

Abstract
In songbirds, nocturnal activity is believed to be a characteristic feature of migration. However, unlike experimental conditions where the onset of nocturnal restlessness is defined as a shift of activity leading up to the dark period, this behaviour has, until now, not been observed in natural conditions. Here we studied the nocturnal behaviour of radio-tagged juvenile Eurasian reed warblers (Acrocephalus scirpaceus) during the pre-migratory period. The birds started nocturnal flights at the age of 38 days, whereas migration did not commence until they were at least 50 days old. The birds left their natal site by nocturnal flights and repeatedly returned to it. Such shuttle movements suggest the existence of a previously unknown period of nocturnal activity. Motivation to perform such night flights gradually increases with age. We relate the function of these nocturnal pre-migratory flights to the development of a stellar compass, necessary for detecting the compass direction towards winter quarters and for the formation of a navigational target, which will be used during return (spring) migration.
Keywords: bird migration, pre-migratory behaviour, nocturnal flights, navigation, telemetry
1. Introduction
A large proportion of birds migrating in the Old and New World do so by night. It is currently believed that in normally diurnal songbirds, nocturnal activity is inseparably linked with migration. Time and age of the onset of nocturnal activity in caged migrants is often used as an identifier of the commencement of autumn migration (Palmgren 1944; Mewaldt et al. 1964; Gwinner et al. 1971). However, back in the late 1960s impressive work done by Emlen (1972) suggested that juvenile nocturnal migrants had to be nocturnal in order to develop their stellar orientation system, as this could only be done at night. Apart from this, studies of captured young Eurasian reed warblers (Acrocephalus scirpaceus), of known age and hatching location, have suggested that they perform nocturnal flights before the onset of autumn migration (Bulyuk et al. 2000; Mukhin 2004). The function of these flights, however, remained obscure.
We assume that in order to successfully complete their unpredictable migration, first-autumn birds have to know two crucial things: where to fly and where to return. Therefore, we suggest that these early nocturnal flights, which occur during a short time window, may be involved in (i) developing their orientation abilities and (ii) imprinting their future breeding area. In order to imprint the locality of future breeding, juveniles have to familiarize themselves with local surroundings before migratory departure (Sokolov et al. 1984; Morton et al.1991). It is not impossible that the best time for nocturnal migrants (which most warblers are) to do this is at night. Therefore, a role of this early nocturnal activity in both these processes is suggested.
Nocturnal pre-migratory activity raises another important question: how do diurnal birds shift their activity to a nocturnal rhythm? Numerous studies on caged birds (Gwinner et al. 1971; Berthold 1996) cannot replace tracking the changes that occur during this crucial period in the behaviour of free-living individuals.
All this suggests that the pre-migratory period in long-distance nocturnal migrants is much more complicated than currently assumed. In this study we tracked locally hatched radio-tagged juvenile Eurasian reed warblers during the post-fledging period to shed light on their pre-migratory behaviour.
2. Material and methods
The study was done near Rybachy village on the south-eastern Baltic coast (55°09′ N, 20°51′ E). The study area is a 6.5 ha breeding habitat of Eurasian reed warblers. Nest searches were done there during the breeding period and nestlings were banded. A small lake supporting some reed-beds (0.4 ha) is situated 1.5 km away. The next closest reed-beds are located 17 km to the south-west.
We fitted locally hatched Eurasian reed warblers that had been banded as nestlings with radio transmitters. The birds were 25–50 days old when tagged. Transmitters (LB-2, 0.5 g, life span three weeks, Holohil Systems Inc., Ont., Canada) were fitted to the birds with Rappole harnesses (Rappole & Tipton 1991). The transmitter mass and harness did not exceed 5–7% of the birds' lean body mass, which is believed to be the upper acceptable limit (Cochran 1980). The detection range of the signal from ground to ground was approximately 0.5 km and the detection range from air to ground was approximately 1.5 km, but both ranges varied considerably with landscape, weather and vegetation. The birds were followed for 24 h. We used a programmable R2000 receiver (Advanced Telemetry Systems, Isanti, MN, USA), which allowed us to track up to 10 radio-tagged birds simultaneously. Nocturnal tracking was done from either of the two towers (12 and 6 m high) in the study area. If the signal intensity changed, the current location of the birds was compared with the one taken 40 min after sunset, after the end of daytime activity. Also, flights were assumed to occur when a signal was located from the sky for a long period or if a signal faded into the distance, suggesting directed flight away from the study site. When a bird left the immediate study site, we attempted to locate it in the vicinity of Rybachy. If the bird was not located during the night, a new search was done in the morning. The transmitter frequency of a disappeared bird was not deleted from the receiver's memory, so that a possible return to the reception area could be detected during the same or a successive night.
Since the lifespan of the transmitters used did not exceed 21 days, we could not cover the whole post-fledging period until the beginning of migration. Therefore, to analyse the beginning of nocturnal activity and the first flights, we used birds tagged at the age of 25–40 days, and to characterize the age of leaving the natal site, the data on birds tagged at the age of 41–50 days were used.
We radio-tagged 22 juvenile Eurasian reed warblers in 2002 and nine individuals in 2003. Of the tagged birds, two died soon after the onset of tracking. Two transmitters were lost by birds and subsequently found by us. If certain parameters were identified in any of these four birds (age of beginning nocturnal activity, age of the first nocturnal flight), they were included in the analysis.
3. Results
(a) First change of signal
Before the first reliable registration of a night flight, we repeatedly recorded changing signals in tagged birds during the night-time. These signal changes could not be identified as flights with certainty but strongly suggested that the birds were awake. In birds radio-tagged before 30 days old, the mean age of the first signal change was 31.7 (s.d.=1.5, n=7). It should be noted that similar signal changes were also recorded during the period of active nocturnal flights.
(b) Nocturnal flights
With increasing age of the birds, we started to record short nocturnal movements which, however, did not result in leaving the reception range (8 ha). Of 21 individuals radio-tagged before 40 days old and surviving until the onset of nocturnal movements, 16 made short night flights over the natal area before the night of departure. These nocturnal flights involved flying in different directions over the study area and also disappearing from the reception range, with subsequent reappearance during the same night. The onset of these flights is shown in figure 1. Nine individuals (56%) took off during the deep night, five (31%) during the astronomical and two (13%) during the nautical twilight. No birds made nocturnal flights during the morning civil twilight. These flights were evenly distributed across the night.
Figure 1.
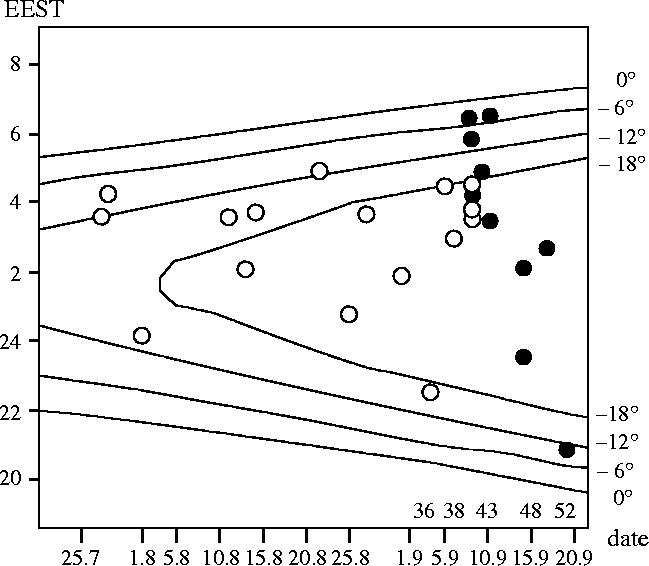
Time of first flights over the natal site by young Eurasian reed warblers radio-tagged before 40 days old (open circles); time shift for beginning of flight activity in one young Eurasian reed warbler (closed circles). Near the date scale is the age of the bird in days. Lines show sun elevation under the horizon: 0°, sunset and sunrise; 6°, limit of civil twilight; 12°, limit of nautical twilight; 18°, limit of astronomical twilight. EEST, Eastern European summer time.
Figure 2 (closed circles) shows first departure times of Eurasian reed warblers marked before 40 days old. Birds that did not return to the study site until sunrise were treated as departed. Birds that took off but returned to the hatching site within several hours (during the same night) were not treated as departed.
Figure 2.
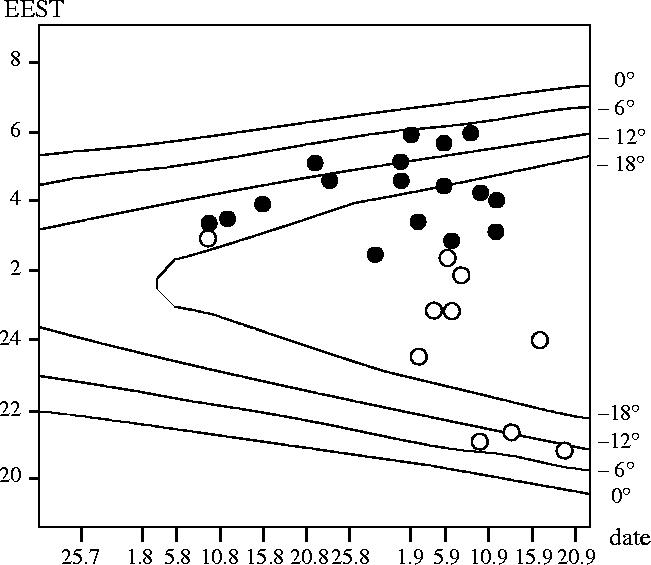
Departure time of young Eurasian reed warblers from the natal site. Closed circles: first departures of birds radio-tagged before 40 days old; open circles: final departure of birds aged over 50 days old. Symbols as in figure 1.
All departures by juveniles occurred in the latter half of the night. Take-off time varied between 351 and 569 min after sunset, on average occurring 460.7 min (s.d.=66.7, n=18), 10th hour, into the night. Seven birds (39%) took off during the deep night, six birds (33%) during the morning astronomical and five (28%) during nautical twilight. No bird took off during the morning civil twilight or after the sunrise. The average age at departure was 41.7 days (s.d.=3.8).
Of 27 radio-tagged birds, 17 (63%) were recorded making return visits to the natal sites after departure. Seven of them (26%) returned more than once. When returns within the same night were excluded, off-site periods varied between 1 and 5 days. Both returns with landing at the natal site and flights over it without stopping were included.
Departure direction was highly variable. As the detection range of a flying bird does not exceed approximately 1.5 km, we could only estimate the direction of pre-migratory flights from Eurasian reed warblers relocated after departure. Our searching efforts were successful in 17 cases (33% of all departures, including the final ones). The birds were relocated up to 2.5 km away from the study area. Our attempts to relocate radio-tagged individuals in remote reed-beds (17–20 km from the hatching area) proved unsuccessful, even though such distant movements do occur in juvenile Eurasian reed warblers in the pre-migratory period (Mukhin 2004).
(c) The last registration
Figure 2 (open circles) shows the time of final departure time for birds whose age at tagging exceeded 40 days (n=7), and for the birds tagged before 40 days old that finally departed from the natal site at the age of approximately 50 days (n=3). Of the ten birds, six took off during the deep night, mainly in its first half, three at dusk and one during the morning astronomical twilight. Take-off time varied between 64 and 358 min after sunset with an average take-off time of 218.2 min (s.d.=123.9, n=9, 4th hour into the night). The exact take-off time of one bird was not known; it departed between 67 and 355 min after sunset. Age of the birds at last registration varied between 49 and 55 days with an average age of 51.5 days (s.d.=1.8; n=10).
The take-off time of birds tagged before 40 days old and departing for the first time differed significantly from the take-off time of birds tagged after 40 days old and finally departing to migrate from the natal site (t=6.65, p<0.001).
(d) Changes in flight activity rhythm
Due to the dispersal of birds during post-fledging movements, only three individuals radio-tagged at the age of 25–40 days were tracked until more than 50 days old; one of these was tracked starting at age 30 days and two were tracked from age 36 days. Figure 1 (closed circles) shows the flight activity pattern of one of them. The onset of flight activity with age gradually shifted towards the earlier part of the night. At the age of 52 days, when juvenile Eurasian reed warblers commence autumn migration, this bird took off during evening nautical twilight.
We analysed the relationship between take-off time and age for all radio-tagged birds tracked. Take-off time shifted significantly towards sunset with age (r=−0.24, p=0.028). All five take-offs of birds older than 51 days occurred before 192 min after sunset (on average 116.8 min).
4. Discussion
In early departing birds, for example, Eurasian reed warblers, the period for preparing for the first migratory departure is rather limited. In this species, fledglings gain independence from their parents when 25–29 days old. Long distance recoveries suggest that juveniles older than 54 days can already be captured on their migratory route, more than 450 km from the natal area (Mukhin 2004). Thus, they have some three weeks to develop the complex behavioural and physiological conditions which characterize a migrant ready for departure (Berthold 2001). Shifting the activity rhythm from a diurnal non-migratory to a nocturnal migratory one is believed to be an important indicator of a developed migratory disposition (Berthold 1996; Fusani & Gwinner 2004). However, nocturnal flights recorded by us contradict this view and suggest a special period of nocturnal activity long before the onset of autumn migration.
Tracking of radio-tagged Eurasian reed warblers showed that the first signs of nocturnal activity (not yet including any flights) occurred at the age of approximately 30 days. The recorded signal changes are interpreted as evidence that in this age, the birds remain awake during a part of the night. At the age of approximately 38 days they already performed nocturnal flights but did not leave the natal site for more than a few hours. The first departures from the natal area occurred on average at the age of 42 days. The last registration of radio-tagged birds took place when they were approximately 52 days old. The bird tracked from 36 days old until 52 days old tended to commence its night flights earlier with increasing age. A similar trend of begining nocturnal activity earlier during the night was reported in caged migrants (Helms 1963). It seems that alterations of nocturnal behaviour are governed by increasing motivation for nocturnal flights.
We suggest that the movements of birds older than 50 days most probably relate to the onset of migration. This is supported by the recovery data (see above) and by the take-off time of birds last recorded older than 50 days. It was on average 218 min after sunset, which is similar to the departure times of Eurasian reed warblers on their migration in southern Sweden (mean=128 min after sunset with the range 32–634 min; Åkesson et al. 2001, 2002).
Preparation for the first migration may serve as an explanation of active pre-migratory night life in juvenile Eurasian reed warblers. Two main (but non-exclusive) reasons may explain the necessity of early nocturnal activity and movements. Juvenile dispersal is believed to be aimed at selecting the future breeding site (Howard 1960; Greenwood & Harvey 1982). The birds have to be able to recognize the selected location(s) upon arrival from their wintering grounds (Ketterson & Nolan 1990). One avian navigation concept, the ‘mosaic map’ hypothesis (Wallraff 1974; Wiltschko & Wiltschko 1982), suggests that birds have to study the surroundings of their natal area during exploratory movements. The navigation target is the familiar area around an individual bird's future breeding site (Baker 1993). The Eurasian reed warbler is a nocturnal migrant (Cramp 1992) that returns to the breeding area by a nocturnal flight that is completed by dawn (Bolshakov et al. 2003). It may be that juveniles familiarize themselves with their future breeding area at night. Evaluation of habitat quality and its suitability for breeding might be done during daytime foraging movements, which normally do not exceed several hundred meters in range.
Shuttle movements of young Eurasian reed warblers agree with this suggested pattern. The natal site may be the focal point in the exploratory movements of a juvenile, around which a mental map of the area is built.
The second reason for nocturnal movements may be the development of orientation abilities. Inherited directional information about the species-specific wintering grounds has to be translated into a certain flight direction. One of the two main compass systems known to be used by nocturnal migrants is based on celestial cues (Wiltschko et al. 1987). At the first stages of migration, the celestial compass has a priority over the magnetic one (Wiltschko & Wiltschko 1988, 1998). To be able to use celestial cues, birds have to develop certain recognition skills during a sensitive period. It has been shown (Emlen 1967; 1970) that migrants have to observe sky rotation prior to their first migration in order to establish a star compass. It is suggested that a sensitive period for this learning should exist. Questions such as how this learning happens in nature and whether the birds have to fly or merely sit and observe the rotation of the celestial sphere remain. Either way, the birds obviously have to remain awake at night, as they can only develop their stellar compass during night-time.
The adaptive significance of pre-migratory flights at night-time might be the same as that suggested to explain nocturnal migration in passerines: temporal segregation of flights and foraging, predator avoidance and calmer atmospheric conditions at night as compared with the daytime (Berthold 2001). Lack of previous flight experience together with the active body moult they are undergoing during this period (Mukhin 2002) may enhance the importance of these factors. However, we believe that the main reasons for nocturnal flights are: (i) building a mental map of the area necessary for successful navigation during return migration next spring and (ii) developing orientation skills needed during the first autumn migration. These processes are crucial for successful migration to, and return from, their wintering grounds.
Pre-migratory night flights of juvenile Eurasian reed warblers are expressed in caged birds as weak nocturnal restlessness (Mukhin 1999). Similar early restlessness was reported for caged pied and collared flycatchers (Ficedula hypoleuca, F. albicollis) (Gwinner & Schwabl-Benzinger 1982) and garden warblers (Sylvia borin) (Widmer 1999). In the latter species and the blackcap (Sylvia atricapilla), nocturnal pre-migratory flights were found, using the telemetry method, to occur in nature (Mukhin unpublished). All this allows us to suggest that nocturnal flights long before migration are not unique for Eurasian reed warblers, but are a common phenomenon among nocturnal songbird migrants.
Acknowledgments
This work was supported by the Russian Foundation for Basic Research (grant no. 02-04-48608). It would not have been possible to carry out the fieldwork without help from a number of students to whom we are indebted. We thank Nikita Chernetsov for his help with improving earlier drafts. We appreciate the improvements in English usage made by Stacy Small, through the Association of Field Ornithologists' programme of editorial assistance, and John Walder.
References
- Åkesson S, Walinder G, Karlsson L, Ehnbom S. Reed warbler orientation: initiation of nocturnal migratory flight in relation to visibility of celestial cues at dusk. J. Anim. Behav. 2001;61:181–189. doi: 10.1006/anbe.2000.1562. [DOI] [PubMed] [Google Scholar]
- Åkesson S, Walinder G, Karlsson L, Ehnbom S. Nocturnal migratory flight initiation in reed warblers Acrocephalus scirpaceus: effect of wind on orientation and timing of migration. J. Avian Biol. 2002;33:349–357. [Google Scholar]
- Baker R.R. The function of post-fledging exploration: a pilot study of three species of passerines in Britain. Ornis Scand. 1993;24:71–79. [Google Scholar]
- Berthold P.Control of bird migration1996Chapman & Hall; London [Google Scholar]
- Berthold P.Bird migration: a general survey2001Oxford University Press [Google Scholar]
- Bolshakov C, Bulyuk V, Chernetsov N. Spring nocturnal migration of reed warblers Acrocephalus scirpaceus: departure, landing and body condition. Ibis. 2003;145:106–112. [Google Scholar]
- Bulyuk V.N, Mukhin A, Fedorov V.A, Tsvey A, Kishkinev D. Juvenile dispersal in reed warblers Acrocephalus scirpaceus at night. Avian Ecol. Behav. 2000;5:45–63. [Google Scholar]
- Cochran W.W. Wildlife telemetry. In: Schemnitz S, editor. Wildlife management techniques manual. 4th edn. The Wildlife Society; Washington: 1980. pp. 507–520. [Google Scholar]
- Cramp S.Handbook of the birds of Europe, the Middle East and North Africa: the birds of the Western Palaearctic1992Oxford University Press [Google Scholar]
- Emlen S.T. Migratory orientation in the indigo bunting, (Passerina cyanea). Evidence for use of celestial cues. Auk. 1967;84:309–342. [Google Scholar]
- Emlen S.T. Celestial rotation: its importance in the development of migratory orientation. Science. 1970;170:1198–1201. doi: 10.1126/science.170.3963.1198. [DOI] [PubMed] [Google Scholar]
- Emlen S.T. The ontogenetic development of orientation capabilities. In: Galler S.R, Schmidt-Koenig K, Jacobs G.J, Belleville R.E, editors. Animal orientation and navigation. US Government Printing Office; Washington: 1972. pp. 191–210. [Google Scholar]
- Fusani L, Gwinner E. Simulation of migratory flight and stopover affects night levels of melatonin in a nocturnal migrant. Proc. R. Soc. B. 2004;271:205–211. doi: 10.1098/rspb.2003.2561. 10.1098/rspb.2003.2561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenwood P.J, Harvey P.H. The natal and breeding dispersal of birds. Annu. Rev. Ecol. Syst. 1982;13:1–21. [Google Scholar]
- Gwinner E, Berthold P, Klein H. Investigation on annual rhythms in warblers II. The effect of photoperiod on the development of plumage, body weight and migratory restlessness in Phylloscopus trochilus and P. collybita. J. Ornithol. 1971;112:253–265. [Google Scholar]
- Gwinner E, Schwabl-Benzinger I. Adaptive temporal programming of molt and migratory disposition in two closely related long-distance migrants, the pied flycatcher (Ficedula hypoleuca) and the collared flycatcher (Ficedula albicollis) In: Papi F, Wallraff H.G, editors. Avian navigation. Springer; Berlin: 1982. pp. 75–89. [Google Scholar]
- Helms C.W. The annual cycle and Zugunruhe in birds. In: Sibleg C, editor. Proceedings of XIIIth International Ornithology Congress. American Ornithologists' Union; Baton Rouge, LA: 1963. pp. 925–939. [Google Scholar]
- Howard W.E. Innate and environmental dispersal of individual vertebrates. Am. Midl. Nat. 1960;63:152–161. [Google Scholar]
- Ketterson E.D, Nolan V. Site attachment and site fidelity in migratory birds: experimental evidence from the field and analogies from neurobiology. In: Gwinner E, editor. Bird migration: physiology and ecophysiology. Springer; Berlin: 1990. pp. 117–129. [Google Scholar]
- Mewaldt L.R, Morton M.L, Brown I.L. Orientation of migratory restlessness in Zonotrichia. Condor. 1964;66:377–417. [Google Scholar]
- Morton M.L, Wakamatsu M.W, Pereyra M.E, Morton G.A. Postfledging dispersal, habitat imprinting and philopatry in a montane, migratory sparrow. Ornis Scand. 1991;22:98–106. [Google Scholar]
- Mukhin A. Nocturnal restlessness in caged juvenile reed warblers Acrocephalus scirpaceus. Avian Ecol. Behav. 1999;3:91–97. [Google Scholar]
- Mukhin A. Post-juvenile moult and change in locomotory activity of caged reed warblers Acrocepahlus scripaceus from the early and the late broods prior migration. Avian Ecol. Behav. 2002;8:71–79. [Google Scholar]
- Mukhin A. Night movements of young reed warblers in summer—is it postfledging dispersal? Auk. 2004;121:203–209. [Google Scholar]
- Palmgren P. Studien über die Tagesrhythmik gekäfigter Zugvögel. Z. Tierpsychol. 1944;6:44–86. [Google Scholar]
- Rappole J.H, Tipton A.R. New harness design for attachment of radio-transmitters to small passerines. J. Field Ornithol. 1991;62:335–337. [Google Scholar]
- Sokolov L.V, Bolshakov K.V, Vinogradova N.V, Dolnik T.V, Lyuleeva D.S, Payevsky V.A, Shumakov M.E, Yablonkevich M.L. The testing of the ability for imprinting and finding the site of future nesting in young Chaffinches. Zool. J. 1984;43:1671–1681. (In Russian.) [Google Scholar]
- Wallraff H.G.Das Navigationssystem der Vögel. Ein theoretischer Beitrag zur Analyse ungeklärter Orientierungsleistungen1974R. Oldenbourg; München, Wien [Google Scholar]
- Widmer, M. 1999 Onset of autumn migratory activity: garden warblers Sylvia borin from the lowlands and the mountains show different responses to photoperiodic changes. Ph.D. thesis, Zürich: University Press, Zürich.
- Wiltschko W, Wiltschko R. The role of outward journey information in the orientation of homing pigeons. In: Papi F, Wallraff H.G, editors. Avian navigation. Springer; Berlin: 1982. pp. 75–89. [Google Scholar]
- Wiltschko W, Wiltschko R. Magnetic versus celestial orientation in migrating birds. Trends Ecol. Evol. 1988;3:13–15. doi: 10.1016/0169-5347(88)90076-6. [DOI] [PubMed] [Google Scholar]
- Wiltschko W, Wiltschko R. Interaction of magnetic and celestial cues in the migratory orientation of passerines. J. Avian Biol. 1998;29:606–617. [Google Scholar]
- Wiltschko W, Daum P, Fergenbauer-Kimmel A, Wiltschko R. The development of the star compass in garden warblers, Sylvia borin. Ethology. 1987;74:285–292. [Google Scholar]