

Abstract
Tight control of cellular growth is essential to ensure normal tissue patterning and prevent pathological responses. Excessive vascular smooth muscle cell (VSMC) proliferation is associated with the pathophysiology of atherosclerosis and restenosis post-angioplasty. Thus, drug targeting of pathological VSMC growth may be a suitable therapeutic intervention in vascular proliferative diseases.
In the present study, we investigated the mechanisms underlying VSMC growth arrest induced by the pharmacological agent PCA-4230. Addition of PCA-4230 to cultured VSMCs blocked the induction of cyclin D1 and cyclin A expression normally seen in serum-restimulated cells. Moreover, PCA-4230 inhibited cyclin-dependent kinase 2 (CDK2) activity and abrogated hyperphosphorylation of the retinoblastoma (Rb) gene product. Similarly, PCA-4230-dependent growth arrest of transformed cell lines correlated with reduced level of cyclin D1 protein and inhibition of CDK2 activity. Consistent with these findings, PCA-4230 repressed serum-inducible cyclin A promoter activity, and overexpression of either cyclin D1 or E2F1 efficiently circumvented this inhibitory effect. Importantly, adenovirus-mediated overexpression of E2F1 restored S-phase entry in PCA-4230-treated VSMCs, demonstrating that PCA-4230 represses cyclin A gene expression and VSMC growth via inhibition of the cyclin D1/E2F pathway.
Because of its ability to inhibit the growth of human VSMCs and transformed cell lines, future studies are warranted to assess whether PCA-4230 may be a suitable therapeutic intervention for the treatment of hyperproliferative disorders, including cardiovascular disease and cancer.
Keywords: PCA-4230; 1,4-dihydropyridine; vascular smooth muscle cells; tumour cells; cell cycle
Introduction
Cell cycle progression is controlled by several cyclin-dependent kinases (CDKs) that associate with regulatory subunits called cyclins (Nurse, 1994; Morgan, 1995). Active CDK/cyclin holoenzymes are presumed to hyperphosphorylate the retinoblastoma (Rb) gene product and the related pocket proteins p107 and p130. The interaction among members of the E2F family of transcription factors and individual pocket proteins is a complex regulatory event that determines whether E2F proteins function as transcriptional activators or repressors (Helin & Harlow, 1993; Weinberg, 1995; Dyson, 1998; Mayol & Graña, 1998; Lavia & Jansen-Durr, 1999). It is accepted that phosphorylation of pocket proteins from mid G1 to mitosis is involved in the transactivation of genes with functional E2F-binding sites. The genes activated by E2F include several growth and cell-cycle regulators (i.e., c-myc, Rb, p34cdc2, cyclin E, cyclin A), as well as genes encoding proteins that are required for nucleotide and DNA biosynthesis (i.e., DNA polymerase α, histone H2A, proliferating cell nuclear antigen, thymidine kinase) (Lavia & Jansen-Durr, 1999).
Excessive proliferation of vascular smooth muscle cells (VSMCs) is considered a key event in the pathophysiology of vascular proliferative diseases (Ross, 1993; Bauters & Isner, 1997; Andrés, 1998). In the adult organism, at homeostasis, VSMCs are found in a ‘contractile' phenotype characterized by the expression of a unique repertoire of differentiation markers and an extremely low proliferation index (Owens, 1995). Numerous observations have suggested that VSMCs in primary atheromas and restenotic lesions have changed to a ‘synthetic' state in which they can respond to different growth factors and cytokines that induce cellular hyperplasia (Campbell & Campbell, 1990; Ross, 1993). Therefore, drug targeting of pathological VSMC growth may be a suitable therapeutic intervention in vascular proliferative diseases.
PCA-4230 is a dihydropyridine derivative showing antithrombotic activity that has long lasting although reversible effects (Gutierrez Diaz et al., 1992; Ortega et al., 1993). A recent study demonstrated the ability of PCA-4230 to reversibly inhibit serum-inducible proliferation of cultured rat VSMCs (del Rio et al., 1997). However, neither the mechanism underlying the antimitogenic effect of PCA-4230 on VSMCs, nor its effect on the proliferation of human VSMCs have been reported. In the present study we demonstrate that PCA-4230 suppresses cyclin A gene transcription and VSMC proliferation via inhibition of the cyclin D1/E2F1 pathway. We also show that PCA-4230 inhibited, in a dose-dependent manner, the growth of human VSMCs and transformed cell lines.
Methods
Cell culture, flow cytometry and 3H-thymidine incorporation
Human VSMCs were isolated from saphenous vein (Pickering et al., 1992). E19P cells (gift from C. Shanahan, Addenbrooke's Hospital, Cambridge, U.K.) were obtained from explant cultures of embryonic day 19 aorta from Fisher rats. These cells express SM22α, calponin and SM-α-actin mRNAs. Human cancer cell lines MCF7 (breast adenocarcinoma) and U2OS (primary bone osteosarcoma) were purchased from American Type Culture Collection (Manassas, VA, U.S.A.). Primary mouse embryonic fibroblasts transformed with the c-Myc oncoprotein were generated as previously described (Perez-Roger et al., 1999). VSMCs were maintained in M199 medium supplemented with 10% FBS. For serum starvation, VSMCs were maintained for 3 days in 0.5% FBS/M199. Transformed cell lines were maintained in Dulbecco's modified Eagle's medium (DMEM) containing 10% FBS. Cultures were maintained at 37°C (95% air and 5% CO2).
PCA-4230 (2(1, 1, 3-trioxo-2, 3-dihydro-1, 2-benzisothiazol-2-yl)ethyl 2, 6-dimethyl-5-(ethoxycarbonyl)-4-methyl-1, 4-dihydropyridincarboxylate) (gift from M.P. Ortega, Laboratorios Alter, Madrid, Spain) was dissolved in dimethyl sulphoxide (DMSO) and immediately used at a dilution giving a final concentration of 0.1% DMSO. Control cells were treated with 0.1% DMSO. Media were changed daily throughout the experiments. For flow cytometric analysis, PCA-4230 or vehicle was administered to starvation-synchronized cells 1 – 2 h before serum restimulation and throughout the 16 h of serum treatment. Flow cytometric analysis was performed as previously described (Sylvester et al., 1998).
For experiments with adenovirus encoding for E2F1 or β-galactosidase (provided by J.R. Nevins, Duke University Medical Center, NC, U.S.A.), cells were seeded in 6-well dishes (25×103 cells/well) and were maintained in 10% v v−1 FBS. Two days later, cells were starvation-synchronized and harvested for FACS analysis, or were pretreated with DMSO or 50 μM PCA-4230 followed by 16 h of serum restimulation. When indicated, starved cells were infected with adenovirus at different multiplicities of infection (MOI) during the last 8 h prior to serum restimulation.
To assess the effect of PCA-4230 on DNA synthesis in MEF-Myc cultures, cells were seeded in 12-well dishes at a density of 60×103 cells/well in 10% FBS/DMEM supplemented with vehicle or PCA-4230. The following day, cells were treated for 2 h with 1 μCi ml−1 of 3H-thymidine (Amersham Pharmacia, Little Chalfont, U.K.). The amount of 3H-thymidine incorporated into DNA was determined by precipitation with trichloroacetic acid and scintillation counting.
Western blot analysis, immunoprecipitation and immune complex kinase assays
Cells were lysed in ice-cold lysis buffer (20 mM HEPES buffer [pH 7.5], 10 mM EGTA, 40 mM β-glycerophosphate, 1% w v−1 NP-40, 2.5 mM MgCl2, 2 mM orthovanadate, 1 mM DTT, 1 mM phenylmethyl sulfonyl fluoride, 10 μg ml−1 aprotinin and 10 μg ml−1 leupeptin). Following centrifugation at 15,000×g for 20 min at 4°C, supernatants were recovered. Western blot and CDK2 assays were performed as previously described (Chen et al., 1997) using the following dilutions of primary antibodies: anti-CDK2 (sc-163, 1/250), anti-cyclin A (sc-751, 1/100), anti-cyclin D1 (sc-450, 1/100), anti-cyclin E (sc-481, 1/250), anti-p27 (sc-528, 1/1000), and anti-p21 (sc-397, 1/250) (Santa Cruz Biotechnology, Santa Cruz, CA, U.S.A.). Rb was detected using a 1/1000 dilution of monoclonal antibody 3C8 (Wen et al., 1994).
For phosphatidylinositol 3-kinase (PI3K) assay, lysates containing 1 mg of total protein were immunoprecipitated with 0.5 μg of anti-PI3K antibody (anti-p85α, sc-423, Santa Cruz Biotechnology). The reaction was started by mixing the immunopellet with 25 μl of a cocktail containing 20 μg L-α-phosphatidylinositol (Avanti Polar Lipids, Alabaster, AL, U.S.A.), 100 mM [γ-32P]ATP (10 μCi) and 40 μM ATP in 25 mM HEPES [pH 7.4]; 10 mM MgCl2 and 0.5 mM EGTA. After 20 min the reaction was stopped by addition of 400 μl of chloroform-methanol (1 : 2 in 1% HCl), plus 125 μl of chloroform and 125 μl of 10 mM HCl. Samples were centrifuged briefly and the lower organic phase was removed and washed once with 500 μl of methanol: 100 mM HCl plus 2 mM EDTA (1 : 1). The organic lower phase was extracted, dried under vacuum and resuspended in 30 μl of chloroform. Samples were applied to silica gel thin-layer chromatography plates (Merck, Darmstadt, Germany). The chromatography was developed with 1-propanol: 2N acetic acid (65 : 35 v v−1), dried and visualized by autoradiography. The amount of radioactivity incorporated into L-α-phosphatidylinositol phosphate was quantified in a Phosphoimager.
Transient transfections assays
E19P cells seeded into 6-well dishes were transiently transfected with Superfect as recommended by the manufacturer (Quiagen, Santa Clarita, CA, U.S.A.) (1 : 2 DNA:Superfect ratio). The luciferase reporter plasmid driven by the human cyclin A promoter region from −924 to +245 (gift of Sobczak-Thépot, INSERM, France) has been described previously (Henglein et al., 1994). Final DNA concentration in experiments involving cotransfected expression vectors for cyclin D1 and E2F1 driven by the cytomegalovirus promoter (gift of K. Walsh, St. Elizabeth's Medical Center of Boston, MA, U.S.A.) was equalized by adding empty vector. To correct for differences in transfection efficiency, luciferase activity was normalized relative to the level of alkaline phosphatase activity produced from cotransfected pSVAPAP plasmid (Henthorn et al., 1988). Luciferase and alkaline phosphatase activities were measured as previously described (Andrés et al., 1995).
Results
PCA-4230 inhibits the proliferation of cultured VSMCs and transformed cell lines
We first examined the effect of PCA-4230 on the kinetics of proliferation of cultured VSMCs. E19P cells, an established cell line of rat embryonic aorta VSMCs, and primary cultures of human VSMCs were maintained in high-mitogen media. Addition of PCA-4230 reduced cell number in a dose-dependent manner over a period of 4 – 6 days in culture (Figure 1A). For example, when compared to control cultures at the latest time points investigated, 5 μM PCA-4230 decreased the number of E19P cells and human VSMCs by 25 and 21%, respectively, whereas 50 μM PCA-4230 reduced E19P and human VSMC number by 85 and 74%, respectively.
Figure 1.

PCA-4230 inhibits serum-inducible proliferation of rat and human VSMCs. Results represent the mean±s.e.mean of three independent measurements. (A) The rat E19P cell line and primary human VSMCs were plated into 6-well dishes and maintained with medium containing 10% FBS plus vehicle or PCA-4230. Cells were trypsinized at different times and cell number was determined with a haemocytometer. (B, C) Cells were starved for 3 days in medium containing 0.5% FBS. After pretreatment with vehicle or PCA-4230, cells were stimulated with 10% FBS plus vehicle or PCA-4230. Cells were harvested for FACS analysis after 16 h of serum stimulation.
We next performed flow cytometry to analyse the effect of PCA-4230 on cell cycle profiles. After pretreatment with vehicle or PCA-4230, starvation-synchronized cells were restimulated with medium containing 10% FBS and vehicle or PCA-4230. Analysis after 16 h of serum restimulation disclosed a dose-dependent inhibition of S-phase in E19P cells treated with PCA-4230 (Figure 1B). Likewise, under conditions where serum-restimulation markedly increased S-phase entry of human VSMCs, 50 μM PCA-4230 completely blocked this response (Figure 1C). Thus, PCA-4230-dependent inhibition of rat and human VSMC growth is associated with G0/G1 arrest.
The effect of PCA-4230 on the growth of transformed cell lines was also investigated. Addition of PCA-4230 to the culture media reduced in a dose-dependent manner the number of MCF7 cells (human breast adenocarcinoma), U2OS cells (human osteosarcoma), and mouse embryonic fibroblast transformed with the proto-oncogene c-Myc (MEF-Myc) (Figure 2A,B). The inhibitory effect of PCA-4230 on the growth of MEF-Myc cells correlated with a marked reduction in 3H-thymidine incorporation (Figure 2C). In agreement with previous studies (del Rio et al., 1997), PCA-4230-treated cultures resumed growth if drug treatment was discontinued (data not shown).
Figure 2.
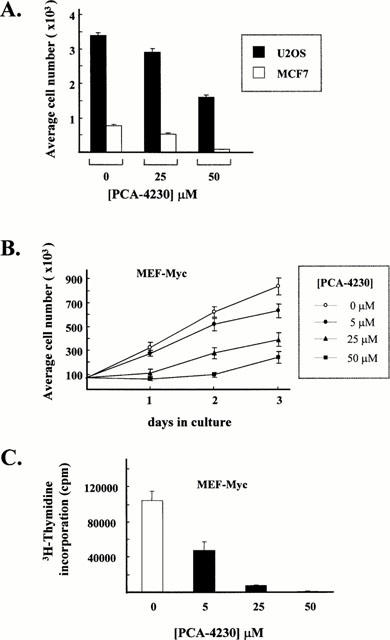
PCA-4230 inhibits proliferation of transformed cells. Cells were maintained in medium containing 10% FBS plus vehicle or PCA-4230. Results represent the mean±s.e.mean of three independent measurements. (A) U2OS and MCF7 cells were plated in 6-well dishes. After 3 days, cells were trypsinized and cell number was determined with a haemocytometer. (B) MEF-Myc cells growing on 6-well dishes were trypsinized at different times and cell number was determined with a haemocytometer. (C) Effect of PCA-4230 on the incorporation of 3H-thymidine into MEF-Myc cells (see Methods for details).
Effects of PCA-4230 on the expression and activity of key cell cycle regulatory proteins
To gain insight into the mechanisms underlying PCA-4230-dependent inhibition of cell proliferation, we investigated the effect of this drug on the activity of CDK2, a key positive regulator of the G1/S transition. Concentrations of PCA-4230 that caused growth arrest markedly reduced CDK2 activity in serum-stimulated VSMCs and MEF-Myc cells (Figure 3A). In contrast, PI3K activity in VSMCs exposed to 20 and 50 μM was 89 and 96% of that seen in control cells, respectively (Figure 3B). Consistent with these findings, PCA-4230 did not affect the phosphorylation status of the PI3K substrate Akt (data not shown). Collectively, these results indicate that PCA-4230 may act specifically to block certain signalling pathways involved in mitogen-induced cell growth.
Figure 3.
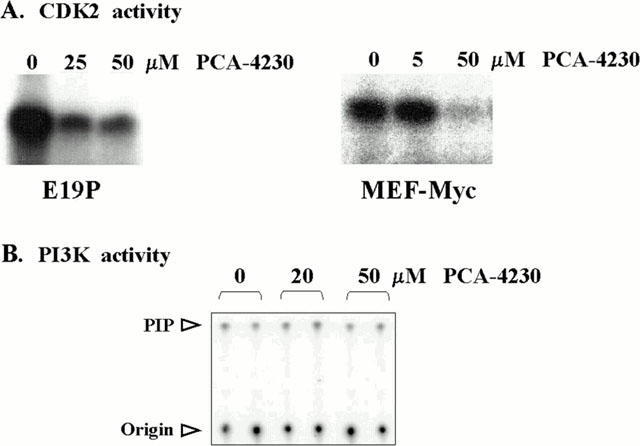
PCA-4230 inhibits CDK2 activity. Subconfluent E19P and MEF-Myc cells maintained in 10% FBS were treated for 15 h with vehicle or the indicated amounts of PCA-4230. (A) Cell lysates were assayed for CDK2 activity using histone H1 as substrate. (B) PI3K activity in E19P lysates using L-α-phosphatidylinositol as substrate. PIP: L-α-phosphatidylinositol phosphate.
We next sought to examine by Western blot analysis the effect of PCA-4230 on key components of the cell cycle machinery (Figure 4). As expected, serum restimulation of starvation-synchronized VSMCs led to a transient induction of cyclin D1 protein expression that showed maximum levels at 8 h and was followed by the upregulation of cyclin A at 16 h (Figure 4A, lanes 1, 3, 5 and 7). Addition of PCA-4230 markedly inhibited serum-inducible cyclin D1 and cyclin A expression (Figure 4A, lanes 2, 4, 6 and 8). Neither serum restimulation nor PCA-4230 treatment affected significantly the expression of cyclin E and CDK2 (Figure 4A). Thus, the remaining CDK2 activity in PCA-4230-treated cells might result from cyclin E-containing CDK2 holoenzymes. PCA-4230-dependent inhibition of the level of cyclin D1 protein was also seen in MEF-Myc cells (Figure 4B).
Figure 4.

Western blot analysis of cell cycle regulatory proteins in PCA-4230-treated cells. Control cultures were exposed to vehicle. (A, C) E19P cells were starvation-synchronized for 3 days in medium containing 0.5% FBS. One-and-a-half hours before serum-restimulation, 50 μM PCA-4230 was added and treatment continued throughout the period of serum restimulation. The antibody used in each blot is shown. pRb: hypophosphorylated Rb; ppRb: hyperphosphorylated Rb. (B) Cyclin D1 expression in asynchronously growing MEF-Myc cells. (D) p27 expression in asynchronously growing MCF7, U2OS and E19P cells.
Progression through the mammalian cell cycle requires the hyperphosphorylation of Rb by active CDK/cyclin holoenzymes (Helin & Harlow, 1993; Weinberg, 1995; Dyson, 1998; Mayol & Graña, 1998). Consistent with its inhibitory effect on CDK2 activity and the reduction in cyclin D1 protein, PCA-4230 abrogated the hyperphosphorylation of Rb normally seen in serum-restimulated VSMCs (Figure 4, compare lanes 1, 3, 5, 7 and 2, 4, 6, 8). Therefore, growth arrest of PCA-4230-treated cells is associated with inhibition of the kinase activity of CDK2 and repression of Rb hyperphosphorylation. Since CDK2 activity and Rb hyperhosphorylation are negatively regulated by specific CDK inhibitors (CKIs) (Elledge & Harper, 1994; Peter & Herskowitz, 1994; Graña & Reddy, 1995; Morgan, 1995), we considered the possibility that members of this family of growth suppressors might underlie the growth inhibitory effect of PCA-4230. However, growth arrest in cells exposed to PCA-4230 did not correlate with increased expression of the CKIs p21 and p27 (Figure 4C,D).
PCA-4230 inhibits serum-inducible cyclin A promoter activity
Hyperphosphorylation of Rb is necessary to induce transcriptional activation of E2F-dependent target genes that are required for DNA synthesis (Helin & Harlow, 1993; Weinberg, 1995; Dyson, 1998; Mayol & Graña, 1998). Since PCA-4230 inhibited Rb hyperphosphorylation (Figure 4), a series of experiments were performed to examine the effect of PCA-4230 on transcriptional activation of the cyclin A gene, a known E2F-regulated gene which is induced by serum in VSMCs (Sylvester et al., 1998). To this end, E19P cells were transiently transfected with a luciferase reporter gene driven by the cyclin A gene promoter. These experiments demonstrated the ability of PCA-4230 to abrogate, in a dose-dependent manner, serum-inducible cyclin A promoter activity in starvation-synchronized cells (Figure 5A). Likewise, addition of PCA-4230 to asynchronously growing E19P cells inhibited in a dose-dependent manner cyclin A promoter activity (Figure 5B). These results suggest that PCA-4230-dependent inhibition of cyclin A gene expression is achieved, at least in part, at the transcriptional level.
Figure 5.

PCA-4230 inhibits serum-inducible cyclin A promoter activity in VSMCs. E19P cells were cotransfected with 2 μg of a luciferase reporter gene driven by the human cyclin A promoter and 0.5 μg of a control plasmid encoding for alkaline phosphatase. Results are expressed as the ratio luciferase/alkaline phosphatase. Bars represent the mean±s.e.mean of three independent transfections. Control cells were treated with vehicle. Results are referred to the activity seen in control untreated cells (=100%). (A) Transfected cells were maintained in 0.5% FBS for 2 days. Cells were then harvested to determine basal cyclin A promoter activity in serum-starved cells (first bar), or were pretreated for 2 h with vehicle or PCA-4230 and then serum-restimulated overnight. (B) Cells were maintained in 10% FBS throughout the experiment.
Ectopic overexpression of cyclin D1 and E2F1 overrides the inhibitory effect of PCA-4230 on cyclin A promoter activity and S-phase entry
The results of our Western blot analysis and transient transfection assays suggested that PCA-4230-dependent transcriptional repression of cyclin A gene expression and growth arrest is due, at least in part, to inhibition of the cyclin D1/E2F pathway. Further evidence in support of this model was provided by rescue experiments in which the luciferase reporter gene driven by the cyclin A gene promoter was cotransfected with either cyclin D1 or E2F1 expression vectors. In agreement with previous studies demostrating the ability of cyclin D1 and E2F1 to induce transcription from the cyclin A gene promoter in fibroblasts (Schulze et al., 1995; Rudolph et al., 1996; Zerfass-Thome et al., 1997; Lavia & Jansen-Durr, 1999), overexpression of cyclin D1 and E2F1 increased cyclin A promoter activity by 4 fold in serum-stimulated VSMCs (Figure 6A,B, respectively; compare gray bars). While PCA-4230 repressed cyclin A promoter activity in control cells, ectopic overexpression of either cyclin D1 (Figure 6A) or E2F1 (Figure 6B) efficiently overcame the inhibitory effect of PCA-4230.
Figure 6.

Ectopic overexpression of either cyclin D1 or E2F1 can overcome the inhibitory effect of PCA-4230 on cyclin A promoter activity. Cells were treated as in Figure 5, except that cultures were maintained throughout the experiment in 10% FBS with vehicle or with 50 μM PCA-4230. Results are expressed as the ratio luciferase/alkaline phosphatase. Bars represent the mean±s.e.mean of three independent transfections. Results are referred to the activity seen in control untreated cells (=100%). Cells were cotransfected with CMV-Cyclin D1 (0.1 μg per transfection, A), or with CMV-E2F1 (0.2 μg per transfection, B).
We next wanted to ascertain whether ectopic overexpression of E2F1 could overcome PCA-4230-dependent growth arrest. For these studies, we used a replication-defective adenovirus encoding for E2F1 (Ad-E2F1). Consistent with the results of Figure 1B, 50 μM PCA-4230 blocked the induction of S-phase normally seen in serum-restimulated E19P cells, and infection with Ad-E2F1, but not control adenovirus encoding for β-galactosidase (Ad-βgal), overcame in a dose-dependent manner PCA-4230-induced growth arrest (Figure 7). Taken together, these results suggest that PCA-4230 represses cyclin A promoter activity and cell proliferation through inhibition of the cyclin D1/E2F1 pathway.
Figure 7.
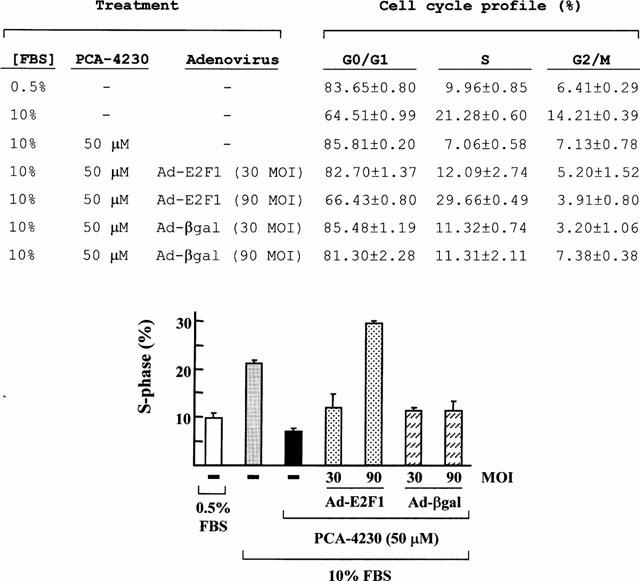
Adenovirus-mediated overexpression of E2F1 restores S-phase entry in PCA-4230-treated VSMCs. E19P cells were maintained for 3 days in 0.5% FBS. Cells were then harvested for FACS analysis to determine basal activity (first bar), or pretreated with vehicle or 50 μM PCA-4230 followed by 16 h of stimulation with 10% FBS. When indicated, starved cells were infected with replication-defective Ad-E2F1 or Ad-βgal at different multiplicities of infection (MOI) during the last 8 h of serum starvation. Results represent the mean±s.e.mean of three experiments.
Discussion
In this study we have examined the molecular mechanisms underlying cellular growth arrest by the pharmacological agent PCA-4230. Our results show that PCA-4230 is a potent inhibitor of serum-inducible S-phase entry in cultures of both rat and human VSMCs. This agent also inhibited the growth of human and murine transformed cells. Potential mechanisms underlying PCA-4230-dependent growth arrest include: (i) inhibition of cyclin D1 and cyclin A protein expression; (ii) abrogation of CDK2 activity and blockade of Rb hyperphosphorylation; and (iii) transcriptional repression of cyclin A gene expression. Neither PI3K activity nor cylin E and CDK2 protein levels were affected by PCA-4230. Moreover, PCA-4230 failed to inhibit CDK2 activity when added directly to kinase reactions (data not shown), demonstrating that the inhibitory effect of this agent was not due to a direct interaction with CDK2. These results indicate that PCA-4230 blocks specifically certain signalling pathways involved in cellular proliferation.
We considered the possibility that members of the CKI family of growth suppressors might contribute to PCA-4230-dependent growth arrest. However, increased expression of the CKIs p21 and p27 was not seen in PCA-4230-treated cells. In contrast, tranilast, a compound with proven clinical efficacy for treating postangioplasty restenosis (Holmes et al., 2000; Tamai et al., 1999), inhibited CDK2/CDK4 activity and Rb hyperphosphorylation by a mechanism that may depend upon increased expression of p21 (Takahashi et al., 1999).
The complex interactions between Rb and members of the E2F family of transcription factors play an important role in the regulation of cell cycle progression (Graña & Reddy, 1995; Dyson, 1998). It is accepted that hyperphosphorylation of Rb by active G1 CDK/cyclin holoenzymes disrupts Rb/E2F interactions thus allowing transcriptional activation of E2F-dependent target genes (i.e., cyclin A, p34cdc2, cyclin E). Thus, inhibition of both cyclin D1 expression and CDK2 activity may account for the blockade of serum-inducible Rb hyperphosphorylation in PCA-4230-treated VSMCs. Accumulation of hypophosphorylated Rb would in turn hinder the accumulation of ‘free E2F', thus preventing cyclin A gene expression and formation of active CDK2/cyclin A complexes. In this regard, we have recently shown that E2F is an important component of the signalling cascade that links Ras activity to cyclin A transcription in VSMCs (Sylvester et al., 1998). Because disruption of cyclin A function inhibits S-phase entry (Girard et al., 1991; Pagano et al., 1992; Zindy et al., 1992), and its overexpression accelerates the G1-to-S transition (Resnitzky et al., 1995; Rosenberg et al., 1995), cyclin A expression appears to be rate limiting for cellular proliferation. Therefore, repression of cyclin A gene transcription may contribute to PCA-4230-dependent inhibition of VSMC proliferation.
To ascertain that PCA-4230 does indeed inhibit cyclin A gene expression and VSMC proliferation through inhibition of the cyclin D1/E2F pathway, we performed rescue experiments by overexpressing cyclins D1 and E2F1. Our results demonstrate that ectopic overexpression of either cyclin D1 or E2F1 efficiently overcame the inhibitory effect of PCA-4230 on cyclin A promoter activity. Moreover, adenovirus-mediated overexpression of E2F1 restored DNA synthesis in PCA-4230-treated VSMCs. These results suggest that PCA-4230 represses cyclin A promoter activity and VSMC proliferation through inhibition of the cyclin D1/E2F1 pathway.
Abnormal VSMC proliferation is an important component of the response to vascular injury, particularly during in-stent restenosis (Ross, 1993; Bauters & Isner, 1997; Libby & Tanaka, 1997). We have previously shown that VSMC proliferation in response to arterial injury in rat and human arteries correlated with the induction of CDK2 and cyclins (Kearney et al., 1997; Wei et al., 1997). Importantly, gene therapy strategies targeting CDK2 and E2F function and Rb hyperphosphorylation have proven efficient at preventing vascular proliferative diseases in animal models (reviewed in Andrés, 1998; Braun-Dullaeus et al., 1998) and human bypass-graft atherosclerosis (Mann et al., 1999). Previous studies have demonstrated that PCA-4230 has platelet aggregation inhibitory activity and in vivo antithrombotic activity (Gutierrez Diaz et al., 1992; Ortega et al., 1993; Sunkel et al., 1988). Our in vitro studies show that inhibition of the cyclin D1/E2F pathway by PCA-4230 is effective at suppressing the growth of human VSMCs and cancer cells. Because PCA-4230 was well tolerated when administered to healthy volunteers (Cillero et al., 1991), future studies are warranted to assess whether PCA-4230 may be a suitable pharmacological agent for the treatment of hyperproliferative disorders, including cardiovascular disease and cancer.
Acknowledgments
We thank M.P. Ortega/Laboratorios Alter (PCA-4230), C. Shanahan (E19P cells), J. Sobczak-Thépot (cyclin A-luciferase reporter gene), J.R. Nevins (Ad-E2F1, Ad-βgal) and K. Walsh (CMV-cyclin D1, CMV-E2F1) for the gift of reagents. We are also grateful to Dr Barbara A. Gilchrest for critical reading of the manuscript, and María J. Andrés for the preparation of figures. This work was supported by Spanish Dirección General de Educación Superior e Investigación Científica grant PM97-0136 and by National Institutes of Health grant HL RO1-57519 (to V. Andrés).
Abbreviations
- Ad-E2F1 (Ad-βgal)
replication-defective adenovirus encoding for E2F1 (β-galactosidase)
- CDK(s)
cyclin-dependent kinase(s)
- DMSO
dimethyl sulphoxide
- MEF
mouse embryonic fibroblast
- MEF-Myc
mouse embryonic fibroblast transformed with the proto-oncogen Myc
- Rb
retinoblastoma susceptibility gene product
- TCA
trichloroacetic acid
- VSMC(s)
vascular smooth muscle cell(s)
References
- ANDRÉS V. Control of vascular smooth muscle cell growth and its implication in atherosclerosis and restenosis. Int. J. Molec. Med. 1998;2:81–89. doi: 10.3892/ijmm.2.1.81. [DOI] [PubMed] [Google Scholar]
- ANDRÉS V., FISHER S., WEARSCH P., WALSH K. Regulation of Gax homeobox gene transcription by a combination of positive factors including MEF2. Mol. Cell. Biol. 1995;15:4272–4281. doi: 10.1128/mcb.15.8.4272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BAUTERS C., ISNER J.M. The biology of restenosis. Prog. Cardiovasc. Dis. 1997;40:107–116. doi: 10.1016/s0033-0620(97)80003-5. [DOI] [PubMed] [Google Scholar]
- BRAUN-DULLAEUS R.C., MANN M.J., DZAU V.J. Cell cycle progression: new therapeutic target for vascular proliferative disease. Circulation. 1998;98:82–89. doi: 10.1161/01.cir.98.1.82. [DOI] [PubMed] [Google Scholar]
- CAMPBELL G.R., CAMPBELL J.H. The phenotypes of smooth muscle expressed in human atheroma. Ann. N.Y. Acad. Sci. 1990;598:143–158. doi: 10.1111/j.1749-6632.1990.tb42286.x. [DOI] [PubMed] [Google Scholar]
- CHEN D., KRASINSKI K., CHEN D., SYLVESTER A., CHEN J., NISEN P.D., ANDRÉS V. Downregulation of cyclin-dependent kinase 2 activity and cyclin A promoter activity in vascular smooth muscle cells by p27Kip1, an inhibitor of neointima formation in the rat carotid artery. J. Clin. Invest. 1997;99:2334–2341. doi: 10.1172/JCI119414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CILLERO F.J., NAVARRO J.L., NARVAIZA J.T., PRIEGO J.G., ORTEGA M.P. Clinical Pharmacology (Phase I) of PCA-4230, a new antithrombotic agent, in healthy volunteers. Thromb. Haemost. 1991;65:1145. [Google Scholar]
- DEL RIO M., SUNKEL C., LARCHER F., ORTEGA M.P. Antiproliferative effects of PCA-4230, a new antithrombotic drug, in vascular smooth muscle cells. Br. J. Pharmacol. 1997;120:1360–1366. doi: 10.1038/sj.bjp.0701035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DYSON N. The regulation of E2F by pRB-family proteins. Genes Dev. 1998;12:2245–2262. doi: 10.1101/gad.12.15.2245. [DOI] [PubMed] [Google Scholar]
- ELLEDGE S.J., HARPER J.W. Cdk inhibitors: on the threshold of checkpoints and development. Curr. Opin. Cell Biol. 1994;6:847–852. doi: 10.1016/0955-0674(94)90055-8. [DOI] [PubMed] [Google Scholar]
- GIRARD F., STRAUSFELD U., FERNÁNDEZ A., LAMB N. Cyclin A is required for the onset of DNA replication in mammalian fibroblasts. Cell. 1991;67:1169–1179. doi: 10.1016/0092-8674(91)90293-8. [DOI] [PubMed] [Google Scholar]
- GRAÑA X., REDDY E.P. Cell cycle control in mammalian cells: role of cyclins, cyclin dependent kinases (CDKs), growth suppressor genes and cyclin-dependent kinase inhibitors (CKIs) Oncogene. 1995;11:211–219. [PubMed] [Google Scholar]
- GUTIERREZ, DIAZ J.A., TOLON R.M., SUNKEL C., PRIEGO J.G., ORTEGA M.P. Interaction of platelets with subendothelium in rats treated with PCA-4230, a new antithrombotic agent. Haemostasis. 1992;22:202–210. doi: 10.1159/000216320. [DOI] [PubMed] [Google Scholar]
- HELIN K., HARLOW E. The retinoblastoma protein as a transcriptional repressor. Trends Cell Biol. 1993;3:43–46. doi: 10.1016/0962-8924(93)90150-y. [DOI] [PubMed] [Google Scholar]
- HENGLEIN B., CHENIVESSE X., WANG J., EICK D., BRÉCHOT C. Structure and cell cycle-regulated transcription of the human cyclin A gene. Proc. Natl. Acad. Sci. U.S.A. 1994;91:5490–5494. doi: 10.1073/pnas.91.12.5490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HENTHORN P., ZERVOS P., RADUCHA M., HARRIS H., KADESCH T. Expression of a human placental alkaline phosphatase gene in transfected cells: use as a reporter for studies of gene expression. Proc. Natl. Acad. Sci. U.S.A. 1988;85:6342–6346. doi: 10.1073/pnas.85.17.6342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HOLMES D., FITZGERALD P., GOLDBERG S., LABLANCHE J., LINCOFF A.M., SAVAGE M., SERRUYS P.W., WILLERSON J., GRANETT J.R., CHAN R., SHUSTERMAN N.H., POLAND M. The PRESTO (Prevention of restenosis with tranilast and its outcomes) protocol: a double-blind, placebo-controlled trial. Am. Heart J. 2000;139:23–31. doi: 10.1016/s0002-8703(00)90304-1. [DOI] [PubMed] [Google Scholar]
- KEARNEY M., PIECZEK A., HALEY L., LOSORDO D.W., ANDRÉS V., SCHAINFIELD R., ROSENFIELD R., ISNER J.M. Histopathology of in-stent restenosis in patients with peripheral artery disease. Circulation. 1997;95:1998–2002. doi: 10.1161/01.cir.95.8.1998. [DOI] [PubMed] [Google Scholar]
- LAVIA P., JANSEN-DURR P. E2F target genes and cell-cycle checkpoint control. Bioessays. 1999;21:221–230. doi: 10.1002/(SICI)1521-1878(199903)21:3<221::AID-BIES6>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- LIBBY P., TANAKA H. The molecular basis of restenosis. Prog. Cardiovasc. Dis. 1997;40:97–106. doi: 10.1016/s0033-0620(97)80002-3. [DOI] [PubMed] [Google Scholar]
- MANN M.J., WHITTEMORE A.D., DONALDSON M.C., BELKIN M., CONTE M.S., POLAK J.F., ORAV E.J., EHSAN A., DELL'ACQUA G., DZAU V.J. Ex-vivo gene therapy of human vascular bypass grafts with E2F decoy: the PREVENT single-centre, randomised, controlled trial. Lancet. 1999;354:1493–1498. doi: 10.1016/S0140-6736(99)09405-2. [DOI] [PubMed] [Google Scholar]
- MAYOL X., GRAÑA X. The p130 pocket protein: keeping order at cell cycle exit/re-entrance transitions. Front. Biosci. 1998;3:11–24. doi: 10.2741/a263. [DOI] [PubMed] [Google Scholar]
- MORGAN D.O. Principles of CDK regulation. Nature. 1995;374:131–134. doi: 10.1038/374131a0. [DOI] [PubMed] [Google Scholar]
- NURSE P. Ordering S phase and M phase in the cell cycle. Cell. 1994;79:547–550. doi: 10.1016/0092-8674(94)90539-8. [DOI] [PubMed] [Google Scholar]
- ORTEGA M.P., SUNKEL C., GÓMEZ M.M., SANTOS L., FAU DE CASA-JUANA M., PRIETO J. A new generation of 1,4-dihydropyridines showing novel pharmacological properties. Curr. Topics Med. Chem. 1993;1:83–91. [Google Scholar]
- OWENS G.K. Regulation of differentiation of vascular smooth muscle cells. Physiol. Rev. 1995;75:487–517. doi: 10.1152/physrev.1995.75.3.487. [DOI] [PubMed] [Google Scholar]
- PAGANO M., PEPPERKOK R., VERDE F., ANSORGE W., DRAETTA G. Cyclin A is required at two points in the human cell cycle. EMBO J. 1992;11:961–971. doi: 10.1002/j.1460-2075.1992.tb05135.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PEREZ-ROGER I., KIM S.H., GRIFFITHS B., SEWING A., LAND H. Cyclins D1 and D2 mediate myc-induced proliferation via sequestration of p27(Kip1) and p21(Cip1) EMBO J. 1999;18:5310–5320. doi: 10.1093/emboj/18.19.5310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PETER M., HERSKOWITZ I. Joining the complex: cyclin-dependent kinase inhibitory proteins and the cell cycle. Cell. 1994;79:181–184. doi: 10.1016/0092-8674(94)90186-4. [DOI] [PubMed] [Google Scholar]
- PICKERING J.G., WEIR L., ROSENFIELD K., STETZ J., JEKANOWSKI J., ISNER J.M. Smooth muscle cell outgrowth from human atherosclerotic plaque: implications for the assessment of lesion biology. J. Am. Coll. Cardiol. 1992;20:1430–1439. doi: 10.1016/0735-1097(92)90259-p. [DOI] [PubMed] [Google Scholar]
- RESNITZKY D., HENGST L., REED S.I. Cyclin A-associated kinase activity is rate limiting for entrance into S phase and is negatively regulated in G1 by p27Kip1. Mol. Cell. Biol. 1995;15:4347–4352. doi: 10.1128/mcb.15.8.4347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ROSENBERG A.R., ZINDY F., LE DEIST F., MOULY H., METEZEAU P., BRÉCHOT C., LAMAS E. Overexpression of human cyclin A advances entry into S phase. Oncogene. 1995;10:1501–1509. [PubMed] [Google Scholar]
- ROSS R. The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature. 1993;362:801–809. doi: 10.1038/362801a0. [DOI] [PubMed] [Google Scholar]
- RUDOLPH B., SAFFRICH R., ZWICKER J., HENGLEIN B., MÜLLER R., ANSORGE W., EILERS M. Activation of cyclin-dependent kinases by Myc mediates induction of cyclin A, but not apoptosis. EMBO J. 1996;15:3065–3076. [PMC free article] [PubMed] [Google Scholar]
- SCHULZE A., ZERFASS K., SPITKOVSKY D., MIDDENDORP S., BERGES J., HELIN K., JANSEN-DÜRR P., HENGLEIN B. Cell cycle regulation of the cyclin A gene promoter is mediated by a variant E2F site. Proc. Natl. Acad. Sci. U.S.A. 1995;92:11264–11268. doi: 10.1073/pnas.92.24.11264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SUNKEL C.E., FAU DE CASA-JUANA M., CILLERO F.J., PRIEGO J.G., ORTEGA M.P. Synthesis, platelet aggregation inhibitory activity, and in vivo antithrombotic activity of new 1,4-dihydropyridines. J. Med. Chem. 1988;31:1886–1890. doi: 10.1021/jm00118a004. [DOI] [PubMed] [Google Scholar]
- SYLVESTER A.M., CHEN D., KRASINSKI K., ANDRÉS V. Role of c-fos and E2F in the induction of cyclin A transcription and vascular smooth muscle cell proliferation. J. Clin. Invest. 1998;101:940–948. doi: 10.1172/JCI1630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- TAKAHASHI A., TANIGUCHI T., ISHIKAWA Y., YOKOYAMA M. Tranilast inhibits vascular smooth muscle cell growth and intimal hyperplasia by induction of p21(waf1/cip1/sdi1) and p53. Circ. Res. 1999;84:543–550. doi: 10.1161/01.res.84.5.543. [DOI] [PubMed] [Google Scholar]
- TAMAI H., KATOH O., SUZUKI S., FUJII K., AIZAWA T., TAKASE S., KUROGANE H., NISHIKAWA H., SONE T., SAKAI K., SUZUKI T. Impact of tranilast on restenosis after coronary angioplasty: tranilast restenosis following angioplasty trial (TREAT) Am. Heart J. 1999;138:968–975. doi: 10.1016/s0002-8703(99)70025-6. [DOI] [PubMed] [Google Scholar]
- WEI G.L., KRASINSKI K., KEARNEY M., ISNER J.M., WALSH K., ANDRÉS V. Temporally and spatially coordinated expression of cell cycle regulatory factors after angioplasty. Circ. Res. 1997;80:418–426. [PubMed] [Google Scholar]
- WEINBERG R.A. The retinoblastoma protein and cell cycle control. Cell. 1995;81:323–330. doi: 10.1016/0092-8674(95)90385-2. [DOI] [PubMed] [Google Scholar]
- WEN S.F., NODELMAN M., NARED HOOD K., DUNCAN J., GERADTS J., SHEPARD H.M. Retinoblastoma protein monoclonal antibodies with novel characteristics. J. Immunol. Methods. 1994;169:231–240. doi: 10.1016/0022-1759(94)90267-4. [DOI] [PubMed] [Google Scholar]
- ZERFASS-THOME K., SCHULZE A., ZWERSCHKE W., VOGT B., HELIN K., BARTEK J., HENGLEIN B., JANSEN-DÜRR P. p27KIP1 blocks cyclin E-dependent transactivation of cyclin A gene expression. Mol. Cell. Biol. 1997;17:407–415. doi: 10.1128/mcb.17.1.407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZINDY F., LAMAS E., CHENIVESSE X., SOBCZAK-THÉPOT J., WANG J., FESQUET D., HENGLEIN B., BRÉCHOT C. Cyclin A is required in S phase in normal epithelial cells. Biochem. Biophys. Res. Commun. 1992;182:1144–1154. doi: 10.1016/0006-291x(92)91851-g. [DOI] [PubMed] [Google Scholar]