

Abstract
A simple argument based on the distribution of individuals amongst discrete resource sites is used to show how the form of single species population models depends on the type of competition between, and the spatial clustering of, the individuals. For scramble competition between individuals, we confirm earlier demonstrations that the Ricker model is a direct consequence of a uniform random distribution of individuals across resources. By introducing spatial clustering of individuals according to a negative binomial distribution, we are able to derive the Hassell model. Furthermore, the tent map model is seen to be a consequence of scramble competition and an ideal-free distribution of individuals. To model contest competition under different degrees of spatial clustering we derive a new three-parameter model, of which the Beverton–Holt and Skellam models are special cases, where one of the parameters relates directly to the clustering distribution. Other population models, such as the quadratic model and the theta-Ricker models, cannot be derived in our framework. Taken together our derivations of population models allows us to make a more rigorous prescription for model choice when fitting to particular datasets.
Keywords: first-principles derivation, negative-binomial distribution, Beverton–Holt model, Skellam model Hassell model Ricker model
1. Introduction
The population dynamics of single species with seasonal reproduction and first-order feedback are often modelled using a single difference equation, at+1=f(at) (May 1976; Berryman 1999; Turchin 2003), with the natural interpretation that at+1 is the expected population in generation t+1 if at is the population in generation t. While there are a large number of these models, most of which are presented in table 1, each of them can be broadly classified as capturing either scramble or contest competition (Nicholson 1954; Hassell 1975). For scramble competition f increases to a maximum and then decreases, reflecting an over-compensation in the density‐dependence at large populations. For contest competition f is non-decreasing, reflecting an increasing utilisation of available resources. Both types of model are illustrated in figure 3. Utida (1967), Maynard-Smith & Slatkin (1973) and Hassell (1975) have all proposed different models incorporating both scramble and contest competition, each with an extra parameter that determines whether the model is scramble or contest. Apart from this scramble/contest classification of models, little is known about how—when faced with a single-species population time series—to choose the appropriate functional form for f.
Table 1.
The major single species population models studied in this paper.
| model | type | name | references | see equation |
|---|---|---|---|---|
| at(1+r(1−at/k)) | S | quadratic | May (1976) | (3.6) |
| at exp(r(1−at/k)) | S | Ricker, discrete logistic | Ricker (1954) | (1.2), (3.2) |
| at exp(r(1−(at/k)θ)) | S | theta-Ricker | Bellows (1981) and Berryman (1999) | (3.5) |
| see equation (3.9) | S | tent | Devaney (1989) | (3.9) |
| k(1−exp(−rat)) | C | Skellam | Skellam (1951) | (3.11) |
| k1at/(1+k2at) | C | Beverton–Holt | Beverton & Holt (1981) | (3.13) |
| see equation (3.12) | C | ‘ramp’ | this paper | (3.12) |
| bn(1−λλ(λ+at/n)−λ) | C | this paper | (3.14) | |
| k1 exp(−k2at)(exp(ck2at)−1)/h | S,C | this paper | (3.15) | |
| k1at/(1+k2at)c | S,C | Hassell | Hassell (1975) | (3.16) |
| S,C | Maynard-Smith–Slatkin | Maynard-Smith (1974) and Maynard-Smith & Slatkin (1973) | (3.17) |
S, scramble competition; C, contest competition.
Figure 3.
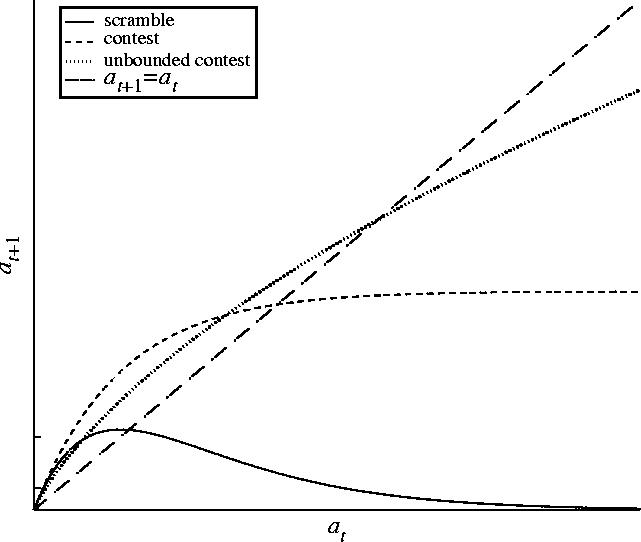
The three types of population model studied in this paper. Models of scramble competition increase to a global maximum and then decrease to 0, while models of contest competition are either increasing and bounded or strictly increasing and unbounded. We denote the latter ‘unbounded contest’. Also shown is the line at+1=at. Note that any biologically realistic unbounded contest model will eventually stay below this line, as it would otherwise be capable of unchecked population growth.
In order to determine the mechanistic underpinnings and thus the applicability, of a mathematical model, one can investigate how the model is derived from first principles. In his influential textbook, Royama (1992) presented a derivation of the Ricker model from first principles. He assumed that in discrete generations at identical individuals are distributed completely at random over a uniform resource environment with area A. He further assumed that the reproductive success of each individual is given by a function r(k) of the number of other individuals within a disc of area s centred on the individual. Since individuals are assumed identical, the expected growth of the population at+1/at equals the expected reproductive success of an individual. Thus,
| 1.1 |
where is the probability of an individual having exactly k neighbours in a disc with area s. Under these assumptions, as A→∞ with the population density at/A fixed, the number of individuals in a disc of area s is Poisson distributed with expectation sat/A and we can take . By assuming that r(k)=bck, equation (1.1) becomes
| 1.2 |
which is the Ricker model from table 1.
Such a ‘first principles’ derivation of a well studied population model gives the model biological grounding. The Ricker model is seen to be a consequence of animals distributed uniformly at random and having reproductive success rapidly reduced by competition with neighbours, i.e. scramble competition. The derivation also relates the parameters that govern individual behaviour to those governing population dynamics. Rewriting equation (1.2) in the more familiar form at+1=at exp(γ(1−at/K)) we see that the populations' carrying capacity K=A log(b)/(s(1−c)), is a function of both total resources (i.e. the area), intensity of competition and rate of reproduction.
Although the Ricker model is far from a unique model of discrete generation, single species population dynamics, Royama did not apply his derivation to any other models. In particular, he was ‘not able to formulate a model of contest competition in discrete time from basic principles’ (Royama 1992, p. 152). This is surprising, since other choices of r(k) produce contest models. For example, if r(k)=b/(k+1) the above derivation will give the Skellam model (Skellam 1951).
Until now, first principles derivations of models other than the Ricker model have mostly been made by integrating a corresponding continuous time model (Gurney 1998; Thieme 2003; Geritz & Kisdi 2004). In the most comprehensive work of this type, Geritz & Kisdi (2004) assume that the dynamics within years are given by a continuous-time resource–consumer system. The Ricker, truncated quadratic, Hassell and Beverton–Holt models are then obtained as the between-year dynamics. An important point to come out of the study is that without influx of resources the system is always over-compensatory, i.e. the discrete-time model is one of scramble competition. Thieme (2003) extends the classic derivation of the Ricker model from a spawner–recruitment context with cannibalism at birth, by considering other distributions of waiting time in the larval stage. The functional response of predators has also been derived from the principle of mass action under various assumptions about the spatial distribution of predators (Cosner et al. 1999).
While the continuous approach is appropriate in modelling, for example, fish populations, it does not capture the types of local interactions explicit in Royama's approach. In particular, the continuous model approach does not reveal how the spatial distribution or competitive interactions give rise to particular functional forms. This is important in modelling insect populations with seasonal reproduction and local resource competition (Berryman 1999). In this paper we extend a site-based framework proposed by Sumpter & Broomhead (2001) and Johansson & Sumpter (2003) and use it to derive most of the models in table 1, including the Beverton–Holt and Hassell models, from basic assumptions on the interactions between, and distribution of, individuals.
2. The site-based framework
Consider a habitat consisting of n discrete resource sites over which the at individuals in a population are distributed and then reproduce. We define pk to be the expected proportion of sites containing k individuals after the distribution, which is then a function of both at and n. Once the individuals are distributed, they reproduce with the success of reproduction dependent only on the number of individuals k at the site or more precisely with reproductive success assumed independent and identically distributed when conditioned on k. We let ϕ(k), henceforth referred to as the interaction function, be the expected number of individuals emerging from a site with k individuals at it. All emerging individuals are assumed to be identical and form the next generation. Figure 1 illustrates this process for the specific case of contest competition, to be discussed in depth later.
Figure 1.
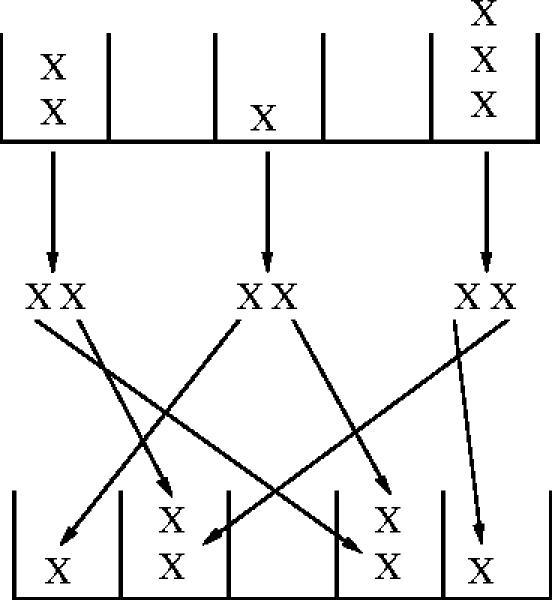
An illustration of the individual based model for contest competition. Here, at=6 individuals are distributed randomly among n=5 discrete resource sites giving the site-count (2, 0, 1, 0, 3). They interact according to the contest interaction function, equation (3.10), giving an intermediate site-count (ϕ(2), 0, ϕ(1), 0, ϕ(3))=(2, 0, 2, 0, 2), giving at+1=6 individuals in the next generation. These are then distributed randomly and the process is repeated.
We can now write the expected population at+1 in generation t+1 given that there are at individuals in generation t as
| 2.1 |
Here, we use the assumption that the number of individuals emerging from sites are independent and identically distributed. Note that in contrast with equation (1.1), this formulation is made in terms of reproduction per site, not reproduction per individual. Equation (2.1) relates a ‘first principles’ model of site-based reproduction, in terms of distribution and reproduction of individuals at resource sites, to a population model, f(at), of how a population changes through time. From a mathematical perspective, our aim now is to relate the probabilities pk and the interaction function ϕ(k) to specific population models such as the Ricker or Beverton–Holt models. From a biological perspective, these choices should realistically reflect both the distribution of individuals across their environment and the way in which individuals interact and in particular compete, for resources. Choosing pk and ϕ(k) will hopefully give us insight into the implicit assumptions that underlie population models.
Although individuals of most species are typically spatially aggregated (McArdle et al. 1976; Taylor et al. 1978; Greig-Smith 1983; He & Gaston 2000), it often suffices to assume a uniform random distribution, where each individual is equally likely to be found any of the resource sites. In this case, as n→∞ with the density at/n fixed, the number of individuals at a given site is Poisson distributed with expectation at/n, so that pk=(at/n)kexp(−at/n)/k!. Taking these as our probabilities pk we get
| 2.2 |
See Johansson & Sumpter (2003) for a more detailed discussion.
If individuals cannot be assumed to be uniformly distributed among sites, the negative binomial distribution is often appropriate for describing spatial aggregation (Boswell & Patil 1970; Perry & Taylor 1985; Krebs 1989). Specific examples include the distribution of red mites on apple leaves (Bliss & Fisher 1953) and parasites on hosts (Anderson & May 1978). By assuming that occupation of a site depends in some way on ‘accessibility’ we can derive, in Appendix A, the negative binomial distribution from first principles, giving
| 2.3 |
where λ is a positive parameter. The Poisson distribution is now the special case when λ→∞ and for λ=1 we obtain the geometric distribution given by
| 2.4 |
Indeed, as illustrated in figure 2, λ determines the degree of clustering among individuals, in the sense that p0 decreases with λ, so that when λ is small there will be more empty sites.
Figure 2.

Three special cases of the negative binomial distribution. (a) The Poisson distribution obtained when λ→∞. (b) A spatially aggregated distribution obtained for λ=3. (c) The geometric distribution, λ=1. The positive parameter λ determines the degree of clustering among individuals, in the sense that p0 decreases with λ, so that when λ is small there will be more empty sites.
Before we continue, in §3, by deriving the various models in table 1, we note that Royama's continuous-space framework, equation (1.1) and our site-based framework, equation (2.1), are equivalent under certain plausible assumptions. This is shown in Appendix B. When individuals are Poisson distributed, the result follows immediately. Assume ϕ(0)=0 and define r(k)=ϕ(k+1)/(k+1). Equation (1.1) then gives
Thus,
If we further substitute n=A/s, observing that ϕ(0)=0, we derive equation (2.2). The reverse derivation follows from the equivalence at each step.
3. Derivation of competition models
We now consider how specific population models of the three types shown in figure 3 relate to the individual-based framework given by equations (2.1) and (2.2).
(a) Scramble competition
Hassell (1975) writes ‘Ideally, scramble [competition] involves the exactly equal partitioning of the resource such that there is an abrupt change from complete survival to 100% mortality’. Assuming that each of the n resource sites in our model can maintain exactly one single individual then, by this definition,
| 3.1 |
so that b offspring are produced at a site containing exactly one individual, while if two or more individuals share a site they fail to reproduce due to interference. Substituting equation (3.1) into equation (2.2) gives,
| 3.2 |
the Ricker model. The Ricker model is thus a direct consequence of the uniform random distribution of individuals amongst resource sites and ‘ideal’ scramble competition. This derivation is identical to that in Sumpter & Broomhead (2001).
As in Royama's derivation of the Ricker model, equation (1.2), the carrying capacity K=n ln(b) is seen to be a function of both the number of available sites and the maximum number of offspring per individual. Recalling how Royama's reproduction function and the interaction function are related, r(k)=ϕ(k+1)/(k+1), Royama's scramble reproduction function r(k)=bck has as corresponding interaction function
| 3.3 |
Thus, equation (3.1) is actually a special case of equation (3.3), used by Royama, when c=0.
The interaction function, equation (3.1), also gives a scramble competition model when {pk} is distributed negative binomially. We substitute equations (3.1) and (2.3) into equation (2.1) to get
| 3.4 |
which is the Hassell model for scramble competition. Equating parameters with Hassell's original model (given in table 1) we see that c=λ+1. Thus, in agreement with Hassell's (1975) original derivation, we have found that c>1 captures scramble dynamics. More interestingly, we have also shown that the degree of spatial clustering, measured by λ in the negative binomial distribution, is determined by the value of the parameter c. A value of c only slightly larger than 1, thus indicates significant levels of spatial aggregation of individuals.
When trying to fit data which is thought to arise from scramble competition, the flexible theta-Ricker is commonly used
| 3.5 |
where θ is a positive constant (Bellows 1981; Berryman 1999). Royama (1992) argues that the case of θ>1 corresponds to an intensification of competition as the number of competing neighbours increases while θ<1 corresponds to a habituation to competition. However, under the assumption that individuals are uniformly distributed there is no interaction function independent of at that can reproduce equation (3.5) for non-integer values of θ, since higher order derivatives of xθ are undefined for x=0. The theta-Ricker model may eventually be derived from a distribution having θ as a parameter, with θ=1 as the Poisson distribution. However, the interaction function cannot be equation (3.1) since p1 would then be proportional to (at/n)exp(−(at/n)θ), which for sufficiently large at and small θ would make p1 exceed 1, thus making the distribution inadmissible. As yet we cannot determine a natural distribution that reproduces the theta-Ricker model.
Another well known scramble competition model is the quadratic (logistic) model,
| 3.6 |
where K is the carrying-capacity (fixed point). This model predicts that the population eventually becomes negative when r>4, but is sometimes used to model population dynamics by assuming r<4 and an initial population below the carrying capacity. Under the assumption of a Poisson distribution of individuals, we can work backwards to find the corresponding interaction function. First observe that K would have to be proportional to the number of sites n as the quadratic model can be written in the form Kg(at/K). Assuming n=γK sites we calculate the interaction function by first writing
then equating coefficients of equation (2.2) we get
| 3.7 |
This interaction function is negative when k>1+(r+1)/(γr). Since a negative number of offspring can occur when more than two individuals share the same resource site even if r<4, the quadratic model does not allow an obvious interpretation in terms of Poisson distributed individuals interacting at discrete sites even in this range.
A closely related model that does not predict negative populations is the truncated quadratic model (Geritz & Kisdi 2004),
The certain extinction predicted by this model is not possible in the site-based framework under a uniform distribution of individuals. To see this it suffices to note that if a Taylor-series vanishes on an interval, then all coefficients (in this case the corresponding interaction function) must be identical zero. Intuitively, in a Poisson distribution all interactions occur with some positive probability and since at least one interaction must produce in expectation a positive number of offspring, it follows that the expected population is also positive. Thus, we expect that any derivation of the truncated quadratic model in a site-based framework would be based on a more even distribution of individuals than the Poisson.
An example of a distribution, where we may have certain extinction for large populations is the ‘ideal-free’ distribution (Sutherland 1983), where every individual receives an equal share of the available resources. Writing at/n=[at/n]+(at/n−[at/n]), where [at/n] is the integer part of at/n, this gives the following distribution
| 3.8 |
With the scramble interaction function, equation (3.1), we get a scaled tent model,
| 3.9 |
It should be noted that since an animal population can at best only approximate the ideal-free distribution, we can only expect to find smooth approximations of the tent model in nature.
(b) Contest competition
Varley et al. (1973) write ‘In contest [competition] each successful animal gets all it requires, the unsuccessful animals get insufficient for survival or reproduction’. In this spirit we let the interaction function
| 3.10 |
so that b offspring are produced at a site containing one or more individuals. Substituting equation (3.10) into equation (2.2) gives
| 3.11 |
This is the Skellam model (Skellam 1951). Provided b>1, the model has a single stable equilibrium bn at which the population will quickly settle. It is a model of contest competition, since f is non-decreasing. The derivation presented here is essentially equivalent to Skellam's original derivation and has also appeared in other works, e.g. Johansson & Sumpter (2003).
With the ideal-free distribution given by equation (3.8), the contest interaction function, equation (3.10), gives linear growth up to a saturating population,
| 3.12 |
which we call the ‘ramp model’.
The standard model of contest competition is the Beverton–Holt model,
| 3.13 |
shown in figure 4 together with the Skellam and ramp model. A comparison between these models shows that they are similar in many ways. They are bounded functions with no inflection points. When at is small eat/n≈1+at/n≈1 so that growth at small populations is identical in the models. Furthermore, as at→∞, both n(1−e−at/n)→n and at/(1+at/n)→n so the populations are bounded by a population bn. However, when at and n are of a similar order of magnitude the three models give different numerical predictions. In particular, the Skellam model, equation (3.11), predicts that the population will grow faster than predicted by the Beverton–Holt model, equation (3.13).
Figure 4.
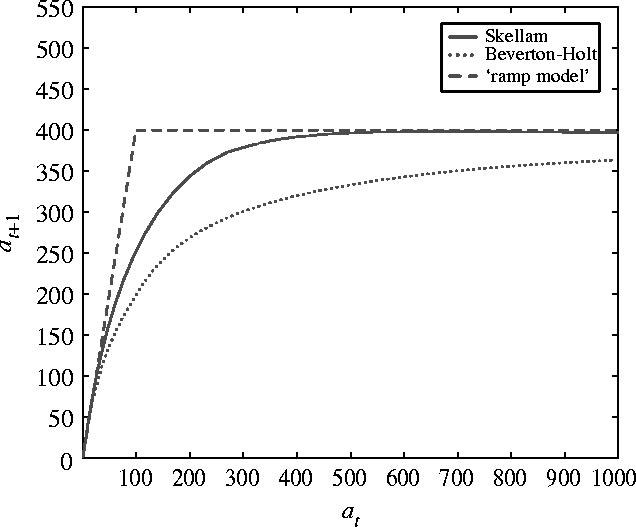
The Beverton–Holt, Skellam and ramp model for b=4 and n=100. The Beverton–Holt is the case λ=1 of equation (3.14) while the Skellam model is obtained in the limit as λ→∞. The ramp model is a consequence if contest competition, equation (3.10), under the ideal-free distribution, equation (3.8).
The Beverton–Holt model can be derived from equation (2.1) if we assume that the probabilities {pk} are distributed negative binomially. Substituting equation (3.10) into equation (2.1) with {pk} given by equation (2.3) we get
| 3.14 |
The direct inclusion of the parameter λ from the underlying distribution offer some interesting insights. For λ=1 this is the Beverton–Holt model. Thus, the Beverton–Holt model is a consequence of clustering and contest competition. As λ increases, the underlying distribution is more uniform and in the limit as λ→∞ the model converges pointwise to the Skellam model.
(c) Generalized competition models
We can derive a competition model encompassing both scramble and contest competition from the interaction function,
where 0<h<1. Equation (2.2) then gives
| 3.15 |
In the limit, as c→0+ or c→1− we recover the Ricker model, equation (3.2) and the Skellam model, equation (3.11), respectively. The main disadvantage of the model is that it does not interpolate between scramble and contest competition in a natural way. Indeed, for any c<1 both equation (3.15) and its interaction function are of scramble competition with exponential decline.
Models that interpolate naturally between scramble and contest can be constructed by taking linear combinations of either scramble or contest models or of their respective interaction functions. However, the algebraic complexity of these models may make them less useful.
Two well known population models that incorporate both scramble and contest competition are the Hassell (Hassell 1975) and the Maynard-Smith–Slatkin (Maynard-Smith & Slatkin 1973) model given by
| 3.16 |
and
| 3.17 |
respectively. The structural similarity of the models is reflected in the identical growth rate of small and large populations, k1 and , respectively. The parameter c>0 determines the competition type. For c>1 both are models of scramble competition and for c=1 both reduce to the Beverton–Holt model of contest competition. For 0<c<1 both are models of unbounded contest competition. This is in contrast with the contest models studied so far, which have all been bounded so that there is a fixed limit on next year's population size independent of the current population.
Although we derived the Hassell model for c≥1 from first principles in §3a, we have not yet seen any example of unbounded contest competition. However, by taking the interaction function to be
| 3.18 |
and the probabilities pk as in equation (2.3) we can derive the Hassell model. Note that for λ=1 this is the contest interaction function, equation (3.10). No corresponding first principles derivation of the Maynard-Smith–Slatkin model has yet been discovered and this remains an open problem.
4. Discussion
The growing interest in the first principles derivations of population models is effecting a change in how population models are viewed. Traditionally, models have been treated phenomenologically, whereas first principles derivations demonstrate that these models are consequences of simple underlying mechanistic principles, most of which can be understood intuitively. Taking a first principles approach leads to new insights, such as relations between disparate models, as well as how individual actions effect population dynamics.
We have extended Royama's (1992) site-based framework to incorporate spatial distributions of aggregated individuals. In particular, we have shown that the degree of spatial aggregation can be related to parameters in a corresponding population dynamic model. This is most clearly illustrated by our three-parameter generalization of the Beverton–Holt model, equation (3.14). The parameter λ in this model can be interpreted as an index of spatial aggregation, with high values corresponding to a more random distribution. Fitting equation (3.14) to ecological time-series of species exhibiting local contest competition may help to determine the level of clustering of the population, without detailed knowledge of how the population is actually structured.
The ideal-free distribution of individuals also produces its own associated population dynamics under different types of competition. Figure 4 shows how we would expect a population under an ideal-free distribution and contest competition to grow in comparison to a population with random or spatially aggregated distribution of individuals. For scramble competition the ideal-free distribution gives rise to the tent model. Of course, the ideal-free distribution is idealized and we would expect the actual dynamics of a naturally ideal-free distributed population to follow something between a tent and a Ricker model.
For scramble competition with individuals distributed according to a negative binomial distribution we derived the Hassell model for scramble competition. Again the parameters of the Hassell model could be related to spatial clustering. In particular, the parameter c was shown not only to relate to competition type, but also to the degree of spatial clustering. The Maynard-Smith–Slatkin model, renowned for its ability to fit data, evaded a derivation altogether and there is no known mechanistic interpretation of this model, even in the continuous time frameworks of Thieme (2003) or Geritz & Kisdi (2004). The theta-Ricker model also evaded derivation and it is difficult to see how a derivation of the type presented here might proceed.
The current paper has limited itself to single generation, single species models with contest and scramble competition. Allee effects can be added to these models in a straightforward manner by, for example, requiring that two or more individuals are needed per resource site for offspring to be produced (Johansson & Sumpter 2003). Such a model would be appropriate in modelling reproduction involving two sexes. The ideas presented here could also be extended to more complex scenarios such as predator–prey or age-structured interactions. Several attempts have been made to derive the functional response of predators in predator–prey models, the mating function and the contact function in sexually transmitted diseases from underlying spatial distribution (Castillo-Chavez et al. 1994; Cosner et al. 1999; Thieme & Yang 2000, see also the references therein).
The primary aim of this paper has been to heighten awareness of the implicit assumptions about competition and clustering that underlie single species population models. The results presented are at their most useful when a particular model has been proved good in fitting population time-series data and we would like to infer something about the ecology of the studied species. Knowledge of the different assumptions that underlie population models should also simplify model choice. By deriving the functional form of a model through a ‘first principles’ description of interactions between, and distribution of, individuals, the assumptions that underlie the model can be better understood. Such a procedure reveals not only facts about the mathematical structure of models but also biological insight into when a particular model is appropriate.
Acknowledgements
The authors would like to thank Éva Kisdi and Anders Johansson for useful comments and discussion and Paul Glendinning for carefully reviewing and correcting an earlier version of the manuscript. We also thank the two anonymous referees for many helpful suggestions.
Appendix A
Appendix A. Derivation of the negative binomial distribution
By revoking the assumption that sites are in all respects identical, we are able to derive the negative binomial distribution from first principles. Assume that an individual chooses a given site with a probability proportional to the site's accessibility, as determined by a number drawn from a continuous probability distribution f with an expectation of unity. As the number of sites n tends to infinity with the density x constant, the number at individuals at a site with value t will be Poisson-distributed with expectation tx. The expected number of individuals at a site for which the value t is not known will be distributed as fx(t)=(1/x)f(t/x). Assuming that accessibility of the sites are distributed as
then the expected number of individuals at a site is Gamma distributed,
| A1 |
Since sites are chosen randomly, albeit with different probabilities, the probability of finding k individuals at a site is given by
This is known as mixing with the Poisson distribution (Kendall & Stuart 1977). We then get
which is the negative binomial distribution formulated in terms of the expectation x, i.e. equation (2.3).
Appendix B
Appendix B. Equivalence of the site-based and Royama's continuous-space framework
The site-based framework
| B1 |
and Royama's continuous-space framework
| B2 |
are equivalent in the sense that (i) for any positive solution {at} of the dynamical system (B 1) with ϕ(0)=0 we can find an admissible distribution and a reproduction function r(k) such that {at} is also a solution of the dynamical system (B 2) and (ii) for any positive bounded solution {at} of the dynamical system (B 2) we can find an admissible distribution pk and a constant n such that {at} is also a solution of the dynamical system (B 1). We consider a distribution pk admissible if it is non-negative and pk and kpk sums to 1 and at/n, respectively. We consider a distribution admissible if it can be realized by distributing individuals for some value of A and s or if it can be realized in the limit as A→∞ with the density at/A constant.
Given any way of distributing individuals among sites in the discrete setting, we can mimic this in Royama's continuous setting by considering n points in a region of total area A. Choose s sufficiently small such that discs of area s centred at these points do not intersect. These points are then interpreted as the sites in the discrete setting. If we distribute at individuals among these ‘sites’ such that the expected fraction of sites with exactly k individuals is pk, then
Thus,
Substituting into equation (B 2) gives
If we let r(k)=ϕ(k+1)/(k+1), change summation indices and observe that ϕ(0)=0, then
which equals equation (B 1).
Assume now that {at} is a solution of the dynamical system (B 2), bounded by some n. Let for k≥1 and , where the sum is taken over the positive integers. This is an admissible distribution, since
Let ϕ(0)=0 and ϕ(k)=kr(k−1) for k≥1. Then, after substituting into equation (B 2) we get equation (B 1).
Footnotes
As this paper exceeds the maximum length normally permitted, the authors have agreed to contribute to production costs.
Present address: Department of Information and Computer Sciences, Nara Women's University, Kita-Uoya Nishimachi, 630-8506 Nara, Japan.
Present address: Department of Zoology, South Parks Road, Oxford OX1 3PS, UK.
References
- Anderson R.M, May R.M. Regulation and stability of host–parasite population interactions: I. Regulatory processes. J. Anim. Ecol. 1978;47:219–247. [Google Scholar]
- Bellows T.S. The descriptive properties of some models for density dependence. J. Anim. Ecol. 1981;50:139–156. [Google Scholar]
- Berryman A.A. Thornes; Cheltenham: 1999. Principles of population dynamics and their applications. [Google Scholar]
- Beverton R.J.H, Holt S.J. Fisheries investment. Series 2, vol. 19. Her Majesty's Stationary Office; London: 1981. On the dynamics of exploited fish populations. [Google Scholar]
- Bliss C.I, Fisher R.A. Fitting the negative binomial distribution to biological data and a note on efficient fitting of the negative binomial. Biometrics. 1953;9:176–200. [Google Scholar]
- Boswell M.T, Patil G.P. Chance mechanisms generating the negative binomial distributions. In: Patil G.P, editor. Random counts in scientific work. vol. 1. Pennsylvania State University Press; University Park, PA: 1970. pp. 3–22. [Google Scholar]
- Castillo-Chavez C, Velasco-Hernandez J.X, Fridman S. Modeling contact structures in biology. In: Levin S.A, editor. Frontiers in theoretical biology Lecture notes in biomathematics. vol. 100. Springer; Berlin: 1994. pp. 454–491. [Google Scholar]
- Cosner C.I, DeAngelis D, Ault J.A, Olson D.B. Effects of spatial grouping on the functional response of predators. Theor. Popul. Biol. 1999;56:65–75. doi: 10.1006/tpbi.1999.1414. 10.1006/tpbi.1999.1414 [DOI] [PubMed] [Google Scholar]
- Devaney R.L. 2nd edn. Addison-Wesley; Redwood City, CA: 1989. An introduction to chaotic dynamical systems. [Google Scholar]
- Geritz S.A.H, Kisdi E. On the mechanistic underpinning of discrete-time population models with complex dynamics. J. Theor. Biol. 2004;228:261–269. doi: 10.1016/j.jtbi.2004.01.003. 10.1016/j.jtbi.2004.01.003 [DOI] [PubMed] [Google Scholar]
- Greig-Smith P. 3rd edn. Blackwell Scientific; Oxford: 1983. Quantitative plant ecology. [Google Scholar]
- Gurney W.S.C. Oxford University Press; New York: 1998. Ecological dynamics. [Google Scholar]
- Hassell M.P. Density-dependence in single-species populations. J. Anim. Ecol. 1975;45:283–296. [Google Scholar]
- He F, Gaston K.J. Estimating species abundance from occurrence. Am. Nat. 2000;5:553–559. doi: 10.1086/303403. 10.1086/303403 [DOI] [PubMed] [Google Scholar]
- Johansson A, Sumpter D.J.T. From local interactions to population dynamics in site-based models of ecology. Theor. Popul. Biol. 2003;64:497–517. doi: 10.1016/s0040-5809(03)00076-5. 10.1016/S0040-5809(03)00076-5 [DOI] [PubMed] [Google Scholar]
- Kendall M, Stuart A. Distribution theory. 4th edn. vol. 1. Macmillan Publishing Co., Inc; New York: 1977. The advanced theory of statistics. [Google Scholar]
- Krebs C.J. Harper Collins; New York: 1989. Ecological methodology. [Google Scholar]
- May R.M. Simple mathematical models with very complicated dynamics. Nature. 1976;261:459–467. doi: 10.1038/261459a0. [DOI] [PubMed] [Google Scholar]
- Maynard-Smith J. Cambridge University Press; Cambridge: 1974. Models in ecology. [Google Scholar]
- Maynard-Smith J, Slatkin M. The stability of predator–prey systems. Ecology. 1973;54:384–391. [Google Scholar]
- McArdle B.H, Gaston K.J, Lawton J.H. Variation in the size of animal populations: patterns, problems and artefacts. J. Anim. Ecol. 1976;59:439–454. [Google Scholar]
- Nicholson A.J. An outline of the dynamics of animal populations. Aust. J. Zool. 1954;2:9–65. 10.1071/ZO9540009 [Google Scholar]
- Perry J.N, Taylor L.R. Adès: new ecological families of species–species frequency distributions that describe repeated spatial samples with an intrinsic power-law variance–mean property. J. Anim. Ecol. 1985;54:931–953. [Google Scholar]
- Ricker W.E. Stock and recruitment. J. Fisheries Res. Board Can. 1954;11:559–623. [Google Scholar]
- Royama T. Chapman & Hall; London: 1992. Analytical population dynamics. [Google Scholar]
- Skellam J.G. Random dispersal in theoretical populations. Biometrika. 1951;38:196–218. [PubMed] [Google Scholar]
- Sumpter D.J.T, Broomhead D.S. Relating individual behaviour to population dynamics. Proc. R. Soc. B. 2001;268:925–932. doi: 10.1098/rspb.2001.1604. 10.1098/rspb.2001.1604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutherland W.J. Aggregation and the ‘ideal free distribution’. J. Anim. Ecol. 1983;52:821–828. [Google Scholar]
- Taylor L.R, Woiwod I.P, Perry J.N. The density dependence of spatial behaviour and the rarity of randomness. J. Anim. Ecol. 1978;47:383–406. [Google Scholar]
- Thieme H.R. Princeton series in theoretical and computational biology. Princeton University Press; Princeton, NJ: 2003. Mathematics in population biology. [Google Scholar]
- Thieme H.R, Yang J. On the complex formation approach in modeling predator prey interactions, mating and sexual disease transmission. Proc. conf. on nonlinear differential equations. Electronic J. Differ. Equations. 2000;5:255–283. [Google Scholar]
- Turchin P. Princeton University Press; Princeton, NJ: 2003. Complex population dynamics: a theoretical/empirical synthesis. [Google Scholar]
- Utida S. Damped oscillation of population density at equilibrium. Res. Popul. Ecol. 1967;9:1–9. [Google Scholar]
- Varley G.C, Gradwell G.R, Hassell M.P. Blackwell Scientific; Oxford: 1973. Insect population ecology. [Google Scholar]