

Abstract
Our experiments, carried out at night and during the day on adults and laboratory-born young of the sandhopper Talitrus saltator, deal with the identification and use of the moon as an orientating factor. Sandhoppers were released in an apparatus (a Plexiglas dome) that produced a scenario similar to the natural one (with artificial sky, moon or sun illuminated at different intensities).
When tested at night, the adult and young sandhoppers used the artificial moon like the natural one, independently of the intensity of illumination of the artificial sky and moon. In other words, sandhoppers tested at night always identified the artificial moon as the moon and never as the sun. In daytime releases, the seaward orientation failed at low intensities of artificial sky and sun illumination (3.07 and 1.55 μW cm2, respectively), whereas the sun compass was used effectively at higher levels of artificial sun and sky illumination. The innate ability of moon compass orientation in inexpert young sandhoppers was demonstrated even under artificial light.
Keywords: sandhoppers, Talitrus saltator, orientation, moon, sun, artificial light
1. Introduction
It has long been known that sandhoppers use astronomical and celestial cues to maintain the shortest route along the sea–land axis of the beach during their excursions (see Pardi & Papi 1953). We recently returned to the topic of moon compass orientation in sandhoppers and were able to fully confirm the use of the moon in orientation along the sea–land axis of the beach (Ugolini et al. 1999a,b, 2002a, 2003). Our findings also showed the separation of the two chronometric mechanisms used to compensate for solar and lunar azimuthal variations (Ugolini et al. 1999b), as well as the innate basis of the moon compass (Ugolini et al. 2003; for the innate ability of direction finding based on the sun compass, see Pardi et al. 1958; Pardi 1960; Pardi & Ercolini 1986).
In various attempts to obtain correct solar orientation under artificial light in supralittoral arthropods (Pardi 1954, 1958; Terracini-Debenedetti 1958), the results failed to conform to expectations: individuals were photopositive or exhibited mean angles smaller than those expected in tests of sun compass orientation in natural conditions carried out at the same time of day. A few years ago (Ugolini et al. 1998), we built an apparatus designed to reproduce sun compass orientation in the laboratory corresponding to that recorded in natural conditions. We used a Plexiglas dome that produced a scenario with variable intensities of artificial sun and sky illumination. Experiments with the sandhopper Talitrus saltator (Ugolini et al. 1998) and the isopod Tylos europaeus (Ugolini & Castellini 2004) were successful when the intensities of the artificial sun and sky exceeded certain threshold values.
In this paper, we test the possibility of obtaining good moon compass orientation under artificial light conditions in adult and young individuals of T. saltator. Our interest was prompted by the fact that nobody has attempted to carry out similar experiments, because of (i) their doubts about the real ability of moon orientation and (ii) the difficulty in evaluating the true use of the moon as an orientating factor rather than the possible use of other celestial cues, like nocturnal skylight polarization (Gal et al. 2001; for the use of moonlight polarization in dung beetle orientation, see Dacke et al. 2003, 2004).
2. Material and Methods
We used adult and young individuals of T. saltator. Adults were collected on a sandy beach in southern Tuscany near the Albegna River mouth (Grosseto, Italy) during the last quarter or new moon. The sea–land direction of the Y-axis (perpendicular to the shoreline) is 268–88°.
The sandhoppers were transported to the laboratory and transferred to Plexiglas containers with wet sand. They were fed weekly with dry fish food on blotting paper. The animals were subjected to an artificial light cycle corresponding in phase and duration to the natural one. There was no artificial dusk or dawn, nor an artificial moon. Daytime illumination was provided by neon tubes (light intensity 65 μW cm2).
The experiments were conducted in a dark room in the laboratory from late spring to early autumn 1996–1998 and 2000, in coincidence with a natural moon phase ranging from 94 to 99% (i.e. nearly full moon). The moon's azimuthal variation during the various experiments is represented outside the circular distributions. Releases were carried out using an experimental apparatus that previously allowed us to reproduce sun compass orientation under artificial light conditions that was comparable to the orientation under the natural sun and sky (figure 1; Ugolini et al. 1998; Ugolini & Castellini 2004). In short, the apparatus consisted of a circular plate of white Plexiglas (diameter 80 cm) with a central part of transparent Plexiglas (29 cm diameter). As there is no inter-individual influence on the directional choice in sandhoppers (Scapini et al. 1981), 8–12 individuals at a time were released in a transparent Plexiglas bowl (18 cm diameter) located on the transparent plate, which allowed vision of the animals and the recording of their directions from below. A single direction per individual was recorded 1–2 min after release by means of a video camera underneath the bowl or by directly reading the direction on a goniometer beneath the bowl. A hemispherical dome of white glazed Plexiglas (diameter 80 cm) covered the circular plate. An optical fibre bundle (8 mm diameter) with a negative lens (focal length 11.7 mm, 21 mm diameter) was placed through the central transparent portion of the dish so that it stuck into the bowl to 28 mm. Its function was to illuminate the inner part of the dome (=artificial sky). A second optical fibre bundle (=artificial moon, 4.5 mm diameter) was inserted into the wall of the dome from the outside, at a height corresponding to around 45° from the plane containing the bowl and the animals. The fibre bundles were lit with two variable intensity illuminators (Schott KL 1500). In some releases a black glazed Plexiglas circular plate was added to the floor of the apparatus to reduce the artificial sky intensity. Some tests were carried out using a black Plexiglas dome. The azimuths of the artificial ‘moon’ or ‘sun’ corresponded to those of the natural ones at the moment of the releases (±5°).
Figure 1.

Experimental apparatus used to reproduce astronomical orientation under artificial light conditions. (a) external view: a, white Plexiglas dome (80 cm in diameter); b, Plexiglas plate; c, optical fibre bundle used to simulate astronomical orientating cues (the sun and the moon). (b), internal view: a, glazed Plexiglas dome; b, Plexiglas plate with c, a central part of transparent Plexiglas; d, optical fibre bundle with a negative lens to illuminate the inner part of the dome (=artificial sky). e, transparent Plexiglas bowl in which the sandhoppers were released.
The different light intensities used in the experiments were measured with a radiometer (Graseby S370, with a silicon photometer and a flat response between 350 and 1100 nm). The light distribution inside the dome was uniform, without asymmetries on the horizontal plane that could have influenced the directional choices of the animals (for a description of the distribution of light intensity inside the dome see Ugolini et al. 1998). Because of the difference in the emitting surfaces, the artificial moon (or sun) was far more brilliant than the artificial sky.
In the previous study (Ugolini et al. 1998), a set of filters was placed in front of the radiometer sensor to better compare the sandhoppers' behaviour with their spectral sensitivity curve (see Ugolini et al. 1996 for a preliminary investigation). For the present experiment, we decided not to use any filters in order to render the experimental conditions easier to reproduce. Moreover, we carried out a new set of measures with a different geometry: the sky intensity was measured while the artificial ‘moon’ was screened, and the intensity of the artificial moon was measured while the sensor head was pointed directly at it.
About one month after being collected, adult females were allowed to mate with males of the same population. The young sandhoppers were kept in the laboratory in the same conditions as the adults, i.e. without any possibility of seeing the natural sky and sun. We only tested inexpert young sandhoppers (born in the laboratory and thus lacking experience in the wild), exposed for the first time to the artificial sky and sun at the time of release, 15–30 days after hatching. They had never been exposed to the natural sky, sun or moon.
The statistical analysis was carried out with the methods described by Batschelet (1981). For each set of directions recorded in each release, the length and mean angle of the mean vector were calculated. The uniformity of the distributions was evaluated by Rao's test (p>0.05). The bimodality was assessed by the method of doubling the angles (Batschelet 1981), even though it does not provide the best description of the distribution of the directions.
For each release, we also calculated the seaward Y axis tendency (YSc), equal to the homeward component: YSc=r cos(YS−α), where r is the mean vector length, YS is the seaward direction of the home beach (theoretically expected for perfect orientation), α is the mean angle of the resulting vector (see Batschelet 1981, p. 42). In the case of bimodality, the YSc was calculated on the basis of the nearest α to the expected seaward direction.
The influence of the different light intensities on orientation was evaluated by comparing the various sets of YSc: sets with the number of releases n≥3 were compared by the Kruskal–Wallis one-way non-parametric analysis of variance, those with the number of releases n=2 by the Wilcoxon Mann–Whitney test for unpaired data (see Siegel & Castellan 1988).
3. Results
The results of the releases of adult sandhoppers at night under the white dome are shown in figure 2. It is quite clear that there is no significant difference in YSc among the various combinations of intensities of artificial sky and ‘moon’ illumination: none of the comparisons between groups with n≥3 releases reaches statistical significance (critical value=40.05, ranks mean difference=39.44 at best, p=0.058, Kruskal–Wallis test). The good YS orientation of sandhoppers is confirmed by the circular distributions (figure 3): the difference between the mean angle and YS is only between 0 and 36°.
Figure 2.
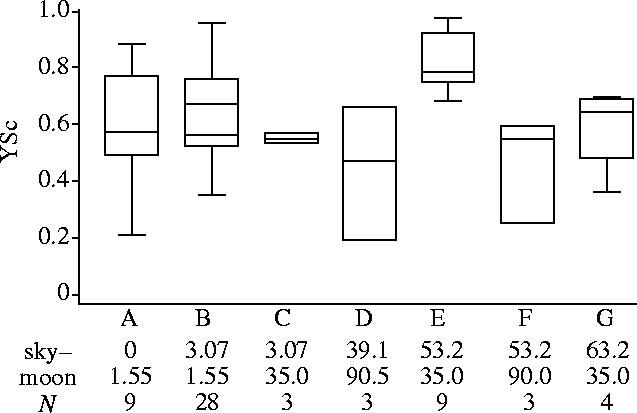
Goodness of orientation (YSc) at different intensities of artificial sky and moon illumination (given in μW cm2) in releases of adult sandhoppers at night under the white dome. N, number of releases. The box represents 50% of the data, the whiskers show the range (largest and smallest observations). The horizontal line within the box represents the median. Letters and data reported below the abscissae refer to the circular distributions of figure 3.
Figure 3.

Circular distributions for each intensity of artificial sky and moon illumination reported in figure 1 (the same letter indicates the same level of illumination). Each dot corresponds to the direction assumed by one individual. The arrow inside the distributions is the mean resultant vector (the length of the radius corresponds to vector length=1) with the mean angle. N, magnetic North; black triangle, seaward direction of the home beach. The line outside each distribution represents the azimuthal variation of the artificial moon during the experiments. n, sample size; u, V-test value with the probability level, p.
The results of releases carried out under the black dome (figure 4) are similar to those obtained at night under the white dome at the same artificial moon and sky intensities (1.55 and 0 μW cm2, respectively, figure 2, A). The mean angle differs from YS by 36° under the black dome, and the comparison of the YSc values does not reach significance (z=−0.486, p=0.627, Wilcoxon Mann–Whitney test).
Figure 4.

Circular distribution of releases of adult sandhoppers under the black dome (sky 0 μW cm2, moon 1.55 μW cm2). For further explanation, see figure 3.
Tests carried out with adults under the white dome during the day (figure 5) clearly show that the seaward orientation depends on the intensities of the artificial sky and sun. There is a significant difference in the goodness of orientation (YSc) at sky and sun intensities of 3.07 and 1.55 versus 63.3 and 35 μW cm2 (figure 5, A versus B) or higher (figure 5, C) (χKW2=7.848, d.f.=2, p=0.019, Kruskal–Wallis test). This is confirmed by the corresponding circular distributions (figure 5): the sandhoppers' orientation towards YS is statistically significant only for ‘diurnal’ artificial sky/sun intensities ≥63.3/35 and 63.3/169.5 μW cm2.
Figure 5.

Releases of adult sandhoppers during the day under the white dome: goodness of orientation (YSc) and circular distributions at different intensities of artificial sky and sun illumination (in μW cm2). For further explanation, see figure 3.
The influence of light intensity on compass orientation is also underlined by the comparison of YSc values obtained in night and day releases of adults at the same light intensity. In fact, while the comparison of YSc at high light intensities is not significant (W=14, p=0.881, Wilcoxon Mann–Whitney test), there is a significant difference at light intensities of 3.07 μW cm2 (sky) and 1.55 μW cm2 (moon, sun) (figure 5, A versus figure 2, A; z=3.252, p=0.0011, Wilcoxon Mann–Whitney test).
To verify that the sandhoppers' ability to maintain the correct seaward direction is due to vision of the artificial moon and not to some other orienting reference, we released some groups of individuals with the azimuth of the artificial moon deviated by 90° with respect to that of the real moon at the same hour (figure 6). The results clearly indicate that controls (tested with the azimuth of the artificial moon equal to that of the real moon at the time of release) are well directed towards the seaward direction of their home beach; the difference between YS and the mean angle is only 6°. However, in the experimental tested with the azimuth of the artificial moon deflected 90° counterclockwise, the mean angle differs from the new expected YS by only 16°, but by 74° from the YS direction of their home beach.
Figure 6.

Circular distributions of releases of adult sandhoppers at night under the white dome (sky 53.2 μW cm2, moon 35.0 μW/cm2). In (a) the artificial moon's azimuth corresponds to the azimuth of the natural moon; in (b) the artificial moon's azimuth is deflected by 90°. Black triangle, seaward direction of the home beach; white triangle, new expected direction based on the deflected lunar azimuth. In (b) the V-test value (u) refers to the white triangle. For further explanation, see figure 3.
The young sandhoppers tested at night under the white dome were also able to use the artificial source of illumination for correct moon compass orientation (figure 7). There is no significant difference among their YSc values (χKW2=0.802, d.f.=2, p=0.669, Kruskal–Wallis test): the young sandhoppers are well concentrated around their seaward direction despite the variations in artificial sky and moon intensities. Moreover, none of the comparisons between the YSc values of young and adult sandhoppers tested at night at the same light intensities reaches significance (figure 2 versus figure 7; W=26, p=0.147 at best, Wilcoxon Mann–Whitney test).
Figure 7.

Releases of inexpert young sandhoppers at night under the white dome: goodness of orientation (YSc) and circular distributions at different intensities of artificial sky and sun illumination (in μW cm2). For further explanation, see figure 3.
4. Discussion
First of all, it should be underlined that we had no intention of reproducing the exact conditions or levels of illumination of the natural sky and moon or sun at different hours of the night or day. Nevertheless, our apparatus is good enough to obtain sandhopper orientation similar to that obtained under the natural sky and sun (see also Ugolini et al. 1998) or moon.
It should also be mentioned that the orientation ability of T. saltator in the experiments carried out at night is independent of the combination of intensities of artificial sky–moon illumination. Moreover, the orientation ability in releases at night does not vary appreciably even when the intensities of artificial sky and moon illumination correspond to the intensities allowing good sun compass orientation in daytime releases. Obviously such orientation is not based on a ‘sun at night’. In fact, at night the chronometric mechanism of the sun compass in T. saltator compensates for a sun moving counterclockwise from West to East, passing through South (Pardi 1954, Ugolini et al. 2002a,b). Therefore, the expected direction of orientation is different.
Tests of adults carried out during the day under the white dome fully confirm that the orientation depends on the artificial sky/sun intensities (see also Ugolini et al. 1998): below certain intensity thresholds, the sun compass no longer operates and the orientation is based on simpler phototactic behaviour.
Talitrus saltator can very probably utilize skylight polarization under natural conditions during the day (Pardi & Papi 1953, Ugolini et al. 1996). Although moonlight is polarized (Gal et al. 2001; Horvath & Varjù 2004) and can be used by dung beetles for nocturnal orientation (Dacke et al. 2003, 2004), our experiments clearly demonstrate that T. saltator does not primarily rely on moonlight polarization to assume the correct seaward orientation (also see the experiments carried out under the light of an electric torch and the natural sky: Papi 1960; Ugolini et al. 1999a; Ugolini 2003). This is not surprising since the level of polarized light at night is very low (Gal et al. 2001; Horvath & Varjù 2004) and even during the day sandhoppers are perfectly able to orient in the ecologically correct direction under artificial illumination (thus without any meaningful information from the polarized light pattern).
In conclusion, the factors by which sandhoppers identify the moon and sun are, for the moment, not clearly understood. Nevertheless, sandhoppers tested at night identify the artificial moon as the moon and never as the sun, independently of the intensity of the artificial moon–sun and sky illumination. The same does not hold true for daytime releases. Finally, young sandhoppers born in the laboratory behave in the same way as adults collected in the wild, which confirms their innate ability of moon compass orientation (Ugolini et al. 1999a, 2003).
Acknowledgments
We would like to thank Drs S. Felicioni, C. Melis and Mrs T. Fantini for their help during the experiments. We are particularly grateful to Mr R. Innocenti for his technical assistance. This research was funded by the University of Firenze (local funds).
References
- Batschelet E. Academic Press; London: 1981. Circular statistics in biology. [Google Scholar]
- Dacke M, Nilsson D.E, Scholtz C.H, Byrne M.J, Warrant E.J. Insect orientation to polarized moonlight. Nature. 2003;424:33. doi: 10.1038/424033a. 10.1038/424033a [DOI] [PubMed] [Google Scholar]
- Dacke M, Byrne M.J, Scholtz C.H, Warrant E.J. Lunar orientation in a beetle. Proc. R. Soc. B. 2004;271:361–365. doi: 10.1098/rspb.2003.2594. 10.1098/rspb.2003.2594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gal J, Horvath G, Barta A, Wehner R. Polarization of the moonlit clear night sky measured by full-sky imaging polarimetry at full moon: comparison of the polarization of moonlit and sunlit skies. J. Geophys. Res. D. 2001;106:22647–22653. [Google Scholar]
- Horvath G, Varjù D. Spinger; Berlin: 2004. Polarized light in animal vision. [Google Scholar]
- Papi F. Orientation by the night: the moon. Cold Spring Harb. Symp. Quant. Biol. 1960;25:475–480. doi: 10.1101/sqb.1960.025.01.049. [DOI] [PubMed] [Google Scholar]
- Pardi L. Esperienze sull'orientamento di Talitrus saltator (Montagu) (Crustacea–Amphipoda): l'orientamento al sole degli individui a ritmo nictiemerale invertito durante la ‘loro notte’. Boll. Ist. Mus. Zool. Univ. Torino. 1954;4:127–134. [Google Scholar]
- Pardi L. Esperienze sull'orientamento solare di Phaleria provincialis Fauv. (Coleoptera): il comportamento a luce artificiale durante l'intero ciclo di 24 ore. Atti Accad. Sci. Torino. 1958;92:1–8. [Google Scholar]
- Pardi L. Innate components in the solar orientation of littoral amphipods. Cold Spring Harb. Symp. Quant. Biol. 1960;25:394–401. doi: 10.1101/sqb.1960.025.01.042. [DOI] [PubMed] [Google Scholar]
- Pardi L, Ercolini A. Zonal recovery mechanism in talitrid crustaceans. Boll. Zool. 1986;53:139–160. [Google Scholar]
- Pardi L, Papi F. Ricerche sull'orientamento di Talitrus saltator (Montagu) (Crustacea, Amphipoda). I. L'orientamento durante il giorno in una popolazione del litorale tirrenico. Z. Vergl. Physiol. 1953;35:459–489. [Google Scholar]
- Pardi L, Ercolini A, Marchionni V, Nicola C. Ricerche sull'orientamento degli anfipodi del litorale: il comportamento degli individui allevati in laboratorio sin dall'abbandono del marsupio. Atti Accad. Sci. Torino. 1958;92:1–8. [Google Scholar]
- Scapini F, Ugolini A, Pardi L. Analysis of astronomical orientation in littoral Amphipods using individual and group tests (Crustacea, Amphipoda) Monitore Zool. Ital. (NS) 1981;15:77–86. [Google Scholar]
- Siegel S, Castellan N.J., Jr . McGraw-Hill; New York: 1988. Nonparametric statistics for the behavioral sciences. [Google Scholar]
- Terracini-Debenedetti E. Orientational response of some Amphipods under artificial light. Boll. Ist. Mus. Zool. Univ. Torino. 1958;6:21–28. [Google Scholar]
- Ugolini A. Activity rhythms and orientation in sandhoppers (Crustacea, Amphipoda) Front. Biosci. 2003;8:s722–s732. doi: 10.2741/1041. [DOI] [PubMed] [Google Scholar]
- Ugolini A, Castellini C. Sun compass orientation of Tylos europaeus (Crustacea, Isopoda) under artificial light. Ethology. 2004;110:581–590. 10.1111/j.1439-0310.2004.00997.x [Google Scholar]
- Ugolini A, Vignali B, Castellini C, Lindstroem M. Zonal orientation and spectral filtering in Talitrus saltator (Amphipoda, Talitridae) J. Mar. Biol. Ass. UK. 1996;76:377–389. [Google Scholar]
- Ugolini A, Tiribilli B, Castellini C. Artificial light and sun compass orientation in the sandhopper Talitrus saltator (Crustacea–Amphipoda) J. Comp. Physiol. A. 1998;182:43–50. 10.1007/s003590050156 [Google Scholar]
- Ugolini A, Melis C, Innocenti R. Moon orientation in adult and young sandhoppers. J. Comp. Physiol. A. 1999a;184:9–12. 10.1007/s003590050301 [Google Scholar]
- Ugolini A, Melis C, Innocenti R, Tiribilli B, Castellini C. Moon and sun compasses in sandhoppers rely on two separate chronometric mechanisms. Proc. R. Soc. B. 1999b;266:749–752. 10.1098/rspb.1999.0700 [Google Scholar]
- Ugolini A, Melis C, Tiribilli B, Castellini C. The sun, the moon, and the sandhopper. In: Wiese K, editor. Crustacean experimental systems in neurobiology. Springer-Verlag; Berlin: 2002a. pp. 247–260. [Google Scholar]
- Ugolini A, Tiribilli B, Boddi V. The sun compass of the sandhopper Talitrus saltator: the speed of the chronometric mechanism depends on the hours of light. J. Exp. Biol. 2002b;205:3225–3230. doi: 10.1242/jeb.205.20.3225. [DOI] [PubMed] [Google Scholar]
- Ugolini A, Fantini T, Innocenti R. Orientation at night: an innate moon compass in sandhoppers (Amphipoda: Talitridae) Proc. R. Soc. B. 2003;270:279–281. doi: 10.1098/rspb.2002.2221. 10.1098/rspb.2002.2221 [DOI] [PMC free article] [PubMed] [Google Scholar]