

Abstract
Understanding El Niño/Southern Oscillation (ENSO) and its biological consequences is hindered by a lack of high-resolution, long-term data from the tropical western Pacific. We describe a preliminary, 6 year dataset that shows tightly coupled ENSO-related bio-physical dynamics in a seawater lake in Palau, Micronesia. The lake is more strongly stratified during La Niña than El Niño conditions, temperature anomalies in the lake co-vary strongly with the Niño 3.4 climate index, and the abundance of the dominant member of the pelagic community, an endemic subspecies of zooxanthellate jellyfish, is temperature associated. These results have broad relevance because the lake: (i) illustrates an ENSO signal that is partly obscured in surrounding semi-enclosed lagoon waters and, therefore, (ii) may provide a model system for studying the effects of climate change on community evolution and cnidarian–zooxanthellae symbioses, which (iii) should be traceable throughout the Holocene because the lake harbours a high quality sediment record; the sediment record should (iv) provide a sensitive and regionally unique record of Holocene climate relevant to predicting ENSO responses to future global climate change and, finally, (v) seawater lake ecosystems elsewhere in the Pacific may hold similar potential for past, present, and predictive measurements of climate variation and ecosystem response.
Keywords: climate, El Niño–Southern Oscillation, marine lake, population dynamics, Scyphozoa, zooxanthellae
1. Introduction
El Niño/Southern Oscillation (ENSO) is the largest perturbation of global and regional climate on inter-annual time scales. Although ENSO events typically originate in the far western equatorial Pacific (Clarke & Van Gorder 2001), limited empirical data exist describing ENSO-related physical and biological dynamics in this key region of the Pacific ocean-climate system.
Palau, situated on the northwest edge of the western Pacific warm pool (at 7°N 134°E), proximate to the western equatorial Pacific origin of ENSO events (Clarke & Van Gorder 2001), appears well positioned to harbour a sensitive indicator of ENSO. ENSO-related meteorological conditions, for example, influence sea surface salinity (Morimoto et al. 2002) and, along with elevated sea temperatures, possibly contribute to coral bleaching (Bruno et al. 2001). During the strong 1998 La Niña event, Jellyfish Lake, a seawater lake situated within one of Palau's karst islands, experienced dramatic bio-physical perturbations including unusually strong stratification, high temperatures, and disappearance of millions of medusae (Dawson et al. 2001), indicating that the lake may be sensitive to ENSO-related climatic conditions. Here, we interpret 6 years of data, collected since 1998, describing physical and biological variation in Jellyfish Lake in relation to ENSO variation.
2. Material and methods
(a) Data collection
Jellyfish Lake, known locally as Ongeim'l Tketau (OTM; 07°09.83′ N 134°22.50′ E), is a brackish lake stratified by an excess of precipitation over evaporation and limited wind and tidal forcing (Hamner et al. 1982; Hamner & Hamner 1998). The upper mixed layer (the ‘mixolimnion’, 0–12 m depth) is physically and biotically variable and floats atop an anoxic, 18 m deep hypolimnion from which aerobic eukaryotes are precluded.
Monthly from December 1998 to February 2005 (bar February–May and July 2004) we measured temperature, salinity and oxygen concentration at 1 m intervals from 0 to 24 m at two sites with hand-held YSI 85 or Hydrolab Quanta meters. For each sample date, we calculated the average temperature from 0 to 12 m (mean mixolimnion temperature, MMT) and the density difference between the top (0 m) and bottom (12 m) of the mixolimnion (mixolimnion density difference, MDD, sigma t). MMT's for February–May and July 2004 were estimated from Onset HOBO Pro-8 thermograph data, recorded at half-hour intervals at 6 and 11 m, by averaging one 24 hour period near the middle of each month and converting this to MMT using the regression MMT=−1.41+1.04HOBO6,11m (r=0.98, p<0.001; calculated using manually collected MMT and HOBO thermograph data from April 2002 to January 2004). Five additional MMTs and MDDs were estimated from profiles taken between 1979 and 1997 (see Hamner & Hamner 1998).
Beginning October 2000, we quantified the abundance of Mastigias cf. papua etpisoni Dawson (hereafter, referred to as Mastigias because the species attribution is unclear) monthly using a 0.5 m diameter, 1 mm mesh net hauled vertically from the bottom of the mixolimnion to the surface. (Visual surveys from December 1998 to September 2000 revealed medusae were absent or in densities too low to quantify (e.g. Dawson et al. 2001)) Net hauls were distributed evenly across the lake at 15 stations, and the set of 15 stations was sampled as many (two or three) times as possible during the course of one day. The bell diameter (BD) of each medusa was measured to the nearest 0.5 cm.
Sea temperatures in Palau were measured at 30 min intervals at lagoon (10 m; 7°19.008 N 134°27.627′ E) and barrier-reef (11, 55, 90 m; 7°17.453′ N 134°14.442′ and/or 7°16.418′ N 134°31.440′ E) sites, using onset thermographs in pressure proof housings, and averaged to produce one mean temperature per calendar month per depth. Sea temperature anomalies in the Niño 3.4 region (05°N–5°S, 120°–170°W), an ENSO sensitive area of the eastern Pacific integral to global climate variation and used extensively in forecasting (iri.columbia.edu/climate/ENSO/background/monitoring.html), were downloaded from www.cpc.ncep.noaa.gov.
(b) Analyses
MMT, barrier-reef and lagoon temperature and MDD anomalies (indicated by subscript A) were calculated by subtracting the monthly mean, calculated for the period of each dataset, from each monthly value. A 12 month cross correlation window was used to assess lead/lag patterns of correlation between Niño 3.4 and MMT and Palau barrier-reef and lagoon anomalies. Except where noted, all datasets were filtered using a 2 month running mean prior to analysis. Correlations of lagoon and Niño 3.4 anomalies were based on 3 month running means to normalize lagoon data. Prior to smoothing, MDDA was ln[X+10] transformed.
The relationship between the abundance of medusae in each 0.5 cm size-class (bar size-classes ≥8 cm which were pooled due to low numbers) and MMT was examined over an 8 month lead/lag window. The same window was used to compare medusae abundance to the Niño 3.4. Prior to analysis, medusae time series were filtered with a 3 month running median to reduce the influence of noise and skew associated with sampling biological populations; for compatibility, lake and ocean temperatures were smoothed with a 3 month running mean. We used the modified Chelton method to adjust sample sizes for autocorrelation (Pyper & Peterman 1998). Normality was assessed using Kolmogorov–Smirnov and Lillefors tests. Medusae densities were ln[X+1] or transformed. Statistica 6.1 was used for all analyses.
3. Results
MMTA was strongly negatively correlated with sea surface temperature anomalies in the Niño 3.4 region (figure 1a). The strongest correlation was observed at an MMTA lead of 3 months (r=−0.91, p<0.05; figure 1b). MMTA was significantly positively correlated with MDDA at a lag of zero (r=0.85, p≤0.05; figure 1c). The five pairs of (unfiltered) pre-1998 values fall within or close to the 95% prediction intervals of the respective filtered regressions (figure 1c,d).
Figure 1.

(a) Mean mixolimnion temperature anomaly (MMTA) in Ongeim'l Tketau and unfiltered Niño 3.4 sea surface temperature anomaly between December 1998 and February 2005. El Niño or La Niña events defined as six consecutive months of anomalies >±0.4 °C (Trenberth 1997). (b) Strength of correlation between water temperature anomalies in Palau when defined as leading (positive) or lagging (negative) the Niño 3.4 by up to 8 months. (c) Relationship between MMTA and mixolimnion density difference anomaly (MDDA; transformed) at a lag of zero (r=0.85, p≤0.05). (d) Relationship between MMTA and Niño 3.4 when MMTA leads Niño 3.4 by 3 months (r=−0.91, p<0.05). In (c) and (d) the crosses (labelled with month and year (m/yr)) show unfiltered single-month measurements from one neutral (1979), three strong (1987, 1991 and 1997), and one weak (1994) El Niño years. Dashed lines define the 95% prediction interval.
Palau barrier-reef and lagoon anomalies were more weakly related to Niño 3.4 (figure 1b). Barrier-reef and Niño 3.4 anomalies were most strongly correlated at a Niño 3.4 lead of 1–2 months (−0.79≤r ≤−0.80; figure 1b). Lagoon and Niño 3.4 anomalies were most strongly associated with the Niño 3.4 at both lag and lead positions (lag −1, r=−0.63, p<0.05; lag +1, r=−0.62, p <0.02; figure 1b).
Densities of Mastigias medusae ≥2 cm BD were significantly and most strongly negatively correlated with MMT at a lag of zero (r=−0.88; p≤0.02; figure 2). Consequently, densities of Mastigias medusae ≥2 cm BD were significantly and most strongly positively associated with Niño 3.4 anomalies three months in the future (r=0.89, p<0.05).
Figure 2.
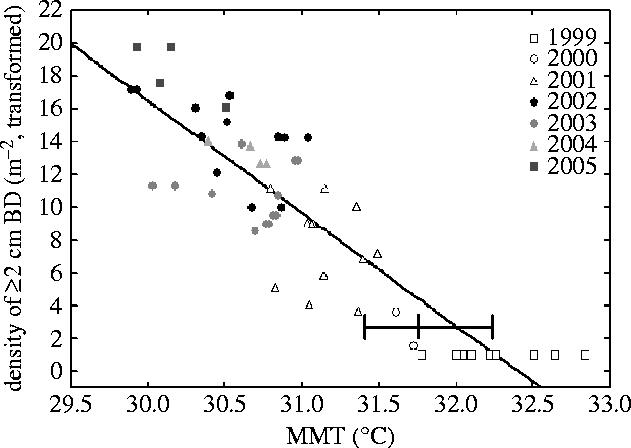
Relationship between abundance of Mastigias medusae ≥2 cm BD and MMT in Ongeim'l Tketau (r=−0.88; p≤0.02). Horizontal bar illustrates the range and mean (cross bar) temperatures between January and September 2000 when medusae were present but in abundances too low to quantify.
4. Discussion
Current understanding of ENSO dynamics is limited by a dearth of data describing physical conditions near the average origin of ENSO events in the far western equatorial Pacific (Clarke & Van Gorder 2001). Detailed studies of population dynamics reflecting the influence of ENSO are also lacking from this region. Our preliminary measurements in OTM thus provide a novel, high resolution and sensitive, regionally unique source of coupled bio-physical data demonstrably related to ENSO.
Although short by climatological standards, our 6 year dataset is complemented by occasional measurements spanning 25 years which together suggest strongly that fluctuations in MMTA are ENSO-related and precede fluctuations in Niño 3.4 by an average of 3 months (figure 1b). This timing is consistent with Palau's early position in the eastward progression of ENSO (Clarke & Van Gorder 2001) and suggests MMTA provides a sensitive, simply measured, and spatially explicit indicator suited to improving Niño 3.4 prediction, including its broader global climate implications, on intra-annual time scales (Clarke & Van Gorder 2003). The predictive skill of our simple linear model (−0.75≤r≤−0.91 for 2–7 month leads, e.g. figure 1d, y=−1.02 (s.e.±0.05)x+0.02 (s.e.±0.04)) equals that of the more complex statistical models of Clarke and Van Gorder (2001, 2003; 0.60≤r≤0.93 for 2–7 month leads) and Penland and Magorian (1993; r=0.84−0.87 for a 3 month lead).
That OTM captures a genuine climate signal attenuated in surrounding coastal waters is supported by the relationship between temperatures in the semi-enclosed shallow lagoon and the Niño 3.4. Here, the leading signal is diminished relative to MMTA, a situation that likely reflects the influence of barrier reef waters whose temperature fluctuations predominantly lag the Niño 3.4 (figure 1b). This observation also helps clarify the physical attributes that likely foster OTM's unique sensitivity. OTM is relatively small with commensurately less thermal inertia than the coastal ocean. OTM is also landlocked with limited tidal flux, which reduces mixing and, critically, physically largely isolates it from two of the three processes that influence temperatures in the surrounding ocean: horizontal advection and deep-water entrainment (Clarke et al. 1998). Thus, MMTA must predominantly be a function of the third process: atmosphere–ocean (in this case atmosphere–lake) interactions. The significant positive association between MMTA and MDDA (figure 1c) is consistent with this interpretation and indicates that temperature is modulated in large part by changes in stratification effected by local weather patterns related to climate fluctuations, as is the case in other tropical Pacific saline lakes (Trichet et al. 2001). Specifically, increased precipitation (relative to evaporation) associated with La Niña conditions increases surface stratification, which by increasing resistance to mixing promotes heat retention and warmer MMTAs (e.g. Dawson et al. 2001). Conversely, decreased rainfall and increased rates of evaporation associated with El Niño conditions de-stratify the water column, likely through a salt finger like mechanism (Schmitt et al. 2005), to promote mixing, heat loss and cooler mixolimnion temperatures.
OTM's sensitivity to ENSO conditions resulted in exceptionally high water temperatures throughout the strong La Niña year of 1999 (the highest single measurement reached 35.2 °C), which also coincided with the absence of the typically millions-strong perennial population of zooxanthellate Mastigias medusae (Dawson et al. 2001). The population's recovery began with a marked cooling of the lake in January 2000 and 6 years of data now demonstrate that the abundance of Mastigias medusae is closely associated with lake temperature and ENSO dynamics (figure 2). This tightly coupled bio-physical relationship is likely mediated by temperature effects on the zooxanthellae–Mastigias photosymbiosis (Dawson et al. 2001), a mechanism that influences population dynamics of other zooxanthellate cnidarians (Bruno et al. 2001). Collectively, the rapid feedback of temperature on medusae survival, the lake's ENSO-linked dynamic thermal regime, and its location in the relatively aseasonal tropics suggest OTM may be a useful model system for long-term study of these ecologically important multi-faceted associations (Fagoonee et al. 1999). (For studies of temperate azooxanthellate scyphozoans and climate in oceanic settings, see Lynam et al. (2004, 2005), Purcell (2005) and Purcell & Decker (2005).)
OTM thus offers a novel high-quality source of coupled bio-physical data from a key but under-studied region of the Pacific, where ENSO related climate variation is much more strongly reflected in sea surface salinity (Morimoto et al. 2002). These data may contribute to better contemporary predictions of ENSO and, therefore, regional and global climate on intra-annual times-scales. In addition, because OTM has a depositional environment ideal for the preservation of a high quality sediment record (Burnett et al. 1989), the lake likely warehouses a highly resolved record of at least Holocene climate and ENSO dynamics that is relevant to understanding how ENSO may vary with climate change (Urban et al. 2000). Moreover, this record is, in part, a biological record, potentially allowing the reconstruction of over 10 000 years of evolution in a tropical marine ecosystem (Dawson & Hamner 2005).
Other marine lake ecosystems, of which many occur across the Pacific, may similarly illustrate local atmospheric changes driven by contemporary global climate fluctuations. They may also harbour high-resolution sedimentary records of climate history (e.g. Trichet et al. 2001), and exhibit ecological responses relevant to understanding climatic influences on tropical marine ecology. Collectively, marine lakes may be one of the most valuable yet under-explored resources for understanding climate and tropical marine ecology and evolution.
Acknowledgments
We thank the many who assisted Coral Reef Research Foundation with data collection, especially Emilio Basilius, Koror State for their long-term support of this work, and A. Clarke and N. Colasacco for discussing ENSO. CRRF supported this work financially, with Marine Resources Pacific Consortium providing partial funding for field costs during 2000–2004. Two anonymous reviewers provided comments that helped improve the clarity of the manuscript.
References
- Bruno J.F, Siddon C.E, Witman J.D, Colin P.L, Toscano M.A. El Niño related coral bleaching in Palau. Western Caroline Islands. Coral Reefs. 2001;20:127–136. doi:10.1007/s003380100151 [Google Scholar]
- Burnett W.C, Landing W.M, Lyons W.B, Orem W. Jellyfish Lake, Palau: a model anoxic environment for geochemical studies. EOS. 1989;70:777–783. [Google Scholar]
- Clarke A.J, Van Gorder S. ENSO prediction using an ENSO trigger and a proxy for equatorial Pacific warm pool movement. Geophys. Res. Lett. 2001;28:579–582. doi:10.1029/2000GL012201 [Google Scholar]
- Clarke A.J, Van Gorder S. Improving El Niño prediction using a space–time integration of Indo-Pacific winds and equatorial Pacific upper heat content. Geophys. Res. Lett. 2003;30:1399. doi:10.1029/2002GL016673 [Google Scholar]
- Clarke A.J, Liu X, Van Gorder S. Dynamics of the biennial oscillation in the equatorial Indian and far western Pacific oceans. Am. Meteorol. Soc. 1998;11:987–1001. [Google Scholar]
- Dawson M.N, Hamner W.M. Rapid evolutionary radiation of marine zooplankton in peripheral environments. Proc. Natl Acad. Sci. USA. 2005;102:9235–9240. doi: 10.1073/pnas.0503635102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawson M.N, Martin L.E, Penland L.K. Jellyfish swarms, tourists and the Christ-child. Hydrobiologia. 2001;451:131–144. doi:10.1023/A:1011868925383 [Google Scholar]
- Fagoonee I, Wilson H.B, Hassell M.P, Turner J.R. The dynamics of zooxanthellae populations: a long-term study in the field. Science. 1999;283:843–845. doi: 10.1126/science.283.5403.843. doi:10.1126/science.283.5403.843 [DOI] [PubMed] [Google Scholar]
- Hamner W.M, Hamner P.P. Stratified marine lakes of Palau (Western Caroline Islands) Phys. Geogr. 1998;19:175–220. [Google Scholar]
- Hamner W.M, Gilmer R.J, Hamner P.P. The physical, chemical, and biological characteristics of a stratified, saline, sulfide, lake in Palau. Limnol. Oceanogr. 1982;27:896–909. [Google Scholar]
- Lynam C.P, Hay S.J, Brierley A.S. Interannual variability in abundance of North Sea jellyfish and links to the North Atlantic Oscillation. Limnol. Oceanogr. 2004;49:637–643. [Google Scholar]
- Lynam C.P, Brierley A.S, Hay S.J. Jellyfish abundance and climatic variation: contrasting responses in oceanographically distinct regions of the North Sea, and possible implications for fisheries. J. Mar. Biol. Assoc. UK. 2005;85:435–450. doi:10.1017/S0025315405011380 [Google Scholar]
- Morimoto M, Osamu A, Kayanne H, Kurita N, Matsumoto E, Yoshida N. Salinity records for the 1997–98 El Niño from western Pacific corals. Geophys. Res. Lett. 2002;29:35.1–35.4. doi:10.1029/2001GL013521 [Google Scholar]
- Penland C, Magorian T. Prediction of Niño 3 sea-surface temperature anomalies using linear inverse modeling. J. Climate. 1993;6:1067–1076. doi:10.1175/1520-0442(1993)006<1067:PONSST>2.0.CO;2 [Google Scholar]
- Purcell J.E. Climate effects on jellyfish and ctenophore blooms: a review. J. Mar. Biol. Assoc. UK. 2005;85:461–476. doi:10.1017/S0025315405011409 [Google Scholar]
- Purcell J.E, Decker M.B. Effects of climate on relative predation by scyphomedusae and ctenophores on copepods in Chesapeake Bay during 1987–2000. Limnol. Oceanogr. 2005;50:376–387. [Google Scholar]
- Pyper B.J, Peterman R.M. Comparison of methods to account for autocorrelation in correlation analyses of fish data. Can. J. Fish. Aquat. Sci. 1998;55:2127–2140. doi:10.1139/cjfas-55-9-2127 [Google Scholar]
- Schmitt R.W, Ledwell J.R, Montgomery E.T, Polzin K.L, Toole J.M. Enhanced diapycnal mixing by salt fingers in the thermocline of the tropical Atlantic. Science. 2005;308:685–688. doi: 10.1126/science.1108678. doi:10.1126/science.1108678 [DOI] [PubMed] [Google Scholar]
- Trenberth K.E. The definition of El Niño. Bull. Am. Meteorol. Soc. 1997;78:2771–2777. doi:10.1175/1520-0477(1997)078<2771:TDOENO>2.0.CO;2 [Google Scholar]
- Trichet J, Defarge C, Tribble J, Tribble G, Sansone F. Christmas Island lagoonal lakes, models for the deposition of carbonate–evaporite–organic laminated sediments. Sediment. Geol. 2001;140:177–189. doi:10.1016/S0037-0738(00)00177-9 [Google Scholar]
- Urban F.E, Cole J.E, Overpeck J.T. Influence of mean climate change on climate variability from a 155-year tropical Pacific coral record. Nature. 2000;407:989–993. doi: 10.1038/35039597. doi:10.1038/35039597 [DOI] [PubMed] [Google Scholar]