

Abstract
Plain-tailed wrens (Thryothorus euophrys) live in groups that sing synchronized choruses, the contributions of females and males alternating with each other in cycles, within which each sex sings two of the four parts, the whole achieving near perfect synchrony. As each bird has a repertoire of ca 20 phrases of each type, the synchrony also requires them to choose the same type at the same time as others of their sex. Songs can last up to 2 min, during which individuals join in and drop out. This must be one of the most complex singing performances yet described in a non-human animal.
Keywords: Thryothorus, wren, duetting, antiphony, chorus
1. Introduction
Song is largely a male prerogative in birds from north temperate regions (Catchpole & Slater 1995) and group singing is unusual. However, song in the tropics is often rather different: females frequently sing (Langmore 1998), in some cases males and females also join together to form duets (Hall 2004), and chorusing occurs in some species (e.g. Seddon 2002).
At their simplest, duets may consist of the two members of a pair singing in loose association with one another, or of one sex replying to the song of the other, usually with remarkably short reaction time so that the two songs overlap (Slater et al. 2002). In other cases, the two sexes sing rapidly repeated antiphonal cycles to which each contributes, often without overlapping with each other so that the song can sound as if produced by a single individual (Levin 1996). In a more complex form of duet, males have two phrases, one of which is sung on its own while the other alternates with that of the female in a series of cycles (Mann et al. 2003). Selection pressures on duet coordination may take two quite different routes depending on the taxonomic group, leading either to such precisely alternated, antiphonal songs or to accurately synchronized, unison songs (e.g. Wickler 1980). Here, we describe a song that incorporates both of these characteristics. It is also one stage more complex than this in that the species concerned, uniquely for its genus, lives in groups, the members of which perform highly coordinated choruses.
2. Methods
In autumn 2002, as part of a comparative study of duetting in many species of the genus Thryothorus, we studied the plain-tailed wren (Thryothorus euophrys) in bamboo thickets on the slopes of Pasachoa volcano, Ecuador. Previously thought to occupy territories in pairs throughout the year (Brewer 2001), as do its congeners, we found that the territories of this species were mostly occupied by groups of adults (mean size=4.2, range 2–7, n=13). In each group most or all individuals were caught by mist-netting and colour ringed with a unique combination. As the sexes are similar, sexing could not be reliably carried out by visual inspection in the hand. A blood sample of approximately 50 μl was therefore taken from each individual by puncturing the brachial vein. Samples were stored in 100 mM Tris, 100 mM EDTA, 2% SDS buffer to prevent DNA degeneration in the field. On return to St Andrews, the DNA was extracted using standard techniques and sex was determined according to the method of Griffiths et al. (1996).
For song analysis we selected four focal groups of different sizes: W (1♀1♂), B (2♀2♂), C (2♀3♂), R (5♀2♂) and took 7–9 h of song recordings from each group (Sennheiser ME66 microphone with Marantz CP430 cassette recorder), combining this with visual observations to determine the contributions of each individual in the group. All birds except one in group R were colour ringed. During observations we used short periods of playback of different songs recorded from other groups approximately every 45 min. This maximized the song output of the group under observation and encouraged it to reveal the full variety of its repertoire. It also attracted the group from the dense vegetation so that individuals gathered close to the loudspeaker where they could be identified and their contributions to the song observed. While it was not possible to monitor the precise contributions of multiple birds during a chorus, we were frequently very close to the birds, and could observe both the phrases produced by a particular bird and that these phrases often continued to be produced by others when that individual ceased to sing. Comments recorded at the time of the song recordings enabled us to identify which individual produced a solo song or, in a chorus, which group member was closest to the microphone. All songs were digitized, identified and compared spectrographically using Avisoft Saslab Pro, and the data entered on spreadsheets.
3. Results
(a) Song structure
The form of the song is shown in figure 1. Where only one male and female are singing (figure 1a), the duet has the general structure ABCDABCD… in a rapidly repeated cycle. The male contributes A and C, the female B and D, with normally no overlap between their contributions. In groups of more than two individuals, all members of each sex contributed, with same-sex birds singing the same phrases as one another in near perfect synchrony (figure 1b). A single continuous bout of such singing may go on for 40 or more such cycles (see table 1 for means) and last up to 2 min. During a bout of song, individuals may drop out or join in rather than contributing throughout. As table 1 shows, the mean number of cycles per song was lowest in the largest group (R), suggesting that it is not just lack of contributors that leads a song to be cut short.
Figure 1.
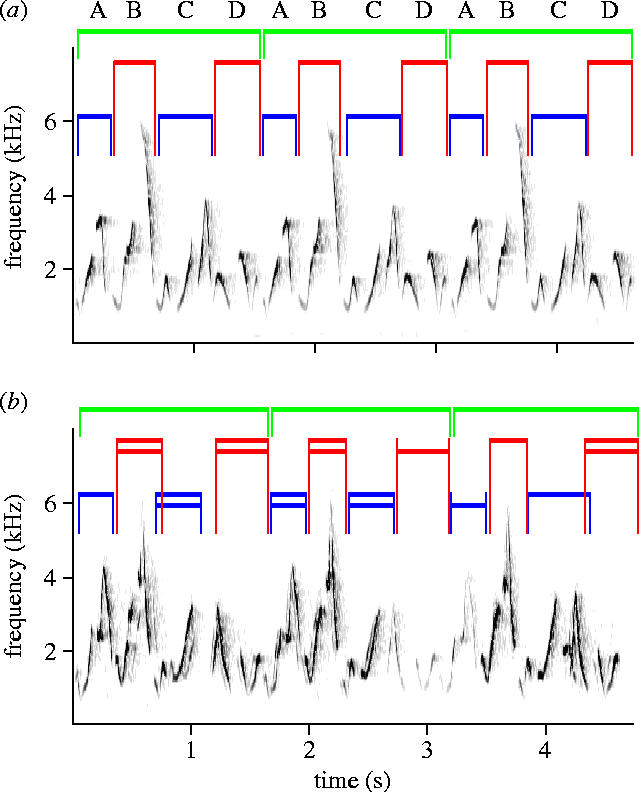
Sonograms (frequency/time plots) of two song sequences produced by the B group of plain-tailed wrens. Male contributions are labelled in blue and female in red. In (a) two birds are singing, (b) shows a different song type, all phrases being distinct from those above. Here, two males and two females are involved in 7 of the 12 phrases as shown by the red and blue lines; one bird of each sex is closer (upper coloured line) and the other more distant (lower coloured line). Where two birds are singing they sing the same phrase in synchrony, the slight time difference between them leading to an ‘echo’ visible on the sonogram trace and audible to the ear (sound files of these two examples and of a longer song sequence are available in the electronic supplementary material).
Table 1.
The song structure of four focal groups of plain-tailed wrens.
| percentages of quadruplets | ||||||
|---|---|---|---|---|---|---|
| group | songsa | mean cycles/song | solo songs ♂/♀ | all phrases match | two phrases conflict | all four phrases conflictb |
| W | 57 | 22.5 | 1/2 | 100 | 0 | 0 |
| C | 90 | 20.0 | 2/4 | 97.2 | 0.9 | 1.9 |
| B | 117 | 25.1 | 0/1 | 95.8 | 0.5 | 3.7 |
| R | 152 | 11.4 | 0/62 | 92.2 | 4.1 | 3.8 |
A song is defined as a continuous series of phrases, whether solos, duets or choruses, uninterrupted by a gap greater than 2 s.
Conflict is where individuals of one sex or both sing different pairs of phrases at the same time.
In rare cases (less than 5%), two members of the same sex within a group produce a different phrase at the same time and, occasionally, where the group consists of two or more of each sex, two songs may be performed at the same time either synchronously or without tight synchrony by different subgroups (see last column in table 1). Group R is particularly interesting (and its output complex), because the sex ratio is highly biased. Unlike other groups, BD combinations were frequently produced solo, without contributions from the two males in the group.
(b) Song repertoires
As table 2 shows, each sex has a separate repertoire of different phrases for each of the two positions it can occupy in the sequence. The form of phrases in each position is distinct: most obviously, B phrases sweep up and down across a broader frequency range than the other three types (see the two examples in figure 1). The phrases are combined such that, within a group, particular A and C phrases nearly always go together, as do particular B and D combinations. Each such combination could be viewed as a single phrase type within which a gap is left for the contribution of the other sex. However, the phrases of the two sexes are also highly associated with each other. As table 2 indicates, there are far fewer ABCD combinations than would be expected from random association of the AC and BD possibilities. Even the relatively small number of combinations found may be inflated because occasionally switches between one phrase type and another appeared to occur within a song in such a way that a quadruplet was a hybrid between the preceding and succeeding ones.
Table 2.
The song repertoires of four focal groups of plain-tailed wrens.
| repertoire (no. of phrase types in each category) | combinations observed (expectedb) | |||||||
|---|---|---|---|---|---|---|---|---|
| group | ABCD bouts recordeda | A(♂) | B(♀) | C(♂) | D(♀) | AC | BD | ABCD |
| W | 45 | 21 | 18 | 22 | 18 | 24 (42.9) | 28 (42.1) | 27 (42.8) |
| C | 69 | 20 | 16 | 19 | 16 | 20 (63.2) | 18 (60.6) | 21 (62.9) |
| B | 114 | 27 | 16 | 27 | 15 | 32 (105.6) | 19(90.9) | 37 (104.1) |
| R | 91 | 26 | 30 | 27 | 28 | 31 (85.4) | 37 (86.3) | 53 (87.5) |
A bout is defined as a series of cycles with the same four phrase types. More than one of these may occur in a single song, while solo songs or incomplete songs may include none.
The expected values given are based on the predictions that A and C are randomly sampled to form AC, B and D to form BD and AC and BD to form ABCD. Matlab was used to resample 10 000 times. As an example, in group W, 21A phrases and 22C phrases gives 462 possible AC combinations. Random sampling among these suggests that 42.9 combinations should be observed on average when 45 bouts are recorded. In fact, only 24 were found indicating that combinations are far from random.
In three of the four groups, the repertoires of the female phrases (B and D) were smaller than those of the male ones (A and C) (see table 2). However, in group R the opposite is the case. This probably results from this group having a sex ratio biased towards females. It may be that individual females in this group do have smaller repertoires, but that these differ somewhat from each other so that their combined repertoire is larger than that of the two males. If this is the case, it implies that there are certain quadruplets to which some individuals are unable to contribute.
(c) Initiation and termination
The different phrases within the song were labelled A, B, C and D because this was consistent with its rhythm, the interval between D and A being longer in all groups than those between the other phrases (table 3). However, as table 3 also shows, song in all groups most often starts with D, and initiations by B or D account overall for nearly 80% of songs. While the overall results on terminations are not significant, again nearly 80% of songs are terminated by either B or D. These results suggest that songs in this species are usually initiated by females and come to an end when males fail to contribute.
Table 3.
Percentage of song cycles started and finished with each phrase, and mean latencies between phrases (Friedmann tests, 3 degrees of freedom).
| initiation (%) | termination (%) | mean latency (ms) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | A | B | C | D | A–B | B–C | C–D | D–A | |
| W | 23 | 14 | 2 | 61 | 8 | 15 | 36 | 41 | 35 | 35 | 48 | 55 |
| C | 23 | 19 | 11 | 47 | 5 | 39 | 5 | 51 | 36 | 39 | 17 | 43 |
| B | 12 | 10 | 2 | 76 | 6 | 21 | 21 | 52 | 40 | 37 | 38 | 55 |
| R | 14 | 32 | 2 | 52 | 4 | 48 | 8 | 40 | 38 | 35 | 28 | 42 |
| χ2=11.1, p<0.02 | χ2=7.6, p<0.10 | χ2=7.7, p<0.10 | ||||||||||
4. Discussion
The song of the plain-tailed wren combines three characteristics that have been separately described in other tropical species. First, male and female components alternate very precisely with little or no overlap between them. This song form resembles the precise interchanges seen in some other Thryothorus species (Levin 1996; Mann et al. 2003), though these only alternate two components. Second, within a sex, individuals sing the same phrase with near perfect synchrony. The unison song of the African forest weaver (Symplectes bicolor) is perhaps the nearest equivalent here (Wickler 1980). Third, more than two individuals sing at the same time to form a chorus. While loose chorusing has been described in many species, perhaps the closest to this tight synchrony within a group of individuals is in the song of the white-browed sparrow weaver (Plocepasser mahali) studied by Wingfield & Lewis (1993). Although this was not described in detail, choruses in groups of different sizes had the same structure, implying tight synchrony. The remarkable feature of song in the plain-tailed wren is that all of these three characteristics are combined in one song performance. In addition, the fact that each song cycle consists of four components, rather than the two usually found in the cyclical duets of other species, adds a further layer of complexity.
While duetting is common in the genus Thryothorus, nothing as complex as the song described here has been reported before. A critical factor that has led to its evolution is likely to be the fact that this species lives in groups, whereas others do not. In other Thryothorus species we have studied, juveniles may remain on the natal territory for several months following independence, and they sometimes join in their parents' duets. Plain-tailed wren groups are probably an extension of this, with young remaining to help their parents' breeding efforts, as occurs in many tropical bird species (Emlen 1991). A low frequency of assistance by members of previous broods has recently been described in another Thryothorus species, the buff-breasted wren (T. leucotis) (Gill 2004).
Why then do the members of a group join to produce a complex chorus? The two hypotheses most commonly put forward for duetting are mate guarding and mutual territorial defence (Hall 2004). The former seems unlikely to be relevant here, as more than two individuals are involved. The latter is more plausible, particularly as playback leads chorusing birds to gather round the speaker. The close presence of several birds singing in a coordinated fashion is then very obvious to a human observer and could be especially intimidating to intruding wrens.
Another hypothesis suggested for duetting is that it may may help to synchronize the reproductive efforts of the members of a pair (e.g. Slater & Mann 2004). Duetting is largely restricted to the tropics, where external cues are less prevalent, and, by its very nature, involves equal amounts of mutual stimulation. The tightly synchronized chorusing of plain-tailed wrens might synchronize the efforts, not just of the breeding pair, but of other members of the group as well. At present these hypotheses remain untested, but we plan further work on this species to understand its breeding system and the role within it of this remarkable song.
Acknowledgments
We thank the Leverhulme Trust and the Royal Society of Edinburgh for financial support, Jeff Graves for help with molecular analysis, him, Vincent Janik and Constantino Macias for suggestions, and Luke Rendell for help with the resampling.
Supplementary Material
References
- Brewer D. Christopher Helm; London: 2001. Wrens, dippers and thrashers. [Google Scholar]
- Catchpole C.K, Slater P.J.B. Cambridge University Press; Cambridge, UK: 1995. Bird song. Biological themes and variations. [Google Scholar]
- Emlen S.T. Evolution of cooperative breeding in birds and mammals. In: Krebs J.R, Davies N.B, editors. Behavioural ecology: an evolutionary approach. Blackwell Scientific Publications; Oxford, UK: 1991. pp. 301–337. [Google Scholar]
- Gill S.A. First record of cooperative breeding in a Thryothorus wren. Wilson Bull. 2004;116:337–341. [Google Scholar]
- Griffiths R, Daan S, Dijkstra C. Sex identification in birds using two CHD genes. Proc. R. Soc. B. 1996;263:1251–1256. doi: 10.1098/rspb.1996.0184. [DOI] [PubMed] [Google Scholar]
- Hall M.L. A review of hypotheses for the functions of avian duetting. Behav. Ecol. Sociobiol. 2004;55:415–430. doi:10.1007/s00265-003-0741-x [Google Scholar]
- Langmore N.E. Functions of duet and solo songs of female birds. Trends Ecol. Evol. 1998;13:136–140. doi: 10.1016/s0169-5347(97)01241-x. doi:10.1016/S0169-5347(97)01241-X [DOI] [PubMed] [Google Scholar]
- Levin R.N. Song behaviour and reproductive strategies in a duetting wren, Thryothorus nigricapillus. I. Playback experiments. Anim. Behav. 1996;52:1107–1117. doi:10.1006/anbe.1996.0258 [Google Scholar]
- Mann N.I, Marshall-Ball L, Slater P.J.B. The complex song duet of the plain wren. Condor. 2003;105:672–682. [Google Scholar]
- Seddon N. The structure, context and possible functions of solos, duets and choruses in the subdesert mesite (Monias benschi) Behaviour. 2002;139:645–676. doi:10.1163/15685390260136753 [Google Scholar]
- Slater P.J.B, Mann N.I. Why do the females of many bird species sing in the tropics? J. Avian Biol. 2004;35:289–294. doi:10.1111/j.0908-8857.2004.03392.x [Google Scholar]
- Slater P.J.B, Gil D, Barlow C.R, Graves J.A. Male led duets in the moho (Hypergerus atriceps) and yellow-crowned gonolek (Lanarius barbarus): mate guarding by females. Ostrich. 2002;73:49–51. [Google Scholar]
- Wickler W. Vocal duetting and the pair bond. II. Unison duetting in the African forest weaver, Symplectes bicolor. Z. Tierpsychol. 1980;52:217–226. [Google Scholar]
- Wingfield J.C, Lewis D.M. Hormonal and behavioural responses to simulated territorial intrusion in the cooperatively breeding white-browed sparrow weaver, Plocepasser mahali. Anim. Behav. 1993;45:1–11. doi:10.1006/anbe.1993.1001 [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.