts was assessed in NIH3T3 mouse fibroblasts (ATCC, Rockville, MD), SW480 human colon cancer cells (ATCC), and HaCaT human keratinocytes. The NIH3T3 mouse fibroblast cell line is an immortalized fibroblast that is a common model for the assessment of cytotoxicity (16). The SW480 colon cancer cells represent a human exposure site after oral ingestion of Hp, whereas the HaCaT keratinocytes represent a human exposure site for the topical administration of Hp preparations. NIH3T3 and SW480 cell lines were cultured in low glucose Dulbecco’s modified Eagles medium (Sigma-Aldrich) supplemented with 3.0 and 3.7 g/L sodium bicarbonate (Sigma-Aldrich), respectively. The HaCaT cells were generously provided by Dr. Tim Bowden (Arizona Cancer Center, University of Arizona) and cultured in high glucose Dulbecco’s modified Eagles medium (4500 mg/L d-glucose) (Invitrogen, Carlsbad, CA) with 3.7 g/L sodium bicarbonate. All cell culture media were also supplemented with 100 UI/mL penicillin/streptomycin antibiotics (Invitrogen) and 10% fetal bovine serum (Invitrogen). The cells were maintained in 70% humidity with 5% CO2 at 37 °C until approximately 80% confluent in 75 cm2 flasks.
Cytotoxicity Screening Assay
Cytotoxic analysis was carried out using the Celltiter96 Aqueous One Solution Cell Proliferation Assay (Promega Corp., Madison, WI). Cells were plated into 48 well plates at 10000 cells/well and allowed to attach for 16–18 h before treatment. Light or dark treatments were randomly assigned to plates, and extract treatments were randomly assigned to wells within a plate. To screen the extracts for their effect on cytotoxicity, cells were treated with 1% of the stock extract solutions for 24 h using media and DMSO as solvent controls and 20 μM hypericin as the positive control. Treatments were performed under limited light conditions, and the plates were immediately exposed to either ambient light (~5.2 J/cm2) or dark conditions at room temperature for 30 min. Upon light exposure following treatment in the dark, cells were exposed to light provided by standard fluorescent lamps in the laboratory where all windows were shaded. Following the light or dark treatment period, the plates were returned to the dark at 37 °C for 24 h. Following the 24 h incubation period, treatment solutions were removed and fresh media and Celltiter96 dye were added for 3 h and 15 min, which was found to be the optimal incubation time for our system. The metabolized dye solutions were then transferred to 96 well plates for absorbance measurement at 490 nm, a wavelength found to not interfere with the excitation or absorption of light by the hypericin compounds. The number of viable cells for each treatment was compared to a standard curve of known cell densities and normalized to the solvent control. The Trypan Blue Exclusion assay (Sigma-Aldrich) was used to corroborate the results of the Celltiter 96 Aqueous One-Solution Cell Proliferation Assay.
The tetrazolium dyes used in assays such as the Celltiter 96 Aqueous One-Solution Cell Proliferation Assay have been known to interfere with some plant compounds (17). This was not an issue with the analysis reported here because all treatment solutions were removed from the cells prior to the addition of the Celltiter 96 dye and cell-free analysis of the treatment residues was found to not induce reduction of the dye (data not shown).
STATISTICAL ANALYSIS
The statistical analysis for the cytotoxicity data was completed using a split plot analysis of variance with plate to plate variation as the main plot error and the well to well variation as the split plot error. Experimental values were compared to the DMSO control, and results are shown with the data. Intergroup comparisons were made between the ethanol extracts and the hexane/chloroform extracts at the extract stock concentration obtained from extraction of 6 g of plant material (Table 4), but differences were not observed and results of these tests are not included. Inspection of the data suggested that some extracts exhibited more variability than others, so a heterogeneous error variances model was fit to the data (18).
Table 4.
Cytotoxicity of Extract Stock Solutions from Dried Frontier Herb Hp in Three Cancer Cell Linesa
| mean ± SE
|
||||||
|---|---|---|---|---|---|---|
| NIH3T3 mouse fibroblasts
|
SW480 human colon cancer cells
|
HaCaT human skin keratinocytes
|
||||
| extraction method and solvents (treatment concn of extracts in media) | light | dark | light | dark | light | dark |
| Soxhlet | ||||||
| 100% EtOH (1161 μg/mL) | 7.7** (±4.2) | 10.3** (±4.9) | 8.3** (±3.5) | 11.7** (±2.8) | 35.4** (±7.5) | 40.3** (±8.9) |
| chloroform (284 μg/mL) | 5.5** (±3.7) | 9.0** (±5.1) | 2.5** (±1.7) | 8.2** (±4.1) | 1.6** (±1.1) | 0.4** (±0.2) |
| hexane (166 μg/mL) | 3.1** (±2.0) | 4.1** (±2.6) | 1.5** (±1.3) | 2.9** (±2.5) | 0.3** (±0.3) | 0.3** (±0.3) |
| 100% ethanol (–chloroform) (589 μg/mL) | 38.1* (±3.4) | 49.0* (±3.3) | 55.1** ## (±2.7) | 77.2 (±1.2) | 35.6** ## (±6.8) | 77.4* (±9.9) |
| 100% ethanol (–hexane) (568 μg/mL) | 12.0** ## (± 4.6) | 32.5** (± 3.1) | 8.7** ## (± 2.7) | 32.4** (± 4.9) | 40.8** (± 11.8) | 50.4** (± 9.6) |
| room temperature shaking | ||||||
| 70% EtOH (740 μg/mL) | 11.1** (±9.1) | 12.5** (±7.4) | 2.5** (±0.9) | 14.2** (±5.5) | 42.8** (±8.9) | 34.3** (±10.6) |
| chloroform (174 μg/mL) | 34.8** (±6.5) | 33.1** (±6.7) | 26.8** (±11.8) | 47.6* (±5.2) | 8.5** (±5.1) | 12.1** (±2.9) |
| hexane (58 μg/mL) | 82.6 (±1.9) | 81.4 (±2.9) | 83.3 (±9.0) | 88.5 (±2.4) | 89.5 (±4.9) | 94.1 (±3.6) |
| 70% ethanol (–chloroform) (692 μg/mL) | 33.6** (±7.4) | 27.6** (±6.7) | 57.7** (±7.5) | 44.6** (±6.7) | 41.6** (±3.6) | 66.2* (±13.7) |
| 70% ethanol (–hexane) (556 μg/mL) | 29.7** (±8.5) | 35.1** (±6.1) | 25.0** (±4.9) | 39.2** (±8.2) | 32.9** (±7.8) | 45.4** (±9.4) |
| water (213 μg/mL) | 84.4* (±3.0) | 85.5* (±1.7) | 95.6 (±5.0) | 85.1 (±5.7) | 73.6 (±16.2) | 87.7 (±8.1) |
| 20 μM hypericin | 10.1** ## (±6.2) | 60.9** (±5.7) | 20.8** ## (±2.4) | 80.5 (±1.3) | 0.87** # (±0.4) | 53.2** (±10.9) |
Cytotoxicity (% survival as compared to vehicle control-treated cells) of Hp extracts screened via the Celltiter96 Aqueous One Solution Cytotoxicity assay (n = 3–5). All extract stock solutions were prepared from 6 g (water extract from 5 g) of dried plant material by either Soxhlet or room temperature extraction and included as 1% of the cell culture media. One set of chloroform and hexane extracts was sequentially extracted in ethanol:ethanol(–chloroform) or ethanol(–hexane). The treatment concentration listed for each extract or fraction (μg/mL) is the amount of extract residue obtained after extraction, diluted in DMSO, and used in the assay. A 20 μM concentration of hypericin was the positive control.
= p < 0.0001 and
= p < 0.01 significantly different cell growth survival as compared to DMSO solvent control.
= p < 0.0001 and
= p < 0.01 significantly different cell growth survival after exposure to 30 min of ambient light as compared to the dark incubation.
RESULTS
Identification and Quantification of Reference Chemicals within Dried Frontier Herb and NCRPIS H. perforatum Extracts
The presence and quantity of several reference chemicals known to be present within Hp were confirmed by HPLC analysis (Table 1) in the extracts prepared from the dried Frontier Herb (Table 2) and NCRPIS (Table 3) plant material. The compounds chosen for detection and quantification include the following: rutin, hyperoside, isoquercitrin, quercitrin, pseudohypericin, hypericin, chlorogenic acid, hyperforin, and quercetin (Tables 1–3). Except for hyperforin, which was not detected, the concentrations of the other compounds were found to be higher within the extracts prepared via Soxhlet extraction as compared to those prepared by shaking at room temperature (Table 2). None of the extracts prepared via Soxhlet extraction were found to contain detectable levels of hyperforin (Table 2). The extracts prepared by room temperature shaking for 24 h were prepared in the light, and because of the known instability of hyperforin upon exposure to light, hyperforin was not expected to be present in these extracts (19, 20). The ethanol, ethanol(–chloroform), and ethanol(–hexane) extracts contained similar concentrations of all compounds tested, whereas the chloroform and hexane extracts did not contain detectable amounts of any reference compound tested. The flavonoids were the most abundant compounds present with rutin, hyperoside, and isoquercitrin generally at the highest concentrations and quercitrin at much lower concentrations. Quercetin was detected but not at concentrations capable of quantification by HPLC analysis. The hypericins were present at lower concentrations than the flavonoids, chlorogenic acid was only detected in one extract, and no evidence of the presence of the hyperforin compounds was found. According to the HPLC analysis, it is likely that many other unidentified compounds similar in structure to the flavonoids and phenolic compounds are also present.
Cytotoxicity of the Frontier Herb H. perforatum Plant Material
The cytotoxicity of extractions prepared from Frontier Herb Hp plant material was screened in three cell lines, NIH3T3 mouse fibroblasts, SW480 human colon cancer cells, and HaCaT human keratinocytes using stock solution obtained from extraction of 6 g of plant material (Table 4). All ethanol and chloroform extracts significantly reduced growth in all three cell lines independent of light exposure. The hexane extract prepared by Soxhlet extraction also significantly reduced cell growth in all three cell lines independent of light exposure, but the hexane extract prepared via room temperature shaking was not toxic. Water was only used as an extraction solvent with room temperature shaking and showed significant toxicity solely in the mouse fibroblasts. The extracts having undergone a sequential extraction via the Soxhlet method in chloroform or hexane first, the residue of these first extractions evaporated to dryness and re-extracted in ethanol [ethanol(–chloroform) and ethanol(–hexane)] showed slightly different effects on cell growth than these extracts prepared by room temperature shaking. The Soxhlet ethanol(–chloroform) extract was significantly cytotoxic in all three cells lines and exhibited significant light-sensitive cytotoxicity in the SW480 cells and HaCaT cells. The Soxhlet ethanol(–hexane) extract was also significantly cytotoxic in all cell lines, showing light sensitivity in the NIH3T3 and SW480 cells but not the HaCaT cells. Unlike the Soxhlet ethanol(–chloroform) and ethanol(–hexane) extracts, those prepared by room temperature shaking showed significant cell growth reduction independent of light exposure in all three cell lines.
As mentioned previously, the Trypan blue exclusion assay was used as a method for testing the validity of the Celltiter96 Aqueous One Solution Cell Proliferation assay in our system using the extracts prepared from Frontier Herb material on the HaCaT keratinocytes (data not shown). Similar cell growth reduction was observed between the two assays with the extracts showing either potent toxicity or little to no influence on cell growth. However, the extracts in which only a moderate amount of toxicity was observed using the Celltiter 96 assay showed more potent reduction of cell growth using the Trypan blue exclusion assay.
The cytotoxicity of the Soxhlet and room temperature extracts prepared from the Frontier Herb material was also assessed at 100 and 10 μg/mL with the HaCaT keratinocytes in order to compare the activity of these extracts at the same concentrations (Table 5). The HaCaT keratinocytes showed the greatest difference in cytotoxicity across the extracts, so this cell line was chosen as the model for the remaining toxicity studies. The Soxhlet extract prepared via extraction in ethanol exhibited significant toxicity after light exposure (p < 0.01) but not in the dark. The Soxhlet chloroform (p < 0.0001) and hexane (p < 0.0001) extracts continued to show significant cell growth reduction at 100 μg/mL independent of light exposure. The Soxhlet ethanol(–hexane) extract lost the light sensitivity exhibited at its undiluted concentration, but it exhibited significant cytotoxicity (p < 0.01) only after light exposure. The ethanol extract prepared by room temperature shaking exhibited significant toxicity at 100 μg/mL but only after incubation in the dark. The extract prepared in chloroform was the only room temperature shaking extract capable of significantly reducing cell growth (p < 0.0001) at 100 μg/mL independent of light exposure. No extract exhibited significant toxicity at 10 μg/mL plant material.
Table 5.
Cytotoxicity of Diluted Extracts Prepared from 6 g of Dried Frontier Herb Hp Material in HaCaT Keratinocytesa
| Soxhlet | ||
|---|---|---|
| HaCaT human skin keratinocytes % control survival (mean ± SE)
|
||
| extraction method and solvents (treatment concn of extracts in media) | light | dark |
| 100% ethanol | ||
| 100 μg/mL | 64.1* (±5.5) | 72.6 (±2.8) |
| 10 μg/mL | 67.8 (±10.3) | 75.4 (±6.7) |
| chloroform | ||
| 100 μg/mL | 16.4** (±6.3) | 20.6** (±6.3) |
| 10 μg/mL | 94.2 (±2.5) | 94.0 (±0.9) |
| hexane | ||
| 100 μg/mL | 0.2** (±0.2) | 1.8** (±1.4) |
| 10 μg/mL | 103.1 (±5.6) | 108.5 (±6.3) |
| 100% ethanol (–chloroform) | ||
| 100 μg/mL | 94.9 (±8.8) | 90.0 (±9.9) |
| 10 μg/mL | 97.0 (±6.8) | 93.2 (±5.4) |
| 100% ethanol (–hexane) | ||
| 100 μg/mL | 76.2* (±4.2) | 83.2 (±9.4) |
| 10 μg/mL | 100.7 (±4.5) | 94.7 (±12.8) |
| Room Temperature Shaking | ||
|---|---|---|
| HaCaT human skin keratinocytes % control survival (mean ± SE)
|
||
| extraction method and solvents (treatment concn of extracts in media) | light | dark |
| 70% EtOH | ||
| 100 μg/mL | 85.6 (±8.8) | 73.1* (±3.8) |
| 10 μg/mL | 104.4 (±7.2) | 92.6 (±8.3) |
| chloroform | ||
| 100 μg/mL | 57.6** (±5.7) | 56.9** (±9.0) |
| 10 μg/mL | 118.3 (±4.2) | 112.6 (±5.7) |
| hexane | ||
| 10 μg/mL | 97.6 (±5.8) | 88.7 (±8.2) |
| 70% ethanol (–chloroform) | ||
| 100 μg/mL | 97.3 (±7.4) | 96.3 (±12.0) |
| 10 μg/mL | 105.4 (±9.9) | 95.8 (±12.1) |
| 70% ethanol (–hexane) | ||
| 100 μg/mL | 79.7 (±2.6) | 80.8 (±6.7) |
| 10 μg/mL | 98.0 (±4.0) | 96.8 (±15.2) |
Cytotoxicity (% survival as compared to vehicle control-treated cells) of the Hp extracts and ethanol fractions of the chloroform and hexane extracts shown in Table 1 diluted to 100 and 10 μg/mL extract residue in DMSO and included at 1% in media with the HaCaT human keratinocytes (n = 6–7). The room temperature hexane extract was only tested at 10 μg/mL because as noted in Table 4, the undiluted extract stock concentration was less than 100 μg/mL.
= p < 0.0001 and
= p < 0.01 significantly different cell growth survival as compared to DMSO solvent control.
To further elucidate the toxic effect of the extracts on the HaCaT keratinocytes, the dose–response toxicity was assessed for the most active Soxhlet extracts prepared from the Frontier Herb material and lethal concentrations were calculated for 50% of the cell population using nonlinear regression curves with GraphPad Prism software. The cytotoxicity dose–response curve generated from the Soxhlet ethanol extract after light exposure showed significant toxicity at 1161 (p < 0.01), 581 (p < 0.0001), 500 (p < 0.0001), and 100 μg/mL (p < 0.01), while the curve for cells not exposed to light showed somewhat less toxicity at 581 (p < 0.01) and 500 μg/mL (p < 0.01) (Figure 1A). However, the extract did not exhibit significantly different toxicity in the light as compared to the dark at any concentration. Because of the slightly greater toxicity of these two concentrations of the ethanol extract in the light, the error surrounding the LC50 concentrations obtained from the curves generated for this extract were larger than that obtained from the other extracts (Table 6). A dose–response in toxicity was exhibited by the ethanol(–chloroform) extract exposed to light; however, the toxicity of this extract in the dark was not lethal for more than 50% of the cell population, so the estimated LC50 value for these data was not obtained (Figure 1B). This extract demonstrated significantly greater cytotoxicity in the light than in the dark (p < 0.0001). The dose–response curves and LC50 values generated for the Soxhlet extracts prepared in chloroform or hexane did not differ between exposure to either light or dark (Figure 1C,D).
Figure 1.
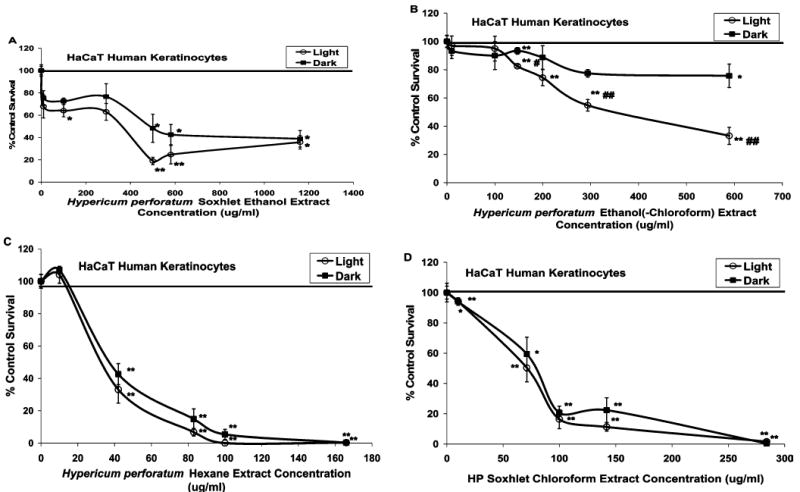
Cytotoxicity of diluted Frontier Herb Hp Soxhlet ethanol and chloroform extracts. Cytotoxicity (mean % control survival as compared to vehicle control ± SE) of diluted Hp extracts prepared via Soxhlet extraction in (A) ethanol, (B) sequential extraction in chloroform first followed by evaporation to dryness of this first extraction and re-extraction in ethanol, (C) hexane, and (D) chloroform (n = 3–6). A 20 μM concentration of hypericin is the positive control showing 8.4 (±4.3)% survival after light exposure and 73.5 (±2.2)% survival relative to the control in the dark, which are significantly different from each other. ** = p < 0.0001 and * = p < 0.01 significantly different cell growth survival as compared to DMSO solvent control. ## = p < 0.0001 and # = p < 0.01 significantly different cell growth survival after exposure to 30 min of ambient light as compared to the dark incubation.
Table 6.
Estimated Lethal Concentration for 50% of the HaCaT Human Keratinocyte Population for the Frontier Herb Extracts Shown in Figure 1a
| LC50 (μg/mL) (mean with 95% CI)
|
||
|---|---|---|
| extraction method and solvents | light | dark |
| 100% EtOH | 145 (58–360) | 552 (254–1203) |
| ethanol (–chloroform) | 248 (165–372) | b |
| chloroform | 66 (59–75) | 66 (49–90) |
| hexane | 36 (29–44) | 39 (33–45) |
Lethal concentration for 50% of the HaCaT human keratinocyte population for the more active Soxhlet extracts prepared from dried Frontier Herb Hp material included at 1% in media (dose–response curves shown in Figure 1). Calculated using nonlinear regression with GraphPad Prism software.
Toxicity was not effective for more than 50% of the cell population.
Cytotoxicity of the NCRPIS H. perforatum Plant Material
The cytotoxicity of extracts prepared from dried aerial Hp material provided by the NCRPIS, consisting of two commercial cultivars and two accessions, extracted by Soxhlet extraction with either ethanol or chloroform, was assessed using the Celltiter96 assay in the human keratinocytes (Table 7). Despite the lower yield in plant material obtained during chloroform extraction of this material as compared to that obtained during ethanol extraction, the chloroform extracts were more toxic than the ethanol extracts. All NCRPIS extracts showed significant cytotoxicity when tested at the concentration of extract in stock solution obtained from extraction of 6 g of plant material. All ethanol extracts, except the PI 371528 accession, retained the ability to significantly reduce cell growth upon dilution of the extract concentration to 100 μg/mL, but activity was lost at 10 μg/mL. The only extract that exhibited significant light sensitivity was prepared from the common commercial cultivar in the ethanol at the highest concentration of the extract examined. The chloroform extracts continued to exhibit significant cytotoxicity at all extract concentrations examined. Both NCRPIS accessions prepared in chloroform, PI 325351 and PI 371528, showed significant light sensitivity at the highest extract concentrations but lost light sensitivity at all other dilutions examined.
Table 7.
Cytotoxicity of Hp Extracts Prepared by Soxhlet Extraction from Dried NCRPIS Materiala
| HaCaT human keratinocytes % control survival (mean ±SE) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| common (μg/mL) | light | dark | Helos (μg/mL) | light | dark | PI 325351 (μg/mL) | light | dark | PI 371528 (μg/mL) | light | dark |
| Soxhlet extraction and solvents: 95% EtOH | |||||||||||
| 607 | 20.4**# (±1.0) | 31.7** (±3.7) | 609 | 20.2** (±1.2) | 26.6** (±4.3) | 885 | 26.8** (±3.0) | 34.5** (±7.2) | 1048 | 27.3** (±3.4) | 33.2** (±4.6) |
| 100 | 46.0** (±6.4) | 47.9** (±5.1) | 100 | 22.9** (±10.2) | 25.6** (±5.2) | 100 | 66.4* (±11.7) | 56.8* (±10.7) | 100 | 103.1 (±9.5) | 88.6 (±2.2) |
| 10 | 100.1 (±8.5) | 87.3 (±2.5) | 10 | 58.2 (±15.5) | 50.9 (±13.4) | 10 | 98.0 (±14.2) | 87.6 (±8.2) | 10 | 104.4 (±5.0) | 93.5 (±2.9) |
| Soxhlet extraction and solvents: chloroform | |||||||||||
| 120 | 5.8** (±1.6) | 8.5** (±2.6) | 153 | 9.8** (±1.9) | 8.2** (±3.3) | 124 | 22.5**## (±0.9) | 33.6** (±1.0) | 97 | 26.9**# (±1.5) | 33.3** (±1.7) |
| 100 | 14.8** (±9.1) | 15.5** (±6.7) | 100 | 24.6** (±6.0) | 29.2** (±3.3) | 100 | 20.4** (±4.4) | 26.4** (±3.6) | |||
| 10 | 44.1** (±6.7) | 40.2** (±3.2) | 10 | 36.5** (±4.7) | 37.6** (±3.0) | 10 | 80.2** (±1.2) | 76.4* (±5.0) | 10 | 82.7** (±2.9) | 81.9* (±3.2) |
Cytotoxicity (% survival as compared to vehicle control-treated cells) of extracts prepared from 6 g of dried Hp aerial plant cultivars or accessions provided by the NCRPIS at Iowa State University and included at 1% in media (n = 3–6). A 20 μM concentration of hypericin was used as the positive control, showing percent survival as compared to the solvent control equal to 0.91** (±0.5) after light exposure and 78.4 (±9.4) in the dark, which are significantly different from each other.
= p < 0.0001 and
= p < 0.01 significantly different cell growth survival as compared to DMSO solvent control.
= p < 0.0001 and
= p < 0.01 significantly different cell growth survival after exposure to 30 min of ambient light as compared to the dark incubation.
Cytotoxicity of the Fractioned Frontier Herb H. perforatum Extract Prepared Via Soxhlet Extraction in 100% Ethanol
The Soxhlet ethanol extract prepared from the Frontier Herb material was chosen for fractionation because HPLC analysis confirmed that it generally contained the greatest concentrations of all detectable reference chemicals. Fractionation of this extract was performed in order to separate the classes of compounds present within this extract according to hydrophobicity and determine which group of chemicals possessed the greatest amount of toxicity and phototoxicity. Each of the three fractions obtained via C18 column separation showed significant light-independent cytotoxicity at the concentration obtained after fractionation of the extract (Table 8). Fraction 1, the 20% ACN elution, demonstrated only a 50–60% reduction in cell growth at its highest concentration, whereas fractions 2 and 3, each containing different combinations of the ACN elutions, exhibited significantly more cytotoxicity at the highest concentrations tested. Diluted concentrations of fractions 2 and 3 continued to show significant toxicity independent of light exposure despite the confirmed presence of pseudohypericin in fraction 2 and hypericin in fraction 3.
Table 8.
Cytotoxicity of Fractioned Portions of the Frontier Herb Hp Soxhlet Ethanol Extracta
| HaCaT human keratinocytes % control survival(mean ±SE) | ||
|---|---|---|
| light | dark | |
| fraction 1 | ||
| 209 μg/mL | 41.9** (±1.8) | 44.9** (±3.0) |
| fraction 2 | ||
| 320 μg/mL | 19.1** (±3.9) | 23.1** (±4.0) |
| 160 μg/mL | 13.2** (±2.7) | 23.5** (±3.4) |
| fraction 3 | ||
| 194 μg/mL | 0.7** (±0.6) | 1.9** (±1.5) |
| 97 μg/mL | 4.8** (±2.0) | 9.9** (±5.9) |
| 48 μg/mL | 43.6** (±4.4) | 48.4** (±5.3) |
Cytotoxicity (% survival as compared to vehicle control-treated cells) of the C18 affinity column fractions derived from the Hp Soxhlet ethanol extract (n = 4–9). Fraction 1, 20% ACN elution; fraction 2, combined 27, 30, and 40% ACN elutions; and fraction 3, combined 50, 60, and 70% ACN elutions. Extracts were added to media at 1%, and 20 μM hypericin was used as the positive control, showing 8.4 (±4.3)% survival after light exposure and 73.5 (±2.2)% survival relative to the control in the dark, which are significantly different from each other.
= p < 0.0001 and
= p < 0.01 significantly different cell growth survival as compared to DMSO solvent control.
Cytotoxicity of the Reference Chemicals Identified within the H. perforatum Extractions
The cytotoxicity of the chemicals chosen for identification within the Hp extracts was assessed in the HaCaT keratinocytes at concentrations roughly equal to what was found to be present within the extracts. No compound tested showed significant cytotoxicity, except hypericin and pseudohypericin, which exhibited significant light-sensitive toxicity as well as some toxicity in the dark (Figure 2).
Figure 2.
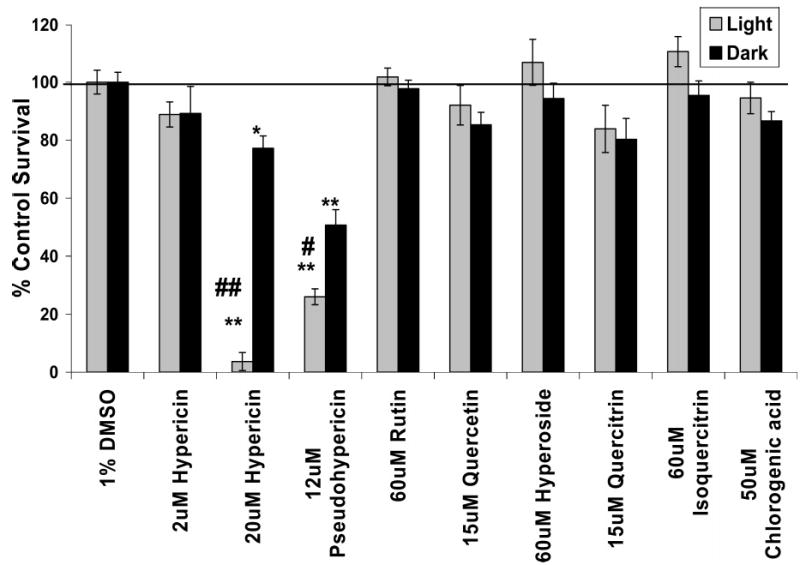
Cytotoxicity of standard compounds known to be present within the Hp plant. Cytotoxicity (mean % control survival as compared to vehicle control ± SE) of purified or synthesized compounds identified within the Hp extracts on the HaCaT human keratinocytes (n = 3–6). A 20 μM concentration of hypericin is the positive control. ** = p < 0.0001 and * = p < 0.01 significantly different cell growth survival as compared to DMSO solvent control. ## = p < 0.0001 and # = p < 0.01 significantly different cell growth survival after exposure to 30 min of ambient light as compared to the dark incubation.
DISCUSSION
Many studies have assessed the toxicity of Hp methanol or ethanol extracts as well as several purified compounds known to be present within this plant species (8, 9, 20–24), but to our knowledge, no studies have been conducted to assess the toxicity of extracts prepared in other solvents. Understanding the toxic potential of all constituents present within Hp is crucial because of its widespread use as an alternative medicine for depression. This evaluation involves identifying and assessing the biological activities of constituents obtained from extraction in both polar and nonpolar solvents. In this study, several Hp extracts prepared from two sources of dried plant material and extracted in several solvents ranging in hydrophobicity, fractions prepared from one ethanol extract, and several reference compounds were tested for cytotoxicity in an effort to determine the role of known compounds to the toxicity of this herb. Three different cell lines were used to screen the toxicity of the extract stock solutions at the concentration obtained from extraction of 6 g of plant material. The NIH3T3 fibroblasts are an immortalized cell line, whereas the HaCaT keratinocytes and SW480 colon cancer cells are both cancer cell lines. Immortalized or cancer cell lines were used because of their enhanced growth rate in culture, which was essential for such a large screening study. In general, the immortalized NIH3T3 fibroblasts exhibited the greatest sensitivity to Hypericum extract cytotoxicity. Because the light-induced cytotoxicity of hypericin and pseudohypericin, unique constituents in Hp, has been previously reported (22, 23, 25–27), the cytotoxicity of all extracts, fractions, and reference chemicals was conducted under both ambient light and dark conditions. Hypericin was used as a positive control, and 20 μM was found to kill between 80 and 100% of the cells after light exposure, so this concentration was used to ensure consistency in the cytotoxicity between the assays.
Quantification of several chemicals within the extracts revealed similar amounts of the identified chemicals in all ethanol extracts but no detectable amounts within the chloroform or hexane extracts (Tables 1–3). Therefore, Hp extracts containing the light-sensitive hypericin and pseudohypericin compounds were expected to exhibit greater cytotoxicity after light exposure than in the dark due to their production of singlet oxygen following photoexcitation (12, 26–29). Despite containing between 1.8 and 3.7 μM hypericin and between 0.6 and 5.2 μM pseudohypericin (these concentrations are 1% of the quantities listed in Tables 2 and 3, representative of the amount of the extract stock solutions added to the cells at 1% of the treatment media), significant light-sensitive toxicity was generally only observed in the ethanol(–chloroform) and ethanol(–hexane) extracts prepared from the Frontier Herb material (Table 4) and with one ethanol extract prepared from the NCRPIS material (Table 7). Similar concentrations of pure hypericin as found within the extracts were also assessed for cytotoxicity in the HaCaT keratinocytes; 2 μM hypericin caused approximately 85% cell survival as compared to the solvent control, and 5 μM hypericin caused between 60 and 70% cell survival as compared to the solvent control. The same concentrations of pure pseudohypericin were found to exhibit slightly less cytotoxicity after light exposure than hypericin. Therefore, the extracts containing higher concentrations of hypericin and pseudohypericin, such as the Soxhlet ethanol extract prepared from the Frontier Herb plant material, were expected to exhibit more toxicity in the light than in the dark. The general absence of light-sensitive toxicity exhibited by the ethanol extracts was counterintuitive due to the confirmed presence of photoactivated compounds within these extracts, but other studies have also reported an absence of light-sensitive toxicity at certain concentrations of Hp commercial extracts prepared in aqueous solvents (8, 9). The light-independent toxicity exhibited by these Hp extracts may suggest the presence of unidentified chemicals capable of reducing the phototoxicity exhibited by the hypericin compounds and the presence of compounds capable of inducing significant toxicity regardless of light exposure. The presence of compounds that attenuate hypericin’s light-dependent toxicity was further suggested by studies where hypericin was added to selected Hypericum extracts (Schmitt et al., submitted for publication).
As stated previously, no detectable levels of any reference chemical were found in the chloroform and hexane extracts; therefore, it was not surprising that these extracts generally did not exhibit light-sensitive toxicity. However, two chloroform extracts prepared from the NCRPIS material did exhibit significant light-sensitive toxicity. The inability to detect any of the known chemicals in the chloroform and hexane extracts indicates that any light-sensitive toxicity exhibited by these extracts was probably not due to the hypericin compounds.
The dose–response cytotoxicity curves generated for the Soxhlet ethanol, ethanol(–chloroform), and chloroform extracts also demonstrated the previously mentioned light-sensitive characteristics of these extracts. The ethanol(–chloroform) extract (Figure 1B) exhibited significant light-sensitive toxicity, whereas the ethanol extract (Figure 1A) showed a trend toward light sensitivity. Neither the chloroform nor the hexane extracts demonstrated light-sensitive toxicity at any of the extract concentrations tested (Figure 1C,D).
The toxicity exhibited by the chloroform and hexane extracts was substantially greater than that observed with the ethanol extracts, despite the lesser amount of extraction residue obtained with these extractions as compared to extraction with ethanol. For example, the chloroform and hexane extracts prepared from the Frontier Herb material maintained significant toxicity when diluted to 100 μg/mL, but the ethanol extracts lost all significant toxicity upon dilution to 100 μg/mL (Table 5). Similarly, the chloroform extracts prepared from the NCRPIS material retained significant toxicity when diluted to 100 and 10 μg/mL, but only two of the ethanol extracts were still significantly toxic at 100 μg/mL, and they completely lost toxicity at 10 μg/mL (Table 7). To reiterate this point, the dose–response curves generated for the Frontier Herb Soxhlet extracts demonstrate the lower concentrations of the chloroform and hexane extracts needed to kill 50% of the cell population as compared to the ethanol extracts. This indicates that the compounds present in the chloroform and hexane extracts exhibit more potent toxicity than those present within the ethanol extracts.
The next step taken to decipher what classes of compounds may be toxic within Hp was to assess the toxicity of fractions obtained from the Frontier Herb Soxhlet ethanol extract, which was chosen for fractionation because it generally contained the highest concentrations of all of the reference chemicals tested. The light-sensitive toxicity of these three fractions was expected to vary, with the first fraction possessing no light sensitivity but increasing sensitivity in fractions 2 and 3 due to the presence of pseudohypericin and hypericin, respectively. Unexpectedly, none of the fractions exhibited significant light-sensitive toxicity despite the presence of the hypericins within two, indicating either a lack of sufficient quantities of the hypericin compounds or the ability of unidentified compounds within the fractions to attenuate hypericin’s light-induced toxicity. All three fractions possessed a significant amount of toxicity, with the fraction containing only one identified chemical, hypericin, possessing the greatest toxicity independent of light exposure. Therefore, the fractionation scheme designed to separate the more hydrophobic hypericin compounds from the flavonoids and phenolic acids did not adequately pinpoint the classes of compounds within this extract that may be responsible for its toxicity or what compounds may be contributing to the attenuation of the light-induced toxicity of the hypericins.
To compliment the toxicity data obtained from Hp extracts and fractions, several pure or synthesized chemicals that were used for determining the presence of known Hp constituents within the extracts were also assessed for cytotoxicity. The pseudohypericin and hypericin compounds possessed significant toxicity after light exposure as well as some toxicity in the dark. The concentration of hypericin needed to induce a significant amount of toxicity after light exposure in the HaCaT keratinocytes was higher in this study than has been reported for other cell lines (16, 23) and other studies using the HaCaT cells (30). Methodological differences between the studies in which fibroblasts (16) and HeLa cells (23) were used, namely, preincubation of cells with hypericin prior to irradiation and use of much greater amounts of light energy, may provide an explanation for the greater concentration of hypericin needed in this study to induce a significant amount of light-induced toxicity. Another explanation may be due to the fact that HaCaT keratinocytes are known to be more resistant to oxidative stress than other types of immortalized or cancer cells (31). However, a study conducted by Schempp et al. showed that 0.5 μM hypericin killed approximately 50% of HaCaT keratinocytes after exposure to 5 J/cm2 visible light (30). The HaCaT keratinocytes in the Schempp study were preincubated with hypericin for 2 h prior to irradiation and treated with hypericin dissolved in a PBS/DMSO solution instead of cell culture media (30). Again, these methodological differences may explain the greater susceptibility of the HaCaT keratinocytes in the Schempp study as compared to this investigation. None of the flavonoids tested nor chlorogenic acid showed significant cytotoxicity, indicating that independently these chemicals are not responsible for the toxicity exhibited by the extracts in which they are present. Although flavonoids, such as quercetin, have been found to exhibit cytotoxicity toward other types of immortalized or cancer cell lines, the effect is not consistent and may be cell specific (21, 24, 32).
In conclusion, the cytotoxicity exhibited by the Hp extracts prepared for this study differed depending upon extraction procedure and solvent, but extracts generally did not possess significant light-sensitive toxicity despite the presence of the hypericin compounds. Fractionation of one ethanol extract did not provide insight into the classes of compounds that may be contributing to toxicity, and the individual reference compounds identified within the extracts showed that no one compound clearly contributed to the toxicity exhibited by the extracts.
Acknowledgments
We thank Dr. Philip Dixon for statistical support and Dr. Timothy Bowden for graciously providing the HaCaT human keratinocytes for this study. We acknowledge the gift of the Hypericum plant material from Frontier Herb (Norway, IA) and the contribution of several accessions of Hp by the NCRPIS at Iowa State University (Ames, IA).
Footnotes
This publication was made possible by Grant P01 ES012020 from the National Institute of Environmental Health Sciences (NIEHS) and the Office of Dietary Supplements (ODS), NIH. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIEHS, NIH.
References
- 1.Bilia AR, Gallori S, Vincieri FF. St. John’s wort and depression: Efficacy, safety and tolerability-an update. Life Sci. 2002;70(26):3077–3096. doi: 10.1016/s0024-3205(02)01566-7. [DOI] [PubMed] [Google Scholar]
- 2.Greeson JM, Sanford B, Monti DA. St. John’s wort (Hypericum perforatum. A review of the current pharmacological, toxicological, and clinical literature. Psychopharmacology (Berlin) 2001;153(4):402–414. doi: 10.1007/s002130000625. [DOI] [PubMed] [Google Scholar]
- 3.Kasper S. Hypericum perforatum–A review of clinical studies. Pharmacopsychiatry. 2001;34(Suppl 1):S51–S55. doi: 10.1055/s-2001-15467. [DOI] [PubMed] [Google Scholar]
- 4.Barnes J, Anderson LA, Phillipson JD. St. John’s wort (Hypericum perforatum L.): A review of its chemistry, pharmacology and clinical properties. J Pharm Pharmacol. 2001;53(5):583–600. doi: 10.1211/0022357011775910. [DOI] [PubMed] [Google Scholar]
- 5.Rodriguez-Landa JF, Contreras CM. A review of clinical and experimental observations about antidepressant actions and side effects produced by Hypericum perforatum extracts. Phytomedicine. 2003;10(8):688–699. doi: 10.1078/0944-7113-00340. [DOI] [PubMed] [Google Scholar]
- 6.Linde K, Berner M, Egger M, Mulrow C. St. John’s wort for depression: Meta-analysis of randomised controlled trials. . Br J Psychiatry. 2005;186:99–107. doi: 10.1192/bjp.186.2.99. [DOI] [PubMed] [Google Scholar]
- 7.Henderson L, Yue QY, Bergquist C, Gerden B, Arlett P. St. John’s wort (Hypericum perforatum): Drug interactions and clinical outcomes. Br J Clin Pharmacol. 2002;54(4):349–356. doi: 10.1046/j.1365-2125.2002.01683.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bernd A, Simon S, Ramirez BA, Kippenberger S, Diaz AJ, Miquel J, Villalba Garcia JF, Pamies MD, Kaufmann R. Phototoxic effects of Hypericum extract in cultures of human keratinocytes compared with those of psoralen. Photochem Photobiol. 1999;69(2):218–221. doi: 10.1562/0031-8655(1999)069<0218:peoeic>2.3.co;2. [DOI] [PubMed] [Google Scholar]
- 9.Hostanska K, Reichling J, Bommer S, Weber M, Saller R. Aqueous ethanolic extract of St. John’s wort (Hypericum perforatum L.) induces growth inhibition and apoptosis in human malignant cells in vitro. Pharmazie. 2002;57(5):323–331. [PubMed] [Google Scholar]
- 10.Meruelo D, Lavie G, Lavie D. Therapeutic agents with dramatic antiretroviral activity and little toxicity at effective doses: Aromatic polycyclic diones hypericin and pseudohypericin. Proc Natl Acad Sci USA. 1988;85(14):5230–5234. doi: 10.1073/pnas.85.14.5230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kraus GA, Melekhov A, Carpenter S, Wannemuhler Y, Petrich J. Phenanthrenequinone antiretroviral agents. Bioorg Med Chem Lett. 2000;10(1):9–11. doi: 10.1016/s0960-894x(99)00589-2. [DOI] [PubMed] [Google Scholar]
- 12.Carpenter S, Kraus GA. Photosensitization is required for inactivation of equine infectious anemia virus by hypericin 5. Photochem Photobiol. 1991;53(2):169–174. doi: 10.1111/j.1751-1097.1991.tb03919.x. [DOI] [PubMed] [Google Scholar]
- 13.Lenard J, Rabson A, Vanderoef R. Photodynamic inactivation of infectivity of human immunodeficiency virus and other enveloped viruses using hypericin and rose bengal: Inhibition of fusion and syncytia formation. Proc Natl Acad Sci USA. 1993;90(1):158–162. doi: 10.1073/pnas.90.1.158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Laakmann G, Schule C, Baghai T, Kieser M. St. John’s wort in mild to moderate depression: The relevance of hyperforin for the clinical efficacy 21. Pharmacopsychiatry. 1998;31(Suppl 1):54–59. doi: 10.1055/s-2007-979346. [DOI] [PubMed] [Google Scholar]
- 15.Carpenter S, Fehr MJ, Kraus GA, Petrich JW. Chemiluminescent activation of the antiviral activity of hypericin: A molecular flashlight. Proc Natl Acad Sci USA. 1994;91(25):12273–12277. doi: 10.1073/pnas.91.25.12273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yu H, Wolford ST, Kegode R, Zhao W, Osweller GD. Hypericin-induced phototoxicity in cultured fibroblasts and swine erythrocytes 1. Photochem Photobiol. 1996;64(1):168–173. doi: 10.1111/j.1751-1097.1996.tb02438.x. [DOI] [PubMed] [Google Scholar]
- 17.Bruggisser R, von DK, Jundt G, Schaffner W, Tullberg-Reinert H. Interference of plant extracts, phytoestrogens and antioxidants with the MTT tetrazolium assay. Planta Med. 2002;68(5):445–448. doi: 10.1055/s-2002-32073. [DOI] [PubMed] [Google Scholar]
- 18.Littell, R. C., Milliken, G. A., Stroup, W. W., Wolfinger, R. D. SAS System for Mixed Models; SAS Institute Inc.: Cary, NC, 1996; 633 pp.
- 19.Bilia AR, Bergonzi MC, Morgenni F, Mazzi G, Vincieri FF. Evaluation of chemical stability of St. John’s wort commercial extract and some preparations. Int J Pharm. 2001;213(1–2):199–208. doi: 10.1016/s0378-5173(00)00660-8. [DOI] [PubMed] [Google Scholar]
- 20.Roscetti G, Franzese O, Comandini A, Bonmassar E. Cytotoxic activity of Hypericum perforatum L. on K562 erythroleukemic cells: Differential effects between methanolic extract and hypericin. Phytother Res. 2004;18(1):66–72. doi: 10.1002/ptr.1369. [DOI] [PubMed] [Google Scholar]
- 21.Wilhelm KP, Biel S, Siegers CP. Role of flavonoids in controlling the phototoxicity of Hypericum perforatum extracts. Phytomedicine. 2001;8(4):306–309. doi: 10.1078/0944-7113-00042. [DOI] [PubMed] [Google Scholar]
- 22.Varriale L, Coppola E, Quarto M, Veneziani BM, Palumbo G. Molecular aspects of photodynamic therapy: Low energy pre-sensitization of hypericin-loaded human endometrial carcinoma cells enhances phototolerance, alters gene expression and affects the cell cycle. FEBS Lett. 2002;512(1–3):287–290. doi: 10.1016/s0014-5793(02)02239-1. [DOI] [PubMed] [Google Scholar]
- 23.Vantieghem A, Assefa Z, Vandenabeele P, Declercq W, Courtois S, Vandenheede JR, Merlevede W, de WP, Agostinis P. Hypericin-induced photosensitization of HeLa cells leads to apoptosis or necrosis. Involvement of cytochrome c and procaspase-3 activation in the mechanism of apoptosis. FEBS Lett. 1998;440(1–2):19–24. doi: 10.1016/s0014-5793(98)01416-1. [DOI] [PubMed] [Google Scholar]
- 24.Kawaii S, Tomono Y, Katase E, Ogawa K, Yano M. Antiproliferative activity of flavonoids on several cancer cell lines. Biosci Biotechnol Biochem. 1999;63(5):896–899. doi: 10.1271/bbb.63.896. [DOI] [PubMed] [Google Scholar]
- 25.Wills NJ, Park J, Wen J, Kesavan S, Kraus GA, Petrich JW, Carpenter S. Tumor cell toxicity of hypericin and related analogues. Photochem Photobiol. 2001;74(2):216–220. doi: 10.1562/0031-8655(2001)074<0216:tctoha>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 26.Dougherty TJ. Photodynamic therapy. Adv Exp Med Biol. 1985;193:313–328. doi: 10.1007/978-1-4613-2165-1_30. [DOI] [PubMed] [Google Scholar]
- 27.Piette J, Volanti C, Vantieghem A, Matroule JY, Habraken Y, Agostinis P. Cell death and growth arrest in response to photodynamic therapy with membrane-bound photosensitizers. Biochem Pharmacol. 2003;66(8):1651–1659. doi: 10.1016/s0006-2952(03)00539-2. [DOI] [PubMed] [Google Scholar]
- 28.Park J, English DS, Wannemuehler Y, Carpenter S, Petrich JW. The role of oxygen in the antiviral activity of hypericin and hypocrellin 5. Photochem Photobiol. 1998;68(4):593–597. [PubMed] [Google Scholar]
- 29.Kubin A, Wierrani F, Burner U, Alth G, Grunberger W. Hypericin–The facts about a controversial agent 1. Curr Pharm Des. 2005;11(2):233–253. doi: 10.2174/1381612053382287. [DOI] [PubMed] [Google Scholar]
- 30.Schempp CM, Simon-Haarhaus B, Heine A, Schopf E, Simon JC. In vitro and in vivo activation of hypericin with the incoherent light source PDT 1200 SOA (520–750 nm) and with solar simulated radiation (290–2500 nm) 21. Photodermatol Photoimmunol Photomed. 1999;15(1):13–17. doi: 10.1111/j.1600-0781.1999.tb00046.x. [DOI] [PubMed] [Google Scholar]
- 31.Leccia MT, Richard MJ, Joanny-Crisci F, Beani JC. UV-A1 cytotoxicity and antioxidant defence in keratinocytes and fibroblasts. Eur J Dermatol. 1998;8(7):478–482. [PubMed] [Google Scholar]
- 32.Wang W, VanAlstyne PC, Irons KA, Chen S, Stewart JW, Birt DF. Individual and interactive effects of apigenin analogues on G2/M cell-cycle arrest in human colon carcinoma cell lines 1. Nutr Cancer. 2004;48(1):106–114. doi: 10.1207/s15327914nc4801_14. [DOI] [PubMed] [Google Scholar]