

Abstract
Many researchers investigating plant-insect interactions maintain their insect colonies on artificial diet and assume that salivary enzymes and elicitors remain representative of natural situations. These salivary elicitors, such as the enzyme glucose oxidase (GOX), play important roles in influencing plant defense responses. In fact, GOX has been implicated in suppressing induced nicotine-production in tobacco plants (Musser et al., 2002). In this study, we investigated the effect of artificial or plant diet on the GOX activity in caterpillars of the beet armyworm, Spodoptera exigua. In the later developmental stages, whole body GOX of S. exigua caterpillars reared on a wheat germ-based artificial diet is over ten times higher than when insects were fed plants of the legume, Medicago trunctula. Labial salivary GOX accounted for most of this whole body activity in 4th instar caterpillars (57.8%), with the remaining activity present in the carcass. Hemolymph GOX levels were below the detection limits of the o-dianisidine-peroxidase assay used to measure activity. Labial salivary GOX activity was significantly higher in 4th larval instars reared on artificial diet compared with plant-fed caterpillars (U/mg per pair labial salivary gland, p = 0.0062), suggesting that diet effects GOX activity. When 4th instar plant-fed caterpillars were transferred to artificial diet, increased labial salivary GOX activity is closely correlated with the amount of time spent feeding on artificial diet. This study shows that the labial salivary GOX activity of S. exigua caterpillars is dependent on diet and developmental stage and that caution must be exercised in the design of plant-insect experiments.
Abbreviations:
GOX: glucose oxidase
Keywords: caterpillar, elicitor, Medicago truncatula, plant-insect interactions, salivary enzyme
Introduction
Salivary glucose oxidase (GOX) has been implicated as a key mechanism used by caterpillars to counteract induced host plant defenses by interfering with jasmonate-dependent signalling (Musser et al., 2002). In phytophagous chewing insects, such as caterpillars, digestion is initiated through the maceration of plant tissues by the mandibles assisted by the laciniae of the maxillae (Elzinga, 1987). Secretions from the tubular labial and mandibular salivary glands solubilize food material and transport it into the mouthparts. Therefore, there is direct contact between caterpillar oral secretions and plant tissue. Elicitors, such as volicitin (N-17-hydroxylinolenoyl)-L-glutamine) or GOX, present in the caterpillar oral secretions are recognized by and influence the plant's responses either by inducing or suppressing defense mechanisms (Halitschke et al., 2001;Voelckel and Baldwin, 2004; Musser et al., 2002). By cauterizing the caterpillar spinneret, thereby impairing labial salivary secretions, Musser et al. (2002) elegantly demonstrated that GOX interferes with herbivore-induced nicotine production in tobacco plants. This action is believed to be mediated through hydrogen peroxide (H2O2), a product of glucose oxidation catalyzed by GOX. In plants, the signalling molecule, H2O2), mediates numerous pleiotrophic responses including localized cell death during the hypersensitive response in incompatible plant-pathogen interactions (De Gara et al., 2003; Vandenabeele et al., 2003). H2O2 produced by secreted caterpillar GOX is believed to act as an upstream signal leading to the biosynthesis of phytohormones, such as salicylic acid or ethylene, that interfere with jasmonate-dependent responses of the plant, such as herbivore-induced nicotine biosynthesis (Felton and Korth, 2000; Musser et al., 2002; Zong and Wang, 2004; Musser et al., 2005a, Musser et al., 2005b).
In the corn earworm, Helicoverpa zea, GOX activity is primarily detected in the labial salivary glands of late larval caterpillars (5th and 6th instars) and associated with feeding activity, in contrast to insect species in other families, where GOX was detected in the hemolymph and cuticle and associated with cuticle hardening during metamorphosis (Candy, 1979; Eichenseer et al., 1999). In actively-feeding caterpillars, GOX is secreted through the caterpillar spinneret (∼0.7 µg/hour, ∼0.095U) (Eichenseer et al., 1999). In addition to its role in modulating induced plant defenses, GOX may act as an oxygen scavenger, which could help maintain a relatively anaerobic midgut reducing the reactivity of plant secondary metabolites such as tannins and phenolics, or as an antimicrobial agent against insect pathogens present on the leaf surface (Eichenseer et al., 1999; Musser et al., 2002).
The activity of salivary GOX in Lepidopteran species is highly variable; enzymatic activity was detected in salivary homogenates of caterpillars of the beet armyworm, Spodoptera exigua, and the Bertha armyworm, Mamestra configurata, but not in caterpillars of other species, including other Noctuids, such as the true armyworm, Pseudaletia unipuncta, or the specialist alfalfa butterfly, Colias eurytheme (Merkx-Jacques and Bede 2004). During these studies, we observed that beet armyworm labial salivary GOX activity was high when caterpillars were reared on artificial diet, but negligible if caterpillars were reared on plants. If caterpillar salivary GOX is a mechanism to thwart induced plant defense responses, then the effect of diet on enzyme activity must be understood. Therefore, this study was conducted to characterize whole body and labial salivary GOX activity in beet armyworm caterpillars reared on either a wheat-germ based artificial diet (BioServ) or Medicago truncatula plants. GOX activity was measured in actively-feeding caterpillars over their larval development. Labial salivary GOX activity was also measured in caterpillars that were transferred from plant to artificial diet over a 12 hour period to determine if the change in GOX activity was diet-dependent.
Materials and Methods
Insect rearing
Eggs of the beet armyworm, Spodoptera exigua, were obtained from the GAST Insect Rearing Facility (Stoneville, MS) and the hatched caterpillars reared on a wheat germ-based artificial diet (Bio-Serv) in a growth chamber (16:8 hrs light: dark, 28-40% relative humidity; 28.5 ± 0.1 °C). Adult moths that eclosed from the collected pupae were allowed to mate and oviposit in a glass jar enclosure on paper towels.
Bio-Serv artificial diet
The Bio-Serv wheat-germ based diet contains: Agar (10.4 gm/L), wheat germ (120 gm/L), casein (25 gm/L), Wesson mineral mix (8 gm/L), methyl paraben (2 gm/L), sodium proprionate (2 gm/L), sorbic acid (2 gm/L), chlortetracycline (0.2 gm/L), cerelose (4.3 gm/L), vitamin A (0.22 gm/L), vitamin E (80 mg/L), pantothenic acid (9.2 mg/L), riboflavin (5 mg/L), folic acid (2.5 mg/L), niacin (10.1 mg/L), thiamine (2.2 mg/L), pyridoxine (2.1 mg/L), vitamin B12 (0.02 mg/L), biotin (0.20 mg/L), choline chloride (0.4 gm/L), ascorbic acid (5.0 gm/L) and inositol (0.2 gm/L). Diet was prepared according to manufacturer's instructions.
Plants
The legume Medicago truncatula A17 Jemalong was grown in a growth chamber (16:8 hrs light: dark, 22° C). Seeds were scarified in sulphuric acid and planted in pasteurized soil (80° C, 2 hr). Plants were watered with a dilute fertilizer solution (0.075% of nitrogen, phosphate and potassium) when needed. At six-weeks, plants were placed in new pots, and after one week were used for caterpillar feeding studies.
Developmental assay
Actively-feeding caterpillars, reared on artificial diet or M. truncatula, were categorized and separated according to instar at every moult. Larval instar was confirmed by colouration and head capsule and body size. GOX activity was measured in whole bodies, labial salivary glands, hemolymph or carcasses throughout larval development.
Whole body
Pooled caterpillars (2nd instar = 20 caterpillars; 3rd instar = 6 caterpillars; 4th and 5th instars = 1 caterpillar) were homogenized in Nathathan's saline (Christensen et al., 1991), containing proteinase inhibitor (Sigma, www.sigmaaldrich.com) to inhibit digestive proteases in the saliva and cellular proteases released during homogenation, and used in whole body GOX experiments. For each larval instar reared on different diets, six to ten separate replicates of the pooled caterpillars were analyzed.
Labial salivary gland dissections
Labial salivary glands were dissected from cooled 3rd, 4th or 5th S. exigua caterpillars under a stereomicroscope and immediately placed in Nathathan's saline containing proteinase inhibitor. GOX activity was determined in 6-9 pairs of pooled labial salivary glands. Each pool represents a single sample and for each larval instar on each diet between six to ten replicates were performed.
Hemolymph analysis
Hemolymph was collected by cutting the second to last caterpillar proleg and the hemolymph was milked into a microfuge tube containing Nathathan's saline and proteinase inhibitor. Total hemolymph collected was analyzed for GOX activity. For 4th and 5th instar caterpillars on the two diets, the hemolymph of six to ten insects was individually assayed for GOX activity.
Carcass dissections
Labial salivary glands were removed from cooled 4th and 5th instar caterpillars. The remaining carcass was homogenized in Nathansons' saline containing proteinase inhibitor and prepared for GOX analysis. The GOX activity was determined in individual caterpillar carcasses and between six to ten individuals were assayed at each larval instar reared on the different diets.
Glucose oxidase assay
Tissue, body homogenates or hemolymph were centrifuged to precipitate large particulates and the supernatant filtered through 0.22 µm low protein binding filters (Millipore, www.waters.com) before being assayed for GOX activity. An aliquot was removed and boiled in the presence of 1.5% SDS or 6 M urea and placed immediately on ice as the negative control. Fungal GOX (Sigma) was used as the positive control for GOX activity.
GOX activity was measured spectrophotometrically using an o-dianisidine-peroxidase coupled assay (Bergmeyer, 1974 (#bergmeyer74)). Solutions of horseradish peroxidase (Sigma) and glucose (Sigma) were prepared in ddH2O one hour in advance to allow mutarotation. Solutions of o-dianisidine (Sigma) were prepared in either 50 mM sodium acetate buffer, pH 5.1, 100 mM potassium phosphate buffer, pH 7.0 or 50 mM bicarbonate/carbonate buffer, pH 9.2. Oral secretions of caterpillars fed on artificial diet were in a neutral pH range (Bede, personal observations). In comparison, those reared on artificial diet had alkaline emissions that may reflect the presence of midgut regurgitant in the oral secretions. Therefore, GOX assays were conducted at pH values of 5.1, 7.0 and 9.2 to evaluate enzymatic activity over a biologically relevant pH range. Solutions were prepared and incubated at 35° C. The reaction cocktail, containing glucose (final concentration 0.1 mM) and o-dianisidine (final concentration 2.1 M), was saturated with oxygen. To this, horseradish peroxidase (final activity 2.2 U) and an aliquot of the pooled samples were added and the oxidation of o-dianisidine measured spectrophotometrically at 460 nm. For each replicate, three subsamples were taken and averaged.
Determination of protein content
Homogenate protein concentrations were measured using the Bradford method (1976) using bovine serum albumin as the protein standard. For each replicate, the average value of two subsamples was used to determine soluble protein amount.
Time course
Caterpillars were reared on artificial diet to maintain sufficient insects for these experiments. Before the assay, 3rd instar caterpillars were placed on plants for 2 days to allow their salivary GOX activity to reach basal levels. Fourth instar caterpillars were transferred to artificial diet for 2, 4, 6 or 12 hours. These times were staggered so that when the labial salivary glands were removed, the developmental time of the caterpillars was approximately the same. Six pairs of labial salivary glands were dissected, pooled, homogenized and analyzed for GOX activity and protein content. For each time point, between six to ten replicates were performed.
Calculation of percentage GOX activity in insect tissues
Total GOX activity per caterpillar in the labial salivary glands, carcass and hemolymph was divided by the total enzyme activity in whole bodies.
Statistical analysis
Means and standard errors were calculated using InStat (Graphpad, version 3.0). Normality residual plots confirmed that data followed an approximate Gaussian distribution. For the developmental assays, significant differences in GOX activity were determined by analysis of variance (ANOVA), followed by Tukey pairwise comparison (InStat, Graphpad). Differences in soluble protein levels were determined by unpaired Student T-test (InStat, Graphpad). For the time course experiment, differences in GOX activity were determined by an ANOVA followed by a Dunnet post-hoc test comparing the activity to GOX activity at T = 0.
Results
Glucose oxidase (GOX) activity
Whole body GOX activity of caterpillars reared either on plant diet or a wheat germ- based artificial diet was compared over larval development (Table 1, Fig. 1A). GOX activity increased throughout development to the highest levels observed in 5th instar caterpillar whole bodies. However, depending on the diet that the caterpillars were reared on, a striking difference in GOX activity was observed; whole body GOX activity of 5th instar caterpillars reared on a wheat germ-based artificial diet was ten times higher than when caterpillars were fed M. truncatula plants.
Table 1. Statistical analysis of glucose oxidase (GOX) activity of caterpillars reared on plant vs artificial diet.
Whole body GOX activities were compared by Student's t-test; labial salivary gland GOX activities were analyzed 1-way ANOVA followed by a Tukey post-hoc test. Significant differences are shown in Fig. 1.
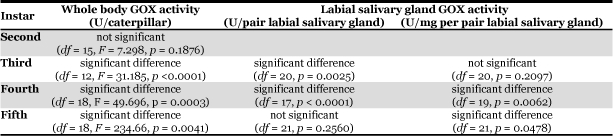
Fig. 1. Glucose oxidase (GOX) activity in Spodoptera exigua whole bodies (1A) and labial salivary glands (1B and 1C). 1A.
GOX activity in whole bodies of S. exigua caterpillars reared on Medicago trunctula or artificial diet over larval development (2nd to 5nd instars). GOX activity measured spectrophotometrically by the o-dianisidine-peroxidase assay (U/caterpillar, 35°C) (Bergmeyer, 1974). Bars represent the mean of 6-10 replicates ± SE. Significantly different means are indicated by alphabetical letters (p ≤ 0.01). Fig 1B and C. GOX activity in labial salivary gland of S. exigua caterpillars fed either plants or artificial diet. Bars represent the mean of between 6-10 replicates ± SE. Fig 1B: Total GOX activity per pair labial salivary gland. Fig 1C: GOX activity per mg soluble protein per pair labial salivary gland.
In many Noctuid caterpillar species, GOX is secreted during feeding by the labial salivary glands (Eichenseer et al., 1999). Labial salivary GOX activity was determined in S. exigua caterpillars reared on the two diets. The oral secretions of caterpillars may depend on diet; those reared on plant diet appear to regurgitate more frequently on their food producing an alkaline secretion, in comparison to those fed artificial diet, (Bede, personal observation). Therefore, labial salivary GOX activity was tested in the neutral range for caterpillars reared on the artificial diet and a neutral and basic pH for those reared on plants to represent biologically relevant situations.
Salivary GOX activity increased over development in caterpillars reared on plants (0.007 ± 0.002 to 0.009 ± 0.002 U/pair labial salivary glands in 3rd to 5th instar caterpillars, pH 9.2) (Table 1, Fig. 1B). In comparison, labial salivary GOX activity increased from 0.087 ± 0.015 U/pair labial salivary glands in 3rd instar caterpillars fed artificial diet to 0.145 ± 0.028 U/pair labial salivary glands in 4th instar caterpillars but decreased GOX levels were seen in 5th instar caterpillars (0.020 ± 0.005 U/pair labial salivary glands, pH 7.0) (Fig. 1B). Soluble protein in whole bodies and labial salivary glands of caterpillars reared on artificial diet was significantly greater than those fed plants (3rd, 4th and 5th; p < 0.002), which may partially account for the difference in labial GOX activity with smaller glands having less activity. However, when activity is standardized to the amount of soluble protein in the salivary gland, labial GOX activity in 4th larval instars reared on artificial diet was still significantly higher than those fed plants (p = 0.0062) (Table 1, Fig. 1C). This suggests that diet influences GOX activity.
In 5th instar caterpillars, GOX activity per mg soluble protein in the labial salivary glands is higher in caterpillars reared on plant diet than on artificial diet (p = 0.048) (Fig. 1C (ref/figure1.html)); however, this only reflects the smaller labial salivary glands of these insects. There is no significant difference in total GOX activity, which is low, in 5th instar caterpillars, regardless of the diet that they were reared on (p = 0.26).
The labial salivary glands of 4th instar caterpillars reared on artificial diet accounts for 57.8% of the GOX activity in whole bodies with the remainder found in the carcass (0.137 ± 0.026 U/caterpillar, pH 7.0). GOX activity was not detected in caterpillar hemolymph using the o-dianisidine-peroxidase spectroscopic assay. In comparison, the labial salivary gland GOX activity of 5th instar S. exigua represented only 6.8% of the whole body activity with the rest of the activity found in the carcass (0.304 ± 0.035 U/caterpillar, pH 7.0).
Time course
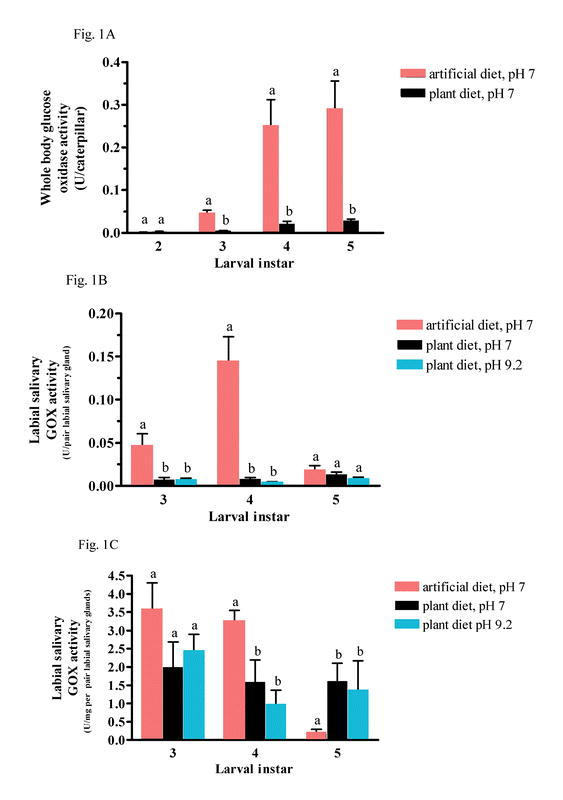
To determine conclusively if diet influences labial salivary GOX activity, plant-fed caterpillars with basal salivary GOX activity were transferred to artificial diet. The time of transfer onto the plants was staggered to ensure that caterpillars were approximately the same developmental age when labial salivary glands were removed and GOX activity determined. S. exigua caterpillars were allowed to feed continuously during the transfer from plant to artificial diet. Labial salivary GOX activity is closely correlated with the amount of time caterpillars spent feeding on artificial diet (Table 2, Fig. 2A). After transfer from plant to artificial diet, labial salivary GOX activity rose ten times from basal levels (0.430 ± 0.119 U/mg soluble protein per pair labial salivary gland, pH 7.0) to 4.59 ± 0.89 U/mg soluble protein per pair of labial salivary gland in 12 hours. A significant difference in protein content was not observed (0.025 ± 0.002 mg/pair labial salivary gland; p = 0.51) (Table 2, Fig. 2B).
Table 2. Statistical analysis of labial salivary glands soluble protein and glucose oxidase (GOX) activity.
Caterpillars were transferred from plant to artificial diet and allowed to feed for specified times. Times were staggered to ensure that caterpillars were approximately the same developmental age when the labial salivary glands were removed and assayed. Differences in protein and enzyme activity were analyzed by 1-way ANOVA followed by a Tukey post-hoc test. Significant differences are shown in Fig. 2.

Fig. 2. Labial salivary gland glucose oxidase (GOX) activity (2a) and protein levels (2b) after transfer of caterpillars from the plant Medicago trunctula to artificial diet.
Fourth instar Spodoptera exigua caterpillars were transferred from plant to artificial diet for 2, 4, 6 or 12 hours. Transfer times were staggered to ensure that the caterpillar developmental time when salivary glands were dissected is approximately equal. Labial salivary glands from six caterpillars were dissected, pooled and homogenate GOX activity and soluble protein amounts determined. Between six to ten replicates were performed. Points represent mean ± SE. Significantly different means are indicated by alphabetical letters (p ≤ 0.01). 2A. GOX activity (U/mg per pair labial salivary glands) 2B. labial salivary gland protein (mg).
Discussion
H2O2 produced by GOX secreted from labial salivary glands of H. zea caterpillars has been implicated in mediating the suppression of induced plant responses (Musser et al., 2002). As in H. zea, high labial salivary GOX oxidase activity has also been detected in caterpillars of the beet armyworm, S. exigua. However, in 4th instar S. exigua caterpillars salivary GOX activity is strongly dependent on diet; labial salivary GOX activity (U/mg soluble protein per pair labial salivary gland) in caterpillars reared on plants was significantly lower than if caterpillars had been fed artificial diet (p = 0.0062) (Fig 1C). This suggests that a factor found in the diet may influence salivary GOX levels. In comparison, total salivary GOX activity of 5th instar caterpillars is low, regardless of the diet the insects were reared on.
When 4th instar S. exigua caterpillars were transferred from plant to artificial diet, salivary GOX activity increased with time spent on the new diet; labial salivary GOX activity increased over three-fold in the 12 hours after caterpillars were transferred from plant to artificial diet. Therefore, a component in the diet influences caterpillar salivary enzyme activity. The activity of digestive enzymes is often regulated by diet; for example, midgut α-amylases are induced in response to starch content of their diet (Silva et al., 2001). This regulation may occur at many levels, including transcriptional and/or covalent modification of the enzymes (Lehane et al., 1995). However, with a few exceptions, the current paradigm is that the regulation of most digestive enzymes occurs at the transcriptional level. Gene expression of the salivary enzyme, α-amylase, is downregulated by dietary glucose in the fruit fly, Drosophila melanogaster (Hickey et al., 1994). Glucose is also a substrate for GOX, suggesting that a similar mechanism of glucose regulation of transcript levels may be present in these caterpillars.
The enzyme GOX has been detected in the salivary secretions and/or labial glands of caterpillars of S. exigua, H. armigera, H. assulta and H. zea (Eichenseer et al., 1999; Merkx-Jacques and Bede 2004; Zong and Wang, 2004). The labial GOX activity of 4th instar S. exigua reared on artificial diet (3.26 ± 0.028 U/mg per pair of labial salivary glands, pH 7.0) is in the range of activity found in other caterpillar species reared on artificial diet, such as H. assulta (0.16 ± 0.07 U/mg per pair of labial salivary glands), H. armigera (1.16 ± 0.78 U/mg per pair of labial salivary glands) and H. zea (2.37 ± 1.27 U/mg per pair of labial salivary glands) (Eichenseer et al., 1999; Zong and Wang, 2004).
This study shows that diet clearly affects 4th instar S. exigua caterpillar salivary GOX activity. The impact of plant diet on labial GOX activity of 5th instar H. zea caterpillars has recently been reported (Peiffer and Felton, 2005). Labial GOX activity was twice as high in caterpillars fed detached tobacco leaves for 48 hr compared to caterpillars fed cotton leaves, 0.118 compared to 0.055 U/ pair of labial salivary glands, respectively. In contrast, labial salivary activity of 4th instar S. exigua caterpillars reared on plant diet was 0.004 ± 0.001 U/pair of labial salivary glands, pH 9.2, compared to caterpillars fed artificial diet, 0.145 ± 0.028 U/pair of labial salivary glands, pH 7.0. Even though both these caterpillar species are generalists and Noctuids, clearly, diet influences the salivary composition of Lepidopteran species distinctly.
For convenience, or because of experimental limitations, many of us rear insects on artificial diet; however, this research shows that we must consider that these manipulations could affect the final outcome. In these experiments, labial salivary GOX activity of 4th instar beet armyworm caterpillars reared on artificial diet was over thirty times higher than if insects were reared on M. truncatula. Thus, if S. exigua reared on artificial diet were placed on plants, observed changes in induced plant defenses due to GOX might not represent the natural situation. Furthermore, salivary GOX activity is developmentally regulated.
The secretion of salivary GOX has been implicated as an adaptive strategy of caterpillars to undermine induced plant defense responses, possibly mediated through cross-talk between downstream pathways initiated by H2O2 and the octadecanoid pathways (Felton and Korth, 2000; Musser et al., 2002). Clearly, plant diet does not seriously impact GOX activity of H. zea caterpillars (Peiffer and Felton, 2005). In comparison, S. exigua labial GOX activity is at basal levels when caterpillars are reared on the legume, M. truncatula. It is unlikely that these levels are high enough to interfere with induced defenses in this plant. Our results indicate that in some Lepidopteran species, larval GOX activity is dependent on diet, which has profound implications for our understanding of plant-insect interactions and for the development of plant protection strategies based on these complex interactions. Future studies will focus on the identification of dietary factor(s) responsible for mediating these changes in GOX activity and determining the molecular mechanisms involved in this regulation.
Acknowledgments
The authors are grateful to Genevieve Nesbitt, Patrick Cournoyer and Annie Hibbert for technical assistance. We thank two anonymous reviewers and the Editor, Dr. Henry Hagedorn, for insightful comments and suggestions. This research was funded through grants from the Natural Sciences and Engineering Research Council (JCB) and Fonds de recherché sur la nature et les technologies (JCB).
References
- Ahmad SA, Hopkins TL. Phenol β-glucosyltransferase and β-glucosidase activities in the tobacco hornworm larvae Manduca sexta (L.): properties and tissue location. Archives of Insect Biochemistry and Physiology. 1992;21:207–224. [Google Scholar]
- Bergmeyer HU. 1974 Reagents for enzymatic analysis. In: Methods of Enzymatic Analysis Vol I. (Eds: Bergmeyer HU, Gawehn K) Verlag Chemie, Weinheim p. 494–495. [Google Scholar]
- Bradford MM. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Analytical Biochemistry. 1976;72:248–254. doi: 10.1006/abio.1976.9999. [DOI] [PubMed] [Google Scholar]
- Candy DJ. Glucose oxidase and other enzymes of hydrogen peroxide metabolism from cuticle of Schistocerca americana gregaria. Insect Biochemistry. 1979;9:661–665. [Google Scholar]
- Christensen TA, Itagaki H, Teal PEA, Jasensky RD, Tumlinson JH, Hildebrand JG. Innervation and neural regulation of the sex-pheromone gland in female Heliothis moths. Proceedings of the National Academy of Sciences USA. 1991;88:4971–4975. doi: 10.1073/pnas.88.11.4971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Gara L, de Pinto MC, Tommasi F. The antioxidant systems vis- á-vis reactive oxygen species during plant-pathogen interaction. Plant Physiology and Biochemistry. 2003;41:863–870. [Google Scholar]
- Eichenseer H, Mathews MC, Bi JL, Murphy JB, Felton GW. Salivary glucose oxidase: multifultional roles for Helicoverpa zea. Archives of Insect Biochemistry and Physiology. 1999;42:99–109. doi: 10.1002/(SICI)1520-6327(199909)42:1<99::AID-ARCH10>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- Elzinga RJ. 1987 Fundamentals of Entomology, 3rd Ed. Prentice Hall, New Jersey. [Google Scholar]
- Felton GW, Korth KL. Trade-offs between pathogen and herbivore resistance. Current Opinion in Plant Biology. 2000;3:309–314. doi: 10.1016/s1369-5266(00)00086-8. [DOI] [PubMed] [Google Scholar]
- Halitschke R, Schittko U, Pohnert G, Boland W, Baldwin IT. Molecular interactions between specialist herbivore Manduca Sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. III Fatty acid-amino acid conjugates in herbivore oral secretions are necessary and sufficient for herbivore-specific plant responses. Plant Physiology. 2001;125:711–717. doi: 10.1104/pp.125.2.711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickey DA, Benkel KI, Fong Y, Benkel BF. A Drosophila gene promoter is subject to glucose repression in yeast cells. Proceedings of the National Academy of Sciences USA. 1994;91:11109–11112. doi: 10.1073/pnas.91.23.11109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehane MJ, Blakemore D, Williams S, Moffatt MR. Regulation of digestive enzyme levels in insects. Comparative Biochemistry and Physiology. 1995;110B:285–289. [Google Scholar]
- Merkx-Jacques M, Bede JC. Caterpillar salivary enzymes: “eliciting” a response. Phytoprotection. 2004;85:33–37. [Google Scholar]
- Musser RO, Hum-Musser SM, Eichenseer H, Peiffer M, Ervin G, Murphy JB, Felton GW. Caterpillar saliva beats plant defenses. Nature. 2002;416:599–600. doi: 10.1038/416599a. [DOI] [PubMed] [Google Scholar]
- Musser RO, Kwon HS, Williams SA, White CJ, Romano MA, Holt SM, Bradbury S, Brown JK, Felton GW. Evidence that caterpillar labial saliva suppresses infectivity of potential bacterial pathogens. Archives of Insect Biochemistry and Physiology. 2005;58:138–144. doi: 10.1002/arch.20031. [DOI] [PubMed] [Google Scholar]
- Musser RO, Cipollini DF, Hum-Musser SM, Williams SA, Brown JK, Felton GW. Evidence that caterpillar salivary enzyme glucose oxidase provides herbivore offense in Solanaceous plants. Archives of Insect Biochemistry and Physiology. 2005;58:128–137. doi: 10.1002/arch.20039. [DOI] [PubMed] [Google Scholar]
- Peiffer M, Felton GW. The host plant as a factor in the synthesis and secretion of salivary glucose oxidase in larval Helicoverpa zea. Archives of Insect Biochemistry and Physiology. 2005;58:106–113. doi: 10.1002/arch.20034. [DOI] [PubMed] [Google Scholar]
- Silva CP, Terra WR, Xavier-Filho J, Grossi de Sá M, Isejimi EM, DaMatta RA, Miguens FC, Bifano TD. Digestion of legume starch granules by larva of Zabrotes subfasciatus (Coleoptera: Bruchidae) and the induction of α-amylases in response to different diets. Insect Biochemistry and Molecular Physiology. 2001;31:41–50. doi: 10.1016/s0965-1748(00)00103-x. [DOI] [PubMed] [Google Scholar]
- Vandenabeele S, Van Der Kelen K, Dat J, Gadjev I, Boonefaes T, Morsa S, Rottiers P, Slooten L, Van Montagu M, Zabeau M, Inzé D, Van Breusegem F.. A comprehensive analysis of hydrogen peroxide-induced gene expression in tobacco. Proceedings of the National Academy of Sciences USA. 2003;100:16113–16118. doi: 10.1073/pnas.2136610100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voelckel C, Baldwin IT. Generalist and specialist lepidopteran larvae elicit different transcriptional responses in Nicotiana attenuata, which correlate with larval FAC profiles. Ecology Letters. 2004;7:770–775. [Google Scholar]
- Zong N, Wang CZ. Induction of nicotine by herbivory and its relation to glucose oxidase activity in the labial gland of three noctuid caterpillars. Chinese Science Bulletin. 2004;49:4596–1601. [Google Scholar]