

Abstract
Climate change has led to shifts in phenology in many species distributed widely across taxonomic groups. It is, however, unclear how we should interpret these shifts without some sort of a yardstick: a measure that will reflect how much a species should be shifting to match the change in its environment caused by climate change. Here, we assume that the shift in the phenology of a species' food abundance is, by a first approximation, an appropriate yardstick. We review the few examples that are available, ranging from birds to marine plankton. In almost all of these examples, the phenology of the focal species shifts either too little (five out of 11) or too much (three out of 11) compared to the yardstick. Thus, many species are becoming mistimed due to climate change. We urge researchers with long-term datasets on phenology to link their data with those that may serve as a yardstick, because documentation of the incidence of climate change-induced mistiming is crucial in assessing the impact of global climate change on the natural world.
Keywords: phenology, climate change, mistiming, food chains
1. Introduction
There is now ample evidence that over the last decades the phenology—the timing of seasonal activities such as timing of flowering or breeding (Walther et al. 2002)—of many plant and animal species has advanced and that these shifts are related to climate change (Hughes 2000; Walther et al. 2002; Parmesan & Yohe 2003; Root et al. 2003; Dunn 2004). What is, however, less clear is how we should interpret these shifts in phenology. The observed changes in phenology may be a positive sign because species are apparently adapting to changing climatic conditions, or they may be a negative sign because they show that climate change is, indeed, impacting living systems (cf. Parmesan & Yohe 2003). Not all species or populations, however, show a shift in phenology (Visser et al. 1998; Visser et al. 2003; Both et al. 2004). Are these the species or populations at risk or are these the lucky ones whose environments are not (yet) affected by climate change? The observed changes in phenology cannot be interpreted without considering the ecological context in which a species lives, and especially how other components of the ecosystem are affected by climate change. What is needed is some sort of a yardstick: a measure of how much a species should shift to match the change in its environment caused by climate change.
Plants and animals exhibit seasonal patterns in their activities because there is a clear seasonality in the suitability of their environment: there is often only a limited period in the year when conditions are favourable enough to successfully reproduce or grow. If reproduction or growth takes place outside this window of favourable conditions, there are often large fitness consequences. Ultimately, the activity that is the most demanding for an organism should take place at the time of optimal conditions. What we need to know to evaluate the observed shifts in phenology is how that period of optimal conditions shifts due to climate change. Thus, we should use the shift in the seasonal changes in the ecological conditions as a yardstick to assess whether the change in phenology observed is sufficient or not.
As these ecological conditions for a species are often set by organisms at other trophic levels it is important to realize that there is no a priori reason why the phenology of different trophic levels will shift at the same rate (Visser et al. 2004): plants are likely to have different mechanisms underlying their phenology than insects, which in turn will have different mechanisms than vertebrates. Although natural selection is expected to have shaped these mechanisms such that the response to temperatures is similar to that of, e.g. food species, this will only be true under the set of abiotic conditions under which these species have evolved, and not for the conditions altered by climate change (see Visser et al. (2004) for a more extended argument).
If the phenology of a species is shifting at a different rate from that of the species that make-up its ecological conditions, this will lead to mistiming of its seasonal activities (Visser et al. 2004) or, to use an alternative terminology, to a mismatch in phenology (Stenseth & Mysterud 2002). Such trophical decoupling of food web phenology may have severe consequences, including biodiversity loss (Visser et al. 2004).
This leaves us with the difficult tasks of, first, assessing the period of optimal conditions for a species and, second, quantifying how much this period has shifted due to climate change and thereby determining whether the response of a species is sufficient. In general, what is needed is identification of the variables that make-up the important selection pressures on the phenology of a species. For many species, the optimal period will be determined by the temporal distribution of food needed during reproduction or growth, as food abundance is often a major determinant of fitness (Martin 1987), and in most of this paper we will assume that the main selection pressure on phenology is food abundance (but see §8).
Given the enormous body of literature reporting on shifts in phenology, it is surprising how rarely the shifts in the period of optimal conditions have also been documented. However, there are now a number of examples, ranging from birds to marine plankton. What is needed, and what we will attempt to do in this review, is to summarize these examples, to draw general conclusions from them and to build a general framework to guide future research.
2. Breeding phenology of birds
Breeding is the most demanding period within the avian life cycle (Martin 1987) and synchrony with food abundance is crucial. One of the food chains with the best studied phenology is the oak (Quercus robur)–winter moth (Opheroptera brumata)–great tit (Parus major) system (Perrins 1970; Visser et al. 1998; Buse et al. 1999), which is typical for a whole group of small insectivorous forest birds. These species mainly feed their nestlings on caterpillars, which are only available during a relatively short period in spring. This poses a clear selection pressure on the timing of reproduction in these species: they have to time their reproduction such that the needs of their offspring match the time of peak abundance of caterpillars. If they lay their eggs earlier or later, they fledge fewer and lighter offspring (Perrins & McCleery 1989; van Noordwijk et al. 1995; Verboven et al. 2001; Visser et al. in press b).
The phenology of caterpillar biomass has been monitored in a few study areas, including Wytham Wood in the UK (Perrins 1991; Cresswell & McCleery 2003) and the Hoge Veluwe in The Netherlands (Visser et al. 1998; Visser et al. in press b). For these populations, the shift in the date of the caterpillar biomass peak can be used as a yardstick for the shifts in timing of reproduction of the birds. The Wytham Wood great tit population has advanced its laying date over the past 39 years, but more so than the shift in caterpillar biomass phenology (Cresswell & McCleery 2003). In contrast, the Hoge Veluwe great tits have not advanced their laying date but the caterpillar peak date has advanced over the past two decades (Visser et al. 1998, in press b) (figure 1c). The Hoge Veluwe pied flycatchers have advanced their laying dates, but not enough to match the shift in caterpillar biomass peak date (Both & Visser 2001). In all cases, the responses of the birds seem to be different from their main food. Especially for the Wytham Wood great tits and the Hoge Veluwe pied flycatchers, where there has been an advance in laying date, it is essential to have a yardstick because otherwise it would be completely unclear whether the observed shifts in laying date have resulted in a mismatch in timing due to a too large (as the Wytham Wood great tits, but see §8) or too small (as the Hoge Veluwe pied flycatchers) response.
Figure 1.
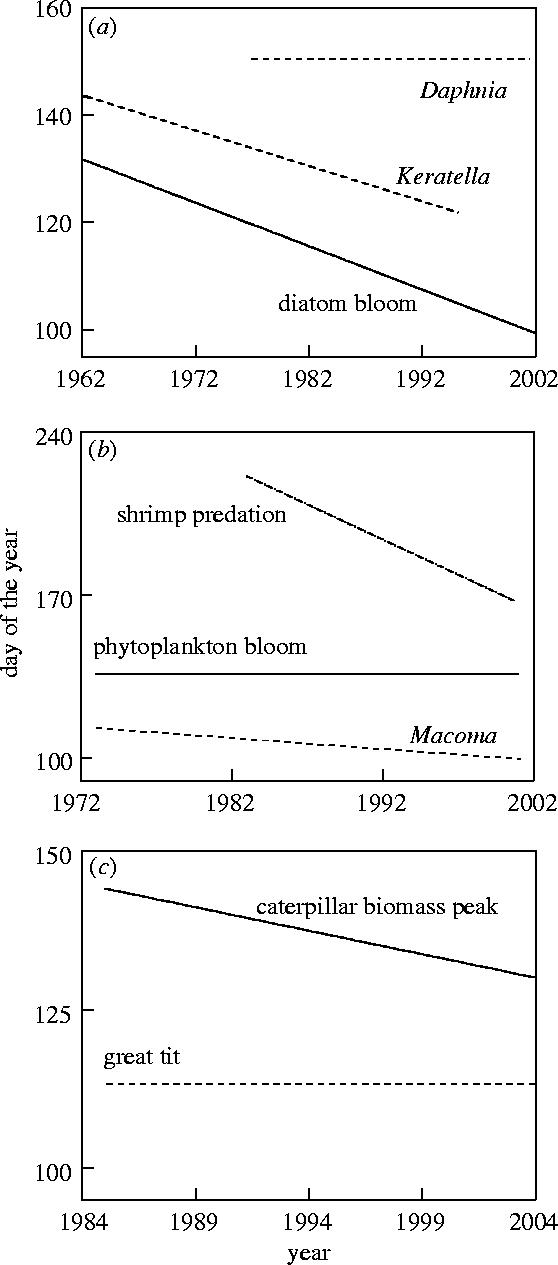
Climate change may lead to different shifts in phenology within food chains. Three of the examples from table 1 are illustrated using the fitted lines of the relationships found. In each example, the solid line is for the species lower down the food chain. (a) diatom bloom–Daphnia peak densities–Keratella peak densities phenology (Winder & Schindler 2004), (b) Macoma spawning–phytoplankton bloom–shrimp predation risk phenology (Philippart et al. 2003), (c) date of caterpillar biomass peak–great tit laying dates phenology (Visser et al. 1998, in press b). Note that in (a) and (b) the time trends for different taxa do not differ significantly when the data are restricted to the period for which data on all three species is available.
In species where the young search for their food themselves, such as golden plovers (Pluvialis apricaria), reproductive success also depends on the synchrony of hatching of their chicks with the phenology of the food, in this case adult tipulids (Tipulidae) (Pearce-Higgins & Yalden 2004). As for the forest caterpillars, tipulids are also abundant for only a 2–3-week period. Unfortunately, no long-term datasets are available to document shifts in the phenology of the birds and their prey. However, the birds' laying dates and tipulid emergence depend on temperatures in different date periods (Pearce-Higgins et al. 2005) and if temperatures in these different periods change at different rates the synchrony between the birds and their food may be distorted. The same may hold for the black grouse (Tetrao tetrix), whose precocious chicks depend critically on abundant insects for their survival during the first two weeks of life (Baines et al. 1996). In Finland, black grouse lekking has advanced over the past four decades, and as a consequence hatching date has shifted as well (G. X. Gilbert, unpublished data). Data on shifts in insect abundance are needed to provide a yardstick for the interpretation of this shift in avian phenology.
Piscivorous birds also experience a clear seasonal peak in the abundance of their prey, which could serve as yardstick. Black-legged kittiwake (Rissa tridactyla) and common guillemot (Uria aalge) laying dates have become later over the past two decades in a North Sea colony, while there has been no change in European shag (Phalacrocorax aristotelis) phenology (Frederiksen et al. 2004). These species all depend to some extent on sandeel (Ammodytes marinus), and the reproductive output of the birds is lower when the sandeels peak earlier (Rindorf et al. 2000). Unfortunately, no data are available on shifts in sandeel phenology, which makes it impossible to compare these with those of the birds. Similarly, the timing of the reproduction of puffins (Fratercula artica) is related to the timing and abundance of herrings (Clupea harengus) migrating north along the Norwegian coast (Durant et al. 2003). Again, no comparison of the shifts in bird and prey phenologies has been made.
3. Avian migration phenology
The phenology of avian migration recorded as departure dates (Lehikoinen et al. 2004), stop-over or passing dates (Huppop & Huppop 2003; Jenni & Kery 2003) and arrival dates (Huin & Sparks 1998, 2000; Sparks 1999; Cotton 2003; Lehikoinen et al. 2004) has also advanced for many, especially short- and medium-distance, migratory bird species (Butler 2003; Lehikoinen et al. 2004; Mills 2005). The timing of the onset of migration and the speed of migration needs to be such that the phenology of food sources are suitable not only at stop-over sites but also at the breeding grounds upon arrival. What makes these migration decisions complicated under climate change is that cues other than temperature, like photoperiod, are often used for the onset of migration (Gwinner 1996) and, even if the birds would use temperature, that climate change is not proceeding at the same rate in the overwintering, staging and breeding areas (Coppack & Both 2002).
Bewick's swans (Cygnus columbianus bewickii) now depart earlier from the Netherlands to their artic breeding areas (J. van Gils, unpublished data), which may be advantageous as the phenology of their stop-over sites in the Baltic area is advancing. In North America, wood warblers (Parulidae) have not advanced their migration phenology, either at a stop-over location or at a location at the southern range of their breeding area (Strode 2003; Marra et al. 2005; Mills 2005). Temperatures north of the stop-over site, however, have significantly increased and as a consequence the phenology of their main prey species, the eastern spruce budworm (Choristoneura fumiferana), has advanced (as deduced from a simple model using temperature). Thus, the wood warblers arrive too late in their breeding grounds to fully profit from the peak in caterpillars, and the same holds for the conditions en route where they need to refuel. In contrast, temperatures south of the stopover site have decreased and, hence, caterpillars there appear later, perhaps hampering an earlier departure date, as birds need to fatten up before starting their migration.
A second example where the advancement of arrival dates of a long distance migrant is insufficient, as deduced using food phenology as a yardstick, is the pied flycatcher (Ficedula hypoleuca) in The Netherlands. While there is a clear advancement in the phenology of their main prey, caterpillars, the birds are not arriving any earlier (Both & Visser 2001). Interestingly, despite this lack of advancement of arrival date, the birds have advanced their laying date. They do this by shortening the interval between arrival and breeding. Currently, this interval has shrunk to only a few days and further advancement of breeding is not possible without an earlier arrival. North American passerines have advanced their arrival date on the breeding grounds, probably as a result of faster spring migration at higher temperatures en route (Marra et al. 2005). However, this response to temperature is about one-third of the advance in flowering of at least one plant species in their breeding areas, and the birds' advance may thus be insufficient. A final example where there is a yardstick available to judge (the lack of) shifts in arrival dates is the honey-buzzard (Pernis apivorus), a bird of prey that is specialized on wasps (Vespula and Dolichovespula spp.). While there is a clear advancement in wasp phenology, there is again no change in Dutch honey-buzzard arrival dates (R. Bijlsma, unpublished data).
An example of an altitudinal, rather than north–south, migrant is the American robin (Turdus migratorius). A good yardstick for the birds' migratory phenology is the date of first bare ground as this correlates tightly with date of first flowering, which in turn determines food availability for the robins. In the Rocky mountains, this bird species is now arriving 14 days earlier than two decades ago, but as there has been no advancement of the date of snow melt, the interval between the first arrival of the Robins and the first date of bare ground has grown by 18 days over this period (Inouye et al. 2000).
4. Insect phenology
Many herbivorous insect species can only develop on young plant material. For these species, the phenology of growth of leaves or growing tips can be used as a yardstick for the shifts in the phenology of their larval development. A clear example of this is the winter moth–oak interaction (Dewar & Watt 1992; Buse & Good 1996; Visser & Holleman 2001). Winter moth eggs which hatch either before or after the oak bud burst have reduced fitness (Feeny 1970; van Dongen et al. 1997) because the first instar will either starve or have to eat older leaves, which contain more tannins, leading to smaller females with a reduced egg load (Buse et al. 1998). Winter moth egg hatch phenology has clearly advanced over the past 15 years in the Netherlands (van Asch, unpublished data; Visser & Holleman 2001). To assess whether this advancement is adaptive we used the oak bud burst phenology as a yardstick. From this, it is clear that the winter moth advancement is too great. Although oak bud burst has also advanced, the winter moth eggs currently hatch well before the bud burst date, leading to mistiming (Visser & Holleman 2001).
An example where climate change might not lead to mistiming is the orange tip butterfly (Anthocharis cardamines) because its mean date of first appearance has a very similar response to March temperatures as the flowering date of garlic mustard (Alliaria petiolata), one of its host plants (Sparks & Yates 1997; Harrington et al. 1999). However, this analysis is based on a period largely prior to climate change (1883–1993) and hence, except in the unlikely event that March temperatures are truly the causal mechanism, climate change may affect insect and plant phenology differently.
Some butterfly species are migratory and hence faced with the same problems as migratory birds. For instance, the red admiral (Vanessa atalanta) has advanced its return date to Britain over the past two decades, while the flowering phenology of one of its host plants the stinging nettle (Urtica dioica), has not advanced (Sparks et al. 2005; T. H. Sparks, unpublished data). As a consequence, the interval between arrival and flowering date has decreased.
5. Phenology in aquatic systems
The timing of spawning in Macoma balthica, a marine intertidal bivalve, is under selection by temporal variation in both its food resources, phytoplankton, and predation risk by juvenile shrimps (Crangon crangon). Over the past two decades, Macoma spawning phenology has advanced by one day per year in the Dutch Waddensea (Philippart et al. 2003) (figure 1b). However, there has been no advancement in the timing of the phytoplankton bloom, leading to a mismatch between Macoma development and resources. The interesting twist is that the timing of the peak in predation risk has advanced by over 3 days per year (Philippart et al. 2003). Because a correct yardstick for Macoma should take into account a resource component that is not shifting and a predation component that is, it is difficult to assess whether or not the shift in Macoma phenology has been sufficient: it is insufficient compared to the shift in predation risk but too strong when compared to the phenology of its food (see §8).
Another marine system where there has been a shift in phenology is the marine pelagic community: diatoms and dinoflagellates (primary producers), copepods (secondary producers), non-copepod holozooplankton and meroplankton (secondary and tertiary producers). Large differences in the extent of the shifts of the phenology of these components have been recorded over the past four decades (Edwards & Richardson 2004): of the producers, diatoms as a group have not shifted (as their phenology depends on photoperiod or light intensity, rather than temperature) but the dinoflagellates have by 23 days. The copepod and non-copepod zooplankton have advanced by 10 days while the largest advancement was observed in the meroplankton (27 days). As a result, there is currently a mismatch between the successive trophic levels and a change in the synchrony in the timing of primary, secondary and tertiary producers (Edwards & Richardson 2004).
There have also been clear phenological shifts in freshwater systems. In Lake Washington, there are two species of zooplankton, Keratella cochlearis and Daphnia pulicaria that feed on phytoplankton (the bloom of the diatom Asterionella formosa). The timing of the diatom bloom has clearly advanced over the past four decades, by 27 days. Keratella's phenology has shifted at roughly the same rate, 21 days, but no shift in Daphnia phenology has been observed (figure 1a, Winder & Schindler 2004). Thus, when using the phenology of the diatom bloom as a yardstick, it is clear that the zooplankton species that is shifting (Keratella) does this at a sufficient rate, while the one that is not shifting (Daphnia) is becoming mistimed with its food resources.
6. Synthesis
From the literature reviewed it is clear that in the majority of cases where a yardstick is available the advances in phenology have been either too much (three out of 11) or too little (five out of 11) compared to the organisms on which the species depend (table 1). This fits in with the general finding that there are different rates of change in the phenology of plants, insects, vertebrates (Parmesan & Yohe 2003; Voigt et al. 2003), leading to mistiming or a mismatch. This result stresses the importance of a yardstick: it would have otherwise been completely unclear which of these species was changing at a sufficient rate and which was not.
Table 1.
Review on studies on shifts in phenology where a yardstick to assess whether these shifts are sufficient can be identified.
| species | phenological event | yardstick | response | reference |
|---|---|---|---|---|
| D. pulicaria (fresh water zooplankton) | abundance | diatom (A. formosa) bloom | too little | Winder & Schindler (2004) |
| K. cochlearis (fresh water zooplankton) | abundance | diatom (A. formosa) bloom | sufficient | Winder & Schindler (2004) |
| M. balthica (a marine intertidal bivalve) | spawning | phytoplankton bloom and settlement of juvenile shrimps (Cragon cragon) | unclear | Philippart et al. (2003) |
| winter moth (O. brumata) | egg hatching | oak (Q. robur) bud burst | too much | Visser & Holleman (2001) |
| orange tip butterfly (A. cardamines) | first appearance | garlic mustard (A. petiolata) flowering date | sufficient (?) | Sparks & Yates (1997), Harrington et al. (1999) |
| red admiral (V. atalanta) | migration–return date | stinging nettle (U. dioica) flowering | too much | Sparks et al. (2005) |
| great tit (P. major) | laying date | caterpillar biomass peak | too little | Visser et al. (1998), Visser et al. (in press b) |
| great tit (P. major) | laying date | caterpillar biomass peak | sufficient | Cresswell & McCleery (2003) |
| golden plover (P. apricaria) | laying date | insect abundance peak | unknown | Pearce-Higgins et al. (2005) |
| black-legged kittiwake (R. tridactyla), common guillemot (U. aalge), European shag (P. aristotelis) | laying date | sandeel (Ammadytes marinus) abundance peak | unknown | Rindorf et al. (2000), Frederiksen et al. (2004) |
| puffin (F. artica) | laying date | herring (C. harengus) abundance peak | unknown | Durant et al. (2003) |
| wood warblers (Parulidae) | migration | eastern spruce budworm (C. fumiferana) density peak | too little | Strode (2003) |
| pied flycatcher (F. hypoleuca) | migration–arrival date | caterpillar biomass peak | too little | Both & Visser (2001) |
| honey-buzzard (P. apivorus) | migration–arrival date | wasp (Vespula sp.) abundance peak | too little | R. Bijlsma (unpublished data) |
| American robin (T. migratorius) | migration–arrival date | snow melt | too much | Inouye et al. (2000) |
Responses are classified as either too strong (the shift is larger than the shift in the yardstick), sufficient or too weak. In some cases, the shift in the yardstick is not reported and these are classified as unknown. In the Macoma example, it is unclear whether the shift is sufficient, see text. For an example on a marine pelagic community (Edwards & Richardson 2004), see text.
At first sight it seems surprising that different parts of a food chain respond differently to variations in annual temperatures. There has always been temperature variation and hence selection on higher trophic levels to keep in line with the annual variation in the phenology of their food source. As a result, many life-history characters are phenotypically plastic: under different conditions the phenotypic expression of, for instance, flowering time differs. It is, however, important to realize that climate may not change uniformly over the season, and hence species which differ in the periods that causally affect their phenology may diverge in timing, with possibly detrimental effects for the higher trophic level and, perhaps, beneficial effects for the lower trophic level.
Species at different trophic levels often differ in the temporal and spatial scale at which decisions are made regarding timing. For at least some vertebrate species reproduction has to be initiated long before their young need provisioning. For instance, in great tits there is a four-week delay before the start of laying and the peak in food demands on the nestlings. If temperatures increase after the birds have started laying, caterpillar development is accelerated but the birds have very limited ability at this stage to advance the hatching of their eggs to match this (see van Noordwijk et al. 1995; Visser et al. 2004, for an elaborated argument). Long distance migrants not only face a temporal problem, but also the spatial problem of being thousands of kilometres away from their breeding site during winter. These birds may not use climatic variables to time their migration and as a consequence the birds do not advance their migration while the food sources at their breeding sites do advance (Coppack & Both 2002). This north–south geographical variation is similar to the variation in climate change at lower and higher elevations, which is responsible for mistiming in the American robin (Inouye et al. 2000).
An additional problem for organisms posed by climate change is that some parts of the food chain use temperature unrelated cues to time their annual cycle, for instance marine diatoms (Edwards & Richardson 2004), Daphnia (Winder & Schindler 2004) and long distance migrants (Gwinner 1996). Moreover, while insects and plants are often temperature sensitive, vertebrates also use photoperiod, a cue that is not affected by climate change. But even in these cases, we would expect natural selection to shape the plasticity of these species in such a way that they shift their phenology in accordance with that of their environment. The fundamental problem is that this plasticity does not work when outside the natural frequency distribution of annual environmental conditions. While a species might be mistimed in the odd warm year in the past, now the warm years are common, and thus what is even more threatening than a general increase in temperature is that the climatic patterns have become disrupted (cf. Visser et al. 2004).
7. Predicting future shifts in phenology
Given that our review shows that most species react differently to climate change from the species on which they depend (see table 1), the outstanding question is how future climate change will affect the phenology of whole ecosystems under different climate change scenarios (Houghton et al. 2001). One of the main challenges is to determine to what extent the reported correlations between phenology and temperatures (usually the mean temperature over a fixed date period) actually reflect the underlying causal mechanisms. This is crucial because extrapolations using climate scenarios now sometimes predict that phenological events will occur outside the date period over which temperatures are used to predict phenology. In this respect, linking studies on the physiological mechanisms underlying phenology is essential for predicting future shifts.
Recently, a method has been developed to describe phenology in a more sophisticated way than just correlating annual phenology to mean temperatures over a fixed period (Gienapp et al. 2005). In this so-called proportional hazard model, developed for survival statistics, the probability of laying is calculated per day depending on the current and previous conditions. This model, albeit still a correlational model, avoids the problems with fixed periods and also allows for interactions, such as that between photoperiod and temperature, to be incorporated. The model has been used to predict great tit laying dates up to 2100 (Visser et al. in press b).
So far we have considered just a single life-history stage, but species have to perform different tasks at different times, all of which may be affected by climate related ecological factors. This is most pronounced in the annual cycle of long distant migrants, where the birds have to decide when to leave their wintering site, how fast to travel, how much energy in the form of body reserves should be brought to the breeding site, and when to start breeding after arrival. Especially for these species it is important to take the entire life cycle into account when predicting future shift in phenology. One way to model this is to use stochastic dynamic programming (McNamara & Houston 1996). This has been applied to the migration of pink-footed geese (Anser brachyrhynchus) from The Netherlands to the Artic by Bauer et al. (submitted), who showed that the onset of spring on the stop-over sites affects the optimal departure times, stressing the need to take the whole migratory journey into account.
Predicting future phenology is not often done, but even more rarely does it take into account evolutionary changes in phenotypic plasticity. In the oak–winter moth system, the predicted phenology of oak bud burst and winter moth egg hatching is getting more and more out of synchrony. However, if an evolutionary change in plasticity in response to selection imposed by mistiming (based on estimated heritabilities of plasticity and fitness consequences of mistiming) is taken into account, synchrony is predicted to be restored within two decades (van Asch et al. submitted).
8. Discussion
Climate change has affected the phenology of a wide range of species but it remains difficult to interpret these shifts and to explain the variation among species and even among populations within species. We argue the need for a yardstick—some measure of how much a species should shift given the changes in its environment—to assess whether shifts in phenology are sufficient. We have brought together all examples we could find (table 1) and have shown that, in the majority of cases, the observed shifts do not seem to match the shifts that would be expected. It remains unclear how we should interpret the thousands of reported shifts in phenology for which we have no yardstick. We would like to encourage researchers to establish links with other researchers or institutions that work on other components of the food chain of their focal species. In many cases, data are available but are often collected by groups working in a different setting, for instance fisheries research may well have data on the phenology of fish needed to interpret shifts in phenology of piscivorous birds. Integration and linkages of long term databases of plant, insect and vertebrate species is both crucial and a major challenge.
In this review, we have considered a very simple form of yardstick: in most of our examples we have assumed that the selection acting on the phenology of a species comes from just a single selection agent: their food source. Furthermore, we have assumed that there is just a single activity (i.e. breeding, spawning) a year for which timing is important. Although we admit that this ‘single critical activity–single selection agent’ scenario is highly unlikely to be the case, we fear that it is the best possible yardstick at this moment. And even so, most examples we reviewed come from simple ecological situations, because more complex trophic interactions are more difficult to study, and long-time series on all relevant species within a food web are rarer than these simple food chains.
When we go beyond measuring the shifts in phenology of a single selection agent it becomes more difficult to define a yardstick, as we then need to integrate these different selection pressures. The one example where there are data on both food and predator phenology (the Macoma example, Philippart et al. 2003) there was no change in food phenology but a strong shift in predator risk phenology, and it is unclear how these two selection pressures should be combined into a single yardstick. Another example where the phenology of predation may be an equally important selection pressure as the phenology of food is the frog (Rana temporaria), and its predators (newts, Triturus sp.). The newts have advanced their entry in ponds, whereas frogs have not substantially altered their reproductive phenology (Beebee 1995). Therefore, embryos and larvae of early breeding frogs are now exposed to higher levels of newt predation (Walther et al. 2002).
Ultimately, the way forwards is to measure selection on phenology and to assess whether there is increased directional selection (Visser et al. in press a). For this however, long-term studies where individual fitness can be calculated are needed rather than just population means. The very few studies that have calculated whether there is increased directional selection are all on birds (Visser et al. 1998; Both & Visser 2001; Cresswell & McCleery 2003) as only for this group such long-term studies are available. Clearly, individual fitness measures will not be available for the Macoma system. But also for such systems, the ultimate way to assess whether their shift in phenology is sufficient is to measure the reproductive success of early and late spawning individuals. Even if we are able to measure the selection acting on the phenology, a yardstick is still essential as the changes in these fitness-based measures may also be caused by other changes in the environment. A full understanding of whether species are reacting sufficiently to the climate change-induced advance of its food requires therefore both these fitness estimates and the yardstick. In the case of great tits, the change in selection differential over the years is consistent with the improved (Cresswell & McCleery 2003) or the deteriorated synchrony (Visser et al. 1998) between food and reproduction.
The second critical assumption of our review is that there is just a single critical activity per year that is under selection. In reality, this will not be the case and a life cycle approach is more appropriate: the entire life cycle has to be fitted into the seasonal changes in suitability of the habitat. This is even more important as different life history stages may each be affected by climate change, as we have already discussed for migrant birds (Winkler et al. 2002; Both & Visser 2005). Such changes of multiple life-history traits have also been reported for resident bird species. The Hoge Veluwe great tits, which have not shifted their laying dates, have responded to climate change by no longer producing second clutches (Visser et al. 2003). In the case of the Wytham Wood great tits, they have prolonged the time between the laying of their last egg and the hatching of their chicks. As a result the interval between hatch date and caterpillar peak has not changed over the years and hatching asynchrony has been reduced (Cresswell & McCleery 2003). It is, therefore, very well possible that the shift in laying date in this population has been sufficient despite the fact that the shift in the phenology of their food, the peak caterpillar biomass date, was weaker than the shift in laying date, and hence we have classified this study as ‘sufficient’ rather than to ‘too much’ in table 1. As for the ‘single selection agent’ assumption, also the ‘single critical activity’ assumption can only be lifted for a handful of studies, again mainly birds.
Finally, there are a few more complications with the assumption that shifts in food phenology is a useful yardstick. It may also be that more generalist species are less affected by climate change, because they can more easily switch to alternative prey if they are out of synchrony with one of their prey species. Another complication is that so far we have just considered changes in the timing of the optimal period, e.g. food peak, but also the width of the optimal window may change as climate changes. These changes have been only rarely mentioned in the literature (Buse et al. 1999) and will make the use of a yardstick more difficult. However, changes in the width of the optimal period may be even more important than the actual date of the peak, because if the window becomes too narrow reproduction may become impossible, whatever the change in phenology.
Our review suggests that an insufficient response to climate change is the rule rather than the exception, and that only in a few cases has the consumer shifted its phenology to the same extent as its food. We, however, want to stress that it may well be possible that insufficient responses are published more frequently than cases where species have adjusted smoothly to the present climate change, i.e. there may be publication bias toward reports of mistiming. We urge researchers with long-tem datasets on phenology to link their data with those that can serve as a yardstick. Despite the complications discussed above, we believe that making a comparison of actual shifts with predicted shifts will be an important step forwards, even if the yardstick is not perfect, as it is crucial to assess the impact of climate change on the natural world. If indeed most species are becoming mistimed this will emphasize the need to take measure to reduce climate change because mistiming is likely to have detrimental effects on species persistence, and thereby on biodiversity.
Acknowledgements
We would like to thank Charles Godfray for inviting us to write this review, Tim Sparks, Margriet van Asch, Rob Bijlsma, Jan van Gils and Silke Bauer for allowing us to cite their unpublished work and Kate Lessells, Phillip Gienapp and Will Cresswell for their comments on the manuscript.
References
- Baines D, Wilson I.A, Beeley G. Timing of breeding in black grouse Tetrao tetrix and capercaillie Tetrao urogallus and distribution of insect food for the chicks. Ibis. 1996;138:181–187. [Google Scholar]
- Bauer, S., Van Dinther, M., Klaassen, M. & Madsen, J. Submitted. Climate change and Arctic breeding geese—past, present and potential future migration schedules. Global Change Biol.
- Beebee T.J.C. Amphibian breeding and climate. Nature. 1995;374:219–220. doi:10.1038/374219a0 [Google Scholar]
- Both C, Visser M.E. Adjustment to climate change is constrained by arrival date in a long-distance migrant bird. Nature. 2001;411:296–298. doi: 10.1038/35077063. doi:10.1038/35077063 [DOI] [PubMed] [Google Scholar]
- Both C, Visser M.E. The effect of climate change on the correlation between avian life history traits. Global Change Biol. 2005;11:1606–1613. [Google Scholar]
- Both C, et al. Large-scale geographical variation confirms that climate change causes birds to lay earlier. Proc. R. Soc. B. 2004;271:1657–1662. doi: 10.1098/rspb.2004.2770. doi:10.1098/rspb.2004.2770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buse A, Good J.E.G. Synchronization of larval emergence in winter moth (Operophtera brumata L.) and budburst in pedunculate oak (Quercus robur L.) under simulated climate change. Ecol. Entomol. 1996;21:335–343. doi:10.1046/j.1365-2311.1996.t01-1-00001.x [Google Scholar]
- Buse A, Good J.E.G, Dury S, Perrins C.M. Effects of elevated temperature and carbon dioxide on nutritional quality of leaves of oak (Quercus robur L.) as food for the winter moth (Operophtera brumata L.) Funct. Ecol. 1998;12:742–749. doi:10.1046/j.1365-2435.1998.00243.x [Google Scholar]
- Buse A, Dury S.J, Woodburn R.J.W, Perrins C.M, Good J.E.G. Effects of elevated temperature on multi-species interactions: the case of Pendunculate Oak, Winter Moth and Tits. Funct. Ecol. 1999;13:74–82. doi:10.1046/j.1365-2435.1999.00010.x [Google Scholar]
- Butler C.J. The disproportionate effect of global warming on the arrival dates of short-distance migratory birds in North America. Ibis. 2003;145:484–495. doi:10.1046/j.1474-919X.2003.00193.x [Google Scholar]
- Coppack T, Both C. Predicting life-cycle adaptation of migratory birds to global climate change. Ardea. 2002;90:369–378. [Google Scholar]
- Cotton P.A. Avian migration phenology and global climate change. Proc. Natl Acad. Sci. USA. 2003;100:12219–12222. doi: 10.1073/pnas.1930548100. doi:10.1073/pnas.1930548100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cresswell W, McCleery R. How great tits maintain synchronization of their hatch date with food supply in response to long-term variability in temperature. J. Anim. Ecol. 2003;72:356–366. doi:10.1046/j.1365-2656.2003.00701.x [Google Scholar]
- Dewar R.C, Watt A.D. Predicted changes in the synchrony of larval emergence and budburst under climatic warming. Oecologia. 1992;89:557–559. doi: 10.1007/BF00317163. [DOI] [PubMed] [Google Scholar]
- Dunn P. Breeding dates and reproductive performance. Adv. Ecol. Res. 2004;35:69–87. [Google Scholar]
- Durant J.M, Anker-Nilssen T, Stenseth N.C. Trophic interactions under climate fluctuations: the Atlantic puffin as an example. Proc. R. Soc. B. 2003;270:1461–1466. doi: 10.1098/rspb.2003.2397. doi:10.1098/rspb.2003.2397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards M, Richardson A.J. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature. 2004;430:881–884. doi: 10.1038/nature02808. doi:10.1038/nature02808 [DOI] [PubMed] [Google Scholar]
- Feeny P. Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter moth caterpillars. Ecology. 1970;51:565–581. [Google Scholar]
- Frederiksen M, Harris M.P, Daunt F, Rothery P, Wanless S. Scale-dependent climate signals drive breeding phenology of three seabird species. Global Change Biol. 2004;10:1214–1221. doi:10.1111/j.1529-8817.2003.00794.x [Google Scholar]
- Gienapp P, Hemerik L, Visser M.E. A new statistical tool to predict phenology under climate change scenarios. Global Change Biol. 2005;11:600–606. doi:10.1111/j.1365-2486.2005.00925.x [Google Scholar]
- Gwinner E. Circannual clocks in avian reproduction and migration. Ibis. 1996;138:47–63. [Google Scholar]
- Harrington R, Woiwod I, Sparks T. Climate change and trophic interactions. Trends Ecol. Evol. 1999;14:146–150. doi: 10.1016/s0169-5347(99)01604-3. doi:10.1016/S0169-5347(99)01604-3 [DOI] [PubMed] [Google Scholar]
- Houghton J.T, Ding Y, Griggs D.J, Noguer M, van der Linden P.J, Xiaosu D. Cambridge University Press; Cambridge, MA: 2001. Climate Change 2001: the scientific basis contribution of working group I to the third assessment report of the intergovernmental panel on climate change (IPCC) [Google Scholar]
- Hughes L. Biological consequences of global warming: is the signal already apparent? Trends Ecol. Evol. 2000;15:56–61. doi: 10.1016/s0169-5347(99)01764-4. doi:10.1016/S0169-5347(99)01764-4 [DOI] [PubMed] [Google Scholar]
- Huin N, Sparks T.H. Arrival and progression of the Swallow Hirundo rustica through Britain. Bird Study. 1998;45:361–370. [Google Scholar]
- Huin N, Sparks T.H. Spring arrival patterns of the Cuckoo Cuculus canorus, Nightingale Luscinia megarhynchos and Spotted Flycatcher Musciapa striata in Britain. Bird Study. 2000;47:22–31. [Google Scholar]
- Huppop O, Huppop K. North Atlantic Oscillation and timing of spring migration in birds. Proc. R. Soc. B. 2003;270:233–240. doi: 10.1098/rspb.2002.2236. doi:10.1098/rspb.2002.2236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inouye D.W, Barr B, Armitage K.B, Inouye B.D. Climate change is affecting altitudinal migrants and hibernating species. Proc. Natl Acad. Sci. USA. 2000;97:1630–1633. doi: 10.1073/pnas.97.4.1630. doi:10.1073/pnas.97.4.1630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenni L, Kery M. Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants. Proc. R. Soc. B. 2003;270:1467–1471. doi: 10.1098/rspb.2003.2394. doi:10.1098/rspb.2003.2394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehikoinen E, Sparks T.H, Zalakevicius M. Arrival and departure dates. Birds Climate Change. 2004;35:1–31. [Google Scholar]
- Marra P.P, Francis C.M, Mulvihill R.S, Moore F.R. The influence of climate on the timing and rate of spring bird migration. Oecologia. 2005;142:307–315. doi: 10.1007/s00442-004-1725-x. doi:10.1007/s00442-004-1725-x [DOI] [PubMed] [Google Scholar]
- Martin T.E. Food as a limit on breeding birds: a life history perspective. Ann. Rev. Ecol. Syst. 1987;18:453–487. doi:10.1146/annurev.es.18.110187.002321 [Google Scholar]
- McNamara J.M, Houston A.I. State-dependent life histories. Nature. 1996;380:215–221. doi: 10.1038/380215a0. doi:10.1038/380215a0 [DOI] [PubMed] [Google Scholar]
- Mills A.M. Changes in the timing of spring and autumn migration in North American migrant passerines during a period of global warming. Ibis. 2005;147:259–269. doi:10.1111/j.1474-919X.2005.00380.x [Google Scholar]
- Parmesan C, Yohe G. A globally coherent fingerprint of climate change impacts across natural systems. Nature. 2003;421:37–42. doi: 10.1038/nature01286. doi:10.1038/nature01286 [DOI] [PubMed] [Google Scholar]
- Pearce-Higgins J.W, Yalden D.W. Habitat selection, diet, arthropod availability and growth of a moorland wader: the ecology of European Golden Plover Pluvialis apricaria chicks. Ibis. 2004;146:335–346. doi:10.1111/j.1474-919X.2004.00278.x [Google Scholar]
- Pearce-Higgins J.W, Yalden D.W, Whittingham M.J. Warmer springs advance the breeding phenology of golden plovers Pluvialis apricaria and their prey (Tipulidae) Oecologia. 2005;143:470–476. doi: 10.1007/s00442-004-1820-z. doi:10.1007/s00442-004-1820-z [DOI] [PubMed] [Google Scholar]
- Perrins C.M. The timing of birds' breeding season. Ibis. 1970;112:242–255. [Google Scholar]
- Perrins C.M. Tits and their caterpillar food supply. Ibis. 1991;133:49–54. [Google Scholar]
- Perrins C.M, McCleery R.H. Laying dates and clutch size in the great tit. Wilson Bull. 1989;101:236–253. [Google Scholar]
- Philippart C.J.M, van Aken H.M, Beukema J.J, Bos O.G, Cadee G.C, Dekker R. Climate-related changes in recruitment of the bivalve Macoma balthica. Limnol. Oceanogr. 2003;48:2171–2185. [Google Scholar]
- Rindorf A, Wanless S, Harris M.P. Effects of changes in sandeel availability on the reproductive output of seabirds. Marine Ecol. Progr. Ser. 2000;202:241–252. [Google Scholar]
- Root T.L, Price J.T, Hall K.R, Schneider S.H, Rosenzweig C, Pounds J.A. Fingerprints of global warming on wild animals and plants. Nature. 2003;421:57–60. doi: 10.1038/nature01333. doi:10.1038/nature01333 [DOI] [PubMed] [Google Scholar]
- Sparks T.H. Phenology and the changing pattern of bird migration in Britain. Int. J. Biometeorol. 1999;42:134–138. doi:10.1007/s004840050096 [Google Scholar]
- Sparks T.H, Yates T.J. The effect of spring temperature on the appearance dates of British butterflies 1883–1993. Ecography. 1997;20:368–374. [Google Scholar]
- Sparks T.H, Roy D.B, Dennis R.L.H. The influence of temperature on migration of Lepidoptera into Britain. Global Change Biol. 2005;11:507–514. doi:10.1111/j.1365-2486.2004.00887.x [Google Scholar]
- Stenseth N.C, Mysterud A. Climate, changing phenology, and other life history and traits: nonlinearity and match-mismatch to the environment. Proc. Natl Acad. Sci. USA. 2002;99:13379–13381. doi: 10.1073/pnas.212519399. doi:10.1073/pnas.212519399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strode P.K. Implications of climate change for North American wood warblers (Parulidae) Global Change Biol. 2003;9:1137–1144. doi:10.1046/j.1365-2486.2003.00664.x [Google Scholar]
- van Asch, M., van Tienderen, P. H., Holleman, L. J. M. & Visser, M. E. Submitted. Predicting shifts in phenology in response to climate change, an insect herbivore example. Global Change Biol
- van Dongen S, Backeljau T, Matthysen E, Dhondt A.A. Synchronization of hatching date with budburst of individual host trees (Quercus robur) in the Winter moth (Operophtera brumata) and its fitness consequences. J. Anim. Ecol. 1997;66:113–121. [Google Scholar]
- van Noordwijk A.J, McCleery R.H, Perrins C.M. Selection of timing of great tit (Parus major) breeding in relation to caterpillar growth and temperature. J. Anim. Ecol. 1995;64:451–458. [Google Scholar]
- Verboven N, Tinbergen J.M, Verhulst S. Food, reproductive success and multiple breeding in the Great Tit Parus major. Ardea. 2001;89:387–406. [Google Scholar]
- Visser M.E, Holleman L.J.M. Warmer springs disrupt the synchrony of oak and winter moth phenology. Proc. R. Soc. B. 2001;268:289–294. doi: 10.1098/rspb.2000.1363. doi:10.1098/rspb.2001.1661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visser M.E, van Noordwijk A.J, Tinbergen J.M, Lessells C.M. Warmer springs lead to mistimed reproduction in great tits (Parus major) Proc. R. Soc. B. 1998;265:1867–1870. doi:10.1098/rspb.1998.0514 [Google Scholar]
- Visser M.E, et al. Variable responses to large-scale climate change in European Parus populations. Proc. R. Soc. B. 2003;270:367–372. doi: 10.1098/rspb.2002.2244. doi:10.1098/rspb.2002.2244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visser M.E, Both C, Lambrechts M.M. Global climate change leads to mistimed avian reproduction. Adv. Ecol. Res. 2004;35:89–110. [Google Scholar]
- Visser, M. E., Both, C. & Gienapp, P. In press a Mistimed reproduction due to global climate change. Acta Zool. Sinica
- Visser, M. E., Holleman, L. J. M. & Gienapp, P. In press b Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird. Oecologia [DOI] [PubMed]
- Voigt W, et al. Trophic levels are differentially sensitive to climate. Ecology. 2003;84:2444–2453. [Google Scholar]
- Walther G.R, Post E, Convey P, Menzel A, Parmesan C, Beebee T.J.C, Fromentin J.M, Hoegh-Guldberg O, Bairlein F. Ecological responses to recent climate change. Nature. 2002;416:389–395. doi: 10.1038/416389a. doi:10.1038/416389a [DOI] [PubMed] [Google Scholar]
- Winder M, Schindler D.E. Climate change uncouples trophic interactions in an aquatic ecosystem. Ecology. 2004;85:2100–2106. [Google Scholar]
- Winkler D.W, Dunn P.O, McCulloch C.E. Predicting the effects of climate change on avian life-history traits. Proc. Natl Acad. Sci. USA. 2002;99:13 595–13 599. doi: 10.1073/pnas.212251999. doi:10.1073/pnas.212251999 [DOI] [PMC free article] [PubMed] [Google Scholar]