

Abstract
Since the production of eosinopoietic cytokines (GM-CSF, IL-3, IL-5) is inhibited by glucocorticoids, while responsiveness to these cytokines is enhanced in bone-marrow of allergic mice, we studied the ability of glucocorticoids to modulate murine bone-marrow eosinopoiesis.
Progenitor (semi-solid) and/or precursor (liquid) cultures were established from bone-marrow of: (a) normal mice; (b) ovalbumin-sensitized and challenged mice or (c) dexamethasone (1–5 mg kg−1) injected mice. Cultures were established with GM-CSF (2 ng ml−1) or IL-5 (1 ng ml−1), respectively, alone or associated with dexamethasone, hydrocortisone or corticosterone. Total myeloid colony numbers, frequency and size of eosinophil colonies, and numbers of eosinophil-peroxidase-positive cells were determined at day 7.
In BALB/c mice, dexamethasone (10−7 M) increased GM-CSF-stimulated myeloid colony formation (P=0.01), as well as the frequency (P=0.01) and size (P<0.01) of eosinophil colonies. Dexamethasone (10−7 M) alone had no effect. Dexamethasone (10−7–10−10 M) increased (P<0.002) eosinophil precursor responses to IL-5. Potentiation by dexamethasone was still detectable: (a) on low density, immature, nonadherent BALB/c bone-marrow cells, (b) on bone-marrow from other strains, and (c) on cells from allergic mice. Hydrocortisone and corticosterone had similar effects. Dexamethasone administered in vivo, 24 h before bone-marrow harvest, increased subsequent progenitor responses to GM-CSF (P=0.001) and precursor responses to IL-5 (P<0.001). These effects were blocked by RU 486 (20 mg kg−1, orally, 2 h before dexamethasone, or added in vitro at 10 μM, P<0.001).
Glucocorticoids, acting in vivo or in vitro, through glucocorticoid receptors, enhance bone-marrow eosinopoiesis in naïve and allergic mice.
Keywords: Glucocorticoids, eosinophil, GM-CSF, IL-5, bone-marrow
Introduction
Glucocorticoids suppress peripheral blood and tissue eosinophilia by inhibiting the production of cytokines required for bone-marrow eosinopoiesis, by inducing apoptosis in mature eosinophils and by antagonizing the in vitro survival-promoting effects of Interleukin-5 (IL-5), granulocyte-macrophage colony stimulating factor (GM-CSF) and Interleukin-3 (IL-3; reviewed in Giembycz & Lindsay, 1999). However, whether they also suppress the effects of IL-5, GM-CSF and IL-3 on bone-marrow eosinophil progenitors and precursors has not been thoroughly investigated, presumably because such interaction is not expected to occur in individuals whose eosinopoietic cytokine production has already been suppressed by therapeutical doses of glucocorticoids (Barnes, 1996; Schleimer & Bochner, 1994; Giembycz & Lindsay, 1999). This remains, nevertheless, an important issue, because a Th2 pattern of cytokine production, including IL-5, may even be induced by glucocorticoids, working synergically with Interleukin-4 (reviewed by Wilckens & De Rijk, 1997), while production of GM-CSF may resist high doses of glucocorticoids (Kunicka et al., 1993; Her et al., 1991).
The study of glucocorticoid effects on eosinopoiesis, which has been centered on human progenitors, has yielded conflicting results, depending on the experimental conditions. Early reports of inhibition of human eosinophil colony formation by glucocorticoids are now considered difficult to interpret because the crude preparations used to stimulate colony growth contain several factors with opposing effects (reviewed by Schleimer & Bochner 1994, and Giembycz & Lindsay, 1999). A careful study by Barr et al. (1987), following the initial observations of Slovick et al. (1985), demonstrated enhancement of eosinophil colony formation from human bone-marrow by physiological and pharmacological concentrations of hydrocortisone. Butterfield et al. (1986) reported that hydrocortisone at pharmacological concentrations increased eosinophil colony formation by bone-marrow cells of normal subjects, but inhibited eosinopoiesis in an allergic patient. Evidence for glucocorticoid resistance of human peripheral blood eosinophil progenitors was also provided by Barr et al. (1987) and Shalit et al. (1995).
We reported that allergen challenge of ovalbumin-sensitized mice increases the responses of eosinophil progenitors and precursors to IL-5 and IL-3 (Gaspar Elsas et al., 1997). Because of the increasing interest in murine models of asthma, it is important to evaluate whether glucocorticoids also modulate responsiveness to cytokines in murine eosinopoiesis. Furthermore, it is necessary to define whether their effects, as those of the eosinopoietic cytokines themselves, are modified by immunization and challenge. To do so, we evaluated the ability of glucocorticoids to modulate responses to GM-CSF, IL-5 and IL-3 in bone-marrow culture established from naïve or from ovalbumin-sensitized and challenged mice of six different strains, including those from the high IgE-producing strain BP-2 (Eum et al., 1995). Furthermore, because of the increased sensitivity of precursor assays, relative to progenitor assays, to the effects of eosinopoietic cytokines (Sanderson, 1992), we have extended our analysis to the effects of glucocorticoids on IL-5-stimulated eosinophil differentiation in liquid culture, an issue which has not been addressed in previous studies. We report that glucocorticoids, either present at relatively low concentrations in bone-marrow culture, or administered in vivo 24 h before bone-marrow harvest, enhance eosinopoiesis in bone-marrow cultures of both naïve and allergic mice.
Methods
Animals and animal procedures
For most of the study, male and female BALB/c mice bred at FIOCRUZ (Rio de Janeiro, Brazil), were used at 6–8 weeks of age. BALB/c mice bred at Elevage Janvier (Le Genest Saint-Isle, France), yielded comparable results. In selected experiments, A/J, CBA/J, DBA/2, and C57BL/10 mice bred at FIOCRUZ, and BP-2 mice, bred at Elevage Janvier, were also used. BALB/c and BP-2 mice were immunized with two s.c. 0.4 ml injections of 100 μg ovalbumin mixed with 4 mg ml−1 Al(OH)3 in 0.9% NaCl, at 7 day intervals. BALB/c mice were intranasally challenged with 10 μg ovalbumin in 50 μl 0.9% NaCl, 1 week after the second injection, a procedure that increases responses to eosinopoietic cytokines (Gaspar Elsas et al., 1997). BP-2 mice were submitted to repeated challenges (twice a day, for 2 days), which induce bronchopulmonary hyperreactivity in this strain (Eum et al., 1995). Both groups were sacrificed 24 h after completing the challenge procedure. Naïve BALB/c mice were injected once with dexamethasone i.p. (1 or 5 mg kg−1), 24 h before bone-marrow harvest. Control animals received an equivalent volume of saline. Where indicated, animals were orally given RU 486 (20 mg kg−1, Perretti & Flower, 1996) in 0.5% methylcellulose, with the help of a spherical end stainless steel catheter, 2 h before dexamethasone administration. Control animals received an equivalent volume of 0.5% methylcellulose. Animal handling followed the standard procedures adopted at the Institut Pasteur, Paris (France).
Reagents
Heat-inactivated foetal calf serum (FCS), L-glutamine and culture media were from Gibco (Life Technologies SARL, Cergy Pointoise, France). Agar Noble from Difco (Detroit, MI, U.S.A.), Diff-Quick from DADE Diagnostika GmbH (Unterschleisshei, Germany) and Harris' Haematoxylin from Réactifs RAL (Paris, France). Recombinant haemopoietic factors (rmIL-3, rmIL-5 and rmGM-CSF) were from Pharmingen (San Diego, CA, U.S.A.). Methylcellulose, dexamethasone 21-phosphate (disodium salt), hydrocortisone (sodium succinate), corticosterone and mifepristone (RU 486, ref. M-8046) were from Sigma (St. Louis, MO, U.S.A.). Dexamethasone 21-phosphate was dissolved in serum-free culture medium and diluted from a 10−3 M stock solution in medium or in saline for culture or in vivo administration, respectively. RU 486 was dissolved in methylcellulose (prepared as a 0.5% gel in water) for oral administration, or used at 10−5 M in culture medium (final concentration) in bone marrow culture (Hardy et al., 1996). Ovalbumin (5× crystallized) was from ICN Biomedicals, Inc. (Costa Mesa, CA, U.S.A.), Al(OH)3 from Merck (Darmstadt, Germany).
Bone-marrow cell studies
Bone-marrow cell harvest, identification and separation
Bone-marrow cells, collected by flushing the two femurs of 5–8 mice with RPMI 1640 medium containing 10% FCS, were washed and counted. The frequency of cells stained for eosinophil peroxidase (EPO) following the protocol of Ten et al. (1989) was determined in cytocentrifuge smears (Gaspar Elsas et al., 1997). The cytochemical pattern of EPO+ cells in stained bone-marrow preparations was identical to that described by Horton et al. (1996). Where indicated, cells were submitted to two cycles of adherence in 100 mm culture dishes (30 min, 37°C, 5% CO2/95% air), and nonadherent cells were further separated by centrifugation for 20 min on discontinuous Percoll (75, 60 and 40%) density gradients, at 100×g, 20°C (Gaspar Elsas et al., 1997). Cells at the three interfaces (Layers I–III from top to bottom) were collected, washed, and counted. Haemopoietic progenitors and precursors were enriched in Layer II (see Results).
Progenitor assays
Semi-solid cultures were established in 35 mm culture dishes, at least in triplicate. One ml of Iscove's modified Dulbecco's medium with 20% FCS, containing 2×105 cells bone-marrow cells and GM-CSF (2 ng ml−1 final concentration), with or without Interleukin-5 (0.2 ng ml−1 final concentration), with or without dexamethasone, hydrocortisone or corticosterone (see Results for dose-response relationships), were mixed with agar to 0.3% final concentration (Metcalf, 1998). Murine bone-marrow colony formation, unlike that in human cord blood (Schibler et al., 1994), was strictly dependent on exogenous growth factors, in agreement with observations from many laboratories (see Bagby, 1994, for an extensive review). A colony was defined as a progenitor-derived ensemble larger than 50 cells, as opposed to a cluster of less than 50 cells (Bagby, 1994; Clutterbuck et al., 1989). Colonies were scored at day 7 under the inverted microscope at low magnification, and the frequency of eosinophil colonies was determined on agar layers dried (50°C), mounted on microscope slides, stained for EPO, and scored under high magnification (Gaspar Elsas et al., 1997). Colony morphology was identical to that described in previous studies and included mixed granulocyte-macrophage-eosinophil colonies, along with pure eosinophil colonies (Clutterbuck et al., 1989; Butterfield et al., 1986; Gaspar Elsas et al., 1997). Dried/stained agar layers were further used for morphometric analysis (see below). We confirmed, in preliminary experiments, that these concentrations of GM-CSF and IL-5 were adequate for colony growth, and that cell density was appropriate for counting and for the morphological study of murine eosinophil colonies. Colony formation was low at 5×104 cells/plate and increased linearly between 1 and 2×105 cells/plate. Glucocorticoid effects on colony growth (see Results) were identical at low and high cell densities, did not require glucocorticoid replacement, in agreement with observations by other groups (Inman et al., 1997; Barr et al., 1987; Butterfield et al., 1986), and culture at 2×105 cells/plate, which approximates the conditions used by many laboratories (Begley et al., 1997; Hisha et al., 1997; Rinehart et al., 1997; Barr et al., 1987; Butterfield et al., 1985) was used for all the experiments shown, because it allowed adequate morphometrical study of eosinophil colonies, even at the low frequency found in naïve mice.
Precursor assays
Liquid bone-marrow cultures (106 cells in a 1 ml volume, in a 24-well cluster, Gaspar Elsas et al., 1997) were seeded in RPMI 1640 medium, with 10% FCS, 2 mM L-glutamine, and penicillin-streptomycin, at 37°C, 5% CO2/95% air, at least in triplicate, in the absence or in the presence of IL-5 (0.01–1 ng ml−1), IL-3 (0.2 ng ml−1), or GM-CSF (2 ng ml−1), with or without dexamethasone, hydrocortisone or corticosterone (see Results for dose-response relationships). The frequency of EPO+ cells was determined after 7 days of culture in cytocentrifuge smears (Clutterbuck et al., 1989; Gaspar Elsas et al., 1997), thus circumventing the problems due to stronger induction of EPO expression by IL-5 as compared to GM-CSF/IL-3 (Oskeritzian et al., 1998). EPO is a constituent of the coreless granules, the earliest morphological marker of the eosinophil lineage, detectable from the promyelocyte stage onwards, which persists in all maturation stages up to mature eosinophils (Egesten et al., 1997; reviewed in Giembycz & Lindsay, 1999). EPO synthesis is an early event and transcription of the EPO gene is decreased or absent in mature forms (Gruart et al., 1992), but EPO protein accumulates and remains stored in specific granules for the lifetime of the cell. Cytokines and glucocorticoids were added only once, at the beginning of the culture, at the same time, and no replacement was necessary, in agreement with observations from other laboratories (Strath et al., 1985; Clutterbuck et al., 1989; Oskeritzian et al., 1998). In preliminary experiments, we verified that rmIL-5 from Pharmingen, at 1 ng ml−1, was as effective as plateau concentrations (2–6 ng ml−1) of rmIL-5 from Immunogenex (Los Angeles, CA, U.S.A.), that had been used in previous studies (Gaspar Elsas et al., 1997). With both sources of IL-5, significantly higher responses were observed in cultures from sensitized and challenged animals, relative to naïve mice (Gaspas Elsas et al., 1997). Hence, rmIL-5 from Pharmingen was used for all experiments shown here. Dose-response relationships for IL-5 in liquid culture are described in Results.
Morphometric analysis of colony size
Computer-assisted morphometric analysis (Hall & Fu, 1985) of pure eosinophil colonies in EPO-stained, haematoxylin-counterstained, dried agar layers was carried out by contouring individual colonies, with the help of the Image Pro Plus (MediaCybernetics, Inc.) software, calibrated with an external standard, using an Olympus B-Max 50 microscope, a Sony Video System and a 486 DX2 100 DPU. The surface area was expressed in mm2. A total of 6–11 colonies were randomly sampled from dried layers originating in at least three separate experiments with naïve or allergic bone-marrow, in order to calculate the average surface area.
Electron microscopic studies
Cell suspensions were prefixed with 1.6% glutaraldehyde in 0.1 M phosphate buffer, pH 7.3. The cells were immobilized by centrifugation at 4500 r.p.m., and after washing in buffer the pellets were dehydrated in ethanol and embedded in epoxy resin. The ultra-thin sections were counter-stained with uranyl acetate and lead citrate, and examined using a JEOL-JEM 1010 electron microscope.
Statistical analysis
The data were analysed with the help of the Systat for Windows version 4 software, using factorial analysis of variance, and with the Tukey (HSD) correction for multiple comparisons between different treatments. Eosinophil colony size was not normally distributed and therefore the Mann-Whitney nonparametric test was used.
Results
Effect of dexamethasone on eosinophil progenitors in naïve bone-marrow cultures
To define the effect of dexamethasone on colony formation by progenitors from naïve BALB/c mice, semi-solid bone-marrow cultures were established in the presence of GM-CSF, alone or with dexamethasone. As shown in Figure 1, a significant increase in the total number of colonies formed in the presence of GM-CSF and dexamethasone (10−7 M) was observed, relative to the control cultures with GM-CSF alone (P<0.001). Dexamethasone by itself induced no colony formation. At 10−8 M, the effect of dexamethasone was not significant (P=0.092). At 10−12 M, dexamethasone had no effect, with or without GM-CSF. Over the entire series of experiments (n=24), dexamethasone at 10−7 M increased colony formation, on the average, by a factor of 1.74. Addition of 0.2 ng ml−1 IL-5 did not further increase the total colony formation in response to 2 ng ml−1 GM-CSF. However, addition of dexamethasone still enhanced significantly colony formation relative to GM-CSF and IL-5-stimulated controls (not shown).
Figure 1.
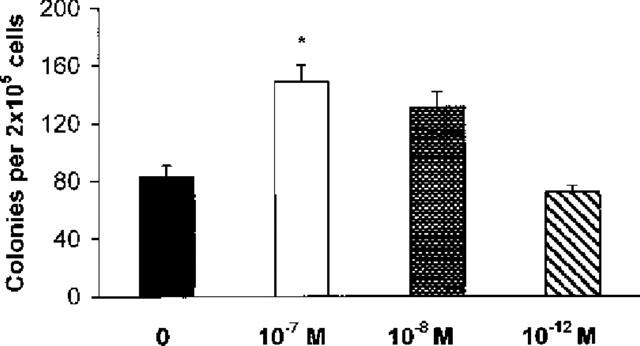
Effect of dexamethasone on GM-CSF-induced colony formation. The data are mean±s.e.mean of the number of myeloid colonies formed by bone-marrow from naïve BALB/c mice. GM-CSF (2 ng ml−1) alone, closed bar. GM-CSF in association with dexamethasone, 10−7 M, open bar, 10−8 M, stippled bar, 10−12 M, cross-hatched bar. Data are derived from a total of 24 experiments, each experiment performed with pooled bone-marrow cells from 5–7 mice. Asterisk indicates significant difference relative to the GM-CSF control (P<0.001).
To evaluate whether dexamethasone affected progenitor commitment to the eosinophil lineage, in addition to a general stimulation of myelopoiesis, differential counts of myeloid colonies in EPO-stained dried agar layers were carried out. The EPO-stained colonies corresponded to the morphological and cytochemical patterns described before (Gaspar Elsas et al., 1997; Butterfield et al., 1986), and consisted of both pure eosinophil colonies, and mixed colonies containing eosinophils, granulocytes and monocytes/macrophages (Bagby, 1994; Clutterbuck et al., 1989). As shown in Table 1(a), dexamethasone (10−7–10−8 M) increased significantly the frequency of pure eosinophil colonies formed in the presence of GM-CSF, relative to GM-CSF-stimulated controls. On the other hand, IL-5 also increased significantly the formation of pure eosinophil colonies, relative to the same GM-CSF-stimulated controls, in agreement with its reported effect in promoting eosinophil commitment of GM-CSF-stimulated colony-forming cells in human bone-marrow (Shalit et al., 1995; Clutterbuck et al., 1989). However, no further increase in the frequency of pure eosinophil colonies was observed, when both IL-5 and dexamethasone were added to GM-CSF-stimulated cultures.
Table 1.
Effect of dexamethasone on the frequency and the size of eosinophil colonies in naïve bone marrow cultures

In order to evaluate whether dexamethasone affected eosinophil colony size, which is a function of the number of cell cycles undergone by the progeny of individual colony-forming cells (Bagby, 1994), pure eosinophil colonies formed in the presence of GM-CSF, alone or in association with dexamethasone (10−7 and 10−8 M), were submitted to morphometric analysis. As shown in Table 1(b), dexamethasone significantly increased the mean area of eosinophil colonies, in cultures stimulated by GM-CSF and dexamethasone (either 10−7 or 10−8 M), relative to GM-CSF-stimulated controls.
Effect of dexamethasone on eosinophil precursors in naïve bone-marrow culture
To evaluate whether synergism between dexamethasone and the eosinopoietic cytokines extended to eosinophil precursor responses in a terminal differentiation assay (Strath et al., 1985; Oskeritzian et al., 1998), which is more sensitive to eosinopoietic cytokines than the commonly used eosinophil colony formation assays (Sanderson, 1992), we established liquid bone-marrow cultures in the presence of IL-5, of GM-CSF, or of IL-3, alone or in association with dexamethasone, and determined the frequency of EPO+ cells on cytocentrifuge smears.
As shown in Figure 2, dexamethasone (10−7–10−10 M) increased significantly (P<0.002 for all four dexamethasone concentrations, as compared to control cultures) the frequency of EPO+ cells in cultures stimulated with 1 ng ml−1 IL-5. Dexamethasone, at these effective concentrations, did not support eosinophil survival or differentiation by itself. Dexamethasone, at 10−6 M, or at 10−11 M, had no significant effect, even in the presence of IL-5. Significant synergism between dexamethasone at the same concentration range (10−7–10−10 M) and 0.1 ng ml−1 IL-5 was also observed (P<0.05 for all four dexamethasone concentrations, relative to control cultures), even though both control responses and the effect of dexamethasone were of a lesser magnitude. Finally, at even lower (0.01 ng ml−1) IL-5 concentrations, eosinophil differentiation was minimal, and no significant enhancing effect of dexamethasone was detected.
Figure 2.
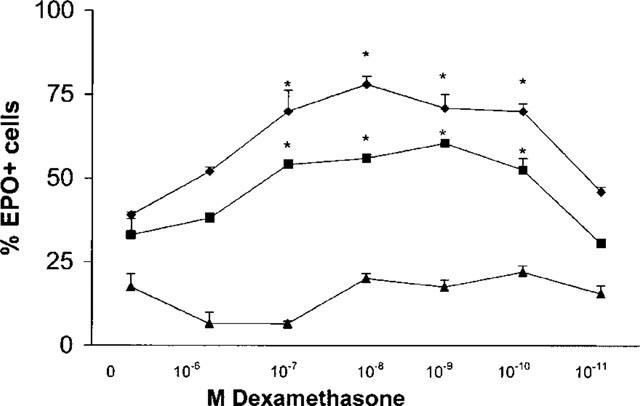
Effect of in vitro exposure to dexamethasone on IL-5-driven eosinophil differentiation in liquid culture. Data are mean±s.e.mean of the per cent EPO+ cells in liquid cultures established from naïve BALB/c mice, in the presence of 1 (losenges), 0.1 (squares) or 0.01 (triangles) ng ml−1 IL-5 and in the absence (0) or in the presence of dexamethasone, at the indicated molar concentrations. Data are derived from four experiments. Asterisks indicate significant differences relative to the respective IL-5 controls (P<0.002 for dexamethasone at 10−7–10−10 M in the upper curve, P<0.05 for the same concentration range in the middle curve).
On the other hand, the total number of EPO+ cells in the culture (which is calculated as the product of the per cent EPO+ cells and the total nucleated cell counts) increased, from (4.91±0.48)×105 ml−1 (mean±s.e.mean), in cultures stimulated by IL-5 (1 ng ml−1) alone, to (12.53± 0.06)×105 ml−1, in cultures stimulated by IL-5 associated with dexamethasone (10−7 M). This increase was significant (P<0.001). This confirms that the increase in per cent EPO+ cells in cytocentrifuge smears is paralleled by actual increases in the total numbers of EPO+ cells in these cultures (as indirectly determined from total and differential cell counts), in agreement with previous observations from our own (Gaspar Elsas et al., 1997) and other groups (Strath et al., 1985; Oskeritzian et al., 1998).
Based on these dose-response relationships, and on the similar results from both methods, subsequent experiments in liquid culture were carried out with 1 ng ml−1 IL-5 (which yielded plateau responses in control cultures, and presented optimal synergism with dexamethasone), and the per cent EPO+ cells was directly determined on cytocentrifuge smears.
Since GM-CSF and IL-3 also stimulate eosinophil precursors (although less efficiently than IL-5, Clutterbuck et al., 1989; Sanderson, 1992), we evaluated the ability of dexamethasone to increase responses to these cytokines in liquid culture. The per cent EPO+ cells increased from 3.33±0.76 (mean±s.e.mean) in IL-3-treated cultures to 8.77±0.76 in cultures treated with IL-3 plus dexamethasone (10−7 M) (P<0.001). Similarly, the per cent EPO+ cells increased from 6±0.86 in GM-CSF-treated control cultures to 17±1.86 in cultures treated with GM-CSF plus dexamethasone (10−7 M) (P<0.001).
We examined the morphology of the EPO+ cells in IL-5 and dexamethasone-stimulated cultures. EPO+ cells in IL-5-stimulated control cultures presented essentially as single cells with strong brown cytoplasmic staining, dispersed among EPO− cells, which are recognizable after counterstaining with haematoxylin, as shown in Figure 3a. In contrast, EPO+ cells in cultures stimulated by IL-5 and dexamethasone were found both as single cells and as clusters of rather uniform appearance, as shown in Figure 3b. Under high magnification, mature eosinophils (small, with abundant peroxidase-containing granules and donut-shaped nucleus) were seen in both control (Figure 3c) and dexamethasone-treated (Figure 3d) cultures. However, EPO+ cells in dexamethasone and IL-5-stimulated cultures also included many immature cells (larger, with fewer peroxidase-containing granules, and with larger, incompletely segmented nuclei), usually presenting in clusters (Figure 3d). Cells with pyknotic nuclei, most of which were EPO−, were found in both control and dexamethasone-treated cultures. Ultrastructural studies demonstrated the characteristic eosinophil granule structure (crystalloid) in morphologically intact, mature cells in cultures stimulated for 7 days with IL-5 and dexamethasone. Cells bearing eosinophil granules could not be distinguished, on ultrastructural criteria, from those found in IL-5-stimulated cultures.
Figure 3.

Morphological features of EPO+ cells grown in the presence of dexamethasone plus IL-5. Cytocentrifuge smears from 7 day-liquid bone-marrow cultures established in the presence of 1 ng ml−1 IL-5 (a and c) or of IL-5 associated with 10−7 M dexamethasone (b and d) were stained for EPO, counterstained with Harris Haematoxilin and photographed under low (100×, (a), 125×, (b) or high (1000×, under immersion, c and d) magnification.
Effects of naturally occurring glucocorticoids on eosinophil progenitors and precursors
To define whether naturally occurring glucocorticoids acted like dexamethasone, bone-marrow cultures were established in the presence of GM-CSF and either hydrocortisone or corticosterone. Hydrocortisone (10−6 M) increased total myeloid colony formation from 82±4.9 (mean±s.e.mean) to 150±6.4 colonies per 2×105 cells (P=0.001, data from three experiments). Hydrocortisone (10−6 M) also increased the per cent EPO+ cells from 49±1 to 82±2 (P<0.001, n=9). This effect of hydrocortisone on frequency of EPO+ cells was paralleled by a significant (P=0.001, n=3) increase in absolute numbers of EPO+ cells from (5.74±0.6)×105 ml−1, in cultures stimulated by IL-5 (1 ng ml−1) alone, to (15.72±1.48)×105 ml−1, in cultures stimulated by IL-5 associated with hydrocortisone (10−6 M). Corticosterone (10−8 M) had similar effects on both progenitors and precursors (not shown).
Factors affecting the effect of glucocorticoids on eosinophil progenitors and precursors
Because adherent, accessory cells play a major role in the stimulation of haematopoiesis by glucocorticoids in human bone-marrow (Rinehart et al., 1997), we examined whether synergism would be detectable in cultures established from murine bone-marrow depleted of adherent cells (as well as of other mature myeloid cells) and enriched in progenitors and precursors. As shown in Table 2, Layer II (lower density) cells were enriched in haemopoietic progenitors, because they formed about four times more colonies than unseparated bone-marrow in identical conditions. Colony formation stimulated by GM-CSF was significantly enhanced by about 2 fold in both unseparated bone-marrow and in Layer II cells, in the presence of dexamethasone (10−7 M).
Table 2.
Effect of dexamethasone on colony formation by nonadherent, low density bone-marrow cells

Because individual differences in responsiveness strongly influence the effect of glucocorticoids on human eosinopoiesis (Barr et al., 1987; Butterfield et al., 1986), we examined the synergism between dexamethasone and GM-CSF, using a broader panel of inbred strains, which differ at both histocompatibility and background genes. As shown in Table 3, dexamethasone enhanced significantly GM-CSF-driven colony formation in bone-marrow from C57BL/10, A/J, DBA/2 and BP-2 mice. Dexamethasone also enhanced IL-5-stimulated differentiation in bone-marrow from all of these strains (not shown).
Table 3.
Effect of dexamethasone on colony formation by naïve bone-marrow cells from inbred strains of different genetic backgrounds

Effects of dexamethasone on bone-marrow from intranasally-challenged, immunized mice
We further determined whether the synergistic effect of dexamethasone would still be detectable when bone-marrow of immune, challenged BALB/c and BP-2 mice was cultured. In a total of 10 experiments, there was no significant increase in the total number of myeloid colonies formed by sensitized/challenged BALB/c bone-marrow, as compared with naïve mice (P=0.595). On the other hand, as shown in Figure 4, dexamethasone (10−7 M), in the presence of GM-CSF, significantly increased colony formation by bone-marrow from ovalbumin-sensitized, challenged mice of this strain, relative to GM-CSF-stimulated cultures. As also shown in Figure 4, significant enhancement was still observed with ovalbumin-sensitized BP-2 mice submitted to repeated intranasal challenge, which induces bronchopulmonary hyperreactivity in this hyper-IgE strain (Eum et al., 1995).
Figure 4.
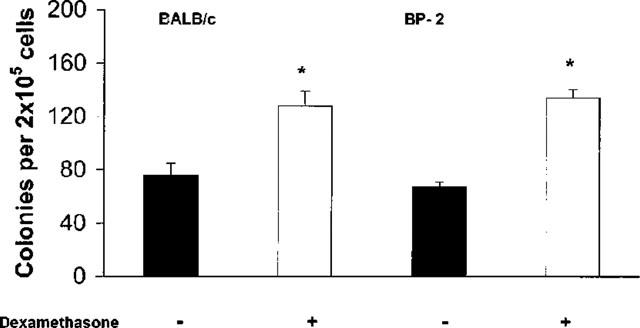
Effect of dexamethasone on GM-CSF-induced colony formation in bone marrow culture from sensitized/challenged mice. The data are mean±s.e.mean of the number of myeloid colonies formed by bone-marrow of ovalbumin-sensitized, challenged BALB/c mice (on the left), challenged once, or BP-2 mice (on the right) challenged four times (2× day−1). GM-CSF (2 ng ml−1) alone, closed bars; GM-CSF plus dexamethasone, 10−7 M, open bars. Data from BALB/c and BP-2 mice are derived from respectively 10 and four experiments. Asterisks indicate a significant difference relative to the respective GM-CSF controls (respectively P=0.002 and P=0.023).
To confirm that the effect of dexamethasone on sensitized/challenged BALB/c mice, as on naïve animals, involved selective modulation of the eosinophil lineage, in addition to a general stimulatory effect on myelopoiesis, we evaluated the frequency and size of eosinophil colonies in bone-marrow cultures. As shown in Table 4, immunization and challenge, by themselves, increased the frequency and size of eosinophil colonies induced by GM-CSF alone, relative to naïve mice (see Table 1), in agreement with previously reported findings (Gaspar Elsas et al., 1997). The increase was significant (P=0.032). Dexamethasone (10−7 M) further increased both the frequency and the mean size of eosinophil colonies relative to GM-CSF-stimulated controls. Dexamethasone (10−8 M) had a significant effect on colony size, even though it did not significantly increase eosinophil colony frequency. Dexamethasone also increased eosinophil precursor responses to IL-5 in sensitized-challenged BALB/c mice (not shown).
Table 4.
Effect of dexamethasone on eosinophil colonies from bone-marrow of sensitized/challenged mice

Effect of the in vivo treatment with dexamethasone on bone-marrow eosinophil progenitors and precursors
To confirm that the enhancing effects of dexamethasone on bone-marrow eosinophil progenitors and precursors were not restricted to in vitro conditions, we injected normal BALB/c mice with dexamethasone (5 or 1 mg kg−1), i.p. Bone-marrow from dexamethasone- or saline-injected mice was harvested 24 h later. Dexamethasone at either dose had no significant effect on the numbers of bone-marrow cells recovered per femur, nor on the frequency of EPO+ cells, as assessed from cytocentrifuge smears of freshly harvested bone-marrow. Semi-solid cultures were established in the presence of GM-CSF, and liquid cultures were established in the presence of IL-5. Bone-marrow cultures were incubated for 7 days before scoring colonies and EPO+ cells. As shown in Figure 5, in vivo treatment with both doses of dexamethasone significantly increased colony formation in the presence of GM-CSF, relative to cultures from saline-injected controls. Addition of dexamethasone (10−7 M) to the cultures significantly enhanced colony formation by bone-marrow from saline-injected controls (P<0.001), but not from both groups of dexamethasone-injected mice. In vivo treatment with both doses of dexamethasone also significantly increased the response of eosinophil precursors in liquid culture relative to saline-injected controls (not shown). Again, no further enhancement was obtained by addition of dexamethasone (10−7 M) to cultures from dexamethasone-treated mice. Finally, a significant increase in eosinophil colony size (P<0.01) was also observed in mice treated with dexamethasone (1 mg kg−1), confirming that the in vivo effect of dexamethasone involves selective stimulation of the eosinophil lineage, in addition to a general enhancing effect on myelopoiesis.
Figure 5.
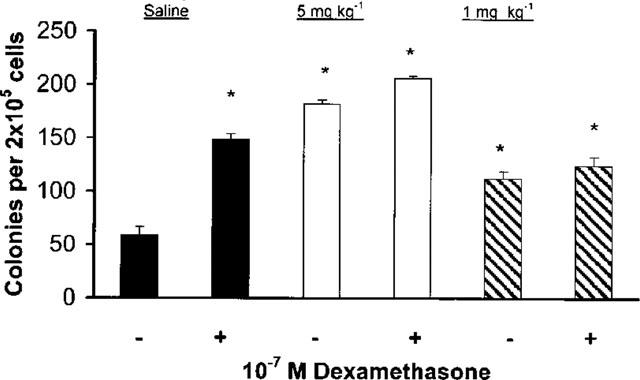
Effect of in vivo administration of dexamethasone on GM-CSF-induced colony formation. The data are mean±s.e.mean of the number of myeloid colonies formed after 7 days by cultured bone-marrow from BALB/c mice that had been injected with saline (closed bars), or dexamethasone, 5 mg kg−1 (open bars) or 1 mg kg−1 (cross-hatched bars), 24 h before bone-marrow harvest. −, cultures with GM-CSF (2 ng ml−1) alone; +, cultures with GM-CSF plus dexamethasone, 10−7 M. Data are derived from seven experiments. Asterisks indicate significant difference relative to the GM-CSF cultures from saline-injected mice (P<0.001 in all cases).
Evidence for involvement of glucocorticoid receptors in the effects of dexamethasone
To evaluate whether stimulation of bone-marrow progenitors and precursors involved glucocorticoid receptors, the ability of RU 486 (mifepristone) to block the effects of dexamethasone was studied in naïve BALB/c mice. RU 486 at 20 mg kg−1 (or 0.5% methylcellulose, used as vehicle) was administered orally to mice 2 h before dexamethasone 5 mg kg−1 (or saline) was injected. Twenty-four hours after dexamethasone injection, bone-marrow was harvested and used for semi-solid culture. Administration of RU 486, or of methylcellulose, as a vehicle, had no effect on bone-marrow cellularity, as assessed from the numbers of bone-marrow cells recovered per animal. As shown in Figure 6, colony formation by GM-CSF-stimulated bone-marrow cultures derived from mice given methylcellulose, or RU 486 in methylcellulose, was comparable to that of untreated naïve mice (see Figure 1). In contrast, cultures from mice given methylcellulose, followed by dexamethasone injection, yielded significantly increased (P<0.001) colony numbers. This effect of dexamethasone was totally abolished in cultures established from bone-marrow of mice given by RU 486 in methylcellulose before glucocorticoid injection. Also, RU 486, at 10−5 M, was also able to block completely the enhancing effect of dexamethasone (10−7 M) on the eosinophil differentiation induced in liquid culture by IL-5 at 1 ng ml−1 (not shown).
Figure 6.
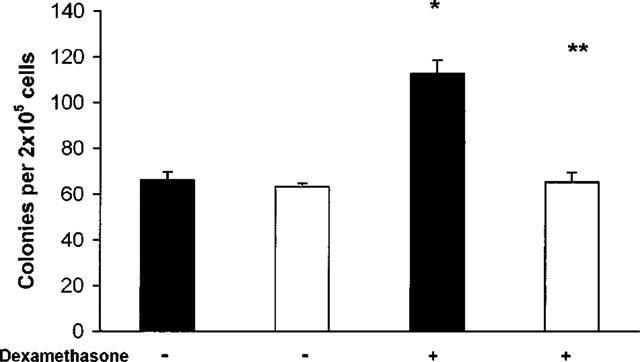
Effect of pretreatment by RU 486 on colony formation by bone-marrow of naïve BALB/c mice injected with dexamethasone. Data are the numbers of colonies formed (mean±s.e.mean) by bone marrow of naive BALB/c mice given methylcellulose (closed bars) or RU 486 (20 mg kg−1 in methylcellulose, open bars), 2 h before injection of dexamethasone (5 mg kg−1, +) or of saline (−), as a control. Data are derived from three experiments. *P<0.001 relative to mice given methylcellulose and saline; **P<0.001 relative to mice given methylcellulose and dexamethasone.
Discussion
The therapeutic effects of anti-inflammatory glucocorticoids are often ascribed to inhibition of eosinopoietic cytokine secretion, and to apoptosis of mature eosinophils (reviewed in Schleimer & Bochner, 1994; Giembycz & Lindsay, 1999). Since airway allergen exposure upregulates the responses of murine bone-marrow to the haemopoietic cytokines after airway allergen exposure in mice (Gaspar Elsas et al., 1997), modulation of these enhanced responses is a potential target for glucocorticoids. We have evaluated whether dexamethasone modulates responses of murine bone-marrow eosinophil progenitors and precursors to eosinopoietic cytokines, and how that relates to glucocorticoid enhancement of human (Rinehart et al., 1997) and murine (Maruyama et al., 1999) myelopoiesis.
In this study, dexamethasone increased the total number of myeloid colonies formed, the fraction of myeloid colonies that were committed to eosinophil production and the size of the eosinophil colonies. By acting simultaneously on these three parameters, it significantly enhanced bone-marrow responses to eosinopoietic cytokines in vitro, in general agreement with earlier observations on humans (Barr et al., 1987; Butterfield et al., 1986). Dexamethasone further potentiated the response of terminally differentiating precursors to IL-5. Both effects were synergistic, because dexamethasone did not induce colony formation nor eosinophil maturation by itself. The effects of dexamethasone were blocked by RU 486, suggesting the involvement of the glucocorticoid receptor (Perretti & Flower, 1996; Hardy et al., 1996).
Differently from human bone-marrow (Rinehart et al., 1997), dexamethasone effects had no obvious requirement for accessory cells. Also, unlike the reported effects of hydrocortisone on human eosinophil colony formation, which were highly dependent on the bone-marrow donor (Barr et al., 1987; Butterfield et al., 1986), we found stimulation of eosinopoiesis in bone-marrow cells from several unrelated inbred strains. In these experiments, C57BL/10 mice yielded higher numbers of colonies in the presence of GM-CSF than the other strains, in comparable conditions. Despite this interstrain variability, which is similar to that described by several laboratories (Roberts et al., 1997; Pozzulo et al., 1993; see Johnson, 1984, for a detailed analysis), dexamethasone had a significant enhancing effect on bone-marrow from all strains.
Our findings confirm and extend these earlier observations in humans, since we have documented an enhancing effect on eosinophil precursors, another potential glucocorticoid target. Taken together, our findings document both a nonselective stimulation of myelopoiesis (as has also been reported by Maruyama et al., 1999) and a selective stimulation of the eosinophil lineage (as described by Barr et al., 1997, and Butterfield et al., 1986, on human bone-marrow). Stimulation of myelopoiesis and of eosinopoiesis are not necessarily linked, as shown by their different dose-response profiles. Significant enhancement of eosinopoiesis in liquid and semi-solid cultures was still observed at dexamethasone concentrations that are insufficient to stimulate myelopoiesis (10−8 M and lower). On the other hand, synergism in stimulation of eosinophil precursors followed a bell-shaped dose-response curve, with the effect disappearing at high dexamethasone concentrations.
Despite stimulation of myelopoiesis, we have focused on the eosinophil lineage because of its relevance to allergy and asthma, which are treated with glucocorticoids. In this context, several studies have documented apoptosis by dexamethasone in mature eosinophils (mostly human, but also from other species), which is, however, prevented by low concentrations of eosinopoietic cytokines (as reviewed in Giembycz & Lindsay, 1999). It may, therefore, be asked whether dexamethasone induces apoptosis in murine bone-marrow eosinophil progenitors and precursors. By definition, eosinophil progenitors are not killed by dexamethasone, since the numbers of eosinophil colonies are increased. This closely parallels observations in human bone-marrow (Barr et al., 1987; Butterfield et al., 1985). Also, human peripheral blood eosinophil progenitors are not killed by dexamethasone (Shalit et al., 1995). We have no evidence of dexamethasone-induced killing of eosinophil precursors in liquid culture. EPO+ cells were viable, increased in number, and morphologically intact. Granule crystalloid ultrastructure indicated normal maturation (Giembycz & Lindsay, 1999). These observations agree entirely with those of Strath et al. (1985), who described intense and continuous eosinopoiesis in liquid culture of murine bone-marrow in the presence of hydrocortisone at 10−6 M. Furthermore, they also agree with observations by Das et al. (1997), who injected eosinopoenic doses of dexamethasone in allergic animals (50–100 μg mouse−1, approximately 15–30 mg kg−1) and observed increased numbers of bone-marrow eosinophils. Lefort et al. (1996) reported similar observations. Hence, dexamethasone does not kill murine bone-marrow eosinophil progenitors and precursors at concentrations in which it causes profound peripheral blood eosinopaenia and prevents eosinophil migration into inflammatory sites (Das et al., 1997).
Synergism between dexamethasone and the eosinopoietic cytokines persisted after ovalbumin-sensitization and challenge of BALB/c and BP-2 mice. The enhancing effect of dexamethasone on the frequency and size of eosinophil colonies seemed less evident in bone-marrow from sensitized and challenged mice, suggesting that the potentiating effects of glucocorticoids are partially lost as the animals become allergic. However, this issue requires further study before the significance of this observation is established.
At present, it is unclear whether the effects of dexamethasone include an increase in IL-5 receptor expression. In preliminary experiments using an antibody specific for IL-5 receptor (kindly provided by Drs R. Devos and J. Tavernier, Roche Research Gent, Belgium), we found increased numbers of receptor-positive cells in dexamethasone plus IL-5-stimulated liquid cultures (as expected from our EPO+ cell counts), but the intensity of staining per cell showed no obvious increase. At any rate, the effect of dexamethasone is not dependent upon modulation of the initial binding of IL-5 to its receptor, which is a very fast process (virtually complete within minutes, as shown by Mita et al., 1993), because dexamethasone added 24 or 48 h after bone-marrow was first stimulated with IL-5 is as effective as dexamethasone added simultaneously with IL-5 (unpublished observations). It may therefore act on cells that have already been stimulated by IL-5 to enhance their growth, as suggested by its positive influence on eosinophil colony growth and eosinophil precursor differentiation.
This work has focused on bone-marrow, rather than blood, because eosinopoiesis in normal individuals takes place predominantly in bone-marrow (as reviewed in Giembycz & Lindsay, 1999). While very small numbers of myeloid progenitors are found in blood of normal mice, their study requires methods distinct from those used in bone-marrow, because they must first be separated from the large numbers of mature blood components. Such studies can be carried out after the numbers of progenitors in peripheral blood are artificially increased by a number of treatments (see, for instance, Roberts et al., 1997), but they are difficult to carry out in normal animals, which were central to our study. Because of the focus on bone-marrow eosinopoiesis, we did not examine the number of mature eosinophils in the blood. However, the doses we used in vivo induce blood eosinopoenia in mice (Das et al., 1997), which is not necessarily related to our findings, because the passage of mature eosinophils from bone-marrow to blood is influenced by adhesive interactions with stromal and endothelial cells, as well as by chemokines and cytokines (reviewed in Giembycz & Lindsay, 1999). On the other hand, an increase in immature eosinophil in bone-marrow is not to be expected, because the production of eosinopoietic cytokines is inhibited by dexamethasone (as reviewed by Giembycz & Lindsay, 1999). However, as GM-CSF-dependent eosinophil progenitors and IL-5-responsive eosinophil precursors are demonstrably increased, when bone-marrow from dexamethasone-treated mice is cultured in the presence of exogenous cytokines, this indicates that dexamethasone has a priming effect on both targets, increasing bone-marrow responsiveness to eosinopoietic cytokines. This may be of relevance in conditions in which the synthesis of these cytokines becomes resistant to glucocorticoids (Kunicka et al., 1993; Her et al., 1991) or is induced by glucocorticoids (Wilckens & De Rijk, 1997; Agarwal & Marshall, 1998).
These results may contribute to the understanding of the effects of glucocorticoids in vivo, because they support a number of studies in which glucocorticoids contribute to allergic reactions, including the induction of a Th2 cytokine secretion pattern (Wilckens & De Rijk, 1997; Agarwal & Marshall, 1998) and enhancement of IgE-dependent immediate and late-phase skin reactions (Katayama et al., 1997). These observations may also be related to the extensive evidence for an association between stress and increased susceptibility to allergy (Michel, 1994; Parker, 1991), which has been recently linked to glucocorticoid effects (Agarwal & Marshall, 1998). Taken together, our results and those of the literature suggest that glucocorticoids have immunoregulatory effects that are not necessarily immunosuppressive, and which may play a role in the pathophysiology of allergic diseases.
Acknowledgments
Supported by grants from PAPES/FIOCRUZ, FINEP, CNPq/RHAE, INSERM/FIOCRUZ, CAPES-COFECUB and by Institut Pasteur/INSERM.
Abbreviations
- EPO
eosinophil peroxidase
- FCS
foetal calf serum
- GM-CSF
granulocyte-macrophage colony-stimulating factor
- IL-3
Interleukin-3
- IL-5
Interleukin-5
References
- AGARWAL S.K., MARSHALL G.D. Glucocorticoid-induced Type 1/Type 2 cytokine alterations in humans: a model for stress-related immune dysfunction. J. Interferon Cytokine Res. 1998;18:1059–1068. doi: 10.1089/jir.1998.18.1059. [DOI] [PubMed] [Google Scholar]
- BAGBY G.C.Hematopoiesis The Molecular Basis of Blood Diseases 1994W.B. Saunders, Philadelphia; 71–106.In: Stamatoyannopoulos, G., Nienhuis, A.W., Majerus, P.W., Varmus H. (eds.)(2nd. Ed.) [Google Scholar]
- BARNES P.J. Molecular mechanisms of steroid action in asthma. J. Allergy Clin. Immunol. 1996;97:159–168. doi: 10.1016/s0091-6749(96)80216-8. [DOI] [PubMed] [Google Scholar]
- BARR D.D., VOLARIC Z., KLIM J.B. Stimulation of human eosinophilopoiesis by hydrocortisone in vitro. Acta Haematol. 1987;77:20–24. doi: 10.1159/000205943. [DOI] [PubMed] [Google Scholar]
- BEGLEY C.G., BASSER R., MANSFIELD R., THOMSON B., PARKER W.R.L., LAYTON J., TO B., CEBON J., SHERIDAN W.P., FOX R.M., GREEN M.D. Enhanced levels and enhanced clonogenic capacity of blood progenitor cells following administration of stem cell factor plus granulocyte colony stimulating factor to humans. Blood. 1997;90:3378–3389. [PubMed] [Google Scholar]
- BUTTERFIELD J.H., ACKERMAN S.J., WEILER D., EISENBREY A.B., GLEICH G.J. Effects of glucocorticoids on eosinophil colony growth. J. Allergy Clin. Immunol. 1986;78:450–457. doi: 10.1016/0091-6749(86)90032-1. [DOI] [PubMed] [Google Scholar]
- CLUTTERBUCK E.J., HIRST E.M.A., SANDERSON C.J. Human Interleukin-5 (IL-5) regulates the production of eosinophils in human bone marrow cultures: comparison and interaction with IL-1, IL-3, IL-6, and GM-CSF. Blood. 1989;73:1504–1512. [PubMed] [Google Scholar]
- DAS A.M., FLOWER R.J., HELLEWELL P.G., TEIXEIRA M., PERRETTI M. A novel murine model of allergic inflammation to study the effect of Dexamethasone on eosinophil recruitment. Br. J. Pharmacol. 1997;121:97–104. doi: 10.1038/sj.bjp.0701122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EGESTEN A., CALAFAT J., WELLER P.F., KNOL E.F., JANSSEN H., WALZ T.M., OLSSON I. Localization of granule proteins in human eosinophil bone marrow progenitors. Int. Arch. Allergy Immunol. 1997;114:130–138. doi: 10.1159/000237657. [DOI] [PubMed] [Google Scholar]
- EUM S.Y., HAILÉ S., LEFORT J., HUERRE M., VARGAFTIG B.B. Eosinophil recruitment into the respiratory epithelium following antigenic challenge in hyper-IgE mice is accompanied by interleukin-5-dependent bronchial hyperresponsiveness. Proc. Natl. Acad. Sci. U.S.A. 1995;92:12290–12294. doi: 10.1073/pnas.92.26.12290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GASPAR ELSAS M.I., JOSEPH D., XAVIER ELSAS P., VARGAFTIG B.B. Rapid increase in bone marrow eosinophil production and responses to eosinopoietic interleukins triggered by intranasal allergen challenge. Amer. J. Resp. Cell. Mol. Biol. 1997;17:404–413. doi: 10.1165/ajrcmb.17.4.2691. [DOI] [PubMed] [Google Scholar]
- GIEMBYCZ M.A., LINDSAY M.A. Pharmacology of the Eosinophil. Pharmacol. Rev. 1999;51:213–339. [PubMed] [Google Scholar]
- GRUART V., TRUONG M.-J., PLUMAS J., ZANDECKI M., KUSNIERZ J.-P., PRIN L., VINATIER D., CAPRON A., CAPRON M. Decreased expression of eosinophil peroxidase and major basic protein messenger RNAs during eosinophil maturation. Blood. 1992;79:2592–2597. [PubMed] [Google Scholar]
- HALL T.L., FU Y.S. Applications of quantitative microscopy to tumor pathology. Lab. Invest. 1985;53:5–21. [PubMed] [Google Scholar]
- HARDY E., FARAHANI M., HALL I.P. Regulation of histamine H1 receptor coupling by dexamethasone in human cultured airway smooth muscle. Br. J. Pharmacol. 1996;118:1079–1084. doi: 10.1111/j.1476-5381.1996.tb15509.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HER E., FRAZER J., AUSTEN K.F., OWEN W.F. Eosinophil hematopoietins antagonize the programmed cell death of eosinophils. Cytokine and glucocorticoid effects on eosinophils maintained by endothelial cell-conditioned medium. J. Clin. Invest. 1991;88:1982–1987. doi: 10.1172/JCI115524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HISHA H., YAMADA H., SAKURAI M.H., KIYOHARA H., LI Y., YU C., TAKEMOTO N., KAWAMURA H., YAMAURA K., SHINOHARA S., KOMATSU Y., ABURADA M., IKEHARA S. Isolation and identification of hematopoietic stem cell-stimulating substances from Kampo (japanese herbal) medicine, Juzen-Taiho-To. Blood. 1997;90:1022–1030. [PubMed] [Google Scholar]
- HORTON M.A., LARSON K.A., LEE J.J., LEE N.A. Cloning of the murine eosinophil peroxidase gene (mEPO): characterization of a conserved subgroup of mammalian hematopoietic peroxidases. J. Leukocyte Biol. 1996;60:285–294. doi: 10.1002/jlb.60.2.285. [DOI] [PubMed] [Google Scholar]
- INMAN M.D., DENBURG J.A., ELLIS R., DAHLBÄCK M., O'BYRNE P.M. The effect of treatment with budesonide or PGE2 in vitro on allergen-induced increases in canine bone marrow progenitors. Am. J. Respir. Cell. Mol. Biol. 1997;17:634–661. doi: 10.1165/ajrcmb.17.5.2746. [DOI] [PubMed] [Google Scholar]
- JOHNSON G.R. Haemopoietic multipotential stem cells in culture. Clinics in Haematology. 1984;13:309–327. [PubMed] [Google Scholar]
- KATAYAMA I., IGAWA K., MINATOHARA K., NISHIOKA K. Topical glucocorticoid augments IgE-mediated passive cutaneous anaphylaxis in Balb/C mice and mast cell deficient WBB6F1 v/v mice. Clin. Exp. Allergy. 1997;27:1477–1483. doi: 10.1046/j.1365-2222.1997.1610974.x. [DOI] [PubMed] [Google Scholar]
- KUNICKA J.E., TALLE M.A., DENHARDT G.H., BROWN M., PRINCE L.A., GOLDSTEIN G. Immunosuppression by glucocorticoids: inhibition of production of multiple lymphokines by in vivo administration of dexamethasone. Cell. Immunol. 1993;149:39–49. doi: 10.1006/cimm.1993.1134. [DOI] [PubMed] [Google Scholar]
- LEFORT J., BACHELET C.M., LEDUC D, VARGAFTIG B.B. Effect of antigen provocation of IL-5 transgenic mice on eosinophil mobilization and bronchial hyperresponsiveness. J. Allergy Clin. Immunol. 1996;97:788–799. doi: 10.1016/s0091-6749(96)80157-6. [DOI] [PubMed] [Google Scholar]
- MARUYAMA S., MINAGAWA M., SHIMIZU T., OYA H., YAMAMOTO S., MUSHA N., ABO W., WEERASINGHE A., HATAKEYAMA K., ABO T. Administration of glucocorticoids markedly increases the numbers of granulocytes and extrathymic T cells in the bone marrow. Cell. Immunol. 1999;194:28–35. doi: 10.1006/cimm.1999.1492. [DOI] [PubMed] [Google Scholar]
- METCALF D. Lineage commitment in the progeny of murine hematopoietic preprogenitor cells: influence of thrombopoietin and Interleukin-5. Proc. Natl. Acad. Sci. U.S.A. 1998;95:6408–6412. doi: 10.1073/pnas.95.11.6408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MICHEL F.B. Psychology of the allergic patient. Allergy. 1994;49:28–30. doi: 10.1111/j.1398-9995.1994.tb04235.x. [DOI] [PubMed] [Google Scholar]
- MITA S., TAKAKI S., TOMINAGA A., TAKATSU K. Comparative analysis of the kinetics of binding and internalization of IL-5 in murine IL-5 receptors of high and low affinity. J. Immunol. 1993;151:6924–6932. [PubMed] [Google Scholar]
- OSKERITZIAN C., LE MAO J., DAVID B. Recombinant murine (rm) interleukin-5 enhances the eosinophil peroxidase content in cells cultured in vitro compared with rmIL-3 and granulocyte-macrophage colony-stimulating factor. Exp. Hematol. 1998;26:472–477. [PubMed] [Google Scholar]
- PARKER C.W. Environmental stress and immunity: possible implications for IgE-mediated allergy. Perspect. Biol. Med. 1991;34:197–212. doi: 10.1353/pbm.1991.0002. [DOI] [PubMed] [Google Scholar]
- PERRETTI M., FLOWER R.J. Measurement of lipocortin 1 levels in murine peripheral blood leukocytes by flow cytometry: modulation by glucocorticoids and inflammation. Br. J. Pharmacol. 1996;118:605–610. doi: 10.1111/j.1476-5381.1996.tb15444.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- POZZULO G.N., SKAMENE E., GERVAIS F. Bone marrow cell response following induction of acute inflammation in different strains of mice. Inflammation. 1993;17:677–685. doi: 10.1007/BF00920473. [DOI] [PubMed] [Google Scholar]
- RINEHART J., KEVILLE L., CLAYTON S., FIGUEROA J.A. Corticosteroids alter hematopoiesis in vitro by enhancing human monocyte secretion of granulocyte colony-stimulating factor. Exp. Hematol. 1997;25:405–412. [PubMed] [Google Scholar]
- ROBERTS A.W., FOOTE S., ALEXANDER W.S., SCOTT C., ROBB L., METCALF D. Genetic influences determining progenitor cell mobilization and leukocytosis induced by granulocyte colony-stimulating factor. Blood. 1997;89:2736–2744. [PubMed] [Google Scholar]
- SANDERSON C.J. Interleukin-5, eosinophils and disease. Blood. 1992;79:3101–3109. [PubMed] [Google Scholar]
- SCHLEIMER R.P., BOCHNER B.S. The effects of glucocorticoids on human eosinophils. J. Allergy Clin. Immunol. 1994;94:1202–1213. doi: 10.1016/0091-6749(94)90333-6. [DOI] [PubMed] [Google Scholar]
- SCHIBLER K.R., LI Y., OHLS R.K., NYE N.C., DURHAM M.C., WHITE W., LIECHTY K.W., LE T., CHRISTENSEN R.D. Possible mechanisms accounting for the growth factor independence of hematopoietic progenitors from umbilical cord blood. Blood. 1994;84:3679–3684. [PubMed] [Google Scholar]
- SHALIT M., SEKHSARIA S., MALECH H.L. Modulation of growth and differentiation of eosinophils from human peripheral blood CD34+ cells by IL5 and other growth factors. Cell. Immunol. 1995;160:50–57. doi: 10.1016/0008-8749(95)80008-7. [DOI] [PubMed] [Google Scholar]
- SLOVICK F.T., ABBOUD C.N., BRENNAN J.K., LICHTMAN M.A. Modulation of in vitro eosinophil progenitors by hydrocortisone: role of accessory cells and interleukins. Blood. 1985;66:1072–1079. [PubMed] [Google Scholar]
- STRATH M., WARREN D., SANDERSON C.J. Detection of eosinophils using an EPO assay. Its use as an assay for eosinophil differentiation factors. J. Immunol. Methods. 1985;83:209–215. doi: 10.1016/0022-1759(85)90242-x. [DOI] [PubMed] [Google Scholar]
- TEN R.M., PEASE L.R., MCKEAN D.J., BELL M.P., GLEICH G.J. Molecular cloning of human eosinophil peroxidase. evidence for the existence of a peroxidase multigene family. J. Exp. Med. 1989;169:1757–1769. doi: 10.1084/jem.169.5.1757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WILCKENS T., DE RIJK R. Glucocorticoids and immune functions: unknown dimensions and new frontiers. Immunol. Today. 1997;18:418–424. doi: 10.1016/s0167-5699(97)01111-0. [DOI] [PubMed] [Google Scholar]