

Abstract
As textbook knowledge has it, bees and ants use polarized skylight as a backup cue whenever the main compass cue, the sun, is obscured by clouds. Here we show, by employing a unique experimental paradigm, that the celestial compass system of desert ants, Cataglyphis, relies predominantly on polarized skylight. If ants experience only parts of the polarization pattern during training but the full pattern in a subsequent test situation, they systematically deviate from their true homeward courses, with the systematics depending on what parts of the skylight patterns have been presented during training. This “signature” of the polarization compass remains unaltered, even if the ants can simultaneously experience the sun, which, if presented alone, enables the ants to select their true homeward courses. Information provided by direct sunlight and polarized skylight is picked up by different parts of the ant’s compound eyes and is channeled into two rather separate systems of navigation.
Keywords: insect vision, polarization compass, sun compass, Cataglyphis
By performing his famous mirror experiment in 1911, Santschi (1) was the first to show that animals (ants, in his case) could use the sun as a compass cue. Moreover, he observed that the apparent position of the sun did not always deflect the ants from their homeward courses. In species of some genera of ants (e.g., in Messor and Monomorium), the mirror experiment worked almost always, but in others (e.g., in Cataglyphis), it worked only under certain conditions (for instance, if the remainder of the sky was experimentally obscured). Santschi (2) concluded from these experiments that at least some species of ants had to be able to derive compass information from some optical property of the sun-free sky. We now know that this property is the polarization of scattered skylight (ref. 3; for reviews, see refs. 4 and 5).
A substantial amount of research has been done on the behavioral neurobiology of the insect’s polarization compass (6, 7), but very few data are available about the relative significance and differential processing of compass information derived from either polarized skylight or direct sunlight. This question was addressed by Frisch (8) in his early work on honey bees, but the results he obtained from bees, which performed their recruitment dances with mirrored positions of the sun, remained ambiguous. Because more recent work tells us that (i) bees and ants process information about direct sunlight and polarized skylight by employing different sensory channels [the latter by using the dorsal rim area (DRA) of the eye (9–14)] and (ii) under particular experimental conditions, bees and ants exhibit systematic navigational errors if presented with restricted parts of the polarization patterns in the sky (10, 13, 15, 16), unique experimental paradigms can be designed that allow us to approach the problem of the relative importance of the two compass cues more directly and in more detail. In the present account, we let desert ants, Cataglyphis fortis, experience different stimulus conditions (i.e., exposure to sunlight, skylight, or both) during either training or testing. We show that, in these long-distance desert navigators, the polarization compass dominates over the sun compass and that the sun compass and the polarization compass form two separate systems of navigation.
Results
In all experiments described in the present account, the ants were trained within narrow channels. These channels restricted their skylight vision to a 90°-wide, strip-like window (“partial sky”). Subsequently, however, the ants were transferred to the open test field, where they experienced “full-sky” conditions. What varied among the four principal experimental paradigms that are described below and compiled in Table 1 was the particular stimulus condition with which the ants were presented in the training and test situation.
Table 1.
Experimental paradigms
| Experimental paradigm | Training (inside the channel) | Test (in the open) | Fig. no. |
|---|---|---|---|
| I | SUN plus SKY | SUN plus SKY | 2 |
| II/1 | SUN | SUN plus SKY | 3 |
| II/2 | SUN | SUN | 3 |
| III | SUN plus SKY | SKY | 4 |
| IV | SUN plus SKY | 1 and 2, SUN | 5 and 6 |
In experimental paradigm IV, tests 1 and 2 differed in the way that the ant’s polarization compass was excluded experimentally. In IV/1, the compass was excluded by occluding the dorsal rim areas of both eyes (and the ocelli); in IV/2, the compass was excluded by optically cutting out the UV range of the spectrum. SUN, direct sunlight; SKY, polarized skylight.
In experimental paradigm I, the ants had access to direct sunlight and polarized skylight in both the training and the test situation. Depending on how the long axis of the training channel was oriented relative to the symmetry plane of the polarization pattern [i.e., relative to the solar and antisolar meridian (Fig. 1)], systematic errors occurred whenever the ants trained within the channel had later to perform their homebound runs under the full sky (Fig. 2). The ants chose the correct home direction only if the symmetry plane of the polarization pattern ran either parallel (α = 0°) or at right angles (α = 90°) to the long axis of the training channel (see open arrowheads in Fig. 2). In all other conditions, they deviated either to the right or to the left from their true homeward courses. Experimentally, the angle α was varied (i) by aligning the training channel in different directions (see double-arrowed lines in Fig. 1) and (ii) by training and testing the ants at different times of day (see red arc in Fig. 1). As, due to the westward movement of the sun during the course of the day, the symmetry plane of the polarization pattern rotates about the zenith (with nonuniform speed), ants running back and forth within one particularly oriented (say, north–south) channel would experience different parts of the skylight polarization pattern at different times of day. Hence, during the course of the day, each orientation of the channel yields a particular “error function” (Fig. 2). Moreover, the results of experiment I clearly show that the sun compass does not dominate the polarization compass. If the sun compass did dominate, no systematic errors would be expected.
Fig. 1.
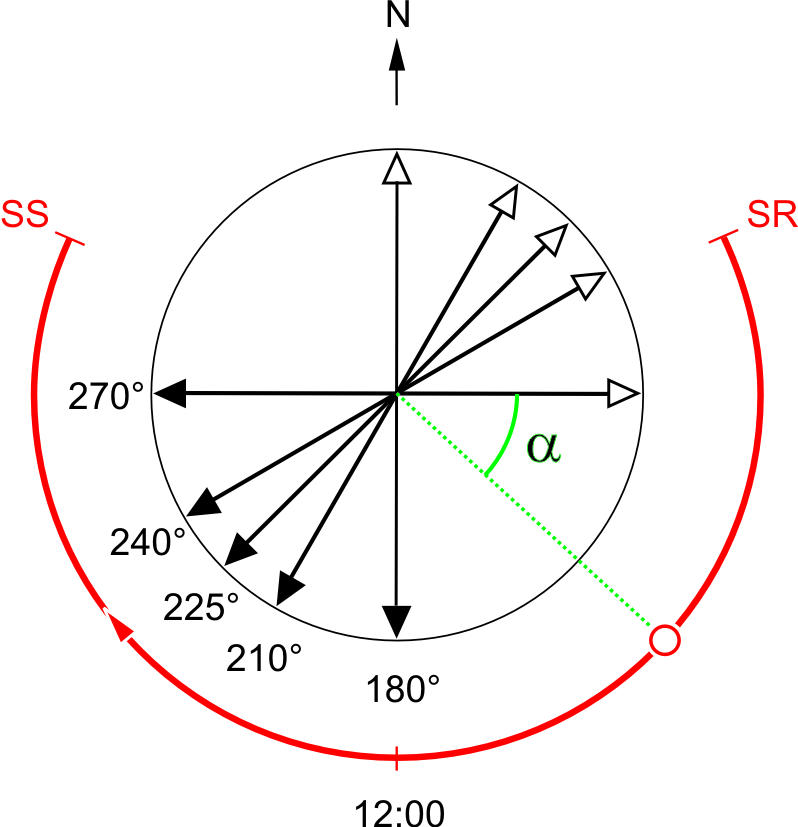
Experimental setup: orientation of training channels within an earthbound system of reference. The double-arrowed lines depict the orientation of the training channels in Fig. 2 (open arrowheads, outbound courses and nest-to-feeder directions; filled arrowheads, inbound courses and feeder-to-nest directions). The red arc indicates the azimuthal position of the sun (the solar meridian) during the course of the day. 12:00 is solar noon. The angle α defines the angular difference between the solar azimuth and the orientation of the training channel (nest-to-feeder direction), depicted here for one particular azimuthal position of the sun and one particular orientation of the training channel. The azimuthal positions of sunrise (SR) and sunset (SS) are given for the summer solstice, June 21. The experiments described in this account were performed during the time period from June 19 to August 5.
Fig. 2.

Experimental paradigm I. Shown are the directions of homing courses, ϕ, taken by ants during the course of the day under full-sky conditions after the ants had been trained under partial-sky conditions within linear channels oriented in five different directions. Means and standard deviations are given in 30-min bins (n = 731). Bold numbers and dotted lines mark the true homeward directions taken by the ants within the training channels (see filled arrowheads in Fig. 1). The open arrowheads at the top of the graphs indicate training situations with α = 0° or 90° (for definition of α, see the legend of Fig. 1). The orange curves show computed polarization-induced error functions (see Methods).
The observed navigational errors are due to the peculiarities of the ant’s polarization compass. As shown first in honey bees (10, 15, 16) and later in desert ants (13), the polarization compass of these insects induces systematic navigational errors whenever the animals are trained and tested under different skylight conditions (e.g., if they are trained under the full pattern of polarized skylight but are later presented only with isolated pixels of that pattern). Based on these studies, theoretical error functions can be computed for all experimental setups used in the present account (see orange curves in Fig. 2; see also Methods). However, irrespective of the validity of these theoretical reconstructions, the point to be made here is that such errors occur and that they occur in a systematic way. Therefore, the systematic time courses of these errors can be regarded as a “signature” of the ant’s polarization compass and thus are indicative of the ant’s use of a polarization compass rather than a sun compass.
We used experimental paradigm II to test whether the latter statement holds true. During training within the channel, the ants had to refrain from using the polarization compass either because the channel was covered by translucent, grease-proof paper that depolarized skylight (paradigm II/1) or because the ants’ DRAs as well as their ocelli had been occluded by light-tight paint (paradigm II/2). Under both conditions, the ants, which during training had experienced only the sun, later selected courses that did not significantly deviate from their true homeward courses (Stevens test, P > 0.1; Fig. 3). The systematic errors observed in experimental paradigm I vanished, implying that the error-inducing system must be the ant’s polarization compass and that the error functions shown in Fig. 2 can really be used as a signature of the ant’s polarization compass.
Fig. 3.

Experimental paradigm II. Shown are the homing courses of ants that experienced only the sun during their channel-bound training. In the open-field test situation, they had access either to the sun plus the polarization of the sky (black symbols, paradigm II/1, n = 161) or to the sun alone (green symbols, paradigm II/2, n = 121). The orange curve shows the computed polarization-induced error function characteristic for the particular orientation of the training channel used in these experiments (homeward direction of 270°; see Fig. 2E). For further explanation of conventions, see the legend of Fig. 2.
Because in paradigm I the ants were presented with both direct sunlight and polarized skylight, it is conceivable that the simultaneously activated sun compass could have decreased the systematic errors induced by the polarization compass. To test this hypothesis, we modified paradigm I by letting the ants perform their homebound runs underneath an experimental trolley, which allowed us to occlude the sun (paradigm III) (Fig. 4) so that the ants were left alone with their polarization compass. As a comparison between Fig. 4 and Fig. 2 D and E shows, the daily courses of the error functions coincide in both cases (one-sided pointwise Watson–Williams test, P > 0.1 in all but one case; in the latter case, P > 0.05).
Fig. 4.

Experimental paradigm III. Shown are the homing courses of ants that were deprived of direct sunlight in the open-field test situation. The polarization-induced errors (black symbols, means and standard deviations, n = 262) do not significantly deviate from the errors exhibited by ants that were able to experience both the sun and the pattern of polarized light in the test situation (blue squares, data taken from Fig. 2 D and E). For further explanation of conventions, see the legend of Fig. 2.
This result can be interpreted in one of two ways. One hypothesis implies that the polarization compass dominates over the sun compass to such an extent that, under the polarized sky present in the training situation, the sun compass is virtually ineffective. The other hypothesis implies that, during training, the ants had taken a mean of the output values of the sun compass and the polarization compass system and that this mean value was later used by either compass system. Experimental paradigm IV/1 was designed to decide between these two hypotheses. After training, the ants’ DRAs (and the ocelli) were occluded so that the animals could use only their sun compass. In this case, the homeward courses scattered widely (Fig. 5) and significantly more so than in the case shown in Fig. 4 (F test, half-hour pairwise comparisons, P < 0.05 in all cases but the ones for 1330–1530). Furthermore, the daily course of the error function shown in Fig. 4 is gone. This result, combined with the outcome of the experiments of paradigm III, clearly shows that the ants do not compute a mean value of the outputs of the two compass systems but that the first hypothesis proposed above holds: Cataglyphis relies primarily on its polarization compass. If it has access to both direct sunlight and polarized skylight during training but only to sunlight when later tested, it encounters extreme difficulties in selecting its homeward course. In this context, it is worth recalling that, in principle, Cataglyphis can navigate with its sun compass as precisely as it can with its polarization compass (compare the scatter in the data of Figs. 2E and 3; they do not differ significantly). However, the ants rely only on their sun compass system if, during training, they had to dispense with polarization cues and were left with the sun alone (paradigm II) (Fig. 3).
Fig. 5.

Experimental paradigm IV/1. Shown are the homing courses of ants that were deprived of polarized skylight (by occlusion of their DRAs) in the open-field test situation. Individual data (open circles) and means ± standard deviations in 30-min bins (n = 189) are shown. For further explanation of conventions, see the legend of Fig. 2.
The outcome of experimental paradigm IV/2 was even more dramatic. Here, the ants were prevented from using their polarization compass not by occluding their DRAs but by letting them perform their home runs under the experimental trolley provided with optical filters that completely cut off the short-wavelength range of the spectrum. Now, the ants performed positive phototactic responses toward the solar azimuth (Fig. 6) (i.e., they did not use the sun as a compass cue at all).
Fig. 6.
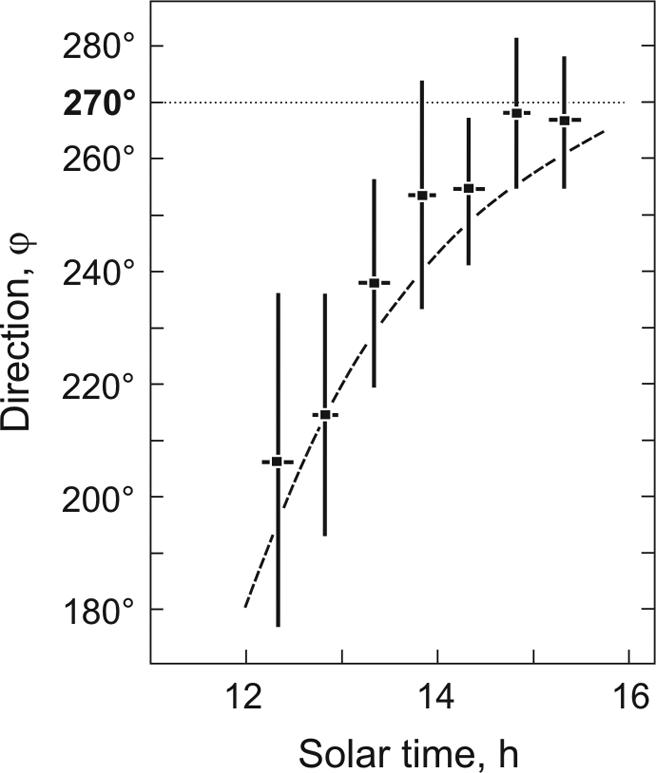
Experimental paradigm IV/2. Shown are the homing courses of ants that were deprived of polarized skylight (by optically cutting off the short-wavelength range of the spectrum) in the open-field test situation. Means and standard deviations (n = 163) are shown. The true homeward direction (ϕ = 270°) is indicated by the dotted line. The dashed line marks the directions that the ants should have taken had they behaved in a purely positive phototactic way (i.e., had they moved toward the azimuthal position of the sun irrespective of their training direction).
Discussion
The Signature of the Polarization Compass.
In analyzing the relative importance of direct sunlight and polarized skylight in the ant’s compass system, we were able to build on a characteristic feature of the polarization compass. Whenever bees and ants that are trained under the full skylight pattern are presented only with a small fraction of this pattern, they exhibit systematic orientation errors depending on what fraction of the entire skylight pattern they have seen. On the other hand, if under full-sky conditions one eye is occluded (by covering it with black paint), the ants can select their true homeward courses, but the trajectories of their courses are more tortuous than those of untreated controls and show frequent deviations to the left and the right (17). The errors induced by presenting bees and ants with individual pixels of polarized skylight have led to hypotheses of how the polarization compass might work. Here, however, it suffices to say that, in our experimental paradigms in which the ants were presented with a partial sky during training and with a full sky in the test situation, such systematic navigational errors indeed occurred and that the error functions corresponded with the predictions outlined in Methods (Fig. 2). Hence, the systematic nature of these errors is a clear indication that the ants have relied on their polarization compass rather than their sun compass system. Whenever the sun is used as a compass cue, systematic errors do not occur (Fig. 3).
The Dominance of the Polarization Compass over the Sun Compass.
If ants have access to both direct sunlight and polarized skylight, their behavior is dominated by their polarization compass. This conclusion follows from a comparison of the results obtained under experimental conditions I and III (Figs. 2 and 4). The navigational errors induced by the ant’s polarization compass do not vanish or even decrease, if, in addition to polarized skylight, the sun is present. If the ants have been trained under sunlight and skylight conditions, and if they later have access only to the sun (paradigm IV), their orientation behavior is severely impaired (Fig. 5), even though their sun compass would have enabled them to select their homeward courses rather precisely (Fig. 3).
Furthermore, the sun is used as a compass cue only if it is presented within the spectral radiance distribution that characterizes the natural sky. If this distribution is destroyed [e.g., by applying optical filters that transmit only the long-wavelength range of the spectrum (λ > 506 nm; paradigm IV/2)], the sun elicits phototactic rather than compass-mediated behavior (Fig. 6). This finding suggests that, in the ant’s celestial navigation system, the sun is not just a bright point-light source in the sky but that its spectral characteristics relative to the remainder of the sky matter. A number of studies performed with honey bees support this suggestion. Bees interpret a spot of unpolarized light as the sun or the antisun depending on whether this spot of light contains long-wavelength or short-wavelength (especially UV) radiation, respectively (18, 19). A reconstruction of the bees’ “spectral image” of the sky showed that bees assume a spot of long-wavelength radiation to lie along the solar meridian and spots of short-wavelength (UV) radiation to lie anywhere within the antisolar half of the sky (11). The observation reported here that the sun is used by Cataglyphis as a compass cue only if it is embedded in the spectral radiance distribution of the sky supports the hypothesis that, also in Cataglyphis, the sun is part of a spectral skylight compass.
Our data further suggest that the ant’s polarization compass works independently of the sun compass. We know already from former behavioral work in which various parts of the ants’ compound eyes have been occluded (9, 13) that, at the photoreceptor level, different input channels are involved: Information about polarized skylight and direct sunlight is picked up by the DRA and the remainder of the dorsal retina, respectively. Our present analysis makes it quite unlikely that the output values of the two compass systems are combined in one way or another. For example, the magnitudes of the systematic errors exhibited by the ants in the open-field test situation do not differ between test paradigms I and III (Fig. 4), in which the ants had access either to both polarized skylight and direct sunlight (paradigm I) or to polarized skylight alone (paradigm III). Hence, in the former test situation, the presence of the sun, which, if presented alone, would have prevented the ants from making any systematic errors at all, did not interfere with the ants’ exclusive use of the polarization compass. The conclusion that information about sunlight and polarized skylight travels in separate channels is further corroborated by experiments designed to test for interocular transfer (IOT). These experiments are performed with monocular ants: One eye is occluded during training; then, in the test situation, the eye cap is moved to the other eye, so that the formerly covered eye becomes the seeing eye and vice versa. Under these conditions, ants that are able to use their polarization compass are still well oriented (17), whereas those that can rely only on their sun compass move in random directions (unpublished data). Hence, Cataglyphis exhibits IOT if it is able to use its polarization compass, but if it is left alone with its sun compass, IOT does not occur.
Methods
Animals and Experimental Site.
All experiments were performed with C. fortis (Formicinae) at our long-term field station near Maharès, Tunisia (lat 34.56°N, long 10.50°E). Because the flat inundation plains near the coastline of Maharès contain wide areas that are free of any vegetation, they constitute ideal test fields for behaviorally unraveling the ant’s celestial compass system without any contamination by landmark-dependent systems of navigation.
Training and Test Procedures.
Ants were trained to run back and forth within linear channels between the nest and an artificial feeder (nest–feeder distance of 15 m). They did so on a horizontal panel of plywood that was inserted into the plastic channels and was covered by a thin layer of sand that was glued to it. The upper borders of the channels restricted the ant’s view of the sky to an elongated, 90°-wide, slit-like window.
Ants to be tested were captured at the feeder and transferred to a test field, where their homebound courses were recorded by means of a grid-work of white lines (mesh width of 1 m) painted on the hard desert floor (for standard techniques, see, for example, ref. 9). In some experimental paradigms, the ants were prevented from using either their sun compass or their polarization compass in either the training or the test situation (Table 1). In test paradigms II/2 and IV/1, the polarization compass was excluded by covering the ants’ polarization-sensitive DRAs as well as their ocelli (20) with light-tight acrylic paint (Lascaux Studio, Bochum, Germany). The ants were immobilized with foam-rubber-coated tweezers, and fine brushes were used to apply the eye covers with the aid of a dissection microscope directly in the field, just before the ants were tested. Another way of excluding the polarization compass during the training procedure was to cover the training channels with depolarizing sheets of grease-proof paper (Utoplex, Basel, Switzerland). Underneath this UV-transmittable paper, the degree of polarization within the UV range of the spectrum was <0.02 [i.e., below the detection threshold for polarized light (8, 21)]. Because the Utoplex paper scattered the direct sunlight to some extent, in experimental paradigm II/1 (Table 1), the ants had to infer the position of the sun from the center of gravity of the celestial intensity distribution rather than from a point-light source. A third way to exclude the polarization compass was applied in the test situation of paradigm IV/2: In this case, the ants were tested underneath an experimental trolley that provided them with a zenith-centered circular skylight window (diameter of 134°; see figure 9 in ref. 9). The circular opening was covered by a UV-absorbing orange Plexiglass disk (transmission < 10−4 for λ < 506 nm, no. 478, Roehm and Haas, Darmstadt, Germany) so that the ants’ polarization channel, which operates only in the UV range of the spectrum (22–24), was shut off. In the test situation of paradigm III, the ants could not see the sun, because they were again tested underneath the experimental trolley. This time, however, the circular opening of the trolley was shaded by a screen mounted on the trolley, so the ants had no access to direct sunlight. Furthermore, the opening of the trolley was covered by a UV-transmitting Plexiglass disk (no. 218, Roehm and Haas).
Data Evaluation and Statistics.
Standard techniques (e.g., ref. 25) were used for determining the direction of the ants’ homebound courses in the test situation. Angular means and standard deviations were computed according to ref. 26. Ref. 26 was also used in testing the following: angular distributions of data for uniformity (Raleigh test), whether a circular distribution of data differed from an expected value (Stephens test), and whether two sets of circularly distributed data differed from each other (Watson–Williams test). An F test adapted to circularly distributed data was applied in testing for differences in the scatter of two data sets.
Computing Polarization-Induced Error Functions.
The theoretical error functions depicted as orange curves in Fig. 2 have been computed on the basis of the navigational errors exhibited by animals that had been presented with isolated pixels of sky (10, 13, 15, 16). Whatever these systematic navigation errors actually mean in functional terms (i.e., whatever the neural mechanisms that underlie the ant’s polarization compass), we have tentatively used these pixel-derived errors to compute the errors that theoretically should result from training the ants within narrow channels. These channels restricted the animal’s view of the sky to 90°-wide, zenith-centered, strip-like windows. In particular, we computed the arithmetic mean of the errors caused by the individual e vectors within the skylight windows and, in doing so, proceeded in 10° steps in both azimuth and elevation (for details see ref. 27). However, we neglected all parts of the sky that had an elevation of <30° above the horizon [because these lower parts of the celestial hemisphere perceived by the ants along the axis of the channel are not covered by the DRA (23)] and that had an angular distance of <30° from the sun [because, in this area around the sun, the degree of polarization is <10% and hence below the detection threshold for polarized skylight (8)]. The resulting theoretical error functions (orange curves in Fig. 2) fit the experimental data sufficiently well. Nevertheless, at least two caveats should be mentioned. First, we have simply taken the arithmetic mean of all pixel-induced errors; second, we have put equal weight on all points of the skylight pattern irrespective of the position of these points within the pattern and irrespective of the degree of polarization prevailing in these points in the sky. All of these assumptions might not fully reflect what is actually going on in the ant’s neural polarization channel. Nevertheless, the at least qualitative agreement between the theoretically derived data and the experimentally established data is striking.
Acknowledgments
We thank Megdalena Götz, Gisela Müller, and Martin Kast for friendship and help in the field; Helmuth Heise for superb construction of the channel systems; and Ursula Menzi for expert help in designing the figures. This work was supported by the Swiss National Science Foundation and the J. and A. Claraz Foundation (R.W.)
Glossary
Abbreviation:
- DRA
dorsal rim area.
Footnotes
Conflict of interest statement: No conflicts declared.
References
- 1.Santschi F. Rev. Suisse Zool. 1911;19:303–338. [Google Scholar]
- 2.Santschi F. Mém. Soc. Vaud. Sci. Nat. 1923;4:127–175. [Google Scholar]
- 3.von Frisch K. Experientia. 1949;5:142–148. doi: 10.1007/BF02174424. [DOI] [PubMed] [Google Scholar]
- 4.Rossel S. In: Facets of Vision. Stavenga D. G., Hardie R. C., editors. Berlin: Springer; 1989. pp. 298–316. [Google Scholar]
- 5.Wehner R. In: Neural Basis of Behavioural Adaptations. Schildberger K., Elsner N., editors. Stuttgart: Fischer; 1994. pp. 103–143. [Google Scholar]
- 6.Wehner R., Srinivasan M. V. In: The Neurobiology of Spatial Behaviour. Jeffery K., editor. Oxford: Oxford Univ. Press; 2003. pp. 9–30. [Google Scholar]
- 7.Wehner R., Labhart T. In: Invertebrate Vison. Warrant E., Nilsson D.-E., editors. Cambridge, U.K.: Cambridge Univ. Press; 2006. in press. [Google Scholar]
- 8.von Frisch K. Tanzsprache und Orientierung der Bienen. Heidelberg: Springer; 1965. [Google Scholar]
- 9.Wehner R. Himmelsnavigation bei Insekten: Neurobiologie und Verhalten. Zürich: Orell Füssli; 1982. [Google Scholar]
- 10.Rossel S., Wehner R. J. Comp. Physiol. A. 1984;154:607–615. [Google Scholar]
- 11.Rossel S., Wehner R. J. Comp. Physiol. A. 1984;155:605–613. [Google Scholar]
- 12.Wehner R., Strasser S. Physiol. Entomol. 1985;10:337–349. [Google Scholar]
- 13.Fent K. Ph.D. thesis. Zürich: University of Zürich; 1985. [Google Scholar]
- 14.Labhart T., Meyer E. P. Microsc. Res. Tech. 1999;47:368–379. doi: 10.1002/(SICI)1097-0029(19991215)47:6<368::AID-JEMT2>3.0.CO;2-Q. [DOI] [PubMed] [Google Scholar]
- 15.Rossel S., Wehner R. Proc. Natl. Acad. Sci. USA. 1982;79:4451–4455. doi: 10.1073/pnas.79.14.4451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wehner R., Rossel S. In: Experimental Behavioral Ecology. Hölldobler B., Lindauer M., editors. Stuttgart: Fischer; 1985. pp. 11–53. [Google Scholar]
- 17.Wehner R., Müller M. Nature. 1985;315:228–229. [Google Scholar]
- 18.Edrich W., Neumeyer C., von Helversen O. J. Comp. Physiol. 1979;134:151–157. [Google Scholar]
- 19.Brines M. L., Gould J. L. Science. 1979;206:571–573. doi: 10.1126/science.206.4418.571. [DOI] [PubMed] [Google Scholar]
- 20.Fent K., Wehner R. Science. 1985;228:192–194. doi: 10.1126/science.228.4696.192. [DOI] [PubMed] [Google Scholar]
- 21.Labhart T. J. Exp. Biol. 1996;199:1467–1475. doi: 10.1242/jeb.199.7.1467. [DOI] [PubMed] [Google Scholar]
- 22.Duelli P., Wehner R. J. Comp. Physiol. 1973;86:37–53. [Google Scholar]
- 23.Labhart T. J. Comp. Physiol. A. 1986;158:1–7. [Google Scholar]
- 24.Labhart T. Naturwissenschaften. 2000;87:133–136. doi: 10.1007/s001140050691. [DOI] [PubMed] [Google Scholar]
- 25.Wehner R., Srinivasan M. V. J. Comp. Physiol. 1981;142:325–338. [Google Scholar]
- 26.Batschelet E. Circular Statistics in Biology. London: Academic; 1981. [Google Scholar]
- 27.Müller M. Ph.D. thesis. Zürich: University of Zürich; 1989. [Google Scholar]