

Abstract
This year, the field of neuroscience celebrates the 50th anniversary of Mountcastle's discovery of the cortical column. In this review, we summarize half a century of research and come to the disappointing realization that the column may have no function. Originally, it was described as a discrete structure, spanning the layers of the somatosensory cortex, which contains cells responsive to only a single modality, such as deep joint receptors or cutaneous receptors. Subsequently, examples of columns have been uncovered in numerous cortical areas, expanding the original concept to embrace a variety of different structures and principles. A ‘column’ now refers to cells in any vertical cluster that share the same tuning for any given receptive field attribute. In striate cortex, for example, cells with the same eye preference are grouped into ocular dominance columns. Unaccountably, ocular dominance columns are present in some species, but not others. In principle, it should be possible to determine their function by searching for species differences in visual performance that correlate with their presence or absence. Unfortunately, this approach has been to no avail; no visual faculty has emerged that appears to require ocular dominance columns. Moreover, recent evidence has shown that the expression of ocular dominance columns can be highly variable among members of the same species, or even in different portions of the visual cortex in the same individual. These observations deal a fatal blow to the idea that ocular dominance columns serve a purpose. More broadly, the term ‘column’ also denotes the periodic termination of anatomical projections within or between cortical areas. In many instances, periodic projections have a consistent relationship with some architectural feature, such as the cytochrome oxidase patches in V1 or the stripes in V2. These tissue compartments appear to divide cells with different receptive field properties into distinct processing streams. However, it is unclear what advantage, if any, is conveyed by this form of columnar segregation. Although the column is an attractive concept, it has failed as a unifying principle for understanding cortical function. Unravelling the organization of the cerebral cortex will require a painstaking description of the circuits, projections and response properties peculiar to cells in each of its various areas.
Keywords: spandrel, barrel, angioscotoma, retinal wave, pinwheel, macaque
1. Introduction
Half a century ago, Mountcastle et al. (1955) made a seminal observation while recording from cat somatosensory cortex. They noted that all cells in a given vertical electrode penetration responded either to superficial (skin, hair) or deep (joint, fascia) stimulation (figure 1). It appeared that for a common receptive field location (e.g. the cat's foreleg), cells were segregated into domains representing different sensory modalities. This discovery led Mountcastle (1957, p. 430) to hypothesize ‘there is an elementary unit of organization in the somatic cortex made up of a vertical group of cells extending through all the cellular layers’. He termed this unit a ‘column’. By making multiple, closely spaced penetrations, Mountcastle concluded that individual columns are no more than 500 μm wide and ‘intermingled in a mosaic-like fashion’. These blocks of tissue contain neurons whose salient physiological properties are identical. The identification of the column has been considered a breakthrough in neuroscience because it seems to simplify the daunting task of understanding one of the most intricate structures in biology: the cerebral cortex.
Figure 1.

Mountcastle's evidence for columnar segregation of skin and deep receptors in areas 1, 2 and 3 of macaque somatosensory cortex. Drawing shows multiple micropipette penetrations from different monkeys, made within 1 mm of the sagittal plane marked by ‘A’ in the surface view below. Horizontal tick marks denote single units; grey shading indicates multiunit recordings. Radial penetrations in area 2 and adjacent area 1 yielded recordings devoted entirely to a single modality, whereas oblique penetrations through areas 1 and 3 resulted in regular switches between skin and deep receptors, presumably as column boundaries were crossed. The width of columns is difficult to judge because parallel penetrations were not shown from the same animal. In this illustration, columns seem to range from 200 μm to more than 1 mm in diameter. Electrode tracks were reconstructed from surface photos of vascular landmarks, electrode carrier dial readings and gliosis in tissue sections. No lesions were made. Mountcastle remarked candidly, ‘The majority of electrode tracks were extremely difficult to find’ (Powell & Mountcastle 1959, p. 135).
Mountcastle's concept of the cortical column has endured as a fundamental principle of brain organization. Yet when one stops to ponder its significance, it becomes apparent that neuroscience has no commonly accepted definition of ‘column’. The term has been used loosely by subsequent investigators to refer to many different entities, some bearing only a vague analogy to the modality-specific columns in the cat somatosensory cortex. Here, we trace the evolution of ideas about columns, challenge the notion that they are essential processing elements within the nervous system and review new insights regarding their formation. Our review deals principally with the visual cortex, where columns have been studied most intensively, but draws freely upon examples from other cortical areas to emphasize certain points.
2. Minicolumns, columns and macrocolumns
Mountcastle proposed the cortical column based on observations made by recording from single cells, but he was influenced strongly by new ideas about the anatomy of the cortex. In the discussion section of his original report, he referred to recent work by Sperry & Lorento de Nó. The latter had reconstructed the complete morphology of single cells using the Golgi method to unravel the circuitry uniting different cortical layers. Tracing synaptic relationships, Lorento de Nó was struck by the predominance of vertical, rather than horizontal connections between cells in the cortical layers. This impression led him to suggest that a vertical cylinder containing all the elements of the cortex ‘may be called an elementary unit, in which, theoretically, the whole process of the transmission of impulses from the afferent fibre to the efferent axon may be accomplished’ (Lorento De Nó 1949, p. 304). Mountcastle's physiological data emphatically supported this view of the cortex as a conglomeration of independent vertical processing machines. It also seemed to explain an amazing series of lesion experiments by Sperry et al. (1955), in which they sliced the visual cortex of cats into pieces with tantalum wires (figure 2). Despite severance of long-range horizontal projections, the cats could still make fine visual discriminations. It was presumed that vision remained normal because the vertically organized columns required for cortical function were still intact.
Figure 2.
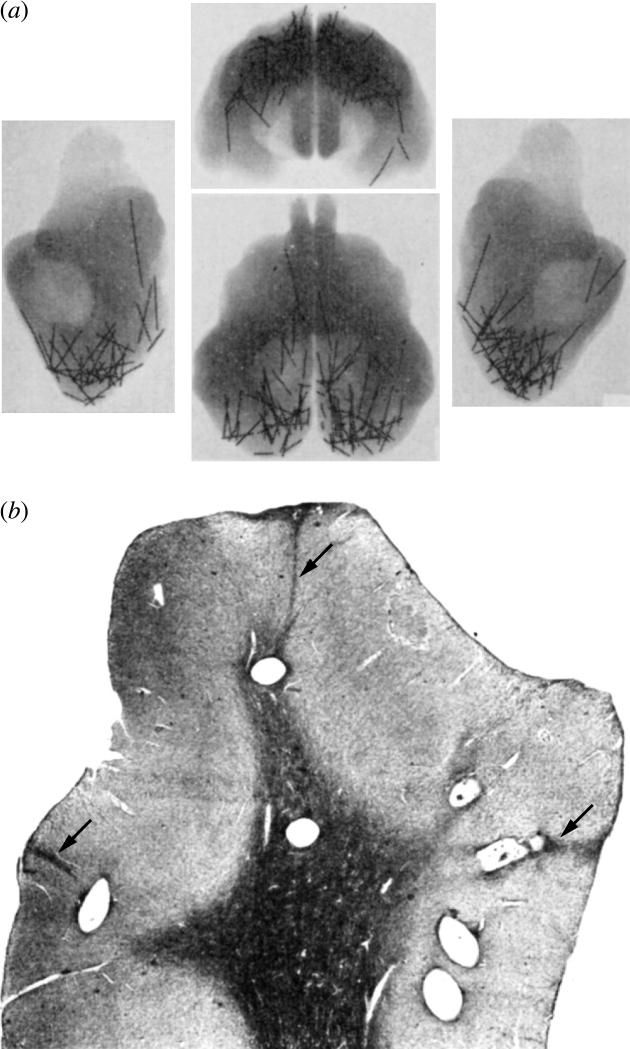
Implanted wires in the cat's visual cortex produce no disturbance in form perception. (a) X-rays of a formalin-fixed cat brain showing segments of tantalum metal inserted months before death to sever horizontal projections. This procedure produced no loss of the ability (measured before and after surgery), to differentiate geometric figures. (b) Coronal section through the posterior lateral gyrus prepared after wire removal, showing seven holes in the primary visual cortex. Note scars (arrows) left from cheese-wiring of the tantalum through the cortex. The wires probably failed to impair visual function simply because they were placed too far apart (Sperry et al. 1955).
The first apparent confirmation of Mountcastle's discovery of cortical columns came from Hubel & Wiesel's (1962, 1968) pioneering studies in the primary visual cortex. They observed that cells recorded simultaneously at any given electrode site invariably had the same receptive field orientation. Moreover, when they advanced an electrode along a penetration perpendicular to the cortical surface, successively recorded cells shared an identical orientation of their receptive field axis. Hubel & Wiesel concluded that striate cortex contains a system of orientation columns superimposed on the 2D representation of the visual field. They drew a parallel with the columns identified by Mountcastle, but noted an important difference. In the somatosensory cortex, the columns arise from different types of peripheral receptors (e.g. Golgi tendon organs versus Meissner's corpuscles). In the visual cortex, by distinction, the orientation columns are generated by patterns of intrinsic wiring between afferents and cortical cells, but originate from the same receptors in the retina.
Mountcastle reported an abrupt transition from one submodality to another in electrode penetrations through the cat somatosensory cortex. This observation established a cardinal property of the cortical column: it is quantal. A column has a sharp (or at least a definite) border and all cells within it share some common, salient property. To see if orientation columns satisfy these criteria, Hubel & Wiesel (1963) plotted the preferred orientation of cells encountered successively during long tangential electrode penetrations through cat visual cortex. As the electrode advanced, they formed the impression by listening to multiunit activity on an audio monitor that the preferred orientation of cells shifted in steps of 15°. Orientation tuning, therefore, appeared to change abruptly from one column to the next, conforming to Mountcastle's stipulation that columns are discrete. In the cat, 12 orientation columns were calculated to exist in a full rotation of 180°. Later, recording in the macaque, Hubel & Wiesel had more difficulty convincing themselves that columns were truly discrete because the preferred orientation seemed to shift with even the smallest electrode advance. Ultimately, they were persuaded by occasional, large jumps in orientation preference to stick with the idea that orientation columns are quantal. However, they conceded that if orientation varies continuously, ‘either one must broaden the definition of the column or decide that the system is not strictly columnar’ (Hubel & Wiesel 1974a, p. 289). Tacitly, the definition of the column has been broadened over subsequent years to include structures without distinct borders. This usage acknowledges the reality that many periodically iterated structures in the cortex have gradual boundaries. It seems unproductive to worry too much about how sharp a column's boundary must be to qualify still as a column.
Hubel & Wiesel (1974a) reported that shifts of 9 to 10° in orientation tuning occurred with electrode advances through the macaque striate cortex of only 25–50 μm (figure 3). Their data suggested that orientation columns might be extremely thin—perhaps only a single cell wide. If so, by definition, orientation columns would be quantal. In Nissl-stained sections, prominent bands of cells are visible running radially through the cortical layers (figure 4). To test the relationship between these palisades of cells and orientation columns, Hubel & Wiesel (1974a) analysed a tangential electrode penetration made through striate cortex. They found 22 orientation shifts along a traverse of 32 cell bands, ruling out a direct, one-to-one correspondence. In retrospect, the comparison was not valid, although it led to the correct conclusion. Subsequent studies have shown that at any given recording site, neurons display scatter in their preferred orientation and are often broadly tuned (Hetherington & Swindale 1999; Ringach et al. 2002). It has become clear that orientation columns are not discrete entities, and therefore that Hubel and Wiesel's designation of 22 orientation shifts was arbitrary. Orientation tuning does exhibit occasional fractures and singularities (pinwheels)—which convinced Hubel and Wiesel that orientation is quantal—but in fact, optimal orientation varies smoothly across most of the cortical surface (Albus 1975; Blasdel & Salama 1986). Therefore, Hubel and Wiesel's classic model showing orientation columns as discrete slabs is misleading because the columns are borderless in real life. The diagram is still useful, however, because it captures an early view of how multiple column systems in the primary visual cortex might be organized.
Figure 3.
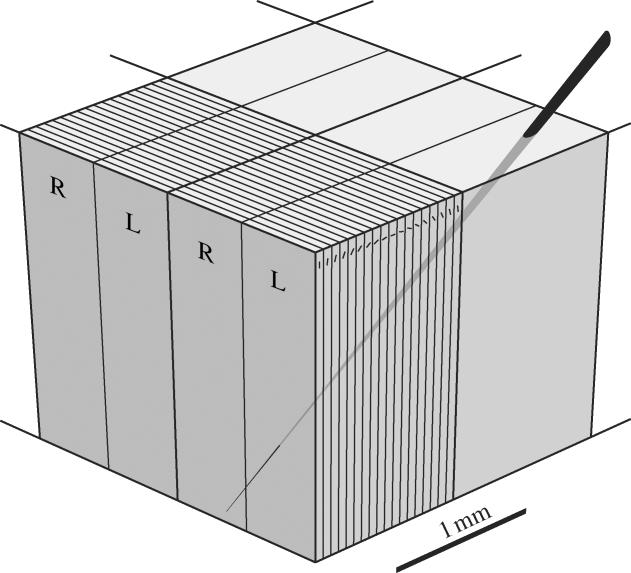
Hubel and Wiesel's ice cube tray model of the striate cortex. In oblique microelectrode penetrations, they attributed the regular shift in orientation preference to an orderly stacking of slab-like orientation columns. Orientation hypercolumns contained 18 discrete columns about 50 μm wide, each 10° apart. Ocular dominance hypercolumns consisted of a single right (R) eye column and left (L) eye column about 500 μm wide. This diagram implies that a module consists of about four orientation and ocular dominance hypercolumns, measuring approximately 2×2 mm. Electrode movement in the cortex from one module to another was thought to produce a non overlapping displacement in the location of the aggregate receptive field of neurons. Hubel & Wiesel (1979, p. 160) later proposed that the combination of a single-orientation hypercolumn and ocular dominance hypercolumn ‘can be considered an elementary unit of the primary visual cortex’ (After Hubel et al. 1976).
Figure 4.
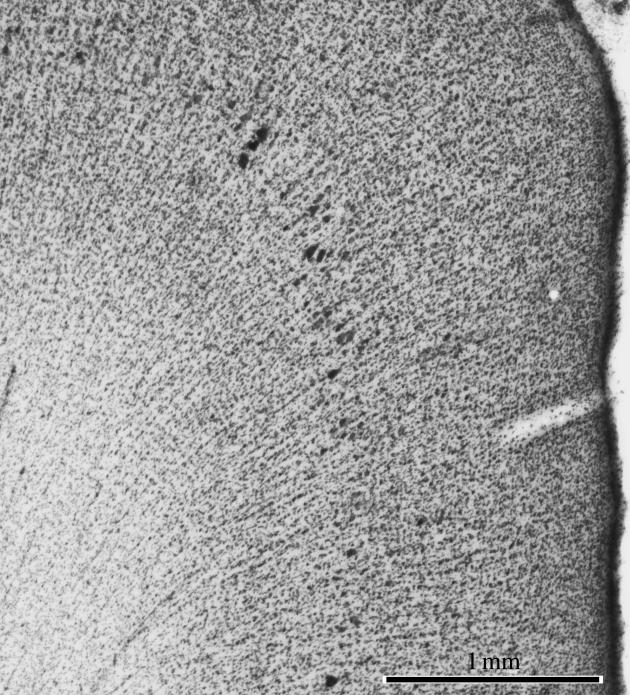
Ontogenetic columns extend from white matter into cortex. A cross-section through macaque somatosensory cortex stained for Nissl substance shows radial stacks of cells extending from the white matter into the cortical layers. These remnants of foetal development have no known functional relationship to columns in the adult cortex.
Although the prominent cell bands seen in Nissl sections do not represent individual orientation columns, they provide persuasive evidence that the adult cortex is composed of discrete, elementary units generated during development. Across many species and cortical regions, these bands have a width of between 30 and 80 μm and contain a few hundred cells (Rockel et al. 1980). Rakic (1971) has shown by [3H]thymidine autoradiography in foetal monkeys that neurons destined for the neocortex are born in a proliferative region near the cerebral ventricles. This ventricular zone contains radial glia, which are stem cells for neurons (Noctor et al. 2001; Tamamaki et al. 2001; Anthony et al. 2004). After birth, neurons migrate successively along a scaffold provided by radial glia into the cortical plate, passing earlier arrivals to generate the layers in an inside-out fashion (Angevine & Sidman 1961). This process produces vertical stacks of cells, divided by glial septa, which extend from the white matter to the pial surface. Rakic (1988) has called these palisades of cells ‘ontogenetic columns’. Studies of cell lineage using a retroviral vector carrying the β-galactosidase marker gene have suggested that tangential migration of neurons also occurs in the developing cortex (Walsh & Cepko 1988, 1992). This observation may be explained by lateral displacement of progenitor cells in the ventricular zone (Reid et al. 1995) and by tangential movement of young neurons through the cortex (O'Rourke et al. 1995). The relative amount that cells travel radially versus laterally through the developing cortex has been controversial; it clearly varies by species, cell type, brain region and developmental stage (Marin & Rubenstein 2003). In the final analysis, although tangential migration of neurons is an important feature of cortical development, the predominant pattern of cell movement is vertical (figure 5), as proposed by the radial unit hypothesis of cortical formation (Rakic 1995, 2003).
Figure 5.
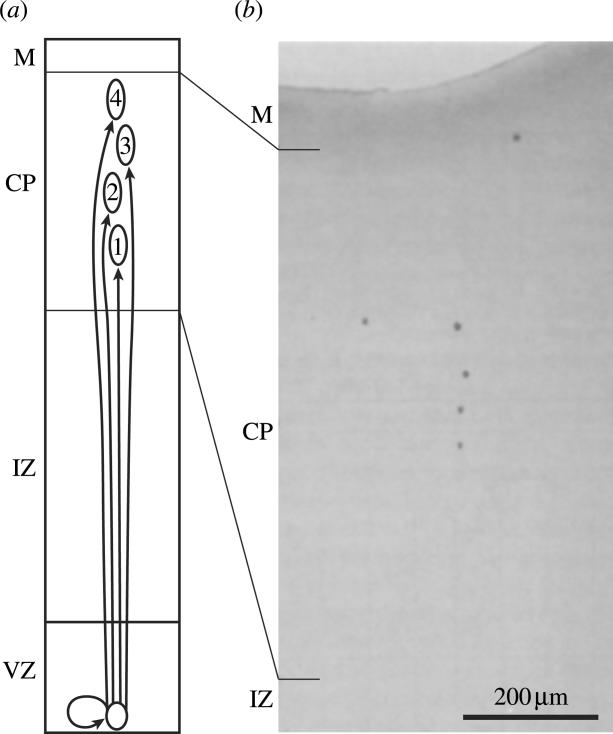
Formation of radial minicolumns or ontogenetic columns. (a) Progenitor cells in the ventricular zone (VZ) give rise to progeny that migrate in succession along a glial scaffold into the cortical plate (CP). These cells remain roughly aligned along their migratory trajectory, giving rise to vertical minicolumns. (b) A β-gal+ clone of cells derived from an asymmetrically dividing stem cell, shown at various stages of vertical migration. The probe was introduced by intraventricular injection of a retroviral probe in the monkey foetus (Rakic 1995).
From the standpoint of cortical processing, the crucial question is whether the radial cell units generated during development correspond in any sense to the functional columns proposed to exist in the mature brain. The original columns identified in the adult cat somatosensory cortex by Mountcastle were much larger than the radial units identified by Rakic—by an order of magnitude. Aware of this dilemma, Mountcastle (1978, p. 37) later invoked a new entity: the minicolumn. He stated, ‘I define the basic modular unit of the neocortex as a minicolumn. It is a vertically oriented cord of cells formed by the migration of neurons from the germinal epithelium of the neural tube along the radial glial cells to their destined location in the cortex, as described by Rakic’. In a subsequent review article, Rakic (1988, p. 176) seemed to agree, asserting that ‘ontogenetic columns become basic processing units in the adult cortex’. He then stipulated, in a rather contradictory footnote, ‘the ontogenetic columns…should not be confused with the minicolumns’. Ultimately, however, Rakic (personal communication) has embraced the idea that the minicolumns of Mountcastle and the radial ontogenetic columns are closely related, as they must be, according to a literal interpretation of Mountcastle's definition.
If the minicolumn is truly the basic unit of the cortex, then what about the column, originally proposed as the elementary physiological unit? If two different structures are defined as the fundamental unit of the cortex, then there is potential for confusion. Mountcastle (1997, p. 701) solved this dilemma by subsequently linking the two structures, declaring that a column is ‘formed by many minicolumns bound together by short-range horizontal connections’. The larger structure, the ‘column’, has been redefined by others as a ‘macrocolumn’ (Buxhoeveden & Casanova 2002). Whichever label one prefers, the concept that minicolumns are bound together like sticks in a fascicle to form larger functional units has several problems. First, nobody seems willing to venture how many minicolumns constitute a column; the number is arbitrary. Second, there is no evidence that short-range connections bind minicolumns into discrete, larger structural entities. In the visual cortex, for example, injection of a tracer at any site labels a diffuse cloud of local connections of about 500 μm in diameter (Callaway 1998; Lund et al. 2003). These short-range connections do not end abruptly along fixed borders in the cortex, as they should if they actually defined the edges of a structurally distinct column. It is true that long-range connections in the cortex terminate in clusters. These clusters unite groups of cells sharing the same receptive field properties (Gilbert & Wiesel 1989; Bosking et al. 1997; Sincich & Blasdel 2001). Such regular, patchy intracortical connections constitute a type of columnar structure, as discussed further below. They lend no support, however, to the theory that the cortical column is a distinct unit composed of an aggregate of minicolumns.
Equally dubious is the concept that minicolumns are basic modular units of the adult cortex, rather than simply remnants of foetal development. The concept derives from the observation that minicolumns are isolated by cell-sparse zones of neuropil into semi-distinct compartments. However, one could also assert that minicolumns are united by synapse-rich zones of neuropil, linking them seamlessly in the cortical sheet. The ‘elementary unit’ hypothesized by Lorento de Nó remains an elusive quarry. No one has demonstrated a repeating, canonical cellular circuit within the cerebral cortex that has a one-to-one relationship with the minicolumn. Moreover, there is little reason to believe that individual minicolumns are discrete functional entities like cortical columns are. If they were, cells in the same minicolumn would share receptive field properties, and these properties would differ from those found in adjacent minicolumns. As mentioned above, Hubel & Wiesel (1974a) made a valiant attempt to correlate orientation columns with minicolumns but found no relationship. Swindale (1990) has pointed out that no stimulus property has ever been shown to be represented in the cortex in a physiologically discrete manner on a scale compatible with the minicolumn. Buxhoeveden & Casanova (2002) have taken issue with this assertion, citing evidence for discrete minicolumn function adduced by Favorov et al. (1987) in cat somatosensory cortex. The latter investigators found occasional abrupt shifts in receptive field location, thought to occur at microelectrode transitions from one ‘segregate’ to another. The ‘segregate’—a new term introduced by Favorov—turns out to be a structure about 350–600 μm wide and resembles precisely a Mountcastle column. Therefore, the study by Favorov and colleagues provides evidence confirming Mountcastle et al.'s (1955) original observations about columns, but does not address the functional properties of minicolumns. It remains true that nothing is known about the physiological correlates (if, indeed, any exist) of the minicolumn.
Why have there been virtually no studies probing the physiology of the cortical minicolumn? A major impediment is the poor spatial resolution of the extracellular recording method. Often, the same spike can be picked up over a traverse of 100 μm, depending to some extent on the electrode impedance, construction and tip shape (Mountcastle et al. 1957; Rosenthal et al. 1966). With such uncertain localization, it would be impossible to show that a receptive field property differs systematically from one minicolumn to the next, even if it did. It is also difficult to orient electrode penetrations in a perfectly radial or tangential trajectory and to reconstruct tracks reliably for post-mortem correlation of physiological properties and histological findings. Such inherent limitations have plagued efforts to show that the minicolumn (or the column for that matter) has distinct properties and borders. Two-photon imaging of calcium fluxes allows one to study intact neuronal circuits in vivo with the resolution of individual cells (Stosiek et al. 2003). The advent of this technique may finally allow one to probe the functional properties of the minicolumn.
3. Hypercolumns and modules
In his original study, Mountcastle found that some electrode penetrations were not modality-pure, but contained a mixture of cells responsive to skin or deep stimulation. Often penetrations were oriented obliquely with respect to column borders (figure 1), explaining the mixture of units. In some cases, the recording electrode might have picked up units from adjacent columns. An alternative interpretation is that the modality columns in somatosensory cortex truly lack precise borders and instead, cells with common receptive field properties vary gradually in relative concentration (Slimp & Towe 1990). Different modalities are represented preferentially in areas 1, 2 and 3, complicating the interpretation of the electrode recordings. The sharpness of the transition from one column to another in somatosensory cortex amounts to a question of degree and remains to be settled definitively. The important concept is the ‘periodicity principle’: cells are clustered according to some common receptive field property and these cell aggregates recur periodically within a given cortical area. In the somatosensory cortex, little is known about the spatial array formed by columns. In the visual cortex, on the other hand, this problem has been investigated extensively, as reviewed by LeVay & Nelson (1991).
In macaque striate cortex, Hubel and Wiesel found that optimal orientation tuning changes systematically through 180° with an electrode advance of between 0.5 and 1.0 mm. They invented the term ‘hypercolumn’ to refer to a complete rotation of columns (e.g. 0, 10, 20…180°; Hubel & Wiesel 1974b). Although the view of orientation columns as discrete slabs has been abandoned, the hypercolumn is still a valid concept, denoting a unit containing a full set of values for any given receptive field parameter. In the visual cortex, optical imaging has provided a vivid impression of the pattern formed by orientation columns (Blasdel & Salama 1986; Bonhoeffer & Grinvald 1991). It has also been used to reveal maps for direction preference, spatial frequency (Weliky et al. 1996; Hübener et al. 1997; Everson et al. 1998; Issa et al. 2000) and perhaps colour (Ts'o et al. 2001b; Landisman & Ts'o 2002; Xiao et al. 2003). For each of these systems, the hypercolumn seems to be approximately the same in size. This common periodicity means that any block of cortex, roughly the size of a single hypercolumn, contains cells tuned to all values of every receptive field variable (Swindale et al. 2000). Hubel & Wiesel (Hubel 1982) applied the term ‘module’ to this tissue block comprising multiple, overlapping hypercolumns. Mountcastle (1997) later defined ‘module’ quite differently, choosing to use the word interchangeably with ‘column’. Eventually, one hopes that these discrepancies in terminology will be resolved.
Typically, studies of columnar organization in visual cortex have systematically varied one receptive field parameter, while keeping all others constant. This approach has generated column maps for different stimulus features, each viewed in isolation. It has been tacitly assumed that cells responsive to any particular combination of stimulus values lie at the intersection of columns belonging to each system. A critical shortcoming of this model is that it becomes increasingly exclusionary with the discovery of each new columnar system. At some point, it becomes impossible for cells tuned to every conceivable combination of stimulus variables to be located physically at the interstices of multiple, overlapping column systems. Previously, the solution to this dilemma was to invoke retinotopic ‘scatter’ in the location of receptive fields. However, the scatter of receptive fields is much less than previously assumed, making this model untenable (Hetherington & Swindale 1999; Blasdel & Campbell 2001; Adams & Horton 2003b).
Recently, Fitzpatrick and colleagues (Basole et al. 2003) have taken a fresh approach to the problem of mapping multiple parameters in the visual cortex by stimulating with varying combinations of orientation and axis of motion. Their data show that radically different optical imaging maps can be produced by short bars of identical orientation moving in different directions (figure 6). The bars' aspect ratio has a critical impact on the pattern of column activation, except in the special case of motion orthogonal to the bars' orientation. Speed of stimulus motion is also a vital parameter. These new findings suggest that population activity in primary visual cortex is better described as a single map of spatio-temporal energy, not as the intersection of multiple, independent maps. The map exists because there is columnar structure to spatio-temporal tuning, as shown by single cell recordings demonstrating that nearby neurons share an extremely similar spatio-temporal receptive field profile (DeAngelis et al. 1999). This conclusion implies that the concept of orientation columns, for example, being rooted in a fixed location in the cortex is an artefact of using impoverished stimuli that vary along only one axis in Fourier space (e.g. long bars drifting perpendicular to their orientation). It does not vitiate the notion of the hypercolumn—whatever combination of stimuli is used, the resulting map always has the same basic periodicity. However, it alters profoundly the interpretation of recent optical imaging studies of striate cortex, which were based on the old model of fixed, independent column maps for each stimulus parameter.
Figure 6.

Different columns are activated by the same stimulus orientation. (a) Optical images of ferret primary visual cortex showing difference images produced by a texture of vertically moving bars oriented at 45°. The inset is shown at higher power below, with iso-orientation contours overlaid from a grating angle map. (b) The same region, after stimulation with horizontally moving bars oriented at 45°. The pattern of activation is quite different, although the stimulus bars were presented at the identical orientation. Red dots are placed to facilitate comparison between (a) and (b) (Basole et al. 2003).
Orientation columns have been demonstrated by single cell recordings in striate cortex of many species including apes, monkeys, marmosets, galagos, cats, tree shrews, ferrets, minks and sheep (LeVay & Nelson 1991). Curiously, there is little evidence for orientation columns in mice, rats, hamsters and rabbits (Chow et al. 1971; Drager 1975; Shaw et al. 1975; Tiao & Blakemore 1976; Murphy & Berman 1979). The absence of orientation columns in some species might be explained by a lower prevalence of oriented cells or coarser orientation tuning. However, Girman and co-workers (1999) have reported that in the rat, for example, more than 90% of cells are tuned with a bandwidth less than or equal to 30° at half-height. This degree of tuning, which exceeds prior estimates in the rat (Shaw et al. 1975; Wiesenfeld & Kornel 1975), rivals the orientation selectivity found in the macaque. This means that poor orientation tuning can be excluded as the explanation for the lack of orientation columns in rats.
An alternative explanation is that species lacking orientation columns have relatively poor visual acuity or a small visual cortex. Perhaps orientation columns develop only in species that depend on form vision and those that have a cortical surface area sufficiently large to require subdivision into columns. This argument is contradicted, however, by a recent study of orientation tuning in the squirrel. This animal has a cone-rich retina and it surpasses the tree shrew and mink in visual acuity and visual cortical area. Although the tree shrew and mink both have beautifully organized orientation columns, the squirrel has none (Van Hooser et al. in press). Nonetheless, its cortex is endowed abundantly with oriented cells, having a median bandwidth of 28° at half-height. The absence of orientation columns in squirrels suggests that they lack some visual attribute present in species that possess orientation columns. However, so far, no such faculty has emerged as a candidate. This inconsistency in the existence of orientation columns across species provides an important piece of circumstantial evidence in support of our thesis that columns may be irrelevant.
4. Ocular dominance columns
In many species, ganglion cells project to distinct, monocular laminae within the retina's principal thalamic target, the lateral geniculate nucleus (LGN). This ocular segregation is perpetuated at the next level of processing, the primary visual cortex. Axon terminals from geniculate cells serving either the right eye or the left eye terminate in a system of alternating stripes called ‘ocular dominance columns’ (Hubel & Wiesel 1969). In a sense, they are more analogous to Mountcastle's columns than orientation columns because they seem to arise as a solution to the problem of representing two different peripheral receptors on the same topographic surface. Instead of deep and skin receptors, however, the end organs are the eyes, whose signals must be dovetailed into a common representation of the visual field. Like modality columns, ocular dominance columns are indisputably discrete and exert their influence through all layers. They differ, however, because the two values they represent are simply left and right, not different classes of sensory receptors. A system of columns that segregates magnocellular and parvocellular geniculate inputs would provide a better analogy, but these are segregated in the visual cortex by layer. Parvocellular neurons terminate in layer 4Cβ, whereas magnocellular neurons supply layer 4Cα (Hubel & Wiesel 1972). Surprisingly, however, the third geniculate channel—koniocellular neurons—appears to segregate both by layer (4A; Chatterjee & Callaway 2003) and by column (cytochrome oxidase (CO) patches; Hendry & Yoshioka 1994). Clearly, geniculate input to primary visual cortex is not constrained to follow any simple rule of laminar versus columnar organization.
Columns are often regarded as a special feature of the cortex, but retinal input to the superior colliculus is segregated into parallel stripes that resemble closely ocular dominance columns (Hubel et al. 1975). The retinal input to the lateral geniculate is also segregated by eye, into at least 10 (the classical six, plus konio) individual monocular laminae. These laminae are stacked and aligned retinotopically—segregating ocular inputs while representing them at common points in the visual field map—the same end met by ocular dominance columns. In the cortex, the equivalent arrangement would be ocular segregation by layer. Instead, the layers are dedicated to other tasks: the elaboration of cell tiers with different receptive field properties and the organization of intrinsic and extrinsic projections. The layers also give rise to binocular cells, by blending the output of monocular units in layer 4C. However, in at least one species, the tree shrew, ocular inputs are partially segregated by layer (Casagrande & Harting 1975; Hubel 1975). Geniculate projections serving the contralateral eye fill layer 4 and a thin tier in the upper layers. Those driven by the ipsilateral eye are confined to layer 4, with a narrow cleft in the middle. Neither eye's input is segregated into vertical columns.
The above examples show variation in anatomical design and provide indirect evidence that columnar organization is not essential in the cortex. If it were, why would konio input (but not magno and parvo input) be segregated into columns (CO patches) in the macaque striate cortex? Why would tree shrews possess ocular dominance layers, rather than ocular dominance columns?
In certain species, the ocular dominance columns are so robust that they have assumed special importance, beyond any intrinsic significance, as a means for probing neural development (Katz & Crowley 2002), exploring the molecular basis of cortical plasticity (Bear & Rittenhouse 1999) or validating new methods in neuroscience (Horton et al. 2000). In normal animals, they can be detected using a diverse array of techniques including single cell recording, silver stains, anterograde transneuronal tracers and optical imaging (LeVay et al. 1975, 1985; Hubel & Wiesel 1977; Blasdel & Salama 1986; Blasdel 1992a). Another approach is to lesion one eye or to silence it with tetrodotoxin and then to process striate cortex for virtually any substance: Nissl, myelin, metabolic enzymes, neurotransmitters, synthetic enzymes or receptors (Haseltine et al. 1979; Horton 1984; Wong-Riley & Carroll 1984; Chaudhuri & Cynader 1993; Hendry & Calkins 1998; Fonta et al. 2000). CO, a mitochondrial enzyme, is one of the easiest techniques to apply (Wong-Riley 1989). After loss of visual function in one eye, the CO level diminishes in columns formerly served by the affected eye. As a result, a pattern of dark and light CO stripes emerges in layer 4C (figure 7). These stripes are identical to those labelled anatomically by injection into one eye of a transneuronal tracer, such as [3H]proline, proving that they represent ocular dominance columns (Sincich & Horton 2003a). The most effective way to visualize their complete pattern is to unfold the occipital lobe and to flat-mount the cortex prior to sectioning. Local regions of layer 4C from individual flattened sections can be montaged to reconstruct the entire mosaic formed by the ocular dominance columns. In macaques, comparisons among normal animals have revealed a surprising, twofold variation in the periodicity of ocular dominance columns (Horton & Hocking 1996b), a finding confirmed in cats (Rathjen et al. 2002). In the latter species, orientation columns also show considerable intrinsic variability in size (Kaschube et al. 2002). It remains unknown, for any given species, if animals with wider ocular dominance columns also have wider orientation columns. They should do if one subscribes to the idea of a common modular structure comprising multiple, overlapping hypercolumns. If they do not, then the periodicity and formation of each column system must be regulated independently within any given cortical area.
Figure 7.

Ocular dominance columns in macaque striate cortex flatmount. (a) Drawing of the left cortex of a normal macaque monkey showing ocular dominance columns supplied by the left (black) and right eye (white). Columns are present everywhere except in the representation of the monocular crescent (MC) and the blind spot (asterisk). The grey sliver extending from 1 to 4° corresponds to a shallow sulcus where the columns could not be reconstructed. (b) CO montage of layer 4C after right eye enucleation, showing the column pattern used to produce (a). (c) Autoradiograph prepared from alternate sections after injection of [3H]proline into the remaining left eye. There is a nearly perfect match between (b) and (c), validating CO histochemistry as a method for labelling ocular dominance columns (Sincich & Horton 2003a).
It seems hard to imagine that the intricate mosaic formed by ocular dominance columns has evolved within the brain without serving a useful purpose. United by this belief, many laboratories have searched for the function of ocular dominance columns. It is known that they partition the striate cortex into monocular and binocular regions, as well as into left eye and right eye domains. Cells located in the centre of ocular dominance columns (‘core zones’) are more likely to be monocular, whereas cells along column borders (‘border strips’) tend to respond equally well to both eyes. This periodic oscillation in binocularity can be revealed by examining the pattern of CO activity in adult macaques after monocular eyelid suture (Horton & Hocking 1998). In normal animals, CO activity in layer 4C is homogeneous. In animals lid-sutured in adulthood, a pattern emerges of thin dark bands alternating with wide pale bands. The pale bands are wider than the ocular dominance columns because metabolic activity is lost not only within the deprived eye's ocular dominance columns, but also along the border strips of the open eye's ocular dominance columns where binocular cells are concentrated (figure 8). Presumably, many of these cells have a lower firing rate because they are no longer stimulated binocularly at the optimal stimulus disparity. Robust enzyme activity is retained only within the core zones of the open eye's ocular dominance columns because cells in this region are relatively monocular and, hence, are still driven effectively when the other eye is closed.
Figure 8.

Monocular core zones and binocular border strips in primary visual cortex. Schematic of layer 4C in normal macaque striate cortex, showing the ocular dominance column borders marked with brackets. Each ocular dominance column consists of a monocular core zone containing CO patches and a binocular border strip. The transition between the two zones, although marked with a dashed line, is gradual. Their CO content is equal, giving layer 4C a homogenous appearance in normal animals. After monocular enucleation, CO activity is lost in the core zones and border strips of the missing eye's ocular dominance columns (below left). After lid suture, CO activity is reduced in the closed eye's ocular dominance columns and in the open eye's border strip regions, creating a pattern of thin dark columns alternating with wide pale columns (below right). Their contrast is low because eye lid suture has less effect on CO activity than enucleation (Horton & Hocking 1998).
The concentration of binocular cells along column boundaries suggests that ocular dominance columns may serve to organize disparity-sensitive units in striate cortex. This issue has been a source of confusion and controversy. Poggio et al. (1988) have identified various classes of disparity-sensitive cells, including tuned excitatory cells, near cells and far cells. Although it is now generally agreed that a continuum exists, this system of classification is still useful. Tuned excitatory cells give an enhanced response to binocular stimulation near zero disparity, whereas far and near cells respond best over a wide range of positive and negative disparities, respectively. When tested monocularly, tuned excitatory cells usually fire equally well to stimulation of either eye, whereas near and far cells tend to be dominated by one eye (Poggio & Fischer 1977; Ferster 1981). Based on this result, one would expect tuned excitatory cells to be located preferentially along border strips and near/far cells in core zones. Gardner & Raiten (1986), however, found a different correlation between disparity sensitivity and ocular dominance. They reported that disparity-tuned cells are dominated by one eye or else require simultaneous, binocular stimulation. Units that respond well to stimulation of each eye alone (typically referred to as ‘binocular’ cells) are less apt to be disparity tuned. This conclusion would place most disparity cells, regardless of class, in the middle of ocular dominance columns where monocular cells are more prevalent. Subsequently, LeVay & Voigt (1988) failed to detect any correlation between overall disparity sensitivity and binocularity, but they did confirm that tuned excitatory cells have balanced ocularity, while near and far cells usually favour one eye. Recently, Read & Cumming (2004), using random-dot stimuli in awake macaques, have denied any correlation between ocular dominance, disparity class (near/far/tuned excitatory/tuned inhibitory) or strength of disparity tuning. Their negative result, based on a careful, quantitative analysis, makes it difficult to ascribe a role in the organization of disparity-tuned cells to ocular dominance columns.
Another way to search for clues to the function of ocular dominance columns is to examine their expression across the animal kingdom. Ocular dominance columns are absent in the mouse, rat, squirrel, rabbit, possum, sheep and goat. They are present in the cat, ferret, mink and more than a dozen primate species (reviewed in Horton & Hocking 1996a). The only salient point to emerge is that species with ocular dominance columns are predators. Among mammals, efficient predation requires high-grade stereopsis, but as outlined in the previous paragraph, disparity-tuned cells appear to have no systematic relationship with ocular dominance columns.
Among primates, the squirrel monkey has often been cited as a puzzling anomaly, because it was reputed to lack ocular dominance columns (Hubel et al. 1976; Hendrickson et al. 1978; Livingstone 1996). However, a recent study of 12 normal squirrel monkeys has revealed an intriguing spectrum in the expression of ocular dominance columns (Adams & Horton 2003a). Some animals have well-developed columns, comparable to those found in macaques and humans, whereas other animals have only rudimentary columns (figure 9). In principle, this natural variability among normal squirrel monkeys could be exploited to establish the function of ocular dominance columns. One would simply need to identify a visual faculty present in animals with columns and absent in those without columns. As mentioned above, stereopsis has been a considered a prime candidate for the function of ocular dominance columns. Motivated by this idea, Livingstone and colleagues (1995) assessed stereopsis in the squirrel monkey by measuring visual evoked responses to random-dot stimuli. They concluded that the squirrel monkey can detect stereoscopic depth. However, only a single monkey was tested and the animal's brain was not actually examined to determine if ocular dominance columns were present. Therefore, it is difficult to infer a definitive conclusion from their study. Further research addressing this issue would be valuable, but one predicts that a correlation is unlikely, based on the following logic. At least 30% of squirrel monkeys essentially lack ocular dominance columns. If these animals also lacked stereopsis, then they would face a major handicap. The selective pressure driving the evolution of stereopsis should have eliminated misfits without ocular dominance columns long ago. As a general principle, it seems unlikely that any visual function vital for survival could be absent among 30% of the members of a free-ranging species. Either stereopsis is over-rated as a visual asset or it is unrelated to ocular dominance columns.
Figure 9.

Variable spectrum of ocular dominance columns in squirrel monkeys. The top row shows CO montages of layer 4C, after enucleation of one eye, from flatmounts of the left cortex of four normal animals. From left to right are examples of large, intermediate, fine and nearly absent columns. In animals with fine or nearly absent columns, a pattern is visible corresponding to the representation of retinal blood vessels (arrow). The blind spot representation is pale in three cases following enucleation of the left eye and dark in one case after enucleation of the right eye. Below each montage is a drawing of the column pattern prepared by Fourier filtering the CO montage. MC, monocular crescent; *, blind spot. Boxed regions in the lower panels are shown in figures 12 and 20 at higher power (Adams & Horton 2003a).
In some squirrel monkeys, ocular dominance columns are present throughout only part of the primary visual cortex (figure 10). Such cases provide the strongest evidence against the idea that ocular dominance columns serve a function. This function, if it exists, can be present only in those portions of the visual field that are represented in regions of striate cortex that contain columns. It is hard to conceive of such a piecemeal implementation of a specific visual function, especially given the fact that the expression of ocular dominance columns, when restricted in striate cortex, can differ locally from animal to animal.
Figure 10.

Regional, mirror-symmetric variation in the expression of ocular dominance columns. The ocular dominance columns were labelled in these flatmounts of the left and right cortex from a normal squirrel monkey by removing one eye. CO montages of layer 4C reveal well-segregated columns beyond an eccentricity of 8°. The columns disappear in the macular representation, but are present within the foveal representation (*). Ophthalmological and histological examination of the eyes showed no evidence of retinitis pigmentosa, nor any abnormality that might erase the columns in this regional fashion. What function could ocular dominance columns serve in such an animal? (Adams & Horton 2003a).
5. Cytochrome oxidase patches
Perhaps one of the most convincing examples of columnar structure is provided by the distribution of CO in the primary visual cortex. A regular array of ‘patches’ or ‘puffs’ of increased enzyme activity is visible in tangential sections (Horton & Hubel 1981; Wong-Riley & Carroll 1984). These structures, renamed ‘blobs’ by later investigators (Livingstone & Hubel 1984a), are present in every layer except 4C and 4A, forming vertical, interrupted pillars. Although defined originally by their increased CO content, they can be identified using many different labelling techniques (figure 11) because they contain increased levels of lactate dehydrogenase, succinate dehydrogenase, acetylcholinesterase, myelin, glutamic acid decarboxylase, GABA receptors, nitric oxide synthase, NMDA receptors, AMPA receptors, sodium–potassium ATPase, glutamate, CAT-301 and microtubule-associated protein 2 (Hendrickson et al. 1981; Hendry et al. 1984, 1994; Horton 1984; Sandell 1986; Hendry & Bhandari 1992; Carder & Hendry 1994; Carder 1997; Fonta et al. 1997; Wong-Riley et al. 1998). In contrast, CO patches have reduced levels of calbindin, zinc, calcineurin and neurofilament protein (Celio et al. 1986; Goto & Singer 1994; Duffy & Livingstone 2003; Dyck et al. 2003). Differential labelling by such a diverse range of techniques makes it clear that CO patches are defined not simply by increased metabolism, but also by their content of neurotransmitters, receptors and structural proteins.
Figure 11.

CO patches in V1 and CO stripes in V2 are revealed by many different labelling techniques. Here, they are shown by histochemical processing for another metabolic enzyme, β-nicotinamide adenine dinucleotide, reduced form (β-NADH). (a) CO patches in layers 2 and 3 of V1 form rows that follow the pattern of ocular dominance columns (arrows). (b) Adjacent section processed for β-NADH. The pattern of patches is identical (arrows). (c) CO-stained section from dorsal V2, with thick (large arrows) and thin (small arrows) stripes separated by pale stripes. (d) Adjacent section processed for β-NADH, showing the same pattern of stripes. Sections were processed for 2 h in a solution of 100 mg NADH (Sigma, grade III), 45 mg nitroblue tetrazolium and 12 ml DMSO in 100 ml of PBS (Horton unpublished work).
When Hubel & Wiesel (1977) proposed that striate cortex consists of modules, each containing overlapping hypercolumns for all stimulus parameters, their module was envisioned as having arbitrary boundaries. It did not correspond to an actual structure with definable morphology that could be labelled using an anatomical method. Modules were thought to be seamless because the visual field is a continuous sensory surface. Their width was estimated at 2 mm, a value derived from the electrode traverse through the cortex required to produce a net displacement of neurons' aggregate receptive field from one location to another in the visual field. Aggregate receptive field size, defined as the product of individual cells' receptive field size and scatter at any given cortical locus, varies considerably by layer. For example, it is less in layer 4C than in layers 2 and 3 (Gilbert 1977). Consequently, the number of hypercolumns comprising a module was never stipulated clearly. The diagram in figure 3 implied a few; however, elsewhere, a module comprised of a single orientation hypercolumn and ocular dominance hypercolumn was called the elementary unit of the primary visual cortex (Hubel & Wiesel 1979).
After CO patches were discovered, it was immediately obvious that they might correspond to the cortical modules proposed by Hubel and Wiesel. In macaques, patches appear oval, about 250 μm long by 150 μm wide, in tangential sections cut through striate cortex. Each patch is aligned along its major axis into rows spaced about 400 μm apart. Each row fits in a precise register with the centre of an ocular dominance column (Horton 1984). This correspondence, also present in human striate cortex (Horton & Hedley-Whyte 1984), proves that patches have a regular relationship with ocular dominance hypercolumns. However, there are two patches per ocular dominance hypercolumn, and hence, two to eight patches per module. The exact numbers and dimensions are perhaps not important; the essential point is that patches are related systematically to columns serving ocular dominance. Strangely, however, this relationship appears to be present in some species, but not in others. In the cat, the evidence is conflicting (Murphy et al. 1995; Boyd & Matsubara 1996). In the squirrel monkey, no relationship exists between ocular dominance columns (when present) and CO patches (Horton & Hocking 1996a). Figure 12 compares the relationship between patches and ocular dominance columns in the macaque and squirrel monkey. This remarkable inconsistency between species is mystifying and sabotages any effort to find an underlying principle uniting CO patches with other elements of the functional architecture within primate striate cortex.
Figure 12.

Conflicting designs: patches and ocular dominance columns in macaque and squirrel monkey. (a) CO patches in layer 3 of a normal macaque. (b) Montage of the [3H]proline-labelled ocular dominance columns in layer 4C. (c) Each row of patches runs down the middle of each ocular dominance column. Arrows in histological images mark vessels used for alignment. (d ) Patches in layer 3 of a squirrel monkey. (e) Ocular dominance columns in layer 4C, labelled by CO after monocular enucleation. This region corresponds to the box in figure 9, lower left. (f) Random relationship between patches and ocular dominance columns in the squirrel monkey (Adams & Horton unpublished work).
The relationship between CO patches and orientation maps has also been tested, with a welter of conflicting results. Livingstone & Hubel (1984a) made tangential penetrations through macaque striate cortex to correlate orientation selectivity with CO patches. They reported that cells in layers 2 and 3 located within CO patches are unoriented and that their receptive fields are centre-surround, colour-opponent. Ts'o & Gilbert (1988) confirmed this result, adding the remarkable observation that individual patches are dedicated to a single type of colour opponency: red/green or blue/yellow. Lennie et al. (1990) concurred that cells most responsive to chromatic stimuli have poor orientation tuning, but reported that cells' chromatic properties in CO patches are not different from those in interpatches. Leventhal et al. (1995) failed to find any relationship whatsoever between colour tuning, orientation selectivity and CO patches. An important drawback of both these negative studies is that no histological sections with lesions were presented to illustrate the correlation between physiological findings and CO activity. It is worth adding that in the galago and owl monkey, no difference has been detected in the orientation tuning of cells in patches versus interpatches (DeBruyn et al. 1993; O'Keefe et al. 1998). Colour information processing is unlikely to constitute the raison d'être for patches, at least in nocturnal primates like owl monkeys, which have poor colour vision (Horton 1984).
Another way to probe the relationship between orientation tuning and patches is to perform optical imaging followed by histological correlation with CO activity in tangential cortical sections. Maps of orientation tuning contain singularities (Blasdel & Salama 1986), also called pinwheels (Bonhoeffer & Grinvald 1991), where orientation columns converge. The resolution of optical imaging is insufficient to distinguish between two possibilities: (i) pinwheels contain cells lacking orientation tuning, or (ii) pinwheels contain intermingled cells tuned to different orientations. In the cat, tetrode penetrations targeted into pinwheel centres favour the second alternative. Sharply tuned neurons of different orientation preference seem to lie in close proximity (Maldonado et al. 1997). Amazingly, in the macaque no comparable study has been performed. Obviously, it would be paramount to learn if pinwheel centres contain clusters of unoriented, centre-surround cells, as one might predict from Livingstone & Hubel's (1984a) microelectrode studies.
Orientation pinwheels tend to be situated in the middle of ocular dominance columns (Bartfeld & Grinvald 1992; Blasdel 1992b; Crair et al. 1997). This result implies a potential spatial correlation between CO patches and orientation pinwheels because CO patches are also located in the middle of ocular dominance columns, at least in the macaque. In the cat, no data are available regarding the relationship between pinwheels and CO patches. The question is difficult to broach in felines because their patches are faint in contrast. In the macaque, a species with vivid patches, the data are once again at odds. Blasdel (1992b) suspected a direct correspondence, but he hesitated to assert a definitive result (figure 13). Subsequently, Bartfeld & Grinvald (1992) and Landisman & Ts'o (2002) reported that pinwheels and patches do not coincide. This lack of spatial correlation has been confirmed in the owl monkey (Xu et al. 2004).
Figure 13.
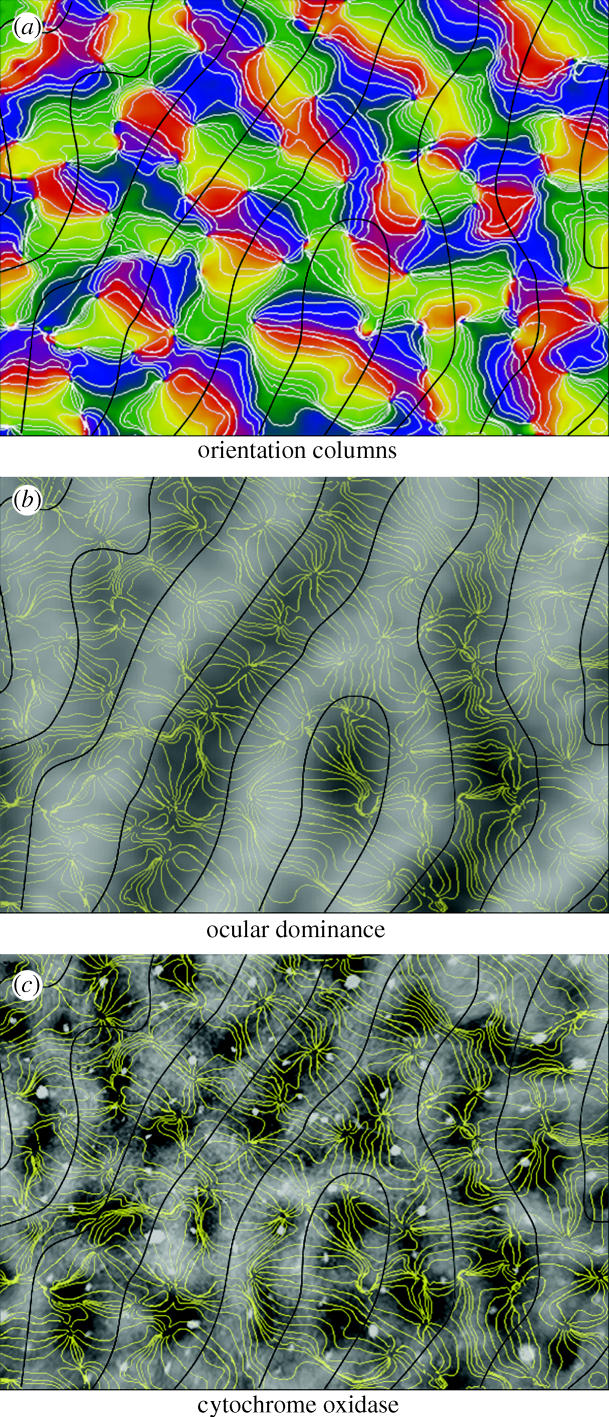
Alignment of singularities (pinwheel centres) and CO patches. (a) Optical imaging map of orientation preference in macaque striate cortex. The black lines represent the borders of ocular dominance columns. (b) Iso-orientation contours from (a) superimposed on an optical imaging map of the ocular dominance columns. Note that orientation singularities tend to be situated in the centres of ocular dominance columns. (c) Comparison between iso-orientation contours and CO patches. In some instances, singularities and patches coincide, but the correlation is far from perfect. There seem to be more singularities than patches, although determining their exact numbers is difficult because neither structure is always well demarcated (Data from Blasdel 1992b with permission).
Before these negative studies are accepted, it is important to consider their limitations. The identification of pinwheels can be arbitrary because not all contain a full rotation of 180° around a single point owing to fractures and irregularities in the orientation map. Photon scatter by brain tissue and optical blur in imaging systems contribute to uncertainty in locating pinwheel centers (Polimeni et al. 2005). CO patches have vague boundaries, are not circular and often blend into each other. These factors make it difficult to define their centres. Finally, matching of optical imaging data with post-mortem CO histology is tricky because it involves comparison of a photograph of the brain surface with a section cut through the actual cortex. The curvature of the tissue usually changes in the transition from living brain to fixed specimen, requiring scrupulous alignment of pial blood vessels imaged in vivo with blood vessel profiles present in the histological section prepared later. Few studies have shown this correspondence, rendering it impossible for the reader to judge the reliability of the alignment and the data.
If CO patches and pinwheel centres are truly unrelated, then a conundrum emerges. The unoriented, centre-surround cells in layers 2 and 3 reported by Livingstone & Hubel (1984a) should give rise to untuned regions in optical imaging maps of orientation selectivity. These regions should correlate post-mortem with CO patches. If pinwheel centres and CO patches do not coincide, then where are Livingstone and Hubel's clusters of unoriented cells in optical imaging maps of primary visual cortex? Here, one can say only that no clear or consistent relationship exists between CO patches and orientation columns in striate cortex, and that it is impossible to say whether cells in CO patches are orientation selective. It is disheartening that such a fundamental question remains unsettled after nearly 20 years of collective labour in visual neuroscience. Perhaps the single-cell resolution afforded by optical maps obtained from in vivo two-photon imaging of calcium fluxes will provide a definitive answer (Ohki et al. 2005). One should certainly be prepared for a negative result, based on the lesson provided by the tree shrew. In this animal, the orientation columns are organized beautifully around pinwheel centres but CO patches are absent (Humphrey et al. 1980; Bosking et al. 1997). This result means that orientation columns and their pinwheel centres have evolved independently from the development of CO patches, at least in some species.
It seems obvious that cells in CO patches and interpatches should differ in their receptive field properties because their anatomical connections are quite different. As mentioned earlier, patches (but not interpatches) receive direct input from konio cells in the LGN (Hendry & Yoshioka 1994). Patches and interpatches partake in a different pattern of connections to CO stripes in V2 (Sincich & Horton 2002a). In addition, intrinsic direct afferents from layer 4 to the supragranular layers differ for patches and interpatches. Lachica et al. (1992) reported that patches in deep layer 3 receive input from layers 4A, 4B, 4Cα and 4Cβ, whereas interpatches receive input from only 4A and 4Cβ. In contrast, Yoshioka et al. (1994) found that patches receive projections from layers 4A and 4B, whereas interpatches receive input from just layer 4C. Although these accounts are discrepant, signalling a need for further studies, both research groups agree that patches and interpatches receive a different pattern of interlaminar projections. However, not all projections are dichotomized by CO patches/interpatches. Projections from striate cortex to the LGN (Horton 1984), superior colliculus (Lia & Olavarria 1996) and middle temporal area (MT or V5) (Sincich & Horton 2003b; but see Boyd & Casagrande 1999) arise randomly with respect to the CO patches.
Within striate cortex, intrinsic horizontal connections are columnar (Lund et al. 2003). Rockland & Lund (1983) first observed this property upon injecting horseradish peroxidase into the macaque striate cortex. Clumps of retrogradely filled cells and anterogradely labelled axons were situated around the injection site. For any given injection, the distribution of transported label matched from one layer to another, indicating columnar organization, although the spatial extent of horizontal projections varied by layer. Interestingly, the clumps of label did not coincide consistently with the location of CO patches. Subsequently, it has become evident that horizontal projections preferentially link similar compartments in striate cortex. For example, CO patches tend to project more strongly to other patches and interpatches with interpatches (Livingstone & Hubel 1984b; Yoshioka et al. 1996; Yabuta & Callaway 1998). In addition, cells with a given orientation tuning project preferentially to cells of the same orientation (figure 14; Gilbert & Wiesel 1989; Malach et al. 1993; Bosking et al. 1997; Sincich & Blasdel 2001). Therefore, the pattern of label surrounding an injection site and its relationship to CO patches depends on precisely where the tracer is deposited in the cortex.
Figure 14.

Patchy horizontal connections in the visual cortex. After biocytin injection into layers 2 and 3 of squirrel monkey striate cortex, clusters of axon terminals surround the injection site. Their distribution is elliptical, with the long axis aligned with the preferred orientation recorded at the injection site. Patches of terminals are sometimes visible in deeper layers. When present, they form columns by virtue of their registration with patches of terminals in layers 2 and 3 (Sincich & Blasdel 2001).
It remains unknown why intrinsic horizontal projections are stronger between similar compartments in striate cortex. The vertical registration of multiple patchy networks of tangential connections gives rise to a type of columnar structure, without boundaries, in the cortex. The relationship to fundamental processing modules, if indeed such modules exist in striate cortex, remains unclear. It is noteworthy that the squirrel, an animal that lacks orientation columns, ocular dominance columns and CO patches, still has a network of patchy horizontal connections within its visual cortex (Kaas et al. 1989). This underscores the lack of any common thread binding features of the functional architecture across species.
6. V2 cytochrome oxidase stripes
Even before periodic horizontal connections were identified within cortical areas, it was known that projections from one cortical area to another are often columnar (Jones et al. 1975; Goldman & Nauta 1977). In the visual system, a single injection of an anterograde tracer in V1 produces clumps of label in V2 (Martinez-Millán & Holländer 1975; Wong-Riley 1979). They are densest in layer 4, but extend vertically in a columnar fashion through layers 2 and 3. Their significance became clearer following studies of CO activity in area V2. The enzyme reveals a pattern of coarse parallel stripes stretching across V2 from the V1 border to the V3 border, organized in repeating cycles of pale-thick–pale-thin (Tootell et al. 1983; Horton 1984). The pale stripes coincide with the strongest columns of input from V1 (Sincich & Horton 2002b), whereas the dark stripes (both thick and thin) are targeted preferentially by the pulvinar (Levitt et al. 1995). These findings indicate that in area V2, just as in area V1, the columnar pattern of CO activity correlates with specific inputs and outputs. From V1, the patches project to thin stripes, whereas the interpatches supply thick and pale stripes (figure 15; Sincich & Horton 2002a). From V2, thin and pale stripes connect principally with area V4, while thick stripes innervate preferentially area MT (DeYoe & Van Essen 1985; Shipp & Zeki 1985; Zeki & Shipp 1989; DeYoe et al. 1994). However, in owl and squirrel monkeys, both sets of dark stripes—thin and thick—seem to contribute to the projection to area MT (Shipp & Zeki 1989; Krubitzer & Kaas 1990).
Figure 15.

Dovetailed, columnar projections from V1 and pulvinar to area V2. The projections from V1 to V2 are divided by cytochrome oxidase. Patches project to thin stripes and interpatches project to pale and thick stripes. The strongest V1 output is to pale stripes, whereas the pulvinar innervates more heavily thin and thick stripes. Controversy abounds regarding the receptive field properties of cells in the V2 stripes.
The distinct pattern of afferent and efferent connections made by the three types of V2 stripe leads irresistibly to the idea that they segregate cells into different functional streams. An early proposal equated colour with thin stripes, form with pale stripes and stereo/motion with thick stripes (Hubel & Livingstone 1987). Subsequently, numerous investigators have made electrode penetrations through V2, attempting to correlate single cell properties with V2 stripe type (Hubel & Livingstone 1987; Peterhans & von der Heydt 1993; Levitt et al. 1994; Roe & Ts'o 1995; Gegenfurtner et al. 1996; Tamura et al. 1996; Yoshioka & Dow 1996; Kiper et al. 1997; Ts'o et al. 2001a; Shipp & Zeki 2002). Collectively, the results from these studies have been largely inconclusive, although some receptive field properties appear to be more prevalent in cells populating certain stripe types. The lack of a clear verdict is reminiscent of the unresolved controversy surrounding the receptive field properties of cells in CO patches. It is easy to demonstrate columnar anatomical structure in the cortex, but functional correlation has often proved to be a far more difficult proposition.
In the macaque, only a thin sliver of V2 is exposed on the cortical surface, making it difficult to reconstruct CO stripes and to recover electrode tracks. Compounding the problem, the pattern of V2 stripes in macaques is often murky, making it impossible to differentiate thin and thick stripes. In contrast, the squirrel monkey consistently demonstrates a crystalline array of CO stripes, separable at a glance into pale, thick and thin. Area V2 is also situated on the cortical surface, making it easy to prepare histological sections. In the macaque, one must unfold the lunate sulcus and artificially flatten the cortex to reveal the CO stripes (Olavarria & Van Essen 1997). At one point, we thought that tissue manipulation during flat-mounting accounts for the poor labelling of CO stripes in some macaques. However, we have since learned that if a robust pattern of CO stripes is present, then it can survive the worst abuses during tissue processing (figure 16). This makes it probable that the variability in the expression of CO stripes in macaques is real. Inexplicably, macaques always have well developed ocular dominance columns but frequently lack crisply defined V2 stripes, whereas in squirrel monkeys, the opposite is true. What significance can be attached to the stripes in V2, if they are well developed in some individuals, and not in others, within a single species? Why are V2 stripes clearly tripartite in the squirrel monkey, but often not so in the macaque, marmoset, owl monkey, galago or human? In species (or members of a species) lacking clear V2 stripes, are the connections from V1 to V2, and from V2 to V4 and MT still neatly divided into separate compartments? These basic questions await definitive answers.
Figure 16.
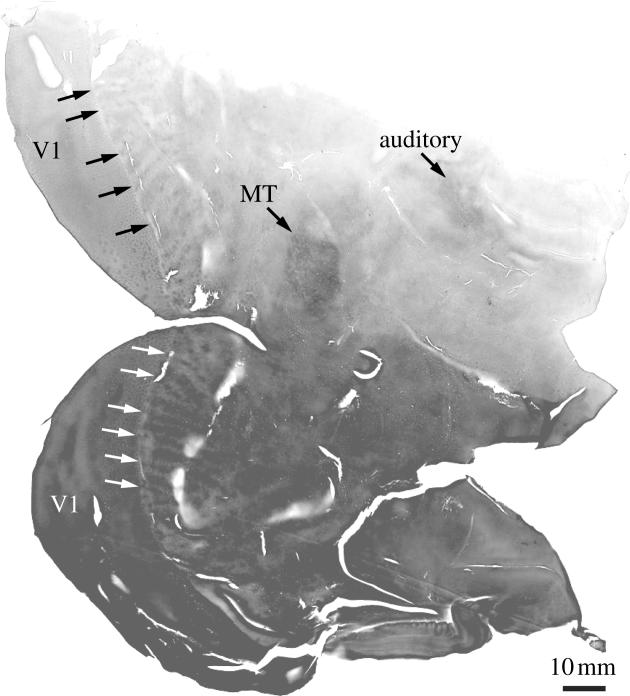
In V2 of many species, it is impossible to recognize two classes of CO stripes: thick and thin. In other species (e.g. squirrel monkeys), they can always be differentiated clearly. Remarkably, even within a species, expression of V2 stripes can be whimsical. In some macaques, thick and thin stripes are obvious, whereas in other macaques, they cannot be distinguished. Failure to visualize thin and thick stripes in some macaques is not an artefact of poor histology. This section shows an example of abysmal histology with uneven section thickness, chatter and tissue cracking, yet a tripartite pattern of CO stripes is still obvious in V2. After flatmounting and staining for CO in area V2 of more than 20 macaques, we have concluded that the variation in stripe expression is real.
7. Columns elsewhere in cortex
Many other cortical areas contain columns, defined as either clusters of cells sharing a common receptive field property or periodic projection fields arising from intra- or inter cortical axonal connections. In area V3, cells with similar disparity tuning are grouped together (Adams & Zeki 2001). Cells in V3 also make columnar projections to other visual areas (Felleman et al. 1997; Angelucci et al. 2002). In area MT, cells with the same directional preference are clustered (Albright et al. 1984), as are cells with the same disparity tuning (DeAngelis & Newsome 1999). Tracer injections in MT reveal patch-like clusters of intrinsic projections (Malach et al. 1997). The pattern of CO staining in MT is also patch-like, but no reliable correlation has been made with any receptive field property or anatomical projection (Born & Tootell 1992). In the inferotemporal cortex, single cell recordings and optical imaging have provided evidence for columns of cells about 400 μm wide, responsive to similar features of objects (Fujita et al. 1992; Wang et al. 1998). Given the nearly infinite variety of objects present in the environment, it is remarkable that specific columns could encode high-order stimulus features.
In the auditory cortex, tracer injections in the upper layers yield patches of label from intrinsic horizontal projections, similar to those seen in V1 and MT. These horizontal projections preferentially link regions with similar bandwidth along isofrequency domains (Read et al. 2001). Auditory cortex is also proposed to contain columns of cells that sum binaural input, interdigitated with columns that suppress binaural input (Imig & Adrian 1977). However, evidence has recently emerged suggesting that binaural suppression and summation may be organized by layer rather than by column (Reser et al. 2000). Commissural projections in cat A1 are periodic and may be related systematically to the proposed binaural map (Imig & Reale 1981). CO mapping of A1 shows intense activity, organized into four to five parallel bands (Tootell et al. 1985; Sincich et al. 2003). Unfortunately, these bands have not been correlated yet with any physiological property or pattern of anatomical connections. Language cortex in humans contains columnar, horizontal, long-range connections that are spaced 20% more widely in the left hemisphere (Galuske et al. 2000).
Columns have been encountered most frequently in sensory regions of the cortex, but they also occur in motor areas. For example, cells in macaque primary motor cortex with the same preferred direction vector are organized into clusters 200 μm apart, interleaved with clusters of cells preferring the orthogonal direction vector (Amirikian & Georgopoulos 2003).
Anterograde tracers injected into many different areas of macaque and squirrel monkey prefrontal cortex produce columns of label between 550 and 700 μm wide in ipsilateral cortical areas, and via the corpus callosum, in contralateral projection fields (Bugbee & Goldman-Rakic 1983). It is probable that patchy interareal projections occur throughout the cerebral cortex. The periodic distribution of projections throughout the cortex remains an enigma.
8. Cortical barrels
The barrels in somatosensory cortex representing the vibrissae are frequently cited as an archetypical example of columnar structure (figure 17). It is common to read introductory remarks to papers such as, ‘Cortical columns are the functional units of the neocortex that are particularly prominent in the “barrel” field of the somatosensory cortex’ (Lübke et al. 2000, p. 5300) or ‘Sensory cortex is ordered into columns, each tuned to a subset of peripheral stimuli. To identify the principles underlying the construction of columnar architecture, we monitored the development of circuits in the rat barrel cortex’ (Bureau et al. 2004, p. 789). Barrels are indisputably modular; in this sense they conform to Mountcastle's original definition more faithfully than other examples of columns, whose borders are nebulous. However, cortical barrels are not really columns. Rather, they are an exquisite example of somatotopic organization, analogous to the retinotopic map in primary visual cortex. Certainly, nobody would cite V1's retinotopic map as an example of columnar structure. It is only because each barrel happens to represent a single round structure on the animal's body, namely, a whisker follicle, that a barrel is endowed with a columnar form. In the case of the barrel, the circular structure and discrete boundary are trivial properties: they derive from the shape and discreteness of the peripheral entity that is being represented.
Figure 17.

A barrel is an isomorphic representation of a whisker, not a cortical column. (a) Flatmount section through the somatosensory cortex of a ground squirrel (Spermophilus beccheyi) processed for CO to show the barrelfield. (b) Masson trichrome-stained section through the vibrissae of a rat. A blood-filled sinus (arrow) surrounds each whisker, which may alter mechanical transduction of whisker motion.
Ironically, Lorento De Nó (1922) observed barrels in somatosensory cortex nearly half a century before they were discovered by Woolsey & Van Der Loos (1970). Lorento de Nó observed vertical septae in cross-sections of cortex, but had no idea that they corresponded to the representation of whiskers. Had Lorento de Nó cut a few sections parallel to the cortical surface, he surely would have recognized, as did Woolsey and Van der Loos, that he had stumbled upon the representation of rodent whiskers in a portion of somatosensory cortex. Thus, Lorento de Nó's elementary unit, which influenced Mountcastle so heavily, was derived from the observation of a structure unique to a specialized region of rodent cortex: the barrelfield. Unfortunately, Lorento de Nó's (1949) observation of the barrel led him ultimately to the general theory that the whole cortex is composed of cylindrical modules.
Another indication that barrels inherit the shape of the peripheral organs they represent, and are not a reflection of a fundamental structure unique to the cortex, comes from the finding of ‘barreloids’ in the ventrobasal thalamic nucleus. These bodies consist of clusters of thalamocortical projection neurons, each bound for a single barrel, separated by lighter-stained, cell-sparse septa (Land et al. 1995). They are organized in rows and columns, just like the cortical barrelfield. The precise barreloid-to-barrel projection makes it clear that the cortical barrelfield is just an isomorphic reiteration of the topography of the peripheral surface.
Following this reasoning, one would predict that any body appendage with a distinctive enough shape might be recognizable in the somatosensory cortex. The most spectacular example uncovered so far is the representation of the proboscis of the star-faced mole (Catania & Kaas 1995). Each of its 11 tentacles is visible as a stripe, separated by a CO-sparse septum, surrounding the representation of the nostril. Another example is provided by the cortical representation of the primate hand. Each digit appears as a dark stripe of myelin separated by a pale septum (Jain et al. 1998). It imparts little insight to refer to the somatosensory map of such body parts as cortical ‘columns’, or to extract any underlying principle beyond that of topographic representation of the body surface.
Body parts that have a highly magnified representation and that are physically separated along most of their surface from surrounding structures (e.g. digits, whiskers) are most likely to be recognizable as distinct structures in the somatosensory cortex. An enduring mystery is why a structure may appear visible in the somatosensory cortex of certain species but not others. For example, the vibrissae give rise to barrelfields in rats, mice, squirrels, rabbits, possums and porcupines, but not in raccoons, beavers or cats (Woolsey et al. 1975). Some animals, such as guinea pigs, do not engage in whisking behaviour, but nonetheless have a barrelfield. The lack of any correlation across species between barrelfields and whisker function recalls the dilemma of ocular dominance columns: a striking but inconstantly expressed anatomical feature that has no obvious relationship to function.
Mutant mice lacking barrelfields have normal cortical topography, as demonstrated by imaging deoxyglucose uptake after whisker stimulation. However, the arbors of individual thalamocortical neurons are altered, ramifying more widely in layer 4 (Cases et al. 1996; Welker et al. 1996). As a result, stronger responses can be recorded from manipulation of surround whiskers than in normal animals. However, the animals seem to display no functional deficit in whisking behaviour.
9. Development of columns
The development of cortical columns has been studied and modelled intensively (Katz & Shatz 1996), making it likely that their formation will be understood long before their purpose, if any. The prevailing view has been that patterns of neuronal activity guide the emergence of columnar structure. This idea derives principally from a series of experiments conducted by Stryker and colleagues, showing that silencing retinal ganglion cell function in kittens by intraocular injection of tetrodotoxin prevents the formation of ocular dominance columns (Stryker & Harris 1986). This result indicates that activity is required for column expression. Does the pattern of activity itself determine the physical structure of the columns, or does blockade of activity simply interfere with the expression of instructions that are encoded in some other form (e.g. genetically programmed secretion of neurotrophic factors)? Evidence bearing on this question has been provided by the celebrated three-eyed frog. Transplantation of an extra eye results in the production of columns in the optic tectum, which normally receives input solely from the contralateral eye (Constantine-Paton & Law 1978). Given that no instructions for column formation should be expected in the normal frog, the favoured interpretation has been that activity-dependent competition between afferents from the two eyes induces the segregation of columns. However, the possibility has not been excluded that instructions do in fact exist for column formation in the normal frog, and that these instructions are unleashed by engrafting a supplementary eye.
The most direct challenge to the theory that neuronal activity gives rise to columnar segregation has come from a study of the formation of ocular dominance columns in the ferret (Crowley & Katz 1999). Animals were binocularly enucleated at birth, before layer 4 of the cortex receives its first afferents. After the animals had matured, tracer injections were made into single LGN laminae. Surprisingly, these injections resulted in patches of label in the cortex that resembled ocular dominance columns (figure 18). The prediction, based on Stryker and Harris' tetrodotoxin experiments, was that loss of retinal activity should block column formation. One could argue that enucleation does not prevent spontaneous LGN activity, and that this activity might be sufficient to direct column formation. If so, why does such LGN activity not promote column formation after intraocular tetrodotoxin injection? The discrepant results obtained from eye enucleation in the ferret and tetrodotoxin injection in the cat are difficult to reconcile.
Figure 18.
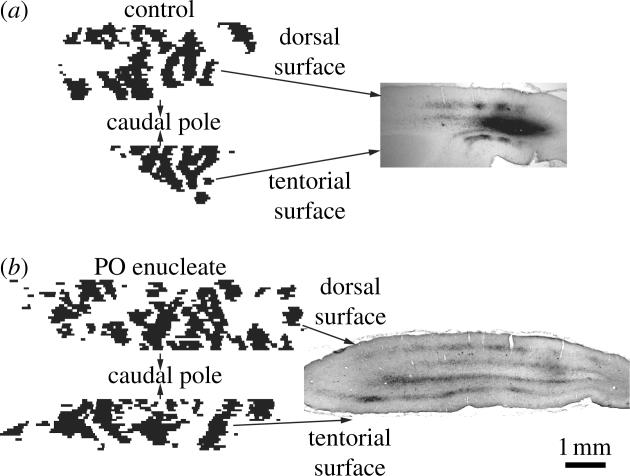
Ocular dominance columns form without eyes. (a) In control ferrets, biotinylated dextran amine was injected into the lateral geniculate nucleus, yielding ocular dominance columns in coronal sections through the visual cortex. Serial sections were aligned and drawn to provide a 2D view of the columns. (b) Enucleation was performed at birth, before geniculate afferents innervate the cortex. Despite this manipulation, ocular dominance columns were present after tracer injection into the lateral geniculate nucleus at 2 months of age (Crowley & Katz 1999).
In late foetal macaques and in week-old kittens, transneuronal labelling of geniculocortical afferents after eye injection with [3H]proline shows diffuse label in layer 4C (Rakic 1977; LeVay et al. 1978; Crair et al. 2001). Following an initial period of diffuse ingrowth of geniculate afferents, formation of ocular dominance columns is believed to occur via a selective process of proliferation of synaptic contacts located within the eye's future columns and retraction of branches located in the other eye's columns. This mechanism has been inferred, however, based on observations made in different animals at different ages. It would be far more powerful to demonstrate by direct serial imaging of geniculocortical afferents in the same animal that column formation occurs by this sequence of events. Crowley & Katz (2000) have provided evidence from ferrets that columns begin to form soon after geniculate afferents reach layer 4, leaving little role for later activity-dependent pruning. Further studies are required to determine if columns form by molecular targeting of geniculocortical afferents to designated locales, or through activity-dependent mechanisms.
In mammals, spontaneous waves of ganglion cell activity sweep periodically across the developing retina (Meister et al. 1991). These waves are transmitted to the LGN and, hence, reflected in the firing patterns of geniculocortical afferents. It has been proposed that retinal waves are well suited to drive the early formation of ocular dominance columns (Mooney et al. 1996). The idea is that correlated activation of one eye's geniculate afferents might reinforce connections onto single cortical cells, promoting the emergence of monocular units in the cortex. However, it is unclear why retinal waves should promote the aggregation of individual monocular cells into columns. For this to occur, the waves would have to sweep repeatedly across the retina in an identical pattern for days. Otherwise, the small effect produced by a single wave would not be sustained, and it would soon be offset moments later by a wave in the other eye. Unless the retinal waves were choreographed, one should not expect them to produce a stereotypic pattern of columns in the cortex.
The pattern formed by the ocular dominance columns in each hemisphere is extremely similar (figure 10). This symmetry poses another major obstacle for the wave hypothesis because it requires coordination of the waves in the two eyes. To produce similar patterns of columns in each striate cortex, a wave in the temporal retina of one eye must occur while a similar wave is occurring in the nasal retina of the other eye (figure 19). Alternatively, wave patterns must be mirror-imaged across the vertical meridian of each retina. Otherwise, without such coordination of the waves in each retina, the column patterns in each hemisphere would show no symmetry. It is difficult to see how waves occurring on a random basis in each eye's retina could give rise to an intricate, matching pattern of columns in the left and right visual cortex. Further doubt has been cast on the importance of spontaneous retinal waves in the segregation of left and right eye afferents by evidence that immunotoxin depletion of starburst amacrine cells (the source of retinal waves) does not interfere with the normal formation of LGN laminae (Huberman et al. 2003).
Figure 19.

Random retinal activity waves cannot generate the patterns formed by ocular dominance columns. Despite variability in column expression among squirrel monkeys, the patterns in the left and right hemispheres are always mirror-image symmetric (see figure 10). Their formation from retinal patterns of neuronal activity would require spontaneous waves (here, scaled from ferret Feller et al. 1996 to human retina) that occur in a symmetric fashion across the vertical meridian (dashed line) of one eye. Alternatively, symmetric column patterns could result if the waves in the temporal retina of one eye and the nasal retina of the other eye were coordinated for all visual field loci. Because the waves are random and independent between the two eyes, neither requirement is feasible.
In some squirrel monkeys, focal deprivation of photoreceptors by retinal blood vessels causes rearrangement of geniculate afferents in striate cortex (Adams & Horton 2002). Cortical territory representing visual field obscured by a retinal vessel is relinquished in favour of the other eye (figure 20). After monocular enucleation in the adult animal, a pattern in striate cortex corresponding to the retinal vascular tree is revealed by CO staining. This phenomenon is analogous, on a local scale, to the wholesale shrinkage of columns reported after monocular eyelid suture (Hubel et al. 1976). The interesting point is that in monkeys with fine ocular dominance columns, each angioscotoma representation in the cortex is fringed by cortex belonging exclusively to the other eye (figure 20a). Why should this ‘frosting’ appear, given that neither eye enjoys a competitive advantage in cortex adjacent to an angioscotoma representation? Perhaps an exchange of cortical territory occurs during early development. One eye takes over a continuous channel of cortical territory, because the other eye's view is obstructed by a retinal blood vessel. In return, it abandons all territory along a strip flanking either side of the angioscotoma representation because an innate cortical mechanism exists that parcels territory in a reciprocal fashion. In animals with essentially no columns, where this innate cortical mechanism is absent, no frosting is seen around angioscotoma representations (figure 20b; Adams & Horton 2003b).
Figure 20.
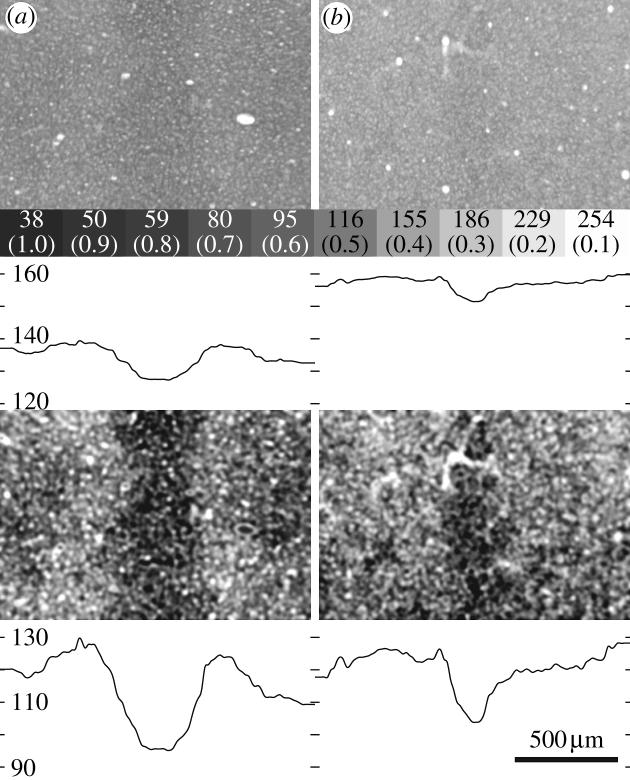
A reciprocal exchange of territory accompanies the formation of angioscotoma representations in animals with ocular dominance columns, but does not occur in those lacking columns. (a) Boxed region from figure 9, showing a dark angioscotoma from an animal with fine columns. It is hardly visible in the raw image at this scale, emphasizing that CO patterns in tissue sections often reflect only a small difference in optical density. The optical density plotted below shows the mean value of each pixel column. Numbers refer to level of grey (0–255) and parentheses indicate optical density of neutral density filters used for calibration (Horton & Hocking 1998). Frame below shows the same image, normalized to full grey scale range, to make the angioscotoma more readily visible. Note the pale zones flanking the angioscotoma, reflected in the inverted Mexican hat profile of the optical density plot. These surrounding zones of opposite contrast are served exclusively by the other eye, whose geniculocortical afferents have been driven out of the cortex corresponding to the angioscotoma representation. (b) A dark angioscotoma representation, from an animal with essentially no ocular dominance columns, without flanking zones dedicated entirely to the other eye (Adams & Horton 2002).
These observations suggest that columnar organization may depend on a cortical mechanism, to date undefined, which involves some form of mutual repellency (Miller et al. 1989). Variation in the strength of this mechanism from animal to animal and from species to species could account for the variability of column architecture found among normal subjects (Horton & Hocking 1996b; Adams & Horton 2003a).
In the cat, the spacing of orientation columns shows remarkable similarity among littermates raised in the same visual environment (Kaschube et al. 2002). For ocular dominance columns, the influence of kinship is less clear (Kaschube et al. 2003). Nonetheless, these observations are important because they provide the first evidence that variation in columnar structure (Horton & Hocking 1996b) may be a heritable feature of the functional architecture of the cortex.
10. Conclusions
The cerebral cortex is an enticing structure of staggering complexity, although it appears as nothing more than a drab sheet of tissue to the naked eye. For the scientist grappling with the challenge of explaining its function, an obvious strategy is to search for underlying principles that govern its organization. The cortical column has been a popular concept for precisely this reason. If one could understand a little piece of the cortex, and if this piece were representative of the whole, then our task would be simplified immensely. Development of the cortex would also be easier to explain: a matter of unravelling the instructions for a single module, and then learning how this module is iterated a few hundred thousand times to cover the surface of the human brain.
Fundamental discoveries, like the action potential or the synaptic vesicle, provided new insight into brain function. It thus became clearer how neurons communicate with each other. Although the column has been offered as the fundamental unit of the cortex, it has not earned this lofty designation. After half a century, it is still unclear what precisely is meant by the term. It does not correspond to any single structure within the cortex. It has been impossible to find a canonical microcircuit that corresponds to the cortical column (Nelson 2002; Douglas & Martin 2004). No genetic mechanism has been deciphered that designates how to construct a column.
At this point, two general properties have materialized that are thought to embody if not the cortical column, then at least the concept of columnar structure. First, cells that share receptive field characteristics are often clustered together. Second, cellular connections within and between areas are usually periodic. These are tantalizing clues but their significance for cortical processing remains unknown. It is telling that over decades, dozens of papers have been written about columns yet no answers have emerged. They have provided grist for theoreticians and spawned innumerable models, but no tangible progress has been made. Only a few commentators have been bold enough to state directly that perhaps columns are not an important feature of brain structure (Swindale 1990; Purves et al. 1992).
We stress that we are not denying the existence of columns, denigrating the impact of their discovery or questioning the utility of studying their properties. As an epiphenomenon, columns have been valuable for probing the development and organization of the cortex. For example, courtesy of columns, we have learned how the cortex responds to manipulation of the sensory environment during the critical period. The spectacular images of geniculocortical afferents shrunken by monocular deprivation owe their existence to ocular dominance columns (Hubel et al. 1976). Their shrinkage was revealed by simply injecting the deprived eye with a transneuronal anterograde tracer. Had geniculate afferents serving each eye been distributed diffusely within the cortex, this classic experiment would have been impossible. Having shown how columns are affected by deprivation, it is natural to assume that the columns themselves have a function. The discovery that geniculate afferents are affected by deprivation was fundamental; the fact that they are aggregated into columns may be trivial.
Species variation in columnar structure is hard to reconcile with ideas about the functional importance of columns. It is problematic when a system such as ocular dominance columns is present in some species but not others, without any obvious functional correlate. It is even worse when some members of a single species have ocular dominance columns but not others, for no apparent reason. The ultimate blow comes with the realization that some members of a single species have ocular dominance columns in only part of the visual cortex, leaving other regions of binocular cortex bereft of columns (Adams & Horton 2003a). At some point, one must abandon the idea that columns are the basic functional entity of the cortex. It now seems doubtful that any single, transcendent principle endows the cerebral cortex with a modular structure. Each individual area is constructed differently, and each will need to be taken apart cell by cell, layer by layer, circuit by circuit and projection by projection to describe fully the architecture of the cortex.
Laboratories work with many different animal species in the hope that each will teach lessons about the function of the human brain. Comparative anatomy provides a glimpse at brain structure during only one moment in evolution. We know little about how brains evolved to take their present form. It is apparent that considerable natural variation occurs among members of a species in normal brain structure. Nature, being a tinkerer (Jacob 1977), probably uses this variation as a substrate for brain evolution. Columnar structures that have no function in some species may acquire a function in others through evolution. This may explain why, in many instances, there seems to be no logic or consistency in Nature's design.
Gould (1997) has used the term ‘spandrel’ to describe structures that arise non adaptively (i.e. as an epiphenomenon) and then later in some species become harnessed secondarily for some purpose. In architecture, a spandrel is the triangular space left when an arch cuts through the rectangular framework that surrounds it. In some medieval churches, the spandrels are decorated so beautifully that one might imagine that they were created for religious displays. In fact, they are nothing more than an obligatory by-product of the architectural decision to build an archway. As we examine the brains of different species, it is possible that the column structures we encounter represent spandrels in various stages of evolution. This could account for the fact that in macaques, the ocular dominance columns are aligned with patches, whereas in squirrel monkeys, the ocular dominance columns (when present) are independent.
Acknowledgments
Supported by grants from the Larry L. Hillblom Foundation and the National Eye Institute. Lawrence Sincich, Stewart Shipp and Vernon Mountcastle provided helpful comments on the manuscript.
Glossary
- CO
cytochrome oxidase
- LGN
lateral geniculate nucleus
- MT
middle temporal area
Footnotes
One contribution of 12 to a Theme Issue ‘Cerebral cartography 1905–2005’.
References
- Adams D.L, Horton J.C. Shadows cast by retinal blood vessels mapped in primary visual cortex. Science. 2002;298:572–576. doi: 10.1126/science.1074887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adams D.L, Horton J.C. Capricious expression of cortical columns in the primate brain. Nat. Neurosci. 2003a;6:113–114. doi: 10.1038/nn1004. [DOI] [PubMed] [Google Scholar]
- Adams D.L, Horton J.C. The representation of retinal blood vessels in primate striate cortex. J. Neurosci. 2003b;23:5984–5997. doi: 10.1523/JNEUROSCI.23-14-05984.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adams D.L, Zeki S. Functional organization of macaque V3 for stereoscopic depth. J. Neurophysiol. 2001;86:2195–2203. doi: 10.1152/jn.2001.86.5.2195. [DOI] [PubMed] [Google Scholar]
- Albright T.D, Desimone R, Gross C.G. Columnar organization of directionally selective cells in visual area MT of the macaque. J. Neurophysiol. 1984;51:16–31. doi: 10.1152/jn.1984.51.1.16. [DOI] [PubMed] [Google Scholar]
- Albus K. A quantitative study of the projection area of the central and the paracentral visual field in area 17 of the cat. II. The spatial organization of the orientation domain. Exp. Brain Res. 1975;24:181–202. doi: 10.1007/BF00234062. [DOI] [PubMed] [Google Scholar]
- Amirikian B, Georgopoulos A.P. Modular organization of directionally tuned cells in the motor cortex: is there a short-range order? Proc. Natl Acad. Sci. USA. 2003;100:12 474–12 479. doi: 10.1073/pnas.2037719100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angelucci A, Levitt J.B, Walton E.J, Hupe J.M, Bullier J, Lund J.S. Circuits for local and global signal integration in primary visual cortex. J. Neurosci. 2002;22:8633–8646. doi: 10.1523/JNEUROSCI.22-19-08633.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angevine J, Sidman R. Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature. 1961;192:766–768. doi: 10.1038/192766b0. [DOI] [PubMed] [Google Scholar]
- Anthony T.E, Klein C, Fishell G, Heintz N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 2004;41:881–890. doi: 10.1016/s0896-6273(04)00140-0. [DOI] [PubMed] [Google Scholar]
- Bartfeld E, Grinvald A. Relationships between orientation-preference pinwheels, cytochrome oxidase blobs, and ocular-dominance columns in primate striate cortex. Proc. Natl Acad. Sci. USA. 1992;89:11 905–11 909. doi: 10.1073/pnas.89.24.11905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basole A, White L.E, Fitzpatrick D. Mapping multiple features in the population response of visual cortex. Nature. 2003;423:986–990. doi: 10.1038/nature01721. [DOI] [PubMed] [Google Scholar]
- Bear M.F, Rittenhouse C.D. Molecular basis for induction of ocular dominance plasticity. J. Neurobiol. 1999;41:83–91. doi: 10.1002/(sici)1097-4695(199910)41:1<83::aid-neu11>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- Blasdel G.G. Differential imaging of ocular dominance and orientation selectivity in monkey striate cortex. J. Neurosci. 1992a;12:3115–3138. doi: 10.1523/JNEUROSCI.12-08-03115.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blasdel G.G. Orientation selectivity, preference, and continuity in monkey striate cortex. J. Neurosci. 1992b;12:3139–3161. doi: 10.1523/JNEUROSCI.12-08-03139.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blasdel G, Campbell D. Functional retinotopy of monkey visual cortex. J. Neurosci. 2001;21:8286–8301. doi: 10.1523/JNEUROSCI.21-20-08286.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blasdel G.G, Salama G. Voltage-sensitive dyes reveal a modular organization in monkey striate cortex. Nature. 1986;321:579–585. doi: 10.1038/321579a0. [DOI] [PubMed] [Google Scholar]
- Bonhoeffer T, Grinvald A. Iso-orientation domains in cat visual cortex are arranged in pinwheel-like patterns. Nature. 1991;353:429–431. doi: 10.1038/353429a0. [DOI] [PubMed] [Google Scholar]
- Born, R. T. & Tootell, R. B. 1992 Segregation of global and local motion processing in primate middle temporal visual area. Nature357, 497–499. [DOI] [PubMed]
- Bosking W.H, Zhang Y, Schofield B, Fitzpatrick D. Orientation selectivity and the arrangement of horizontal connections in tree shrew striate cortex. J. Neurosci. 1997;17:2112–2127. doi: 10.1523/JNEUROSCI.17-06-02112.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyd J.D, Casagrande V.A. Relationships between cytochrome oxidase (CO) blobs in primate primary visual cortex (V1) and the distribution of neurons projecting to the middle temporal area (MT) J. Comp. Neurol. 1999;409:573–591. doi: 10.1002/(sici)1096-9861(19990712)409:4<573::aid-cne5>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- Boyd J.D, Matsubara J.A. Laminar and columnar patterns of geniculocortical projections in the cat: relationship to cytochrome oxidase. J. Comp. Neurol. 1996;365:659–682. doi: 10.1002/(SICI)1096-9861(19960219)365:4<659::AID-CNE11>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- Bugbee N.M, Goldman-Rakic P.S. Columnar organization of corticocortical projections in squirrel and rhesus monkeys: similarity of column width in species differing in cortical volume. J. Comp. Neurol. 1983;220:355–364. doi: 10.1002/cne.902200309. [DOI] [PubMed] [Google Scholar]
- Bureau I, Shepherd G.M, Svoboda K. Precise development of functional and anatomical columns in the neocortex. Neuron. 2004;42:789–801. doi: 10.1016/j.neuron.2004.05.002. [DOI] [PubMed] [Google Scholar]
- Buxhoeveden D.P, Casanova M.F. The minicolumn hypothesis in neuroscience. Brain. 2002;125:935–951. doi: 10.1093/brain/awf110. [DOI] [PubMed] [Google Scholar]
- Callaway E.M. Local circuits in primary visual cortex of the macaque monkey. Annu. Rev. Neurosci. 1998;21:47–74. doi: 10.1146/annurev.neuro.21.1.47. [DOI] [PubMed] [Google Scholar]
- Carder R.K. Immunocytochemical characterization of AMPA-selective glutamate receptor subunits: laminar and compartmental distribution in macaque striate cortex. J. Neurosci. 1997;17:3352–3363. doi: 10.1523/JNEUROSCI.17-09-03352.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carder R.K, Hendry S.H. Neuronal characterization, compartmental distribution, and activity-dependent regulation of glutamate immunoreactivity in adult monkey striate cortex. J. Neurosci. 1994;14:242–262. doi: 10.1523/JNEUROSCI.14-01-00242.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casagrande V.A, Harting J.K. Transneuronal transport of tritiated fucose and proline in the visual pathways of tree shrew Tupaia glis. Brain Res. 1975;96:367–372. doi: 10.1016/0006-8993(75)90749-0. [DOI] [PubMed] [Google Scholar]
- Cases O, Vitalis T, Seif I, De Maeyer E, Sotelo C, Gaspar P. Lack of barrels in the somatosensory cortex of monoamine oxidase A-deficient mice: role of a serotonin excess during the critical period. Neuron. 1996;16:297–307. doi: 10.1016/s0896-6273(00)80048-3. [DOI] [PubMed] [Google Scholar]
- Catania K.C, Kaas J.H. Organization of the somatosensory cortex of the star-nosed mole. J. Comp. Neurol. 1995;351:549–567. doi: 10.1002/cne.903510406. [DOI] [PubMed] [Google Scholar]
- Celio M.R, Scharer L, Morrison J.H, Norman A.W, Bloom F.E. Calbindin immunoreactivity alternates with cytochrome c-oxidase-rich zones in some layers of the primate visual cortex. Nature. 1986;323:715–717. doi: 10.1038/323715a0. [DOI] [PubMed] [Google Scholar]
- Chatterjee S, Callaway E.M. Parallel colour-opponent pathways to primary visual cortex. Nature. 2003;426:668–671. doi: 10.1038/nature02167. [DOI] [PubMed] [Google Scholar]
- Chaudhuri A, Cynader M.S. Activity-dependent expression of the transcription factor Zif268 reveals ocular dominance columns in monkey visual cortex. Brain Res. 1993;605:349–353. doi: 10.1016/0006-8993(93)91765-k. [DOI] [PubMed] [Google Scholar]
- Chow K.L, Masland R.H, Stewart D.L. Receptive field characteristics of striate cortical neurons in the rabbit. Brain Res. 1971;33:337–352. doi: 10.1016/0006-8993(71)90107-7. [DOI] [PubMed] [Google Scholar]
- Constantine-Paton M, Law M.I. Eye-specific termination bands in tecta of three-eyed frogs. Science. 1978;202:639–641. doi: 10.1126/science.309179. [DOI] [PubMed] [Google Scholar]
- Crair M.C, Ruthazer E.S, Gillespie D.S, Stryker M.P. Ocular dominance peaks at pinwheel center singularities of the orientation map in cat visual cortex. J. Neurophysiol. 1997;77:3381–3385. doi: 10.1152/jn.1997.77.6.3381. [DOI] [PubMed] [Google Scholar]
- Crair M.C, Horton J.C, Antonini A, Stryker M.P. Emergence of ocular dominance columns in cat visual cortex by 2 weeks of age. J. Comp. Neurol. 2001;430:235–249. doi: 10.1002/1096-9861(20010205)430:2<235::aid-cne1028>3.0.co;2-p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crowley J.C, Katz L.C. Development of ocular dominance columns in the absence of retinal input. Nat. Neurosci. 1999;2:1125–1130. doi: 10.1038/16051. [DOI] [PubMed] [Google Scholar]
- Crowley J.C, Katz L.C. Early development of ocular dominance columns. Science. 2000;290:1321–1324. doi: 10.1126/science.290.5495.1321. [DOI] [PubMed] [Google Scholar]
- DeAngelis G.C, Newsome W.T. Organization of disparity-selective neurons in macaque area MT. J. Neurosci. 1999;19:1398–1415. doi: 10.1523/JNEUROSCI.19-04-01398.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeAngelis G.C, Ghose G.M, Ohzawa I, Freeman R.D. Functional micro-organization of primary visual cortex: receptive field analysis of nearby neurons. J. Neurosci. 1999;19:4046–4064. doi: 10.1523/JNEUROSCI.19-10-04046.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeBruyn E.J, Casagrande V.A, Beck P.D, Bonds A.B. Visual resolution and sensitivity of single cells in the primary visual cortex (V1) of a nocturnal primate (Bush Baby): correlations with cortical layers and cytochrome oxidase patterns. J. Neurophysiol. 1993;69:3–18. doi: 10.1152/jn.1993.69.1.3. [DOI] [PubMed] [Google Scholar]
- DeYoe E.A, Van Essen D.C. Segregation of efferent connections and receptive field properties in visual area V2 of the macaque. Nature. 1985;317:58–61. doi: 10.1038/317058a0. [DOI] [PubMed] [Google Scholar]
- DeYoe E.A, Felleman D.J, Van Essen D.C, McClendon E. Multiple processing streams in occipitotemporal visual cortex. Nature. 1994;371:151–154. doi: 10.1038/371151a0. [DOI] [PubMed] [Google Scholar]
- Douglas R, Martin K. Neuronal circuits of the neocortex. Annu. Rev. Neurosci. 2004;27:419–451. doi: 10.1146/annurev.neuro.27.070203.144152. [DOI] [PubMed] [Google Scholar]
- Drager U.C. Receptive fields of single cells and topography in mouse visual cortex. J. Comp. Neurol. 1975;160:269–290. doi: 10.1002/cne.901600302. [DOI] [PubMed] [Google Scholar]
- Duffy K.R, Livingstone M.S. Distribution of non-phosphorylated neurofilament in squirrel monkey V1 is complementary to the pattern of cytochrome-oxidase blobs. Cereb. Cortex. 2003;13:722–727. doi: 10.1093/cercor/13.7.722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dyck R.H, Chaudhuri A, Cynader M.S. Experience-dependent regulation of the zincergic innervation of visual cortex in adult monkeys. Cereb. Cortex. 2003;13:1094–1109. doi: 10.1093/cercor/13.10.1094. [DOI] [PubMed] [Google Scholar]
- Everson R.M, Prashanth A.K, Gabbay M, Knight B.W, Sirovich L, Kaplan E. Representation of spatial frequency and orientation in the visual cortex. Proc. Natl Acad. Sci. USA. 1998;95:8334–8338. doi: 10.1073/pnas.95.14.8334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Favorov O.V, Diamond M.E, Whitsel B.L. Evidence for a mosaic representation of the body surface in area 3b of the somatic cortex of cat. Proc. Natl Acad. Sci. USA. 1987;84:6606–6610. doi: 10.1073/pnas.84.18.6606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felleman D.J, Burkhalter A, Van Essen D.C. Cortical connections of areas V3 and VP of macaque monkey extrastriate visual cortex. J. Comp. Neurol. 1997;379:21–47. doi: 10.1002/(sici)1096-9861(19970303)379:1<21::aid-cne3>3.0.co;2-k. [DOI] [PubMed] [Google Scholar]
- Feller M.B, Wellis D.P, Stellwagen D, Werblin F.S, Shatz C.J. Requirement for cholinergic synaptic transmission in the propagation of spontaneous retinal waves. Science. 1996;272:1182–1187. doi: 10.1126/science.272.5265.1182. [DOI] [PubMed] [Google Scholar]
- Ferster D. A comparison of binocular depth mechanisms in areas 17 and 18 of the cat visual cortex. J. Physiol. (Lond.) 1981;311:623–655. doi: 10.1113/jphysiol.1981.sp013608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonta C, Chappert C, Imbert M. N-Methyl-d-aspartate subunit R1 involvement in the postnatal organization of the primary visual cortex of Callithrix jacchus. J. Comp. Neurol. 1997;386:260–276. doi: 10.1002/(sici)1096-9861(19970922)386:2<260::aid-cne7>3.0.co;2-#. [DOI] [PubMed] [Google Scholar]
- Fonta C, Chappert C, Imbert M. Effect of monocular deprivation on NMDAR1 immunostaining in ocular dominance columns of the marmoset Callithrix jacchus. Vis. Neurosci. 2000;17:345–352. doi: 10.1017/s0952523800173031. [DOI] [PubMed] [Google Scholar]
- Fujita I, Tanaka K, Ito M, Cheng K. Columns for visual features of objects in monkey inferotemporal cortex. Nature. 1992;360:343–346. doi: 10.1038/360343a0. [DOI] [PubMed] [Google Scholar]
- Galuske R.A, Schlote W, Bratzke H, Singer W. Interhemispheric asymmetries of the modular structure in human temporal cortex. Science. 2000;289:1946–1949. doi: 10.1126/science.289.5486.1946. [DOI] [PubMed] [Google Scholar]
- Gardner J.C, Raiten E.J. Ocular dominance and disparity-sensitivity: why there are cells in the visual cortex driven unequally by the two eyes. Exp. Brain Res. 1986;64:505–514. doi: 10.1007/BF00340488. [DOI] [PubMed] [Google Scholar]
- Gegenfurtner K.R, Kiper D.C, Fenstemaker S.B. Processing of color, form, and motion in macaque area V2. Vis. Neurosci. 1996;13:161–172. doi: 10.1017/s0952523800007203. [DOI] [PubMed] [Google Scholar]
- Gilbert C.D. Laminar differences in receptive field properties of cells in cat primary visual cortex. J. Physiol. (Lond.) 1977;268:391–421. doi: 10.1113/jphysiol.1977.sp011863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert C.D, Wiesel T.N. Columnar specificity of intrinsic horizontal and corticocortical connections in cat visual cortex. J. Neurosci. 1989;9:2432–2442. doi: 10.1523/JNEUROSCI.09-07-02432.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Girman S.V, Sauve Y, Lund R.D. Receptive field properties of single neurons in rat primary visual cortex. J. Neurophysiol. 1999;82:301–311. doi: 10.1152/jn.1999.82.1.301. [DOI] [PubMed] [Google Scholar]
- Goldman P.S, Nauta W.J. Columnar distribution of cortico-cortical fibers in the frontal association, limbic, and motor cortex of the developing rhesus monkey. Brain Res. 1977;122:393–413. doi: 10.1016/0006-8993(77)90453-x. [DOI] [PubMed] [Google Scholar]
- Goto S, Singer W. Laminar and columnar organization of immunoreactivity for calcineurin, a calcium- and calmodulin-regulated protein phosphatase, in monkey striate cortex. Cereb. Cortex. 1994;4:636–645. doi: 10.1093/cercor/4.6.636. [DOI] [PubMed] [Google Scholar]
- Gould S.J. The exaptive excellence of spandrels as a term and prototype. Proc. Natl Acad. Sci. USA. 1997;94:10 750–10 755. doi: 10.1073/pnas.94.20.10750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haseltine E.C, DeBruyn E.J, Casagrande V.A. Demonstration of ocular dominance columns in Nissl-stained sections of monkey visual cortex following enucleation. Brain Res. 1979;176:153–158. doi: 10.1016/0006-8993(79)90878-3. [DOI] [PubMed] [Google Scholar]
- Hendrickson A.E, Wilson J.R, Ogren M.P. The neuroanatomical organization of pathways between the dorsal lateral geniculate nucleus and visual cortex in Old World and New World primates. J. Comp. Neurol. 1978;182:123–136. doi: 10.1002/cne.901820108. [DOI] [PubMed] [Google Scholar]
- Hendrickson A.E, Hunt S.P, Wu J.Y. Immunocytochemical localization of glutamic acid decarboxylase in monkey striate cortex. Nature. 1981;292:605–607. doi: 10.1038/292605a0. [DOI] [PubMed] [Google Scholar]
- Hendry S.H, Bhandari M.A. Neuronal organization and plasticity in adult monkey visual cortex: immunoreactivity for microtubule-associated protein 2. Vis. Neurosci. 1992;9:445–459. doi: 10.1017/s0952523800011251. [DOI] [PubMed] [Google Scholar]
- Hendry S.H, Calkins D.J. Neuronal chemistry and functional organization in the primate visual system. Trends Neurosci. 1998;21:344–349. doi: 10.1016/s0166-2236(98)01245-4. [DOI] [PubMed] [Google Scholar]
- Hendry S.H, Yoshioka T. A neurochemically distinct third channel in the macaque dorsal lateral geniculate nucleus. Science. 1994;264:575–577. doi: 10.1126/science.8160015. [DOI] [PubMed] [Google Scholar]
- Hendry S.H, Hockfield S, Jones E.G, McKay R. Monoclonal antibody that identifies subsets of neurones in the central visual system of monkey and cat. Nature. 1984;307:267–269. doi: 10.1038/307267a0. [DOI] [PubMed] [Google Scholar]
- Hendry S.H, Huntsman M.M, Viñuela A, Möhler H, de Blas A.L, Jones E.G. GABAA receptor subunit immunoreactivity in primate visual cortex: distribution in macaques and humans and regulation by visual input in adulthood. J. Neurosci. 1994;14:2383–2401. doi: 10.1523/JNEUROSCI.14-04-02383.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hetherington P.A, Swindale N.V. Receptive field and orientation scatter studied by tetrode recordings in cat area 17. Vis. Neurosci. 1999;16:637–652. doi: 10.1017/s0952523899164046. [DOI] [PubMed] [Google Scholar]
- Horton J.C. Cytochrome oxidase patches: a new cytoarchitectonic feature of monkey visual cortex. Phil. Trans. R. Soc. B. 1984;304:199–253. doi: 10.1098/rstb.1984.0021. [DOI] [PubMed] [Google Scholar]
- Horton J.C, Hedley-Whyte E.T. Mapping of cytochrome oxidase patches and ocular dominance columns in human visual cortex. Phil. Trans. R. Soc. B. 1984;304:255–272. doi: 10.1098/rstb.1984.0022. [DOI] [PubMed] [Google Scholar]
- Horton J.C, Hocking D.R. Anatomical demonstration of ocular dominance columns in striate cortex of the squirrel monkey. J. Neurosci. 1996a;16:5510–5522. doi: 10.1523/JNEUROSCI.16-17-05510.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horton J.C, Hocking D.R. Intrinsic variability of ocular dominance column periodicity in normal macaque monkeys. J. Neurosci. 1996b;16:7228–7239. doi: 10.1523/JNEUROSCI.16-22-07228.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horton J.C, Hocking D.R. Monocular core zones and binocular border strips in primate striate cortex revealed by the contrasting effects of enucleation, eyelid suture, and retinal laser lesions on cytochrome oxidase activity. J. Neurosci. 1998;18:5433–5455. doi: 10.1523/JNEUROSCI.18-14-05433.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horton J.C, Hubel D.H. Regular patchy distribution of cytochrome oxidase staining in primary visual cortex of macaque monkey. Nature. 1981;292:762–764. doi: 10.1038/292762a0. [DOI] [PubMed] [Google Scholar]
- Horton J.C, Hocking D.R, Adams D.L. Rapid identification of ocular dominance columns in macaques using cytochrome oxidase, Zif268, and dark-field microscopy. Vis. Neurosci. 2000;17:495–508. doi: 10.1017/s0952523800174024. [DOI] [PubMed] [Google Scholar]
- Hubel D.H. An autoradiographic study of the retino-cortical projections in the tree shrew (Tupaia glis) Brain Res. 1975;96:41–50. doi: 10.1016/0006-8993(75)90568-5. [DOI] [PubMed] [Google Scholar]
- Hubel D.H. Exploration of the primary visual cortex, 1955–78. Nature. 1982;299:515–524. doi: 10.1038/299515a0. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, Livingstone M.S. Segregation of form, color, and stereopsis in primate area 18. J. Neurosci. 1987;7:3378–3415. doi: 10.1523/JNEUROSCI.07-11-03378.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Receptive fields, binocular interaction and functional architecture in the cat's visual cortex. J. Physiol. (Lond.) 1962;160:106–154. doi: 10.1113/jphysiol.1962.sp006837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Shape and arrangement of columns in cat's striate cortex. J. Physiol. (Lond.) 1963;165:559–568. doi: 10.1113/jphysiol.1963.sp007079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Receptive fields and functional architecture of monkey striate cortex. J. Physiol. (Lond.) 1968;195:215–243. doi: 10.1113/jphysiol.1968.sp008455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Anatomical demonstration of columns in the monkey striate cortex. Nature. 1969;221:747–750. doi: 10.1038/221747a0. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Laminar and columnar distribution of geniculo-cortical fibers in the macaque monkey. J. Comp. Neurol. 1972;146:421–450. doi: 10.1002/cne.901460402. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Sequence regularity and geometry of orientation columns in the monkey striate cortex. J. Comp. Neurol. 1974a;158:267–293. doi: 10.1002/cne.901580304. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Uniformity of monkey striate cortex: a parallel relationship between field size, scatter, and magnification factor. J. Comp. Neurol. 1974b;158:295–305. doi: 10.1002/cne.901580305. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. The Ferrier lecture: functional architecture of macaque monkey visual cortex. Proc. R. Soc. B. 1977;198:1–59. doi: 10.1098/rspb.1977.0085. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N. Brain mechanisms of vision. Sci. Am. 1979;241:150–162. doi: 10.1038/scientificamerican0979-150. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, LeVay S, Wiesel T.N. Mode of termination of retinotectal fibers in macaque monkey: an autoradiographic study. Brain Res. 1975;96:25–40. doi: 10.1016/0006-8993(75)90567-3. [DOI] [PubMed] [Google Scholar]
- Hubel D.H, Wiesel T.N, LeVay S. Functional architecture of area 17 in normal and monocularly deprived macaque monkeys. Cold Spring Harb. Symp. Quant. Biol. 1976;40:581–589. doi: 10.1101/sqb.1976.040.01.054. [DOI] [PubMed] [Google Scholar]
- Hübener M, Shoham D, Grinvald A, Bonhoeffer T. Spatial relationships among three columnar systems in cat area 17. J. Neurosci. 1997;17:9270–9284. doi: 10.1523/JNEUROSCI.17-23-09270.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huberman A.D, Wang G.Y, Liets L.C, Collins O.A, Chapman B, Chalupa L.M. Eye-specific retinogeniculate segregation independent of normal neuronal activity. Science. 2003;300:994–998. doi: 10.1126/science.1080694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphrey A.L, Skeen L.C, Norton T.T. Topographic organization of the orientation column system in the striate cortex of the tree shrew (Tupaia glis). II. Deoxyglucose mapping. J. Comp. Neurol. 1980;192:549–566. doi: 10.1002/cne.901920312. [DOI] [PubMed] [Google Scholar]
- Imig T.J, Adrian H.O. Binaural columns in the primary field (A1) of cat auditory cortex. Brain Res. 1977;138:241–257. doi: 10.1016/0006-8993(77)90743-0. [DOI] [PubMed] [Google Scholar]
- Imig T.J, Reale R.A. Ipsilateral corticocortical projections related to binaural columns in cat primary auditory cortex. J. Comp. Neurol. 1981;203:1–14. doi: 10.1002/cne.902030102. [DOI] [PubMed] [Google Scholar]
- Issa N.P, Trepel C, Stryker M.P. Spatial frequency maps in cat visual cortex. J. Neurosci. 2000;20:8504–8514. doi: 10.1523/JNEUROSCI.20-22-08504.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacob F. Evolution and tinkering. Science. 1977;196:1161–1166. doi: 10.1126/science.860134. [DOI] [PubMed] [Google Scholar]
- Jain N, Catania K.C, Kaas J.H. A histologically visible representation of the fingers and palm in primate area 3b and its immutability following long-term deafferentations. Cereb. Cortex. 1998;8:227–236. doi: 10.1093/cercor/8.3.227. [DOI] [PubMed] [Google Scholar]
- Jones E.G, Burton H, Porter R. Commissural and cortico-cortical “columns” in the somatic sensory cortex of primates. Science. 1975;190:572–574. doi: 10.1126/science.810887. [DOI] [PubMed] [Google Scholar]
- Kaas J.H, Krubitzer L.A, Johanson K.L. Cortical connections of areas 17 (V-I) and 18 (V-II) of squirrels. J. Comp. Neurol. 1989;281:426–446. doi: 10.1002/cne.902810308. [DOI] [PubMed] [Google Scholar]
- Kaschube M, Wolf F, Geisel T, Löwel S. Genetic influence on quantitative features of neocortical architecture. J. Neurosci. 2002;22:7206–7217. doi: 10.1523/JNEUROSCI.22-16-07206.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaschube M, Wolf F, Puhlmann M, Rathjen S, Schmidt K.F, Geisel T, Lowel S. The pattern of ocular dominance columns in cat primary visual cortex: intra- and interindividual variability of column spacing and its dependence on genetic background. Eur. J. Neurosci. 2003;18:3251–3266. doi: 10.1111/j.1460-9568.2003.02979.x. [DOI] [PubMed] [Google Scholar]
- Katz L.C, Crowley J.C. Development of cortical circuits: lessons from ocular dominance columns. Nat. Rev. Neurosci. 2002;3:34–42. doi: 10.1038/nrn703. [DOI] [PubMed] [Google Scholar]
- Katz L.C, Shatz C.J. Synaptic activity and the construction of cortical circuits. Science. 1996;274:1133–1138. doi: 10.1126/science.274.5290.1133. [DOI] [PubMed] [Google Scholar]
- Kiper D.C, Fenstemaker S.B, Gegenfurtner K.R. Chromatic properties of neurons in macaque area V2. Vis. Neurosci. 1997;14:1061–1072. doi: 10.1017/s0952523800011779. [DOI] [PubMed] [Google Scholar]
- Krubitzer L.A, Kaas J.H. Cortical connections of MT in four species of primates: areal, modular, and retinotopic patterns. Vis. Neurosci. 1990;5:165–204. doi: 10.1017/s0952523800000213. [DOI] [PubMed] [Google Scholar]
- Lachica E.A, Beck P.D, Casagrande V.A. Parallel pathways in macaque monkey striate cortex: anatomically defined columns in layer III. Proc. Natl Acad. Sci. USA. 1992;89:3566–3570. doi: 10.1073/pnas.89.8.3566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Land P.W, Buffer S.A, Jr, Yaskosky J.D. Barreloids in adult rat thalamus: three-dimensional architecture and relationship to somatosensory cortical barrels. J. Comp. Neurol. 1995;355:573–588. doi: 10.1002/cne.903550407. [DOI] [PubMed] [Google Scholar]
- Landisman C.E, Ts'o D.Y. Color processing in macaque striate cortex: relationships to ocular dominance, cytochrome oxidase, and orientation. J. Neurophysiol. 2002;87:3126–3137. doi: 10.1152/jn.2002.87.6.3126. [DOI] [PubMed] [Google Scholar]
- Lennie P, Krauskopf J, Sclar G. Chromatic mechanisms in striate cortex of macaque. J. Neurosci. 1990;10:649–669. doi: 10.1523/JNEUROSCI.10-02-00649.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LeVay S, Nelson S.B. Columnar organization of the visual cortex. In: Leventhal A.G, editor. The neural basis of visual function. CRC Press; Boston: 1991. pp. 266–315. [Google Scholar]
- LeVay S, Voigt T. Ocular dominance and disparity coding in cat visual cortex. Vis. Neurosci. 1988;1:395–414. doi: 10.1017/s0952523800004168. [DOI] [PubMed] [Google Scholar]
- LeVay S, Hubel D.H, Wiesel T.N. The pattern of ocular dominance columns in macaque visual cortex revealed by a reduced silver stain. J. Comp. Neurol. 1975;159:559–575. doi: 10.1002/cne.901590408. [DOI] [PubMed] [Google Scholar]
- LeVay S, Stryker M.P, Shatz C.J. Ocular dominance columns and their development in layer IV of the cat's visual cortex: a quantitative study. J. Comp. Neurol. 1978;179:223–244. doi: 10.1002/cne.901790113. [DOI] [PubMed] [Google Scholar]
- LeVay S, Connolly M, Houde J, Van Essen D.C. The complete pattern of ocular dominance stripes in the striate cortex and visual field of the macaque monkey. J. Neurosci. 1985;5:486–501. doi: 10.1523/JNEUROSCI.05-02-00486.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leventhal A.G, Thompson K.G, Liu D, Zhou Y, Ault S.J. Concomitant sensitivity to orientation, direction, and color of cells in layers 2, 3 and 4 of monkey striate cortex. J. Neurosci. 1995;15:1808–1818. doi: 10.1523/JNEUROSCI.15-03-01808.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levitt J.B, Kiper D.C, Movshon J.A. Receptive fields and functional architecture of macaque V2. J. Neurophysiol. 1994;71:2517–2542. doi: 10.1152/jn.1994.71.6.2517. [DOI] [PubMed] [Google Scholar]
- Levitt J.B, Yoshioka T, Lund J.S. Connections between the pulvinar complex and cytochrome oxidase-defined compartments in visual area V2 of macaque monkey. Exp. Brain Res. 1995;104:419–430. doi: 10.1007/BF00231977. [DOI] [PubMed] [Google Scholar]
- Lia B, Olavarria J.F. The distribution of corticotectal projection neurons correlates with the interblob compartment in macaque striate cortex. Vis. Neurosci. 1996;13:461–466. doi: 10.1017/s0952523800008130. [DOI] [PubMed] [Google Scholar]
- Livingstone M.S. Ocular dominance columns in New World monkeys. J. Neurosci. 1996;16:2086–2096. doi: 10.1523/JNEUROSCI.16-06-02086.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livingstone M.S, Hubel D.H. Anatomy and physiology of a color system in the primate visual cortex. J. Neurosci. 1984a;4:309–356. doi: 10.1523/JNEUROSCI.04-01-00309.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livingstone M.S, Hubel D.H. Specificity of intrinsic connections in primate primary visual cortex. J. Neurosci. 1984b;4:2830–2835. doi: 10.1523/JNEUROSCI.04-11-02830.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livingstone M.S, Nori S, Freeman D.C, Hubel D.H. Stereopsis and binocularity in the squirrel monkey. Vision Res. 1995;35:345–354. doi: 10.1016/0042-6989(94)00133-7. [DOI] [PubMed] [Google Scholar]
- Lorento De Nó R. La corteza cerebral del ratón. Trab. Lab. Invest. Biol. (Madrid) 1922;20:41–78. [Google Scholar]
- Lorento De Nó R. Cerebral cortex: architecture, intracortical connections, motor projections. In: Fulton J, editor. Physiology of the nervous system. Oxford University Press; New York: 1949. pp. 288–330. [Google Scholar]
- Lübke J, Egger V, Sakmann B, Feldmeyer D. Columnar organization of dendrites and axons of single and synaptically coupled excitatory spiny neurons in layer 4 of the rat barrel cortex. J. Neurosci. 2000;20:5300–5311. doi: 10.1523/JNEUROSCI.20-14-05300.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lund J.S, Angelucci A, Bressloff P.C. Anatomical substrates for functional columns in macaque monkey primary visual cortex. Cereb. Cortex. 2003;13:15–24. doi: 10.1093/cercor/13.1.15. [DOI] [PubMed] [Google Scholar]
- Malach R, Amir Y, Harel M, Grinvald A. Relationship between intrinsic connections and functional architecture revealed by optical imaging and in vivo targeted biocytin injections in primate striate cortex. Proc. Natl Acad. Sci. USA. 1993;90:10 469–10 473. doi: 10.1073/pnas.90.22.10469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malach R, Schirman T.D, Harel M, Tootell R.B, Malonek D. Organization of intrinsic connections in owl monkey area MT. Cereb. Cortex. 1997;7:386–393. doi: 10.1093/cercor/7.4.386. [DOI] [PubMed] [Google Scholar]
- Maldonado P.E, Godecke I, Gray C.M, Bonhoeffer T. Orientation selectivity in pinwheel centers in cat striate cortex. Science. 1997;276:1551–1555. doi: 10.1126/science.276.5318.1551. [DOI] [PubMed] [Google Scholar]
- Marin O, Rubenstein J.L. Cell migration in the forebrain. Annu. Rev. Neurosci. 2003;26:441–483. doi: 10.1146/annurev.neuro.26.041002.131058. [DOI] [PubMed] [Google Scholar]
- Martinez-Millán L, Holländer H. Cortico-cortical projections from striate cortex of the squirrel monkey (Saimiri sciureus). A radioautographic study. Brain Res. 1975;83:405–417. doi: 10.1016/0006-8993(75)90833-1. [DOI] [PubMed] [Google Scholar]
- Meister M, Wong R.O.L, Baylor D.A, Shatz C.J. Synchronous bursts of action potentials in ganglion cells of the developing mammalian retina. Science. 1991;252:939–943. doi: 10.1126/science.2035024. [DOI] [PubMed] [Google Scholar]
- Miller K.D, Keller J.B, Stryker M.P. Ocular dominance column development: analysis and simulation. Science. 1989;245:605–615. doi: 10.1126/science.2762813. [DOI] [PubMed] [Google Scholar]
- Mooney R, Penn A.A, Gallego R, Shatz C.J. Thalamic relay of spontaneous retinal activity prior to vision. Neuron. 1996;17:863–874. doi: 10.1016/s0896-6273(00)80218-4. [DOI] [PubMed] [Google Scholar]
- Mountcastle V.B. Modality and topographic properties of cat's somatic sensory cortex. J. Neurophysiol. 1957;20:408–434. doi: 10.1152/jn.1957.20.4.408. [DOI] [PubMed] [Google Scholar]
- Mountcastle V. In: An organizing principle for cerebral function: the unit module and the distributed system. In The mindful brain. Edelman G.M, Mountcastle V.B, editors. MIT Press; Massachusetts: 1978. pp. 7–50. [Google Scholar]
- Mountcastle V.B. The columnar organization of the neocortex. Brain. 1997;120:701–722. doi: 10.1093/brain/120.4.701. [DOI] [PubMed] [Google Scholar]
- Mountcastle V.B, Berman A.L, Davies P.W. Topographic organization and modality representation in first somatic area of cat's cerebral cortex by method of single unit analysis. Am. J. Physiol. 1955;183:464. [Google Scholar]
- Mountcastle V, Davies P, Berman A. Response properties of neurons of cat's somatic sensory cortex to peripheral stimuli. J. Neurophysiol. 1957;20:374–407. doi: 10.1152/jn.1957.20.4.374. [DOI] [PubMed] [Google Scholar]
- Murphy E.H, Berman N. The rabbit and the cat: a comparison of some features of response properties of single cells in the primary visual cortex. J. Comp. Neurol. 1979;188:401–427. doi: 10.1002/cne.901880305. [DOI] [PubMed] [Google Scholar]
- Murphy K.M, Jones D.G, Van Sluyters R.C. Cytochrome-oxidase blobs in cat primary visual cortex. J. Neurosci. 1995;15:4196–4208. doi: 10.1523/JNEUROSCI.15-06-04196.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson S. Cortical microcircuits: diverse or canonical. Neuron. 2002;36:19–27. doi: 10.1016/s0896-6273(02)00944-3. [DOI] [PubMed] [Google Scholar]
- Noctor S.C, Flint A.C, Weissman T.A, Dammerman R.S, Kriegstein A.R. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 2001;409:714–720. doi: 10.1038/35055553. [DOI] [PubMed] [Google Scholar]
- Ohki K, Chung S, Ch'ng Y.H, Kara P, Reid R.C. Functional imaging with cellular resolution reveals precise micro-architecture in visual cortex. Nature. 2005;433:597–603. doi: 10.1038/nature03274. [DOI] [PubMed] [Google Scholar]
- O'Keefe L.P, Levitt J.B, Kiper D.C, Shapley R.M, Movshon J.A. Functional organization of owl monkey lateral geniculate nucleus and visual cortex. J. Neurophysiol. 1998;80:594–609. doi: 10.1152/jn.1998.80.2.594. [DOI] [PubMed] [Google Scholar]
- Olavarria J.F, Van Essen D.C. The global pattern of cytochrome oxidase stripes in visual area V2 of the macaque monkey. Cereb. Cortex. 1997;7:395–404. doi: 10.1093/cercor/7.5.395. [DOI] [PubMed] [Google Scholar]
- O'Rourke N.A, Sullivan D.P, Kaznowski C.E, Jacobs A.A, McConnell S.K. Tangential migration of neurons in the developing cerebral cortex. Development. 1995;121:2165–2176. doi: 10.1242/dev.121.7.2165. [DOI] [PubMed] [Google Scholar]
- Peterhans E, von der Heydt R. Functional organization of area V2 in the alert macaque. Eur. J. Neurosci. 1993;5:509–524. doi: 10.1111/j.1460-9568.1993.tb00517.x. [DOI] [PubMed] [Google Scholar]
- Poggio G.F, Fischer B. Binocular interaction and depth sensitivity in striate and prestriate cortex of behaving rhesus monkey. J. Neurophysiol. 1977;40:1392–1405. doi: 10.1152/jn.1977.40.6.1392. [DOI] [PubMed] [Google Scholar]
- Poggio G.F, Gonzalez F, Krause F. Stereoscopic mechanisms in monkey visual cortex: binocular correlation and disparity selectivity. J. Neurosci. 1988;8:4531–4550. doi: 10.1523/JNEUROSCI.08-12-04531.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polimeni, J. R., Granquist-Fraser, D., Wood, R. J. & Schwartz, E. L. 2005 Physical limits to spatial resolution of optical recording: clarifying the spatial structure of cortical hypercolumns. Proc. Natl Acad. Sci. USA102, 4158–4163. [DOI] [PMC free article] [PubMed]
- Powell T.P.S, Mountcastle V.B. Some aspects of the functional organization of the cortex of the postcentral gyrus of the monkey: a correlation of findings obtained in a single unit analysis with cytoarchitecture. Bull. Johns Hopkins Hosp. 1959;105:133–162. [PubMed] [Google Scholar]
- Purves D, Riddle D.R, LaMantia A.S. Iterated patterns of brain circuitry (or how the cortex gets its spots) Trends Neurosci. 1992;15:362–368. doi: 10.1016/0166-2236(92)90180-g. [DOI] [PubMed] [Google Scholar]
- Rakic P. Guidance of neurons migrating to the fetal monkey neocortex. Brain Res. 1971;33:471–476. doi: 10.1016/0006-8993(71)90119-3. [DOI] [PubMed] [Google Scholar]
- Rakic P. Prenatal development of the visual system in rhesus monkey. Phil. Trans. R. Soc. B. 1977;278:245–260. doi: 10.1098/rstb.1977.0040. [DOI] [PubMed] [Google Scholar]
- Rakic P. Specification of cerebral cortical areas. Science. 1988;241:170–176. doi: 10.1126/science.3291116. [DOI] [PubMed] [Google Scholar]
- Rakic P. Radial versus tangential migration of neuronal clones in the developing cerebral cortex. Proc. Natl Acad. Sci. USA. 1995;92:11 323–11 327. doi: 10.1073/pnas.92.25.11323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rakic P. Developmental and evolutionary adaptations of cortical radial glia. Cereb. Cortex. 2003;13:541–549. doi: 10.1093/cercor/13.6.541. [DOI] [PubMed] [Google Scholar]
- Rathjen S, Schmidt K.E, Lowel S. Two-dimensional analysis of the spacing of ocular dominance columns in normally raised and strabismic kittens. Exp. Brain Res. 2002;145:158–165. doi: 10.1007/s00221-002-1086-9. [DOI] [PubMed] [Google Scholar]
- Read J.C, Cumming B.G. Ocular dominance predicts neither strength nor class of disparity selectivity with random-dot stimuli in primate V1. J. Neurophysiol. 2004;91:1271–1281. doi: 10.1152/jn.00588.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Read H.L, Winer J.A, Schreiner C.E. Modular organization of intrinsic connections associated with spectral tuning in cat auditory cortex. Proc. Natl Acad. Sci. USA. 2001;98:8042–8047. doi: 10.1073/pnas.131591898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reid C.B, Liang I, Walsh C. Systematic widespread clonal organization in cerebral cortex. Neuron. 1995;15:299–310. doi: 10.1016/0896-6273(95)90035-7. [DOI] [PubMed] [Google Scholar]
- Reser D.H, Fishman Y.I, Arezzo J.C, Steinschneider M. Binaural interactions in primary auditory cortex of the awake macaque. Cereb. Cortex. 2000;10:574–584. doi: 10.1093/cercor/10.6.574. [DOI] [PubMed] [Google Scholar]
- Ringach D.L, Shapley R.M, Hawken M.J. Orientation selectivity in macaque V1: diversity and laminar dependence. J. Neurosci. 2002;22:5639–5651. doi: 10.1523/JNEUROSCI.22-13-05639.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rockel A.J, Hiorns R.W, Powell T.P. The basic uniformity in structure of the neocortex. Brain. 1980;103:221–244. doi: 10.1093/brain/103.2.221. [DOI] [PubMed] [Google Scholar]
- Rockland K.S, Lund J.S. Intrinsic laminar lattice connections in primate visual cortex. J. Comp. Neurol. 1983;216:303–318. doi: 10.1002/cne.902160307. [DOI] [PubMed] [Google Scholar]
- Roe A.W, Ts'o D.Y. Visual topography in primate V2: multiple representation across functional stripes. J. Neurosci. 1995;15:3689–3715. doi: 10.1523/JNEUROSCI.15-05-03689.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenthal F, Woodbury J.W, Patton H.D. Dipole characteristics of pyramidal cell activity in cat postcruciate cortex. J. Neurophysiol. 1966;29:612–625. doi: 10.1152/jn.1966.29.4.612. [DOI] [PubMed] [Google Scholar]
- Sandell J.H. NADPH diaphorase histochemistry in the macaque striate cortex. J. Comp. Neurol. 1986;251:388–397. doi: 10.1002/cne.902510309. [DOI] [PubMed] [Google Scholar]
- Shaw C, Yinon U, Auerbach E. Receptive fields and response properties of neurons in the rat visual cortex. Vision Res. 1975;15:203–208. doi: 10.1016/0042-6989(75)90208-4. [DOI] [PubMed] [Google Scholar]
- Shipp S, Zeki S. Segregation of pathways leading from area V2 to areas V4 and V5 of macaque monkey visual cortex. Nature. 1985;315:322–325. doi: 10.1038/315322a0. [DOI] [PubMed] [Google Scholar]
- Shipp S, Zeki S. The organization of connections between areas V5 and V2 in macaque monkey visual cortex. Eur. J. Neurosci. 1989;1:333–354. doi: 10.1111/j.1460-9568.1989.tb00799.x. [DOI] [PubMed] [Google Scholar]
- Shipp S, Zeki S. The functional organization of area V2. I. Specialization across stripes and layers. Vis. Neurosci. 2002;19:187–210. doi: 10.1017/s0952523802191164. [DOI] [PubMed] [Google Scholar]
- Sincich L.C, Blasdel G.G. Oriented axon projections in primary visual cortex of the monkey. J. Neurosci. 2001;21:4416–4426. doi: 10.1523/JNEUROSCI.21-12-04416.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sincich L.C, Horton J.C. Divided by cytochrome oxidase: a map of the projections from V1 to V2 in macaques. Science. 2002a;295:1734–1737. doi: 10.1126/science.1067902. [DOI] [PubMed] [Google Scholar]
- Sincich L.C, Horton J.C. Pale cytochrome oxidase stripes in V2 receive the richest projection from macaque striate cortex. J. Comp. Neurol. 2002b;447:18–33. doi: 10.1002/cne.10174. [DOI] [PubMed] [Google Scholar]
- Sincich L.C, Horton J.C. An albino-like decussation error in the optic chiasm revealed by anomalous ocular dominance columns. Vis. Neurosci. 2003a;19:541–545. doi: 10.1017/s0952523802194132. [DOI] [PubMed] [Google Scholar]
- Sincich L.C, Horton J.C. Independent projection streams from macaque striate cortex to the second visual area and middle temporal area. J. Neurosci. 2003b;23:5684–5692. doi: 10.1523/JNEUROSCI.23-13-05684.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sincich L.C, Adams D.L, Horton J.C. Complete flatmounting of the macaque cerebral cortex. Vis. Neurosci. 2003;20:663–686. doi: 10.1017/s0952523803206088. [DOI] [PubMed] [Google Scholar]
- Slimp J.C, Towe A.L. Spatial distribution of modalities and receptive fields in sensorimotor cortex of awake cats. Exp. Neurol. 1990;107:78–96. doi: 10.1016/0014-4886(90)90065-z. [DOI] [PubMed] [Google Scholar]
- Sperry R.W, Miner N, Myers R.E. Visual pattern perception following subpial slicing and tantalum wire implantations in the visual cortex. J. Comp. Physiol. Psychol. 1955;48:50–58. doi: 10.1037/h0043456. [DOI] [PubMed] [Google Scholar]
- Stosiek C, Garaschuk O, Holthoff K, Konnerth A. In vivo two-photon calcium imaging of neuronal networks. Proc. Natl Acad. Sci. USA. 2003;100:7319–7324. doi: 10.1073/pnas.1232232100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stryker M.P, Harris W.A. Binocular impulse blockade prevents the formation of ocular dominance columns in cat visual cortex. J. Neurosci. 1986;6:2117–2133. doi: 10.1523/JNEUROSCI.06-08-02117.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swindale N.V. Is the cerebral cortex modular? Trends Neurosci. 1990;13:487–492. doi: 10.1016/0166-2236(90)90082-l. [DOI] [PubMed] [Google Scholar]
- Swindale N.V, Shoham D, Grinvald A, Bonhoeffer T, Hubener M. Visual cortex maps are optimized for uniform coverage. Nat. Neurosci. 2000;3:822–826. doi: 10.1038/77731. [DOI] [PubMed] [Google Scholar]
- Tamamaki N, Nakamura K, Okamoto K, Kaneko T. Radial glia is a progenitor of neocortical neurons in the developing cerebral cortex. Neurosci. Res. 2001;41:51–60. doi: 10.1016/s0168-0102(01)00259-0. [DOI] [PubMed] [Google Scholar]
- Tamura H, Sato H, Katsuyama N, Hata Y, Tsumoto T. Less segregated processing of visual information in V2 than in V1 of the monkey visual cortex. Eur. J. Neurosci. 1996;8:300–309. doi: 10.1111/j.1460-9568.1996.tb01214.x. [DOI] [PubMed] [Google Scholar]
- Tiao Y.C, Blakemore C. Functional organization in the visual cortex of the golden hamster. J. Comp. Neurol. 1976;168:459–481. doi: 10.1002/cne.901680403. [DOI] [PubMed] [Google Scholar]
- Tootell R.B.H, Silverman M.S, De Valois R.L, Jacobs G.H. Functional organization of the second cortical visual area in primates. Science. 1983;220:737–739. doi: 10.1126/science.6301017. [DOI] [PubMed] [Google Scholar]
- Tootell R.B, Hamilton S.L, Silverman M.S. Topography of cytochrome oxidase activity in owl monkey cortex. J. Neurosci. 1985;5:2786–2800. doi: 10.1523/JNEUROSCI.05-10-02786.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ts'o D.Y, Gilbert C.D. The organization of chromatic and spatial interactions in the primate striate cortex. J. Neurosci. 1988;8:1712–1727. doi: 10.1523/JNEUROSCI.08-05-01712.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ts'o D.Y, Roe A.W, Gilbert C.D. A hierarchy of the functional organization for color, form and disparity in primate visual area V2. Vision Res. 2001a;41:1333–1349. doi: 10.1016/s0042-6989(01)00076-1. [DOI] [PubMed] [Google Scholar]
- Ts'o D.Y, Roe A.W, Gilbert C.D. A hierarchy of the functional organization for color, form and disparity in primate visual area V2. Vision Res. 2001b;41:1333–1349. doi: 10.1016/s0042-6989(01)00076-1. [DOI] [PubMed] [Google Scholar]
- Van Hooser S.D, Heimel J.A.F, Chung S, Nelson S.B, Toth L.J. Orientation selectivity without orientation maps in visual cortex of a highly visual mammal. J. Neurosci. 2005;25:19–28. doi: 10.1523/JNEUROSCI.4042-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walsh C, Cepko C.L. Clonally related cortical cells show several migration patterns. Science. 1988;241:1342–1345. doi: 10.1126/science.3137660. [DOI] [PubMed] [Google Scholar]
- Walsh C, Cepko C.L. Widespread dispersion of neuronal clones across functional regions of the cerebral cortex. Science. 1992;255:434–440. doi: 10.1126/science.1734520. [DOI] [PubMed] [Google Scholar]
- Wang G, Tanifuji M, Tanaka K. Functional architecture in monkey inferotemporal cortex revealed by in vivo optical imaging. Neurosci. Res. 1998;32:33–46. doi: 10.1016/s0168-0102(98)00062-5. [DOI] [PubMed] [Google Scholar]
- Weliky M, Bosking W.H, Fitzpatrick D. A systemic map of direction preference in primary visual cortex. Nature. 1996;379:725–728. doi: 10.1038/379725a0. [DOI] [PubMed] [Google Scholar]
- Welker E, Armstrong-James M, Bronchti G, Ourednik W, Gheorghita-Baechler F, Dubois R, Guernsey D.L, Van der Loos H, Neumann P.E. Altered sensory processing in the somatosensory cortex of the mouse mutant barrelless. Science. 1996;271:1864–1867. doi: 10.1126/science.271.5257.1864. [DOI] [PubMed] [Google Scholar]
- Wiesenfeld Z, Kornel E.E. Receptive fields of single cells in the visual cortex of the hooded rat. Brain Res. 1975;94:401–412. doi: 10.1016/0006-8993(75)90225-5. [DOI] [PubMed] [Google Scholar]
- Wong-Riley M.T.T. Columnar cortico-cortical interconnections within the visual system of the squirrel and macaque monkeys. Brain Res. 1979;162:201–217. doi: 10.1016/0006-8993(79)90284-1. [DOI] [PubMed] [Google Scholar]
- Wong-Riley M.T. Cytochrome oxidase: an endogenous metabolic marker for neuronal activity. Trends Neurosci. 1989;12:94–101. doi: 10.1016/0166-2236(89)90165-3. [DOI] [PubMed] [Google Scholar]
- Wong-Riley M, Carroll E.W. Effect of impulse blockage on cytochrome oxidase activity in monkey visual system. Nature. 1984;307:262–264. doi: 10.1038/307262a0. [DOI] [PubMed] [Google Scholar]
- Wong-Riley M, Anderson B, Liebl W, Huang Z. Neurochemical organization of the macaque striate cortex: correlation of cytochrome oxidase with Na+K+ATPase, NADPH-diaphorase, nitric oxide synthase, and N-Methyl-d-aspartate receptor subunit 1. Neuroscience. 1998;83:1025–1045. doi: 10.1016/s0306-4522(97)00432-6. [DOI] [PubMed] [Google Scholar]
- Woolsey T.A, Van Der Loos H. The structural organization of layer IV in the somatosensory region (S1) of mouse cerebral cortex: the description of a cortical field composed of cytoarchitectonic units. Brain Res. 1970;17:205–242. doi: 10.1016/0006-8993(70)90079-x. [DOI] [PubMed] [Google Scholar]
- Woolsey T.A, Welker C, Schwartz R.H. Comparative anatomical studies of the SmL face cortex with special reference to the occurrence of “barrels” in layer IV. J. Comp. Neurol. 1975;164:79–94. doi: 10.1002/cne.901640107. [DOI] [PubMed] [Google Scholar]
- Xiao Y, Wang Y, Felleman D.J. A spatially organized representation of colour in macaque cortical area V2. Nature. 2003;421:535–539. doi: 10.1038/nature01372. [DOI] [PubMed] [Google Scholar]
- Xu X, Bosking W, Sary G, Stefansic J, Shima D, Casagrande V. Functional organization of visual cortex in the owl monkey. J. Neurosci. 2004;24:6237–6247. doi: 10.1523/JNEUROSCI.1144-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yabuta N.H, Callaway E.M. Functional streams and local connections of layer 4C neurons in primary visual cortex of the macaque monkey. J. Neurosci. 1998;18:9489–9499. doi: 10.1523/JNEUROSCI.18-22-09489.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshioka T, Dow B.M. Color, orientation and cytochrome oxidase reactivity in areas V1, V2 and V4 of macaque monkey visual cortex. Behav. Brain Res. 1996;76:71–88. doi: 10.1016/0166-4328(95)00184-0. [DOI] [PubMed] [Google Scholar]
- Yoshioka T, Levitt J.B, Lund J.S. Independence and merger of thalamocortical channels within macaque monkey primary visual cortex: anatomy of interlaminar projections. Vis. Neurosci. 1994;11:467–489. doi: 10.1017/s0952523800002406. [DOI] [PubMed] [Google Scholar]
- Yoshioka T, Blasdel G.G, Levitt J.B, Lund J.S. Relation between patterns of intrinsic lateral connectivity, ocular dominance, and cytochrome oxidase-reactive regions in macaque monkey striate cortex. Cereb. Cortex. 1996;6:297–310. doi: 10.1093/cercor/6.2.297. [DOI] [PubMed] [Google Scholar]
- Zeki S, Shipp S. Modular connections between areas V2 and V4 of macaque monkey visual cortex. Eur. J. Neurosci. 1989;1:494–506. doi: 10.1111/j.1460-9568.1989.tb00356.x. [DOI] [PubMed] [Google Scholar]