

Abstract
Rasagiline [N-propargyl-1R(+)-aminoindan], was examined for its monoamine oxidase (MAO) A and B inhibitor activities in rats together with its S(−)-enantiomer (TVP 1022) and the racemic compound (AGN-1135) and compared to selegiline (1-deprenyl). The tissues that were studied for MAO inhibition were the brain, liver and small intestine.
While rasagiline and AGN1135 are highly potent selective irreversible inhibitors of MAO in vitro and in vivo, the S(−) enantiomer is relatively inactive in the tissues examined.
The in vitro IC50 values for inhibition of rat brain MAO activity by rasagiline are 4.43±0.92 nM (type B), and 412±123 nM (type A). The ED50 values for ex vivo inhibition of MAO in the brain and liver by a single dose of rasagiline are 0.1±0.01, 0.042±0.0045 mg kg−1 respectively for MAO-B, and 6.48±0.81, 2.38±0.35 mg kg−1 respectively for MAO-A.
Selective MAO-B inhibition in the liver and brain was maintained on chronic (21 days) oral dosage with ED50 values of 0.014±0.002 and 0.013±0.001 mg kg−1 respectively.
The degree of selectivity of rasagiline for inhibition of MAO-B as opposed to MAO-A was similar to that of selegiline. Rasagiline was three to 15 times more potent than selegiline for inhibition of MAO-B in rat brain and liver in vivo on acute and chronic administration, but had similar potency in vitro.
These data together with lack of tyramine sympathomimetic potentiation by rasagiline, at selective MAO-B inhibitory dosage, indicate that this inhibitor like selegiline may be a useful agent in the treatment of Parkinson's disease in either symptomatic or L-DOPA adjunct therapy, but lack of amphetamine-like metabolites could present a therapeutic advantage for rasagiline.
Keywords: Monoamine oxidase (MAO) A and B, brain, liver, small intestine, selegiline (l-deprenyl), clorgyline, rasagiline, irreversible inhibitors, Parkinson's disease, dopamine
Introduction
The knowledge that dopamine is equally well oxidatively deaminated by monoamine oxidase (MAO) types A and B (Collins et al., 1970), the dominance of MAO-B (80%) as compared to MAO-A in the extrapyramidal regions of human brain (Collins et al., 1970; Squires, 1972; O'Carroll et al., 1983) and the absence of the ‘cheese reaction' in whole animal and isolated tissue preparations by the selective irreversible MAO-B inhibitor selegiline (deprenyl; Knoll & Magyar, 1972) led to the introduction of selegiline as an adjunct to L-DOPA therapy of Parkinson's disease (Birkmayer et al., 1975; 1977; Lees et al., 1977). Selegiline has proved to be a useful antiparkinson drug both in monotherapy (Parkinson Study Group, 1989) and as adjunct to L-DOPA therapy, and has L-DOPA sparing action (Birkmayer et al., 1977; Riederer & Rinne, 1992). Selegiline is a propargyl derivative of 1-amphetamine. It irreversibly inhibits MAO-B by binding mole per mole covalently to the N5 position of the isoalloxazine moiety of FAD, the cofactor of MAO-B (Maycock et al., 1976; Youdim, 1978). It is metabolized in vivo to 1-amphetamine and to 1-methamphetamine (Reynolds et al., 1978) and has amphetamine-like sympathomimetic actions (Simpson, 1978; Finberg et al., 1981).
In this paper we report on the in vitro and acute and chronic in vivo inhibitory activity of a highly potent and selective inhibitor of MAO-B, rasagiline [N-propargyl-1R(+)-aminoindan; TVP-1012; Youdim et al., 1995; Sterling et al., 1998] with structural similarity to selegiline (Sabbagh & Youdim, 1978; Kalir et al., 1981; see Figure 1. Rasagiline has been developed as an antiparkinson drug (Youdim et al., 1995; Finberg et al., 1996; 1999) and is an analogue of selegiline, but unlike the latter is metabolized to aminoindan, which is largely devoid of amphetamine-like properties. It was isolated from the racemic form of the selective MAO-B inhibitor, AGN1135 (Sabbagh > Youdim, 1978; Kalir et al., 1981), since the latter drug was shown by us to be devoid of sympathomimetic activity and does not produce the ‘cheese reaction' in isolated tissue preparations or in vivo in rats and cats in doses selective for inhibition of MAO-B (Finberg et al., 1981; Finberg & Youdim, 1985).
Figure 1.
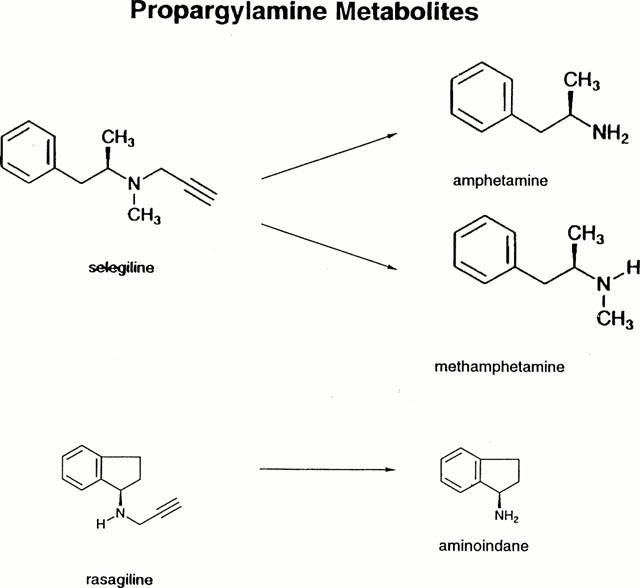
Chemical structures of rasagiline, selegiline (deprenyl) and their metabolites.
Methods
Animals and drug treatments
Male Sprague-Dawley rats (Charles-River) were housed in wire-mesh cages at an environmental temperature of 19 – 21°C, 12 h light – dark cycle (lights on at 0700), and were fed rat chow ad libitum for the duration of the experiment.
Determination of MAO inhibitory activity in vitro
The activities of MAO-A and -B were determined by the adapted method of Tipton & Youdim (1983). Rat or human cerebral cortical tissue was homogenized in 0.3 M sucrose (one part tissue to 20 parts sucrose) using a glass-teflon motor-driven homogenizer (brain and liver), or Ultraturrax (gut). The inhibitor under test was added to a suitable dilution of the enzyme preparation in 0.05 M phosphate buffer (pH 7.4) and incubated together with selegiline 0.1 μM (for determination of MAO-A) or clorgyline 0.1 μM (for determination of MAO-B). Incubation was carried on for 60 min at 37°C before addition of labelled substrates (14C-5-hydroxytryptamine creatinine disulphate 100 μM for determination of MAO-A, or 14C-phenylethylamine 10 μM for determination of MAO-B) and incubation continued for 30 or 20 min respectively. The reaction was then stopped by addition of citric acid (2 M). Radioactive metabolites were extracted into toluene/ethyl acetate (1 : 1 v v−1), a solution of 2,5-diphenyloxazole was added to a final concentration of 0.4% (w v−1), and metabolite content estimated by liquid scintillation counting. Activity in presence of drug was expressed as a percentage of that in control samples.
The preincubation was carried out in the presence of clorgyline or selegiline because phenylethylamine is also metabolized quite effectively by MAO-A (O'Carroll et al., 1983), leading to inhibition curves for MAO-B, which showed a plateau at about 80% inhibition with selegiline or rasagiline if MAO-A was not inactivated. For comparison between two inhibitors with potentially different inhibitory effects on MAO-A and MAO-B, therefore, it was thought necessary to employ the system in which opposite enzyme forms are inactivated before assay.
Determination of inhibition of MAO activity in vivo
In in vivo studies, drugs were administered orally by gavage (p.o.). The animals weighed 250 – 300 g at the time of killing. For estimation of in vivo inhibitory effect, varying doses of the inhibitors were administered to groups of five or six rats for the stated times, the animals were killed by decapitation, tissues removed and frozen at −20°C, and enzyme activity determined subsequently as above. Enzyme activity in drug-treated tissues were expressed as a percentage of that in control tissues.
Protein determination
Protein determination for standardization of MAO activity was performed by the method of Lowry et al. (1951) employing human serum albumin as standard.
Statistical analysis
Values of IC50 and ED50 in in vitro and in vivo analyses were calculated in each separate experimental run by non-linear regression analysis using the Graphpad Prism programme, and the mean values of all runs for each drug treatment and tissue were compared using one-way ANOVA, with Tukey-Kramer post-hoc test. A value of P=0.05 was taken as cut-off for significance.
Materials
N-propargyl-1R(+)-aminoindan (rasagiline, TVP1012), N-propargyl-1S(−)-aminoindan (TVP1022), the racemic form of rasagiline (AGN-1135), and selegiline [(−)-deprenyl]hydrochloride were generously supplied by Teva (Israel). The radioisotopes β-[ethyl-1-14C]-phenylethylamine hydrochloride and 5-[2-14C]-hydroxytryptamine binoxalate were obtained from New England Nuclear (Boston, U.S.A.) at a specific activity in the range 1.48 – 2.22 GBq mmol−1. All other reagents and chemicals were analytical grade and were purchased from either Sigma (Israel) or Merck (Germany).
Results
Determination of MAO-A and -B inhibition in vitro
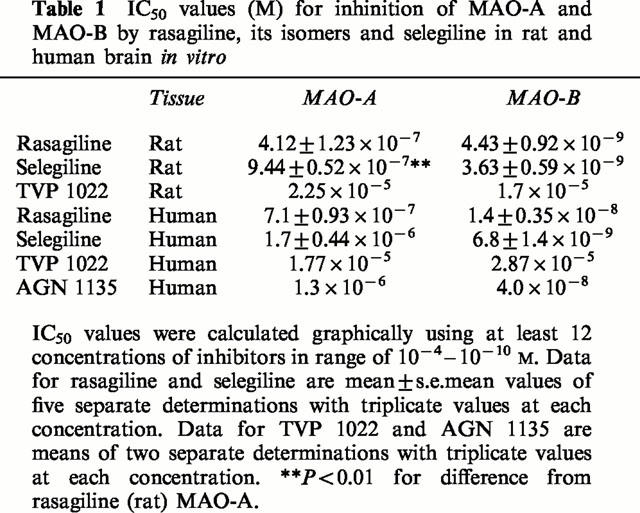
MAO-A and -B activities were determined in rat brain homogenate in vitro following incubation with varying concentrations of rasagiline, TVP1022, and selegiline (Figure 2 and Table 1). The IC50 values calculated for each inhibitor are presented in Table 1. The optical enantiomers of N-propargyl-1-aminoindan can be seen to possess a striking degree of stereoselectivity, since the (−)-isomer (TVP 1022) is 1/3,800 as active as the (+)-isomer (rasagiline) for inhibition of MAO-B. Whereas rasagiline is 93 times more potent for inhibition of MAO-B than MAO-A, TVP 1022 shows little or no selectivity for MAO-A or -B. Rasagiline and selegiline had similar IC50 values for inhibition of MAO-B (P>0.05), but selegiline was significantly less potent than rasagiline for inhibition of MAO-A (P<0.01). In human tissue, there was no significant difference between the IC50 values for rasagiline and selegiline for inhibition of MAO-A or MAO-B (Table 1), and TVP 1022 was again seen to be largely devoid of inhibitory activity. The racemic form of N-propargyl-1-aminoindan (AGN 1135) had approximately half the potency of rasagiline, indicating that no interaction occurred between the optical isomers at the MAO active site (similar data were obtained in preliminary studies using rat tissue).
Figure 2.
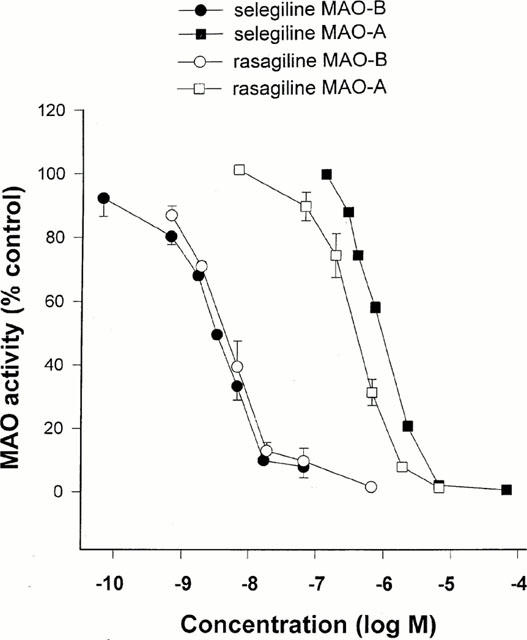
In vitro inhibitory action of rasagiline and selegiline against rat brain MAO. The test compounds were added to buffer containing 10−7 M clorgyline (for assay of MAO-B) or 10−7 M selegiline (for assay of MAO-A) and were incubated with the tissue homogenate for 60 min at pH 7.4 before addition of 14C-β-phenylethylamine or 14C-5-hydroxytryptamine respectively.
Table 1.
IC50 values (M) for inhinition of MAO-A and MAO-B by rasagiline, its isomers and selegiline in rat and human brain in vitro
Determination of brain and liver MAO-A and -B inhibition ex vivo
Acute p.o. administration
The MAO inhibitory activities of rasagiline and selegiline were compared in rats 2 h after a single oral dose (0.01 – 100 mg kg−1); inhibition curves are shown in Figure 3, and ED50 values are shown in Table 2. A comparison of potency between these inhibitors is difficult due to the fact that the slopes of the dose-response curves were rather different (Figure 3), however from ED50 values it can be seen that rasagiline was significantly more potent than selegiline for inhibition of both brain and liver MAO-A and MAO-B (Table 2). A clinically-relevant degree of inhibition of MAO-B (i.e. over 90%) with minimal inhibition of MAO-A was produced by a dose of 0.5 mg kg−1 rasagiline, but for selegiline a dose of 5 mg kg−1 was required. Similar inhibition curves were obtained for the liver enzymes, demonstrating that rasagiline maintains its selectivity for MAO-B inhibition on oral administration, both in brain and periphery. From comparison of inhibition curves shown in Figure 3 it can be seen that the selectivity of rasagiline for inhibition of MAO-B over MAO-A was similar to that of selegiline.
Figure 3.
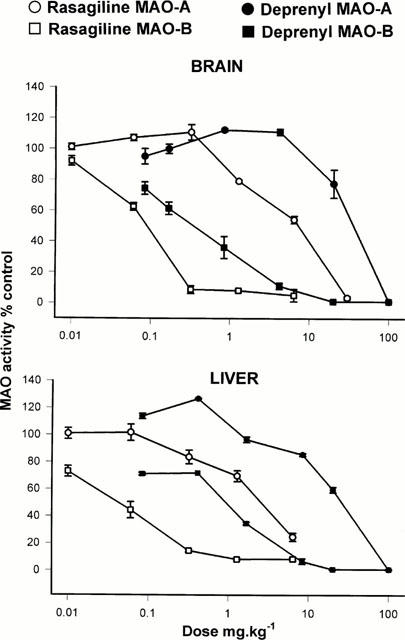
In vivo MAO inhibitory activity of rasagiline and selegiline in rat brain and liver following a single dose of the compound administered orally. Groups of rats (3 – 5 at each dose level) were injected with the doses of the compounds shown and sacrificed 2 h later. Activity of MAO was determined in a suitably diluted sucrose homogenate of whole brains (minus cerebellum) and liver using 14C-β-phenylethylamine or 14C-5-hydroxytryptamine for assay of MAO-B or MAO-A activity respectively. Enzyme activity in tissues from drug-treated animals was expressed as a percentage of the enzyme activity in tissues of animals treated with normal saline. Mean values shown±s.e.mean.
Table 2.
ED50 values for inhibition of rat brain and liver MAO-A and MAO-B by rasagiline and selegiline in vivo

Chronic p.o. administration
Rats given oral rasagiline and selegiline at the doses indicated in Figure 4, one dose daily for 21 days, were decapitated 24 h after the last dose and brains and livers were examined for their MAO activity. Since the higher doses were omitted in this experiment, determination of ED50 was not feasible in every case, but those values with could be obtained are shown in Table 2B. Again rasagiline was significantly more potent than selegiline (from ED50 data) for inhibition of MAO-B in brain and liver by a factor of 5 and 3 respectively, however, a clinically relevant degree of brain MAO-B (84±2%, n=6) was produced by a dose of 0.05 mg kg−1 daily rasagiline, whereas the same degree of inhibition (77±4%, n=6) required a dose of 0.5 mg kg−1 selegiline. The inhibitory potency of the racemic form of rasagiline was also examined in similar dose ranges, and rasagiline was twice as active as the racemate (data not shown). In the liver similar data for MAO-A and -B inhibition were obtained for both inhibitors with rasagiline showing greater selectivity of MAO-B inhibition at doses lower than 1 mg kg−1 (Figure 4).
Figure 4.
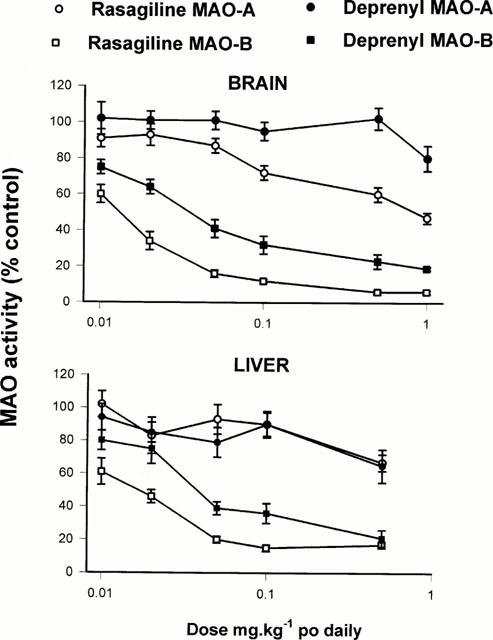
In vivo MAO inhibitory activity of rasagiline and selegiline in rat brain and liver following chronic treatment with the compounds p.o. Groups of rats (4 – 6 at each dose level) were treated daily by gavage with the doses of the compounds shown for 21 days and sacrificed 24 h after the last dose. Activity of MAO was determined in suitable dilutions of a sucrose homogenate of whole brain (minus cerebellum) or liver using 14C-β-phenylethylamine or 14C-5-hydroxytryptamine for assay of MAO-B or MAO-A activity respectively. Enzyme activity in drug-treated rats is expressed as a percentage of the enzyme activity in brain or liver tissue of control animals treated with normal saline. Mean values shown±s.e.mean.
The effect of rasagiline on small intestine MAO
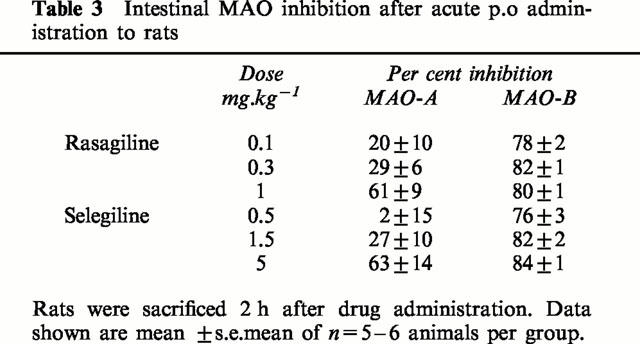
MAO-A and -B activities were estimated in the small intestine 2 h after an oral dose of rasagiline or selegiline. Selectivity of inhibition of MAO-B to MAO-A was seen also in this tissue (Table 3).
Table 3.
Intestinal MAO inhibition after acute p.o administration to rats
Recovery of MAO-A and -B activities after i.p. and chronic p.o. rasagiline
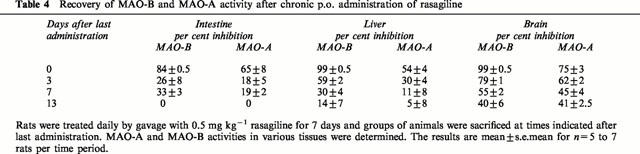
In a chronic study where rasagiline was given at an oral dose of 0.5 mg kg−1 for 7 days once daily, brain MAO-B and MAO-A were inhibited by 79±1% and 62±2% (n=6) 3 days after the last dose. Both enzymes in the brain were still inhibited by some 40% after 13 days, but recovery of enzyme activity in the small intestine and liver was more rapid (Table 4).
Table 4.
Recovery of MAO-B and MAO-A activity after chronic p.o. administration of rasagiline
Discussion
In this paper we have examined the in vitro and in vivo MAO-A and -B inactivating properties of rasagiline, the R(+) enantiomer of the selective irreversible MAO-B inhibitor AGN 1135 (Kalir et al., 1981), its S(−) enantiomer (TVP1022) and the racemic compound AGN1135. The inhibitory potency of rasagiline has been examined in brain, liver and intestine of rats after acute and chronic oral treatment and compared to the selective irreversible MAO-B inhibitor, selegiline (Knoll & Magyer, 1972). The results demonstrate a high degree of stereoselectivity for MAO inhibition, in that the R(+) enantiomer (rasagiline) was three orders of magnitude more effective in inhibition of MAO-B than the S(−) enantiomer. Selegiline, by comparison, shows only a small degree of stereoselectivity, and in this case it is the S(−) enantiomer which is more potent than the R(+) enantiomer. The full explanation for this markedly different degree of stereoselectivity between closely allied chemical compounds will only be apparent when three-dimensional modelling of the MAO active site is available.
Used in Parkinson's Disease, selegiline is believed to potentiate the pharmacological action of dopamine formed from L-DOPA, since in human brain dopamine is equally well metabolized by both MAO-A and -B (Green et al., 1977; Maitre et al., 1976; O'Carroll et al., 1983). The latter enzyme is enriched in the extrapyramidal regions of human brain (Collins et al., 1970; O'Carroll et al., 1983) and primate brain (Gotz et al., 1998) including the caudate nucleus, globus pallidus, substantia nigra and thalamus. Rasagiline, like selegiline, is a propargyl containing drug and is an analogue of the latter. However, the propargyl component in rasagiline unlike that of selegiline is bound to an aminoindan moiety. Whereas the major metabolite of 1-selegiline is 1-methamphetamine (Reynolds et al., 1978) rasagiline gives rise to aminoindan derivatives (Speiser et al., 1998).
The propargyl-containing MAO inhibitors such as clorgyline and selegiline are known as suicide inhibitors since they are substrates of the enzyme (Maycock et al., 1976). They inactivate MAO-A and -B selectively and irreversibly by interacting covalently with the cysteinyl-FAD cofactor of the enzymes at their active centers (Kearney et al., 1971; Bach et al., 1988). This interaction takes place between the propargyl group and the N5 of the FAD isoalloxazine component of liver (Maycock et al., 1976) and brain MAO (Salach et al., 1976). It has been shown for these inhibitors that 1 mole of the inhibitor inactivates 1 mole of the enzyme (Salach et al., 1978) and since rasagiline has the same enzyme inactivating moiety as that of clorgyline, selegiline and pargyline, its most likely mechanism of MAO inactivation is identical to these inhibitors
The present study has demonstrated that rasagiline, like selegiline, is an irreversible inhibitor of MAO-B. This was demonstrated in experiments where rasagiline MAO inhibitory activity was examined in vitro and in vivo when it was given p.o. and MAO-A and -B were then estimated ex vivo in various tissues, at time intervals up to 13 days after rasagiline treatment. It is apparent that rasagiline is a very potent selective MAO-B inhibitor and has a good uptake across the blood-brain barrier, as shown by the similarity of inhibition curves between liver and brain. Although when compared in vitro, rasagiline had similar potency to selegiline for inhibition of MAO-B, the in vivo study showed a greater potency of rasagiline. This greater potency of rasagiline is even more marked if, instead of 50% enzyme inhibition, the dose required for 80% inhibition is measured. The reason for this is not currently known, but may be due to different rates of metabolism of the parent compounds in vivo, or to improved tissue penetration of rasagiline. Interestingly, preliminary studies in humans show an approximately 5 fold greater potency for rasagiline over selegiline for inhibition of platelet MAO-B (unpublished data). Although rasagiline has a greater potency than selegiline, its selectivity for MAO-A and -B inhibition is very similar to what has been reported for selegiline. However, in contrast to selegiline which does not show selectivity between its optical isomers for inhibition of MAO-A and -B, AGN 1135 shows roughly 4 and 2 orders of magnitude between its optical isomers for inhibition of MAO-B and -A respectively.
The present results complement findings in non-human primate (monkey) brains (Gotz et al., 1998) where rasagiline was given chronically for 7 days at various doses and MAO-A and -B activities were measured in several brain regions, including caudate nucleus, globus pallidus, cerebral cortex and hippocampus. Rasagiline was shown to be a potent selective inhibitor of MAO-B in the caudate nucleus and globus pallidus where the activity of MAO-B is 4 fold higher than that of MAO-A (Gotz et al., 1998).
The recovery of the MAO-A and -B activities after in vivo inhibition, which is related to the synthesis of enzyme apoprotein, differs between the tissues (liver, intestine and brain) examined. The small intestine MAO-B activity has the fastest recovery, while the brain MAO-B activity shows the slowest recovery. These differences in rat tissue enzyme activity recovery after rasagiline treatment, are not unusual since similar findings have been reported for enzyme recovery after inhibition by selegiline and clorgyline (Neff & Goridis, 1972; Della Corte & Tipton, 1980). Indeed, in primate (monkeys and human) brains the half-life for recovery of MAO-B after selegiline treatment has been reported to be well over 30 days (Fowler et al., 1994), and for the rat brain 13 days (Neff & Goridis, 1972; Della Corte & Tipton, 1980).
In conclusion, the present study has shown that rasagiline is a potent irreversible inhibitor of MAO-B and it is 3 – 15 times more potent than selegiline in the rat in vivo with a similar selectivity for inhibition of MAO-B to MAO-A. Because of its cleaner pharacological profile, with absence of amphetamine-like properties, formation of the metabolite aminoindan rather than 1-methamphetamine, and recently described neuroprotective and antiapoptotic properties (Finberg et al., 1998; Huang et al., 1999; Youdim et al., 1999), we can conclude that this drug may have a preferential activity to that of selegiline in the treatment of Parkinson's disease.
Acknowledgments
We wish to thank Teva Pharmaceutical Co (Israel) for generous support of this work and the supply of rasagiline. We wish to thank Rachel Freedman, Dr Gila Lavian, and Dr Ester Pirak for their excellent technical assistance at various stages of this project.
Abbreviations
- AGN 1135
racemic N-propargyl-1-aminoindan
- ANOVA
analysis of variance
- ED50
50% effective dose
- FAD
flavine adenine dinucleotide
- ID50 50% inhibitory concentration; L-DOPA
L-dihydroxyphenylalanine
- MAO
monoamine oxidase
- TVP 1012
N-propargyl-1R(+)aminoindan
- TVP 1022
N-propargyl-1S(−)-aminoindan
References
- BACH A.W.J., LAN N.C., JOHNSON D.L, , ABELL C.W., BEMBEREK M.E., KWAN S.W., SEEBERG P.H., SHIH J. cDNA cloning of human liver monoamine oxidase A and B: Molecular basis of differences in enzymatic properties. Proc. Nat. Acad. Sci. U.S.A. 1988;85:4934–4938. doi: 10.1073/pnas.85.13.4934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BIRKMAYER W., RIEDERER P., AMBROZI L., YOUDIM M.B.H. Implications of combined treatment with Madopar and 1-deprenyl in Parkinson's disease. Lancet. 1977;2:439–443. doi: 10.1016/s0140-6736(77)91940-7. [DOI] [PubMed] [Google Scholar]
- BIRKMAYER W., RIEDERER P., YOUDIM M.B.H., LINAUER W. The potentiation of anti-akinetic effect after L-dopa treatment by an inhibitor of MAO-B, deprenyl. J. Neural Transm. 1975;36:303–326. doi: 10.1007/BF01253131. [DOI] [PubMed] [Google Scholar]
- COLLINS G.G.S., SANDLER M., WILLIAMS E.D., YOUDIM M.B.H. Multiple forms of human brain mitochondrial monoamine oxidase. Nature. 1970;225:817–820. doi: 10.1038/225817a0. [DOI] [PubMed] [Google Scholar]
- DELLA CORTE L., TIPTON K.F. The turnover of the A and B-forms of monoamine oxidase in rat liver. Biochem. Pharmacol. 1980;29:811–815. doi: 10.1016/0006-2952(80)90219-1. [DOI] [PubMed] [Google Scholar]
- FINBERG J.P.M., YOUDIM M.B.H. Modification of blood pressure and nictitating membrane response to sympathetic amines by selective monoamine oxidase inhibitors of types A and B in the cat. Br. J. Pharmacol. 1985;85:541–546. doi: 10.1111/j.1476-5381.1985.tb08891.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FINBERG J.P.M., LAMENSDORF I., COMMISSIONG J.W., YOUDIM M.B.H. Pharmacology and neuroprotective properties of rasagiline. J. Neural. Transm. 1996;48 Suppl.:95–103. doi: 10.1007/978-3-7091-7494-4_9. [DOI] [PubMed] [Google Scholar]
- FINBERG J.P.M., LAMENSDORF I., WEINSTOCK M., SCHWARTZ M., YOUDIM M.B.H. Pharmacology of rasagiline (N-propargyl-IR-aminoindan) Adv. Neurol. 1999;80:495–501. [PubMed] [Google Scholar]
- FINBERG J.P.M., TAKESHIMA T., JOHNSTON J.M., COMMISSIONG J.W. Increased survival of dopaminergic neurons by rasagiline, a monoamine oxidase B inhibitor. Neuroreport. 1998;9:703–707. doi: 10.1097/00001756-199803090-00026. [DOI] [PubMed] [Google Scholar]
- FINBERG J.P.M., TENNE M., YOUDIM M.B.H. Tyramine antagonistic properties of AGN 1135, an irreversible inhibitor of monoamine oxidase type B. Br. J. Pharmacol. 1981;73:65–74. doi: 10.1111/j.1476-5381.1981.tb16772.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FOWLER J.S., VOLKOW N.D., LOGAN J., WANG G.J., MACGREGOR R.R., SCHYLER D., WOLF A.D., PAPPAS M., ALEXOFF D., SHEA C. Slow recovery of human brain MAO B after deprenyl (selegiline) withdrawal. Synapse. 1994;18:86–93. doi: 10.1002/syn.890180203. [DOI] [PubMed] [Google Scholar]
- GOTZ M.E., BREITHAUPT W., SAUTTER J., KUPSCH A., SCHWARZ J., OERTEL W.H., YOUDIM M.B.H., RIEDERER P., GERLACH R.Y. Chronic TVP-1012 (rasagiline) dose-activity response of monoamine oxidases A and B in the brain of the common marmoset. J Neural Transm. 1998;52 Suppl.:271–278. doi: 10.1007/978-3-7091-6499-0_27. [DOI] [PubMed] [Google Scholar]
- GREEN A.R., TORDOFF A., MITCHEL B., YOUDIM M.B.H. Evidence for dopamine deamination by both type A and type B monoamine oxidase in rat brain in vivo and for degree of inhibition of enzyme necessary for increased function and activity of dopamine and 5-hydroxytryptamine. Br. J. Pharmacol. 1977;60:343–349. doi: 10.1111/j.1476-5381.1977.tb07506.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HUANG W., CHEN Y., SHOHAMI E., WEINSTOCK M. Neuroprotective effect of rasagiline, a selective monoamine oxidase B inhibitor, against head injury in the mouse. Eur. J. Pharmacol. 1999;366:127–135. doi: 10.1016/s0014-2999(98)00929-7. [DOI] [PubMed] [Google Scholar]
- KALIR A., SABBAGH A., YOUDIM M.B.H. Selective ‘suicide' acetylenic and reversible inhibitors of monoamine oxidase A and B. Br. J. Pharmacol. 1981;73:55–64. doi: 10.1111/j.1476-5381.1981.tb16771.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KEARNEY F.B., SALACH J.I., WALKER W.H., SENG R.L., KENNEY W., ZESZOLEK E., SINGERT P. The covalently bound flavin of hepatic monoamine oxidase I. Isolation and sequence of a flavin peptide and evidence for binding to the 8∞-position. Eur. J. Biochem. 1971;24:321–327. doi: 10.1111/j.1432-1033.1971.tb19689.x. [DOI] [PubMed] [Google Scholar]
- KNOLL J., MAGYAR K. Some puzzling pharmacological effects of monoamine oxidase inhibitors. Adv. Biochem. Psychopharmacol. 1972;5:393–408. [PubMed] [Google Scholar]
- LEES A., SHOW K.M., KAHOUT L.J., STERN G., ELSWORTH J.D., SANDLER M., YOUDIM M.B.H. Deprenyl in Parkinson's Disease. Lancet. 1977;(ii):791–795. doi: 10.1016/s0140-6736(77)90725-5. [DOI] [PubMed] [Google Scholar]
- LOWRY O.G., ROSEBROUGH N.J., FARR A.L., RANDALL R.J. Protein measurement with Folin phenol reagent. J. Biol. Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- MAITRE L., DELINI-STULA A., WALDMEIER P.C.Relation between the degree of monoamine oxidase inhibition and some psychopharmacological responses to monoamine oxidase inhibitors 1976Amsterdam: Elsevier; 247–267.Ciba Foundation Symposium No. 39 [Google Scholar]
- MAYCOCK A.L., ABELES R.H., SALACH J., SINGER T.P. The action of acetylenic inhibitors on mitochondrial monoamine oxidase: Structure of the flavin site in the inhibited enzyme. 1976Amsterdam: Elsevier; 33–49.Ciba Foundation Symposium No. 39 [Google Scholar]
- NEFF N.H., GORDIS C. Neuronal monoamine oxidase: Specific enzyme types and their rates of formation. Adv. Biochem. Psychopharmacol. 1972;5:307–324. [PubMed] [Google Scholar]
- O'CARROLL A.M., FOWLER C.J., PHILLIP J.P., TOBIA I., TIPTON K.F. The deamination of dopamine by human brain monoamine oxidase. Specificity for the two forms in seven brain regions. Naunyn Schmiedebergs Arch. Pharmacol. 1983;322:198–207. doi: 10.1007/BF00500765. [DOI] [PubMed] [Google Scholar]
- PARKINSON STUDY GROUP (DATATOP) Selegiline and Parkinson's disease. N. Engl. J. Med. 1989;321:1364–1371. doi: 10.1056/NEJM198911163212004. [DOI] [PubMed] [Google Scholar]
- REYNOLDS G.P., RIEDERER P., SANDLER N.R., JELLINGER K., SEEMALI D. Amphetamine and 2-phenylethylamine in post mortem Parkinson brains after 1-deprenyl administration. J. Neural. Transm. 1978;43:271–278. doi: 10.1007/BF01246964. [DOI] [PubMed] [Google Scholar]
- RIEDERER P., RINNE U.K. Selegiline in Parkinson's Disease: An update. Mov. Disords. 1992;8(Suppl. 1):51–544. [Google Scholar]
- SABBAGH A., YOUDIM M.B.H. Selective inhibition of monoamine oxidase type B by propargyl-containing drugs. Israel J. Med. Sci. 1978;14:1097. [Google Scholar]
- SALACH J., DETMER K., YOUDIM M.B.H. The reaction of bovine and rat liver monoamine oxidase with [14C]-clorgyline and [14C]-deprenyl. Mol. Pharmacol. 1978;16:234–241. [PubMed] [Google Scholar]
- SALACH J., SINGER T.P., YAUNOBU K.T., MONAMURA N., YOUDIM M.B.H.Cysteinyl flavin in monoamine oxidase from the central nervous system 1976Amsterdam: Elsevier; 49–57.Ciba Foundation Symposium No. 39 pp [Google Scholar]
- SIMPSON L.L. Evidence that deprenyl, a type B monoamine oxidase inhibitor is an indirectly acting sympathomimetic amine. Biochem. Pharmacol. 1978;27:1591–1595. doi: 10.1016/0006-2952(78)90490-2. [DOI] [PubMed] [Google Scholar]
- SPEISER Z., LEVY R., COHEN S. Effect of N-propargyl-1R-aminoindan (rasagiline) in models of motor and cognition disorders. J. Neural Transm., Suppl. 1998;52:287–300. doi: 10.1007/978-3-7091-6499-0_29. [DOI] [PubMed] [Google Scholar]
- SQUIRES R.F. Multiple forms of monoamine oxidase in intact mitochondria as characterized by selective inhibitors and thermal stability. Adv. Biochem. Psychopharmacol. 1972;5:355–370. [PubMed] [Google Scholar]
- STERLING J., VEINBERG A., LERNER D., GOLDENBERG W., LEVY R., YOUDIM M.B.H., FINBERG J.P.M. (R)(+)-N-Propargyl-1-aminoindan (rasagiline) and derivatives: highly selective and potent inhibitors of monoamine oxidase B. J. Neural. Transm. 1998;52(Suppl):301–305. doi: 10.1007/978-3-7091-6499-0_30. [DOI] [PubMed] [Google Scholar]
- TIPTON K.F., YOUDIM M.B.H.The assay of monoamine oxidase activity Methods in Biogenic Amine Research 1983Amsterdam: Elsevier; 441–467.Parvez, S. Nagatsu, T., Nagatsu, I. & Parvez, H. (eds). pp [Google Scholar]
- YOUDIM M.B.H. The active centers of monoamine oxidase types A and B; binding with 14C-clorgyline and 14C-deprenyl. J. Neural. Transm. 1978;43:199–208. doi: 10.1007/BF01246956. [DOI] [PubMed] [Google Scholar]
- YOUDIM M.B.H., FINBERG J.P.M., LEVY R., STERLING J., LERNER D., BERGER-PASKIN T., YELLIN H. R-Enantiomers of N-propargyl-amino indian compounds. Their preparation and pharmaceuticals containing them. U.S. patent. 1995;5:457–133. [Google Scholar]
- YOUDIM M.B.H., TATTON W.G.Nueroprotective properties of anti-Parkinson drug rasagiline and its optical isomer Neurosci. Lett. 1999. In press