

Abstract
Leopard, a well-known zebrafish mutant that has a spotted skin pattern instead of stripes, is a model for the study of pigment patterning. To understand the mechanisms underlying stripe formation, as well as the spot variation observed in leopard, we sought to identify the gene responsible for this phenotype. Using positional cloning, we identified the leopard gene as an orthologue of the mammalian connexin 40 gene. A variety of different leopard alleles, such as leot1, leotq270 and leotw28, show different skin-pattern phenotypes. In this manuscript we show that the mutation in allele leot1 is a nonsense mutation, whereas alleles leotq270 and leotw28 contain the missense mutations I202F and I31F, respectively. Patch-clamp experiments of connexin hemichannels demonstrated that the I202F substitution in allele leotq270 disrupted the channel function of connexin41.8. These results demonstrate that mutations in this gene lead to a variety of leopard spot patterns.
Keywords: connexin, leopard, pattern formation, positional cloning, zebrafish
Introduction
Colour patterns such as eye spots, dots, mottle and cross stripes are visually prominent features of many organisms and can be involved in camouflage, threatening behaviour and mate choice. Among a large number of pattern types, stripes and dots are the most well known and attractive. Zebrafish (Danio rerio)—a model organism for genetic and developmental studies—is known, as its name indicates, for its stripes, comprising black (melanophore), orange (xanthophore) and silver (iridophore) pigment cells. Consequently the zebrafish is also a model organism for colour pattern studies (Johnson et al, 1995a; Hirata et al, 2003, 2005; Kelsh, 2004). Several zebrafish genes involved in the development of pigments and/or pigment cells have been identified. For example, c-kit (sparse), ednrb1 (rose), c-fms (panther), mitf (nacre) and slc24a5 (golden) have been isolated from mutant fish and investigated for their roles in pigment cell development (Lister et al, 1999; Parichy et al, 1999, 2000a, 2000b; Rawls & Johnson, 2000; Lamason et al, 2005). Although these studies have contributed to our understanding of the development and migration of individual pigment cells, how these cells form a pattern in the skin remains largely unclear. Although the reaction–diffusion model, originally proposed by Turing (Turing, 1952; Kondo & Asai, 1995; Asai et al, 1999; Kondo, 2002), provides an explanation for how pigment cells make striped patterns, little is known about the molecular basis of this theoretical model. Thus, analysis of zebrafish mutants that do not show any defect in pigment cell development but have altered pigment cell localization would contribute to our understanding of pattern formation.
Leopard is one of the most well-known zebrafish mutants; it was introduced in the well-known textbook Developmental Biology as a mutant awaiting identification of its corresponding gene (Gilbert, 2003). In this study, we show that the zebrafish connexin41.8 gene (Cx41.8) is responsible for the leopard phenotypes. Connexin is a component of gap junctions (Kumar & Gilula, 1996). In most eukaryotic tissues and organs, gap junction channels allow direct intercellular transfer of ions, secondary messengers (such as cyclic AMP, ATP, inositol 1,4,5-trisphosphate) and small metabolites of up to ∼1,500 Da (Saez et al, 2005). Gap junctions are an aggregate of intercellular channels, each of which is composed of two connexons. Connexons are hexamers of connexin subunits, which are encoded by a multigene family, and more than 20 connexin genes have been identified in humans (Gerido & White, 2004). In zebrafish, almost 40 connexin genes were found through database searches (Eastman et al, 2006).
Here, we report that a variety of leopard allele phenotypes are generated with a change in connexin function, reflecting the effects of different mutations in Cx41.8.
Results And Discussion
Zebrafish leopard phenotype
We examined three leopard alleles: leot1, leotw28 and leotq270. Fig 1 illustrates the zebrafish phenotypes used in this study. Wild-type (WT) zebrafish (strain Tu) have several well-organized stripes consisting of melanophores and xanthophores (Fig 1A). The leot1 heterozygote has a similar phenotype to the WT (Fig 1B), whereas the leot1 homozygote shows a spot pattern consisting of well-organized melanophores (Fig 1C). The leotw28 heterozygote shows a pattern of undulating stripes (Fig 1D), and the leotw28 homozygote shows an interrupted stripe pattern (Fig 1E). In addition, there are fewer melanophores in the leotw28 homozygote than in the leotw28 heterozygote. The leotq270 heterozygote shows a spot pattern similar to the leot1 homozygote, whereas the leotq270 homozygote has smaller spots (Fig 1F,G). These observations demonstrate that the leotq270 mutant has a more pronounced phenotype than does the leotw28 mutant.
Figure 1.

Stripe patterns of zebrafish. (A) Wild-type (WT) zebrafish with normal horizontal stripes. (B,D,F) Zebrafish heterozygous for leot1, leotw28 or leotq270. (C,E,G) Zebrafish homozygous for leot1, leotw28 or leotq270. The inset boxes in each image show magnified images of stripes or spots on the skin of the fish.
Cx41.8 is the leopard gene
To understand the role of the leopard gene in stripe pattern development in zebrafish skin, we used positional cloning to identify the corresponding gene (supplementary information online). Meiotic mapping with chromosomal walking identified the critical region for leopard (Fig 2A). This region yielded two candidate genes, bcl9, the B-cell chronic lymphocytic leukaemia/lymphoma 9 gene, and Cx41.8, the connexin41.8 gene. We sequenced the coding regions of both genes from two WT strains, AB and Tu, and the three leopard alleles. No mutation was detected in the bcl9 gene when the deduced amino-acid sequences of the WT and leopard alleles were compared. However, mutations were identified in the Cx41.8 sequence of each of the leopard mutant alleles leot1, leotw28 and leotq270, as shown in Fig 2B and supplementary Fig S1 online. leot1 carries a nonsense mutation in the first extracellular loop domain, which reduces the predicted molecular size of Cx41.8 encoded by this allele from 42 to 8 kDa. Because the leot1 heterozygote had the same phenotype as the WT fish, leot1 represents a recessive allele caused by a null mutation of Cx41.8. A missense mutation, I31F, was identified in the allele leotw28, and another missense mutation, I202F, was identified in the allele leotq270 (supplementary Fig S1 online). Because the I31F mutation lies in the first transmembrane domain of Cx41.8, and I202F lies in the fourth transmembrane domain, these substitutions were presumed to be critical mutations for connexin function. leotw28 and leotq270 heterozygotes have phenotypes different from that of WT; thus, they represent dominant-negative alleles.
Figure 2.
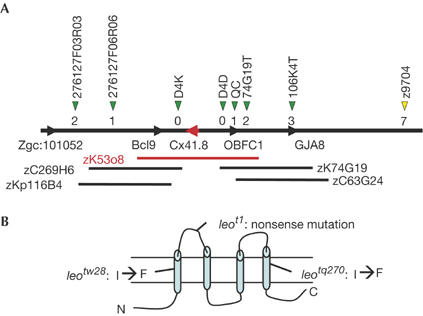
Zebrafish connexin41.8 as a leopard gene. (A) Map of the leopard region. The yellow arrowhead indicates the microsatellite marker z9704. The red bar indicates the bacterial artificial chromosome (BAC) clone zK53o8, which includes the critical region for leopard. The black bars show the BAC clones located around this region. Green arrowheads indicate single nucleotide polymorphism marker sites. Each site was named as shown above the arrowhead. The numbers below the green arrowheads refer to the number of recombination events (supplementary information online). This region includes two genes, bcl9 and Cx41.8. Cx41.8 is indicated by a red arrowhead. (B) Schematic of zebrafish connexin41.8 showing predicted structural motifs and mutations detected in leopard alleles.
To confirm that this critical region includes the leopard gene, we performed transgenic experiments. We injected the bacterial artificial chromosome (BAC) clone, zK53o8, into 500 fertilized eggs of homozygous leot1 mutants, and observed that some transgenic mosaic fish showed melanophore spots that are larger than those of leot1 homozygote fish (supplementary Fig S2 online). This observation is consistent with a transplantation experiment in which transplanted mosaic fish show a larger or irregular form of melanophore spots (Maderspacher & Nusslein-Volhard, 2003). Accordingly, this result indicates that this BAC clone actually contains a factor for melanophore development and/or stripe pattern formation (supplementary Fig S2 online).
Functional analysis of Cx41.8
To determine whether the mutations identified in the Cx41.8 gene from the leopard mutants affect the ability of connexin to form functional channels, we performed patch-clamp analysis to record hemichannel currents. We first prepared HeLa cells transfected with either WT or mutant Cx41.8 complementary DNA. After establishing the conditions for whole-cell recording, the membrane potential was clamped to +20 mV. To measure the current flowing through the connexin hemichannel, bipolar voltage pulses of 100 ms were delivered to alter the Vm in 20 mV increments from −80 to +40 mV (Fig 3A). Hemichannels composed of connexins are known to open on removal of extracellular Ca2+. Concordantly, the presence of 2 mM extracellular Ca2+ elicited a very small current, whereas depolarization produced activating currents in the absence of Ca2+. The hemichannel blocker carbenoxolone also blocked the hemichannel ability (Fig 3A). Fig 3B represents a superimposed Im trace of HeLa cells transfected with the WT, leotw28 or leotq270 connexin41.8 gene, or of untransfected HeLa cells (negative control). After establishing conditions for whole-cell recording, Vm was clamped to −40 mV. Voltage pulses of 100 ms were then delivered to depolarize Vm up to +15 mV. Currents were recorded in the absence of Ca2+. Fig 3C summarizes the patch-clamp data for the WT, leotw28 and leotq270 connexin hemichannels compared with untransfected HeLa cells. Interestingly, connexin41.8 from leotq270 had the same low-level basal current as untransfected HeLa cells, indicating that connexin hemichannel function was disrupted by the leotq270 mutation. This result led us to conclude that the I202F substitution has a crucial effect on the ability of connexin to conduct ions and that the phenotypes of zebrafish skin pattern might depend on connexin channel function. Conversely, the trace from zebrafish allele leotw28 could not be distinguished from that of WT, indicating that the I31F substitution does not have a substantial effect on ion channel function. Small differences in current might not be detected using this assay and/or this mutation might only manifest its effect after the formation of intercellular channels (Castro et al, 1999; Bao et al, 2004). Further analysis might demonstrate whether the mutation I31F actually affects the function of connexin gene. Therefore, we concluded that the leopard allele with the most severe spot phenotype, leotq270, showed the most pronounced disruption in channel function.
Figure 3.
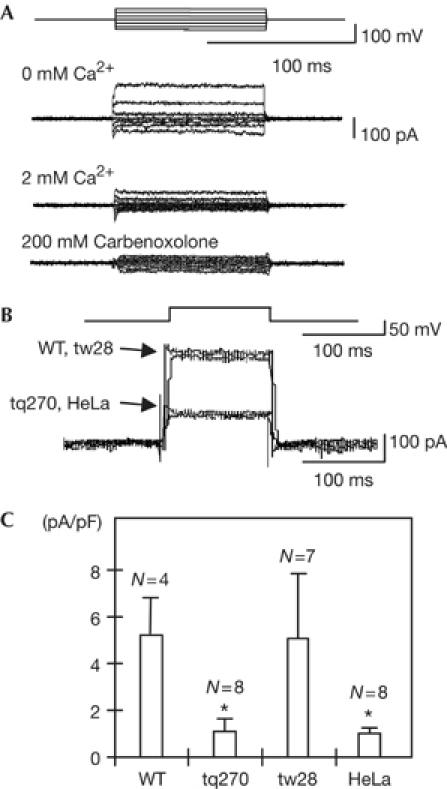
Patch-clamp analysis of Cx41.8 hemichannels expressed in HeLa cells. (A) Establishment of conditions for patch-clamp analysis with HeLa cells transfected with the wild-type (WT) Cx41.8 cDNA. Im responses recorded in solution with no added Ca2+ (upper), with 2 mM Ca2+ (middle) and with 200 μM carbenoxolone (lower) in the bath solution. (B) Superimposed Im traces from HeLa cells expressing WT, leotw28 or leotq270 connexin41.8, and untransfected HeLa cells (negative control). (C) Bar graph of WT (N=4), mutant leotw28 (N=7) and mutant leotq270 (N=8), compared with untransfected HeLa cells (N=8, negative control). Data represent the means±s.e.m. Each value was compared with that of WT. *P<0.005.
The dominant-negative phenotypes of leotw28 and leotq270 could be explained by the generation of heterohexamers of connexin to form the gap junctions, although we did not obtain functional data about leotw28. Homotypic heterohexamers can be generated from mutated and normal Cx41.8, and heterotypic heterohexamers can be formed from mutated Cx41.8 and another zebrafish connexin, such as Cx43 or Cx45.6 (Christie et al, 2004). It is likely that the phenotype of the heterozygous leotw28 mutant is due to formation of homotypic heterohexamers that consequently cause a wavy striped pattern in zebrafish. The leotq270-encoded protein showed disrupted channel function, predicting that the skin pattern phenotype of the leotq270 heterozygote would be the same as that of the null mutant leot1 homozygote. In fact, the dominant-negative leotq270 homozygote has a more severe phenotype than the null mutant leot1 homozygote. This more severe phenotype of leotq270 homozygotes might be explained by the formation of heterotypic heterohexamers composed of leotq270 connexin41.8 and other connexins, such as Cx43 or Cx45.6, that form irregular connexons in the homozygous leotq270 mutant. Another report supports this hypothesis: connexin 26 and connexin 32 can form a heterohexamer connexon that has different signalling molecule selectivity compared with connexin 26 or connexin 32 homohexamer connexons (Bevans et al, 1998). In addition, a mouse gap junction consisting of WT connexin 43 and a dominant-negative mutant connexin 43 has a smaller channel pore size than normal gap junctions, thereby allowing the transfer of relatively smaller molecules (Upham et al, 2003). This hypothesis of heterohexamer formation further suggests that other connexin genes contribute to stripe pattern formation.
Expression of the Cx41.8 gene
Because gene expression levels of connexins are generally too small to detect using in situ hybridization, we performed reverse transcription–PCR (RT–PCR) analysis to determine the tissue specificity of Cx41.8 expression (supplementary information online). RT–PCR was carried out using cDNA generated from adult eye, brain, heart, liver, skin, fin, ovary, testis, intestine and gill. The fin was divided into the dark portion (melanophore) and light portion (light portion) of each stripe, and each fraction was subjected to RT–PCR. Cx41.8 expression was strongly detected in the eye, brain, heart, skin and fin. The gene was also expressed in melanophore and xanthophore portions of the fin (Fig 4). To confirm these results further, and to exclude the possibility of cross contamination of the pigment cells in each fraction, an alternative approach was undertaken as described in the supplementary text and supplementary Fig S3 online. These data indicate that the gene is expressed in melanophores and that expression in xantophores is also likely.
Figure 4.
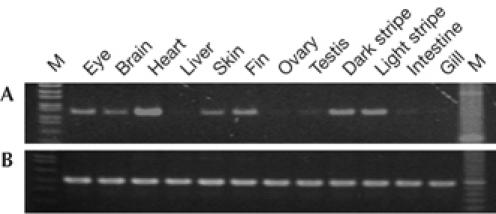
Cx41.8 expression in adult tissues. Reverse transcription–PCR was performed with RNA samples isolated from the indicated adult fish tissues. M indicates molecular size marker. Total RNAs were reverse-transcribed with oligo-d(T)18 primer, and the resultant complementary DNAs were amplified by PCR using specific primers for (A) Cx41.8 and (B) β-actin.
Speculation on connexin functions in pattern formation
It is possible that the change in skin pattern can be explained by the loss of cell–cell interactions owing to the gap-junction defect of connexin41.8. Maderspacher & Nusslein-Volhard (2003) provided an important observation by transplantation experiments that showed that the spots became larger in transplanted fish, suggesting that the leopard gene might have a crucial role in cell–cell interactions of melanophore–melanophore, xanthophore–xanthophore and melanophore–xanthophore during the process of stripe pattern formation. This observation is consistent with our results, because gap junction proteins are responsible for the formation of cell–cell interactions. Conversely, Hirata et al (2003, 2005) showed that there are layer structures of pigment cells, such as xanthophores/iridophores/melanophores, on zebrafish skin and this structure might prevent direct interaction between melanophores and xanthophores. These observations indicate that the leopard gene might also have an indirect effect on cell-to-cell interactions between melanophores and xanthophores. Indirect effects might be explained by the function of the connexin hemichannel. Pearson et al (2005) proposed a model in which ATP release and its diffusion from chick embryonic retinal pigment epithelium through connexin 43 hemichannels activates mitosis of the progenitor cells in the adjacent neural retina. In this model, connexin hemichannels are important for differentiation of neighbouring cells. Although there is no evidence that a similar ATP signalling pathway through connexin hemichannels contributes to melanophore differentiation in zebrafish, defects in the function of mutant Cx41.8 hemichannels could explain the observation that zebrafish leopard mutants have a reduced number of melanophores (Fig 1; Johnson et al, 1995a).
Recently, we identified a jaguar/obelix gene as a potassium channel (Iwashita M, Watanabe M, Ishii M, Chen T, Johnson SL, Kurachi Y, Okada N & Kondo S, unpublished data). Mutations in this gene alter the localization of pigment cells, causing broadened stripe patterns. Although the functions of the leopard and jaguar/obelix genes in pigment cell localization in zebrafish skin remain unknown, it is a novel and fascinating finding that channels that form gap junctions and ion channels contribute to pigment cell localization. This leads to further questions on how small molecules such as ATP and ions contribute to the positioning of pigment cells on the skin to yield a specific pattern.
Methods
Positional cloning. leot1 homozygous fish were crossed with WT strains Tu, AB, and India to produce a reference mapping cross. Heterozygous F1 were intercrossed, and separate pools of F2 homozygous leot1 fish (1,738 individuals) were used for sequence length polymorphism and single nucleotide polymorphism analyses. Because the linkage number in which the leopard gene was mapped was previously predicted to be linkage group 1 (LG1; Johnson et al, 1995b), we started screening using LG1 polymorphism markers that were obtained from the Z-FIN database. (See the supplementary information online for the positional cloning strategy for the leopard gene.)
HeLa cell transfection. Patch-clamp experiments were performed with HeLa cell clones that were transfected with cDNA constructs containing the coding sequence of WT or mutant Cx41.8. HeLa cells were transfected with 5 μg of recombinant Cx41.8-pcDNA plasmid using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA). The cells were seeded onto sterile glass coverslips placed in 12-well dishes and used 24–48 h after plating (see the supplementary information online for further details).
Hemichannel current measurement. Experiments were carried out on single cells attached to a pipette. The channel current was recorded using a patch-clamp amplifier, Axon200B (Axon Instrument, CA, USA), and analysed with a commercially available software, pCLAMP9 (Axon Instrument). Results are shown as mean values, and error bars represent ±s.e.m. (see the supplementary information online for further details).
Supplementary information is available at EMBO reports online (http://www.emboreports.org).
Supplementary Material
Supplementary Information
Acknowledgments
This work was supported by a Grant-in-Aid for Scientific Research on Priority Areas ‘Molecular Mechanism of Speciation' and ‘Comparative Genomics' from the Ministry of Education, Culture, Sports, Science and Technology of Japan to N.O.
References
- Asai R, Taguchi E, Kume Y, Saito M, Kondo S (1999) Zebrafish leopard gene as a component of the putative reaction–diffusion system. Mech Dev 89: 87–92 [DOI] [PubMed] [Google Scholar]
- Bao X, Reuss L, Altenberg GA (2004) Regulation of purified and reconstituted connexin 43 hemichannels by protein kinase C-mediated phosphorylation of Serine 368. J Biol Chem 279: 20058–20066 [DOI] [PubMed] [Google Scholar]
- Bevans CG, Kordel M, Rhee SK, Harris AL (1998) Isoform composition of connexin channels determines selectivity among second messengers and uncharged molecules. J Biol Chem 273: 2808–2816 [DOI] [PubMed] [Google Scholar]
- Castro C, Gomez-Hernandez JM, Silander K, Barrio LC (1999) Altered formation of hemichannels and gap junction channels caused by C-terminal connexin-32 mutations. J Neurosci 19: 3752–3760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christie TL, Mui R, White TW, Valdimarsson G (2004) Molecular cloning, functional analysis, and RNA expression analysis of connexin45.6: a zebrafish cardiovascular connexin. Am J Physiol Heart Circ Physiol 286: H1623–H1632 [DOI] [PubMed] [Google Scholar]
- Eastman SD, Chen TH, Falk MM, Mendelson TC, Iovine MK (2006) Phylogenetic analysis of three complete gap junction gene families reveals lineage-specific duplications and highly supported gene classes. Genomics 87: 265–274 [DOI] [PubMed] [Google Scholar]
- Gerido DA, White TW (2004) Connexin disorders of the ear, skin, and lens. Biochim Biophys Acta 1662: 159–170 [DOI] [PubMed] [Google Scholar]
- Gilbert SF (2003) Developmental Biology 7th edn. Sunderland, MA, USA: Sinauer Associates Inc. [Google Scholar]
- Hirata M, Nakamura K, Kanemaru T, Shibata Y, Kondo S (2003) Pigment cell organization in the hypodermis of zebrafish. Dev Dyn 227: 497–503 [DOI] [PubMed] [Google Scholar]
- Hirata M, Nakamura K, Kondo S (2005) Pigment cell distributions in different tissues of the zebrafish, with special reference to the striped pigment pattern. Dev Dyn 234: 293–300 [DOI] [PubMed] [Google Scholar]
- Johnson SL, Africa D, Walker C, Weston JA (1995a) Genetic control of adult pigment stripe development in zebrafish. Dev Biol 167: 27–33 [DOI] [PubMed] [Google Scholar]
- Johnson SL, Africa D, Horne S, Postlethwait JH (1995b) Half-tetrad analysis in zebrafish: mapping the ros mutation and the centromere of linkage group I. Genetics 139: 1727–1735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelsh RN (2004) Genetics and evolution of pigment patterns in fish. Pigment Cell Res 17: 326–336 [DOI] [PubMed] [Google Scholar]
- Kondo S (2002) The reaction–diffusion system: a mechanism for autonomous pattern formation in the animal skin. Genes Cells 7: 535–541 [DOI] [PubMed] [Google Scholar]
- Kondo S, Asai R (1995) A reaction–diffusion wave on the skin of the marine angelfish Pomacanthus. Nature 376: 765–768 [DOI] [PubMed] [Google Scholar]
- Kumar NM, Gilula NB (1996) The gap junction communication channel. Cell 84: 381–388 [DOI] [PubMed] [Google Scholar]
- Lamason RL et al. (2005) SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 310: 1782–1786 [DOI] [PubMed] [Google Scholar]
- Lister JA, Robertson CP, Lepage T, Johnson SL, Raible DW (1999) nacre encodes a zebrafish microphthalmia-related protein that regulates neural-crest-derived pigment cell fate. Development 126: 3757–3767 [DOI] [PubMed] [Google Scholar]
- Maderspacher F, Nusslein-Volhard C (2003) Formation of the adult pigment pattern in zebrafish requires leopard and obelix dependent cell interactions. Development 130: 3447–3457 [DOI] [PubMed] [Google Scholar]
- Parichy DM, Rawls JF, Pratt SJ, Whitfield TT, Johnson SL (1999) Zebrafish sparse corresponds to an orthologue of c-kit and is required for the morphogenesis of a subpopulation of melanocytes, but is not essential for hematopoiesis or primordial germ cell development. Development 126: 3425–3436 [DOI] [PubMed] [Google Scholar]
- Parichy DM, Mellgren EM, Rawls JF, Lopes SS, Kelsh RN, Johnson SL (2000a) Mutational analysis of endothelin receptor b1 (rose) during neural crest and pigment pattern development in the zebrafish Danio rerio. Dev Biol 227: 294–306 [DOI] [PubMed] [Google Scholar]
- Parichy DM, Ransom DG, Paw B, Zon LI, Johnson SL (2000b) An orthologue of the kit-related gene fms is required for development of neural crest-derived xanthophores and a subpopulation of adult melanocytes in the zebrafish, Danio rerio. Development 127: 3031–3044 [DOI] [PubMed] [Google Scholar]
- Pearson RA, Dale N, Llaudet E, Mobbs P (2005) ATP released via gap junction hemichannels from the pigment epithelium regulates neural retinal progenitor proliferation. Neuron 46: 731–744 [DOI] [PubMed] [Google Scholar]
- Rawls JF, Johnson SL (2000) Zebrafish kit mutation reveals primary and secondary regulation of melanocyte development during fin stripe regeneration. Development 127: 3715–3724 [DOI] [PubMed] [Google Scholar]
- Saez JC, Retamal MA, Basilio D, Bukauskas FF, Bennett MV (2005) Connexin-based gap junction hemichannels: gating mechanisms. Biochim Biophys Acta 1711: 215–224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turing AM (1952) The chemical basis of morphogenesis. Philos Trans R Soc Lond B Biol Sci 327: 37–72 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Upham BL, Suzuki J, Chen G, Wang Y, McCabe LR, Chang CC, Krutovskikh VA, Yamasaki H, Trosko JE (2003) Reduced gap junctional intercellular communication and altered biological effects in mouse osteoblast and rat liver oval cell lines transfected with dominant-negative connexin 43. Mol Carcinog 37: 192–201 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Information