

Abstract
Eye lenses of various diurnal geckos contain up to 12% ι-crystallin. This protein is related to cellular retinol-binding protein type I (CRBP I) but has 3,4-didehydroretinol, rather than retinol, as a ligand. The 3,4-didehydroretinol gives the lens a yellow color, thus protecting the retina by absorbing short-wave radiation. ι-Crystallin could be either the gecko's housekeeping CRBP I, recruited for an additional function in the lens, or the specialized product of a duplicated CRBP I gene. The finding of the same CRBP I-like sequence in lens and liver cDNA of the gecko Lygodactylus picturatus now supports the former option. Comparison with ι-crystallin of a distantly related gecko, Gonatodes vittatus, and with mammalian CRBP I, suggests that acquiring the additional lens function is associated with increased amino acid changes. Compared with the rat CRBP I structure, the ι-crystallin model shows reduced negative surface charge, which might facilitate the required tight protein packing in the lens. Other changes may provide increased stability, advantageous for a long-living lens protein, without frustrating its role as retinol transporter outside the lens. Despite a number of replacements in the ligand pocket, recombinant ι-crystallin binds 3,4-didehydroretinol and retinol with similar and high affinity (≈1.6 nM). Availability of ligand thus determines whether it binds 3,4-didehydroretinol, as in the lens, or retinol, in other tissues. ι-Crystallin presents a striking example of exploiting the potential of an existing gene without prior duplication.
Keywords: adaptive evolution, gene sharing, protein modeling
To function properly, the vertebrate eye lens must contain high concentrations of water-soluble proteins, the crystallins (1, 2). In most vertebrates, the bulk of the lens proteins is formed by the β/γ-crystallins, a superfamily of stable proteins of obscure evolutionary origins. Large quantities of αA- and αB-crystallin are generally also present in the lens. These belong to the small heat-shock protein superfamily and may function as molecular chaperones to prevent aggregation of proteins in the lens. In addition to these ubiquitous crystallins, many species also express high levels of one or more of the so-called taxon-specific crystallins. All but one of the 11 characterized taxon-specific crystallins are identical or closely related to regular house-keeping enzymes, mostly oxidoreductases such as lactate dehydrogenase B and α-enolase (1–6). Some of these enzyme crystallins may have been recruited for their additional function in the lens because of the associated high levels of pyridine nucleotide cofactors, which may confer protection against oxidative damage (2, 7).
The only nonenzyme taxon-specific crystallin observed thus far is ι-crystallin (6), which is present at levels ranging from 2 to 12% in certain species of diurnal geckos (8). A partial protein sequence of this monomeric 16-kDa lens protein showed that it belongs to the family of intracellular lipid-binding proteins (iLBPs), being most closely related to cellular retinol-binding protein type I (CRBP I). This protein is ubiquitous and is involved in retinoid storage and transport (9, 10). However, although retinol is the normal ligand for CRBP I, ι-crystallin from gecko lens contains 3,4-didehydroretinol (vitamin A2) as a ligand. The 3,4-didehydroretinol⋅ι-crystallin complex gives the lenses of these geckos a yellow coloring and absorbs short-wave radiation. This characteristic improves the optical quality of the lens, and provides an excellent protection against ultraviolet damage to the retina. The latter seems particularly useful for diurnal geckos, which have no eyelids and cannot regulate the aperture of their pupil and live in habitats where they are exposed to high intensities of light (6).
The expression in gecko lenses of a CRBP I-like protein with the unusual ligand 3,4-didehydroretinol raises important evolutionary questions. First of all, is ι-crystallin the product of a duplicated CRBP I gene, allowing specific adaptations for functioning as a lens protein? Or rather, could ι-crystallin be the gecko's housekeeping CRBP I, overexpressed in the lens to gain a dual function? Second, has the structure of ι-crystallin been adapted to harbor 3,4-didehydroretinol as a ligand, and does it have other properties that might support its functioning in the lens? Finally, can the evolutionary pathways that led to the recruitment of ι-crystallin in various genera of geckos be reconstructed? To answer these questions, we determined the cDNA sequences for eye lens ι-crystallin and for its housekeeping homologue in the liver of the gecko Lygodactylus picturatus and found them to be identical. The obtained DNA sequence was used to reconstruct the phylogenetic relationship of ι-crystallin with other known CRBPs. A partial protein sequence of ι-crystallin from a distantly related gecko, Gonatodes vittatus, was obtained to reconstruct the evolutionary history of this protein more accurately. A three-dimensional ι-crystallin model was constructed to understand its ligand-binding and structural properties, and recombinant ι-crystallin of L. picturatus was produced to study its retinoid-binding behavior.
Materials and Methods
Cloning of ι-Crystallin.
Total RNA was isolated from three lenses of L. picturatus, with the TRIzol reagent (GIBCO/BRL), and cDNA was synthesized with the 5′/3′ rapid amplification of cDNA ends kit (Roche Molecular Biochemicals). Amplification was performed with a primer based on the partial protein sequence of ι-crystallin (ref. 6; ATGCCNCCNAAYTTYACNGG, corresponding to positions 1–6 in Fig. 1 and including the start codon) and an anchor primer from the rapid amplification of cDNA ends kit. The resulting cDNA was cloned into the pGEM-T vector (Promega) to give ι/pGEM-T, and three independent clones were sequenced (ALF Express, Amersham Pharmacia).
Figure 1.

ι-Crystallin sequences aligned with mammalian CRBP I and II. The topmost sequence, from position 7 onward, is deduced from the ι-crystallin mRNA of L. picturatus. It corrects the previously reported incomplete sequence (6) at positions 64–67, 69, and 79. The second sequence is the partial sequence of G. vittatus ι-crystallin, as determined by Edman degradation. Residues that are identical to the topmost sequence are indicated by dots. Positions of β-strands A–J and two α-helices in the CRBP structure (12) are indicated. Accession numbers for CRBP I are G809309 for rat, Q00915 for mouse, AAB34275 for cow, and P09455 for human; accession numbers for CRBP II are A92065 for rat, Q08652 for mouse, P50121 for pig, and AAC50162 for human.
Cloning of Gecko CRBP.
Isolation of RNA from two L. picturatus livers and cDNA synthesis were performed as described above. A first amplification was performed with a primer based on nucleotide sequences of human, rat, and mouse CRBP I and ι-crystallin (CTGAAGCCRGACAARGAGATC, corresponding to positions 36–42 in Fig. 1) and an anchor primer from the rapid amplification of cDNA ends kit. This PCR product was the template for another round of amplification with the primer ACCAYATGAYCATCCGCAC (corresponding to positions 47–53 in Fig. 1) and the anchor primer. The resulting cDNA was cloned and sequenced as described above.
Protein Sequence of Gonatodes ι-Crystallin.
Water soluble proteins from G. vittatus eye lenses were separated by SDS/PAGE, blotted onto poly(vinylidene difluoride) membrane (Immobilon-P, Millipore), stained with Coomassie blue, and partially destained. The band corresponding to ι-crystallin, migrating at 16 kDa, was excised and further destained. Amino acid sequencing was performed on a Hewlett–Packard G1000A protein sequencer.
Modeling.
A structural model for L. picturatus ι-crystallin was made by homology modeling (whatif; ref. 11) with rat CRBP I (PDB entry 1CRB; ref. 12) as the template. Rotamers of conserved residues were left unchanged. All other residues were mutated initially to alanines, and rotamers then were modeled with the whatif backbone-dependent rotamer libraries (13). The resulting model was energy minimized by using the steepest descents algorithm in the gromos87 programs (14). Calculations were performed in vacuo with crystallographic waters by using the gromos reduced charges force field. All calculations included positional restraints on the Cα atoms to the template coordinates (force constant = 9,000 kJ⋅mol−1⋅nm−2), and 3,4-didehydroretinol was included in the calculations with a modified gromos87 retinol topology. After energy minimization, 5 ps of molecular dynamics at 300 K was performed to relax the structure further, followed by conjugate gradient energy minimization until no significant energy difference could be observed. Analysis of electrostatic surface potential was performed by using grasp (15), and other pictures were made with bobscript (16) and raster3d (17).
Expression and Purification of Recombinant ι-Crystallin.
The QuikChange Site-Directed Mutagenesis Kit (Stratagene) was used to introduce an NdeI site (containing the initiation codon ATG of ι-crystallin) in the vector ι/pGEM-T. This vector, which already contains an NdeI site downstream of the ι-crystallin sequence, was digested with NdeI. The 665-bp fragment containing the ι-crystallin cDNA was isolated from a 1% agarose gel with the QiaEx II Gel Extraction Kit (Qiagen, Chatsworth, CA) and cloned into a pET3a vector for transfection into Escherichia coli BL21(DE3). Protein expression was induced by isopropyl β-d-thiogalactoside. Water-soluble lysate was produced (18) in phosphate buffer (4 mM Na2HPO4/4 mM KH2PO4/14 mM KCl/0.1 mM EDTA/20 mg/liter NaN3/3 mg/liter DTT/500 mg/liter PMSF, pH 6.8), and 25-ml fractions were applied to a DEAE-Sepharose Fast Flow column (50 ml; Amersham Pharmacia). ι-Crystallin was eluted with a second phosphate buffer (6.6 mM Na2HPO4/6.6 mM KH2PO4/23.3 mM KCl/0.166 mM EDTA/33.3 mg/liter NaN3/5 mg/liter DTT, pH 6.8), whereas almost all E. coli proteins remained bound. Suitable fractions were concentrated (Nova 3K disc membrane; Filtron, Karlstein, Germany) and purified further over Superdex-75 PG 16/60 (Amersham Pharmacia). Peak fractions of ι-crystallin were stored at 4°C for further use. Purity was assessed by SDS/PAGE, and identification was performed by Western blotting with a monoclonal antibody against bovine CRBP I (8).
Fluorescence Assays for Retinoid Binding.
The precise concentration of active ι-crystallin was determined by fluorescence titration with all-trans-retinol as described (19), except that retinol instead of protein fluorescence was measured. Stoichiometric binding of ligand was ensured by using protein concentrations far above the dissociation constant for retinol⋅CRBP I (19, 20). Fluorescence was measured at 458 nm (bandpass 20 nm) with excitation at 348 nm (bandpass 3 nm) in the absence of light on a Shimadzu RF-5301PC spectrofluorophotometer. Stock retinoid solutions in ethanol were prepared in the dark, and concentrations were determined on a Perkin–Elmer lambda-15 spectrometer with the absorption coefficients: all-trans-retinol, ɛ = 46,000 M−1⋅cm−1 at 325 nm (21); all-trans-retinoic acid, ɛ = 45,000 M−1⋅cm−1 at 350 nm (21); and all-trans-3,4-didehydroretinol, ɛ = 41,300 M−1⋅cm−1 at 350 nm (22). For direct binding assays, increasing amounts of all-trans-retinol were added to 1.5 ml of a 20 nM solution of ι-crystallin in phosphate buffer. Fluorescence was measured twice on two samples for each data point. The increase in retinol fluorescence was plotted as a function of retinol concentration, and the dissociation constant of the retinol⋅ι-crystallin complex was calculated by nonlinear least-squares fitting (19). For competition binding assays, increasing amounts of either all-trans-3,4-didehydroretinol or all-trans-retinoic acid were added to 1.5 ml of a 250 nM solution of ι-crystallin and a saturating amount (350 nM) of all-trans-retinol. This ι-crystallin concentration is well above the value of the dissociation constant of the retinol⋅ι-crystallin complex. Fluorescence was measured twice on a single sample for each data point. All samples were allowed to equilibrate for at least 15 min at room temperature in the dark before measurement. The final amount of ethanol in all samples never exceeded 1%.
Results
Sequence Characteristics of L. picturatus ι-Crystallin.
During the initial characterization of ι-crystallin, as isolated from the eye lenses of L. picturatus, the N-terminal sequence up to position 83 was determined by automated Edman degradation (6). To deduce the complete amino acid sequence, we performed reverse transcription–PCR on total RNA from L. picturatus lenses with a primer designed on the basis of the very N-terminal protein sequence of ι-crystallin. This procedure provided the sequence of the coding region, from nucleotide position 21 onward, and the 149 nucleotides of the 3′ untranslated region of ι-crystallin cDNA. The deduced amino acid sequence is 134 residues in length, giving a calculated molecular mass of 15,788 Da. Alignment with all available sequences of CRBP types I and II, which are known only for mammals, confirms that the protein is most closely related with CRBP I, with 59–60% sequence identity (Fig. 1). As compared with rat CRBP I, the number of charged residues is reduced in ι-crystallin: Asp + Glu from 25 to 21 and Lys + Arg from 19 to 17. An extra proline in ι-crystallin, next to the N-terminal proline, which is also present in CRBP I, is of interest. According to the “N-end rule” (23), an N-terminal proline may increase the average lifetime of a protein by protecting against degradation. A second proline might enhance this effect by providing a secondary barrier. There also is a trend to replace smaller with larger aliphatic residues: Ala + Val from 14 to 9 and Leu + Ile from 18 to 25. Proteins from thermophilic bacteria indicate a correlation between thermostability and a so-called aliphaticity index, calculated from the relative volume occupied by aliphatic residues (24). ι-Crystallin has a higher aliphaticity index (83.58) than that of rat CRBP I (77.01), which suggests a somewhat increased stability.
ι-Crystallin Is the Gecko CRBP I.
If ι-crystallin were the gecko housekeeping CRBP I, up-regulated in the lens, one would expect that CRBP I-like proteins in other tissues are identical to ι-crystallin. On the other hand, if ι-crystallin were the product of a duplicated CRBP I gene, the genuine housekeeping CRBP I should be found in other organs, possibly together with ι-crystallin. Reverse transcription–PCR was therefore performed on total RNA from liver, an organ with high CRBP I levels (9, 10). Two CRBP I-specific primers, designed on the basis of conserved regions in mammalian CRBP I and ι-crystallin cDNA sequences, were used in consecutive PCRs. These PCRs yielded a single cDNA sequence, identical to the last 266 nucleotides of the coding sequence and the entire 3′ untranslated region of the ι-crystallin mRNA as isolated from the lens. The fact that the only CRBP I-like gene product detected in gecko liver is identical with ι-crystallin indicates that it is indeed the housekeeping CRBP I that is up-regulated as a crystallin in the lens. ι-Crystallin/CRBP I is thus a convincing example of “gene sharing,” a phenomenon in which a protein is recruited to serve an additional function without loosing its original one (3, 25, 26).
Evolution of ι-Crystallin.
Western blotting has identified the presence of ι-crystallin/CRBP I-like proteins in lens extracts from diurnal geckos belonging to two other Old World genera, Quedenfeldtia and Pristurus, and to the New World genus Gonatodes (8). The CRBP I-like lens protein of G. vittatus deviates from that of the Old World genera by having a slightly larger apparent molecular mass and by having both 11-cis and all-trans 3,4-didehydroretinol as natural ligands (8). In lenses of the Old World geckos, only the all-trans form is found. Because of these differences, we performed automatic Edman degradation on Gonatodes ι-crystallin, isolated by SDS/PAGE and blotting onto poly(vinylidene difluoride) membrane. This procedure identified 58 residues in the N-terminal sequence, of which 51 are identical to Lygodactylus ι-crystallin (Fig. 1). Over this sequence, the Gonatodes and Lygodactylus ι-crystallins are approximately equidistant to mammalian CRBP I (66% and 64% sequence identity, respectively). This finding indicates that the G. vittatus protein is the orthologue of L. picturatus ι-crystallin.
The data set of Fig. 1 was used to obtain an impression of the relationship of ι-crystallin to mammalian CRBP I and II. Fig. 2 presents the tree that was obtained by neighbor-joining analysis (27) of the protein alignment. As expected, ι-crystallins are closest to CRBP I. Interestingly, the branch lengths indicate a considerably faster rate of change at the protein level in the gecko ι-crystallin/CRBP I lineage, because it diverged from the lineage toward mammalian CRBP I. This faster rate of change may in part reflect the accumulation of selectively advantageous amino acid changes enabling the additional function of CRBP I as a lens protein in geckos.
Figure 2.

ι-Crystallins in the phylogenetic tree of the CRBPs. This neighbor-joining tree (27) is constructed from the aligned protein sequences in Fig. 1 by using a Tajima-Nei distance matrix (28) and midpoint rooting. Unassigned positions in Gonatodes ι-crystallin are introduced as missing data. Bootstrap scores are based on 1,000 replicates. Branch lengths are proportional to the number of amino acid replacements.
Structural Model of ι-Crystallin.
One wonders whether such adaptive changes can be observed when the ι-crystallin sequence is modeled by using rat CRBP I as the template structure. CRBP I has, like the other iLBPs, a β-barrel structure made up of 10 antiparallel strands, with the ligand localized inside the barrel (10, 29). In agreement with the amino acid composition of ι-crystallin, analysis of electrostatic surface potential shows a reduction of—mostly negative—surface charge, especially at the front (strands A–F; Fig. 3). Reduction of negative surface charge reduces repulsive behavior, thereby allowing a protein to achieve high concentrations, in line with the close packing of proteins needed in the eye lens to obtain a high refractive index (30). The mutations in ι-crystallin are concentrated in a few areas (Fig. 4A). In strands F–J, forming the β-sheet behind the retinoid, strings of residues running perpendicular to the strand directions have been mutated. All these residues point toward the solvent. The residues lining the pocket of the retinoid tail, i.e., pointing inward on the two β-sheets, have been completely conserved. However, there is a cluster of mutations around the retinoid ring, on the tips of the C–D and E–F turns, and on the inside of the helices. These areas are compared for ι-crystallin and rat CRBP I in Fig. 4 B and C. The residue that is in closest contact with the retinoid is at position 33 (3.7 Å for both ι-crystallin and CRBP I). This residue is an alanine in mammalian CRBP I and II but a valine in ι-crystallin, both in Lygodactylus and Gonatodes. Further mutations around the retinoid ring are Phe-16 → Met and Phe-57 → Leu. These three mutations could represent adaptive modifications to optimize the fit for 3,4-didehydroretinol in ι-crystallin. Alternatively, they may equally well be selectively neutral changes.
Figure 3.
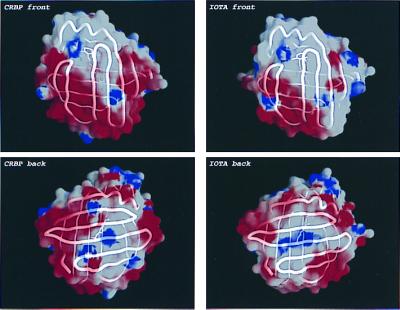
Reduced surface charge of ι-crystallin (Right) as compared with rat CRBP I (Left). (Upper) Front view. (Lower) Back view. Molecular surfaces, calculated in grasp (15), are colored according to electrostatic potential: red < −5.0 kT and blue > 5 kT. The Cα backbone worm is shown in white, and retinoids are in a stick representation (3,4-didehydroretinol for ι-crystallin and retinol for CRBP I).
Figure 4.

Mutational hotspots in ι-crystallin. (A) Ribbon model of the ι-crystallin β-barrel, with positions of amino acid replacements (compared with rat CRBP I) in red and with 3,4-didehydroretinol in green stick representation. Orientation as in Fig. 3 Upper. (B and C) Comparison of the retinoid binding pockets of ι-crystallin and rat CRBP I, respectively. Amino acid side chains are drawn as sticks, colored by atom type. The retinoid is shown in purple sticks.
Ligand-Binding Properties of Recombinant ι-Crystallin.
To assess the actual retinoid-binding properties of ι-crystallin, the recombinant L. picturatus protein was expressed in E. coli and purified. A solution containing a precisely determined concentration of active ι-crystallin was used in a direct binding assay with all-trans-retinol. Retinol fluorescence was measured and plotted as a function of total retinol concentration (Fig. 5A). Nonlinear least-squares fitting of the data gave Kd = 1.606 nM and n = 1.043 as best estimates (R2 = 0.997) for the apparent dissociation constant of the all-trans-retinol⋅ι-crystallin complex and the correction factor for the ι-crystallin concentration, respectively. This dissociation constant is similar to values published for bovine CRBP I (19), although higher values have also been reported (20). The ability of all-trans-3,4-didehydroretinol to compete with all-trans-retinol for binding to ι-crystallin is shown in Fig. 5B. The binding curve of all-trans-3,4-didehydroretinol drops to half the fluorescence intensity [i.e., 1/2(F0 + F∞)] at a molar ratio to all-trans-retinol equal to one. This value indicates that ι-crystallin has similar and high affinity for both of these retinoids. Fig. 5B also shows that all-trans-retinoic acid, another physiologically relevant retinoid, is unable to compete with all-trans-retinol. These observations are comparable to those reported for rat CRBP I, which also has similar affinities for all-trans-retinol and all-trans-3,4-didehydroretinol, and where all-trans-retinoic acid is unable to compete for binding (20).
Figure 5.

Ligand binding of recombinant ι-crystallin. (A) Determination of the dissociation constant of ι-crystallin for all-trans-retinol. Open circles indicate the fraction of liganded ι-crystallin at increasing total retinol concentrations, as calculated from measured retinol fluorescence. The continuous line is the nonlinear least-squares fitting of these data, giving a Kd of 1.606 nM. (B) Competition binding assays, presenting remaining retinol fluorescence after competitive binding of all-trans-retinoic acid (closed circles) and all-trans-3,4-didehydroretinol (open circles).
Discussion
Our results suggest the following scenario for the evolutionary origin of gecko lens ι-crystallin. The mostly nocturnal gecko family is supposed to have descended from a diurnal lizard ancestor (31). Several genera of geckos have reverted to diurnal habits, often living in environments where they are exposed to intense ambient light. The eyelids of these geckos are fused to form a transparent spectacle, and they are unable to regulate incident light by changing the pupil diameter. Consequently, the only means to avoid potentially harmful radiation, especially UV and short-wave blue light, that reaches the retina is to absorb it in the lens. This absorption can be accomplished by the accumulation of suitable lens pigments. Diurnal geckos of the genera Lygodactylus, Gonatodes, Quedenfeldtia, and Pristurus use 3,4-didehydroretinol, bound to ι-crystallin/CRBP I, for this purpose (6, 8). This situation required complex evolutionary processes that had to occur in concert to have any adaptive value. Although not normally occurring in the lens, one can imagine that CRBP I became expressed in the gecko lens, initially at low levels, by chance mutations in the promoter of its gene. Interestingly, another iLBP, related to epidermal fatty acid binding protein, has been found at low levels in the bovine lens (32). It may be relevant in this respect that iLBP genes have the potential to become highly expressed, given the presence of 18% muscle fatty acid-binding protein in adult locust flight muscle (33). Interesting, too, is the discovery that O-crystallin from the octopus eye lens is related to the phosphatidylethanolamine-binding proteins, another family of lipid-binding proteins (34).
A gradually increasing expression of CRBP I in the lens would probably be selectively neutral, because CRBP I, like other iLBPs, is a very stable protein (29); as such, it is acceptable as a lens protein. However, retinol, the natural ligand of CRBP I, is unsuitable as a lens chromophore, because it is not photostable and fluoresces in the range of visible light (35). In contrast, 3,4-didehydroretinol is photostable and does not fluoresce but is rarely found in terrestrial vertebrates. Vitamin A2-based visual pigments do in fact occur in some lizards (36), but both nocturnal and diurnal geckos use retinol for visual pigment generation (6). However, in the eye cups of diurnal Lygodactylus geckos, but not in those of nocturnal geckos, 3,4-didehydroretinol and its ester are found (8). It therefore seems that the synthesis of a suitable lens chromophore as a ligand for ι-crystallin required the activation of an enzyme that converts retinol into 3,4-didehydroretinol specifically for this purpose. Because this conversion apparently occurs in the retinal pigment epithelium (8), the chromophore must be transported into the lens—by ways that are as yet unknown—before it can be liganded to ι-crystallin. Considering the complexity of this suite of events, it seems most likely that it occurred only once in some ancestral gecko lineage. This conclusion does not necessarily imply that the presence of ι-crystallin in four gecko genera indicates their monophyly; the expression of ι-crystallin may have faded again in certain daughter lineages, as has been proposed for other taxon-specific crystallins (1, 2).
Acquiring the additional function as a lens protein, while maintaining its role as retinol transporter in other tissues, may have imposed specific demands on gecko ι-crystallin/CRBP I. In the lens, ι-crystallin/CRBP I should bind its chromophore without the need to release it again, and an increased stability would be favorable for a long-living lens protein. To function as a retinol transporter, ι-crystallin/CRBP I should be able to bind retinol and release it again as needed (37), thus requiring a certain degree of structural flexibility (38). Like mammalian CRBP I (20), ι-crystallin binds all-trans-3,4-didehydroretinol equally as well as all-trans-retinol (Fig. 5B). Because 3,4-didehydroretinol is the only retinoid present in the gecko eye lens (8), there is no real requirement for specific adaptations of ι-crystallin to obtain preferential binding of this chromophore. Indeed, although our modeling studies identified replacements in the binding pocket of ι-crystallin around the retinoid ring, suggesting that it is optimized for all-trans 3,4-didehydroretinol, we could not demonstrate any higher affinity in our retinoid-binding assays. Furthermore, any optimization in this direction would almost certainly affect the retinol transporter functions of ι-crystallin in nonlens tissues. Ruling out this option, therefore, it seems that ι-crystallin/CRBP I has sustained the following adaptive changes to make it a better lens protein. First, trends in amino acid substitutions and the proline at position 2 lead to increased stability and lowered turnover. Second, diminished negative surface charge reduces repulsive interactions, thereby facilitating close packing of proteins, in line with the need for a high refractive index of the eye lens (30). The apparently increased rate of change in the ι-crystallin lineage after its divergence from the lineage to mammalian CRBP I may in part reflect these adaptive trends.
During evolution, various other chromophores have been recruited in vertebrate lenses to reduce the deleterious effects of short-wave radiation. The yellowish lenses of human, squirrel, and some fishes contain kynurenine derivatives (39, 40), whereas mycosporine-like amino acids occur in the lenses of many marine fishes (40). Lipid-soluble carotenoids also serve this purpose and are bound to the lens protein α-crystallin in the hatchetfish Argyropelecus affinis (41) and to γs-crystallin in the gecko Quedenfeldtia trachyblepharus (42). The recruitment of CRBP I as a lens protein and the concomitant synthesis of a stable and nonfluorescent chromophore further indicate the versatility of organisms to cope with novel environmental demands and present a convincing example of adaptive evolution at the molecular level.
Acknowledgments
We thank Willem de Grip and Petra Geurts for help with the retinoid-binding experiments, Ole Madsen for phylogenetic tree constructions, and Hoffmann–La Roche for the gift of vitamin A2.
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
Data deposition: The sequences reported in this paper have been deposited in the GenBank database (accession nos. AJ249753 for Lygodactylus picturatus ι-crystallin and P82025 for Gonatodes vittatus ι-crystallin).
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.050500597.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.050500597
References
- 1.de Jong W W, Lubsen N H, Kraft H J. Prog Retin Eye Res. 1994;13:391–442. [Google Scholar]
- 2.Wistow G. Molecular Biology and Evolution of Crystallins: Gene Recruitment and Multifunctional Proteins in the Eye Lens. Austin, TX: Landes; 1995. [Google Scholar]
- 3.Piatigorsky J, Wistow G. Cell. 1989;57:197–199. doi: 10.1016/0092-8674(89)90956-2. [DOI] [PubMed] [Google Scholar]
- 4.Jimenez-Asensio J, Gonzalez P, Zigler J S, Jr, Garland D L. Biochem Biophys Res Commun. 1995;209:796–802. doi: 10.1006/bbrc.1995.1570. [DOI] [PubMed] [Google Scholar]
- 5.Röll B, Van Boekel M A, Amons R, de Jong W W. Biochem Biophys Res Commun. 1995;217:452–458. doi: 10.1006/bbrc.1995.2797. [DOI] [PubMed] [Google Scholar]
- 6.Röll B, Amons R, de Jong W W. J Biol Chem. 1996;271:10437–10440. doi: 10.1074/jbc.271.18.10437. [DOI] [PubMed] [Google Scholar]
- 7.Zigler J S, Jr, Rao P V. FASEB J. 1991;5:223–225. doi: 10.1096/fasebj.5.2.2004667. [DOI] [PubMed] [Google Scholar]
- 8.Röll B, Schwemer J. J Comp Physiol A. 1999;185:51–58. [Google Scholar]
- 9.Saari J C. Prog Retin Eye Res. 1990;9:363–381. [Google Scholar]
- 10.Newcomer M E. FASEB J. 1995;9:229–239. doi: 10.1096/fasebj.9.2.7781925. [DOI] [PubMed] [Google Scholar]
- 11.Vriend G. J Mol Graphics. 1990;8:52–56. doi: 10.1016/0263-7855(90)80070-v. [DOI] [PubMed] [Google Scholar]
- 12.Cowan C W, Newcomer M E, Jones T A. J Mol Biol. 1993;230:442–446. doi: 10.1006/jmbi.1993.1238. [DOI] [PubMed] [Google Scholar]
- 13.Vriend G, Sander C. J Appl Crystallogr. 1993;26:47–60. [Google Scholar]
- 14.van Gunsteren W F, Berendsen H C J. gromos, BIOMOS, Biomolecular Software. Univ. of Groningen, Groningen, The Netherlands: Laboratory of Physical Chemistry; 1987. [Google Scholar]
- 15.Nicholls A, Sharp K, Honig B. Proteins. 1991;11:281–296. doi: 10.1002/prot.340110407. [DOI] [PubMed] [Google Scholar]
- 16.Esnouf R M. J Mol Graphics. 1997;15:132–134. doi: 10.1016/S1093-3263(97)00021-1. [DOI] [PubMed] [Google Scholar]
- 17.Bacon D, Anderson W F. J Mol Graphics. 1988;6:219–220. [Google Scholar]
- 18.Werten P J L, Carver J A, Jaenicke R, de Jong W W. Protein Eng. 1996;9:1021–1028. doi: 10.1093/protein/9.11.1021. [DOI] [PubMed] [Google Scholar]
- 19.Malpeli G, Stoppini M, Zapponi M C, Folli C, Berni R. Eur J Biochem. 1995;229:486–493. [PubMed] [Google Scholar]
- 20.MacDonald P N, Ong D E. J Biol Chem. 1987;262:10550–10556. [PubMed] [Google Scholar]
- 21.Horwitz J, Heller J. J Biol Chem. 1973;248:6317–6324. [PubMed] [Google Scholar]
- 22.von Planta C, Schweitzer U, Chopard-dit-Jean L, Ruegg R, Kofler M, Isler O. Helv Chim Acta. 1962;45:548–561. [Google Scholar]
- 23.Gonda D K, Bachmair A, Wunning I, Tobias J W, Lane W S, Vashavsky A. J Biol Chem. 1989;264:16700–16712. [PubMed] [Google Scholar]
- 24.Ikai A. J Biochem. 1980;88:1895–1898. [PubMed] [Google Scholar]
- 25.Piatigorsky J, O'Brien W E, Norman B L, Kalumuck K, Wistow G J, Borras T, Nickerson J M, Wawrousek E F. Proc Natl Acad Sci USA. 1988;85:3479–3483. doi: 10.1073/pnas.85.10.3479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hendriks W, Mulders J W, Bibby M A, Slingsby C, Bloemendal H, de Jong W W. Proc Natl Acad Sci USA. 1988;85:7114–7118. doi: 10.1073/pnas.85.19.7114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Saitou N, Nei M. Mol Biol Evol. 1987;4:406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 28.Tajima F, Nei M. Mol Biol Evol. 1984;1:269–285. doi: 10.1093/oxfordjournals.molbev.a040317. [DOI] [PubMed] [Google Scholar]
- 29.Banaszak L, Winter N, Xu Z, Bernlohr D A, Cowan S, Jones T A. Adv Protein Chem. 1994;45:89–151. doi: 10.1016/s0065-3233(08)60639-7. [DOI] [PubMed] [Google Scholar]
- 30.Tardieu A, Vérétout F, Krop B, Slingsby C. Eur Biophys J. 1992;21:1–12. doi: 10.1007/BF00195438. [DOI] [PubMed] [Google Scholar]
- 31.Walls G L. The Vertebrate Eye and Its Adaptive Radiation. Bloomfield Hills, MI: Cranbrook Inst. Sci.; 1942. [Google Scholar]
- 32.Jaworski C J, Wistow G J. Invest Ophthalmol Visual Sci. 1994;35:1706. [Google Scholar]
- 33.Haunerland N H, Chen X, Andolfatto P, Chisholm J M, Wang Z. Mol Cell Biochem. 1993;123:153–158. doi: 10.1007/BF01076487. [DOI] [PubMed] [Google Scholar]
- 34.Zinovieva R D, Piatigorsky J, Tomarev S I. Biochim Biophys Acta. 1999;1431:512–517. doi: 10.1016/s0167-4838(99)00066-7. [DOI] [PubMed] [Google Scholar]
- 35.Ong D E, Chytil F. J Biol Chem. 1978;253:828–832. [PubMed] [Google Scholar]
- 36.Provencio I, Loew E R, Foster R G. Vision Res. 1992;32:2201–2208. doi: 10.1016/0042-6989(92)90084-v. [DOI] [PubMed] [Google Scholar]
- 37.van Aalten D M F, Findlay J B C, Amadei A, Berendsen H J C. Protein Eng. 1995;8:1129–1135. doi: 10.1093/protein/8.11.1129. [DOI] [PubMed] [Google Scholar]
- 38.Jamison R S, Newcomer M E, Ong D E. Biochemistry. 1994;33:2873–2879. doi: 10.1021/bi00176a017. [DOI] [PubMed] [Google Scholar]
- 39.Zigman S, Paxhia T. Exp Eye Res. 1988;47:819–824. doi: 10.1016/0014-4835(88)90065-6. [DOI] [PubMed] [Google Scholar]
- 40.Thorpe A, Douglas R H, Truscott R J. Vision Res. 1993;33:289–300. doi: 10.1016/0042-6989(93)90085-b. [DOI] [PubMed] [Google Scholar]
- 41.Yu N T, Cai M Z, Lee B S, Kuck J F, Jr, McFall-Ngai M, Horwitz J. Exp Eye Res. 1991;52:475–479. doi: 10.1016/0014-4835(91)90045-g. [DOI] [PubMed] [Google Scholar]
- 42.Röll, B. (2000) Comp. Biochem. Physiol., in press.