

Abstract
The last European fossil occurrence of a coelacanth is from the Mid-Cretaceous of the English Chalk (Turonian, 90 million years ago). Here, we report the discovery of a coelacanth from Late Cretaceous non-marine rocks in southern France. It consists of a left angular bone showing structures that imply close phylogenetic affinities with some extinct Mawsoniidae. The closest relatives are otherwise known from Cretaceous continental deposits of southern continents and suggest that the dispersal of freshwater organisms from Africa to Europe occurred in the Late Cretaceous.
Keywords: Actinistia, fossil, Cretaceous, palaeobiogeography, dispersal
1. Introduction
Since Agassiz (1839) described a tail of Coelacanthus from the Permian of England, coelacanth fishes have long been known as fossils. They were thought to have become extinct some 75 million years ago. The living coelacanth, Latimeria, from the western Indian Ocean, was described by Smith (1939), as recently was another from the Celebes Sea (Erdmann et al. 1998). Marine representatives of the Latimeria lineage are known from the Mid-Cretaceous (Albian–Turonian) chalk of England and Czech Republic with Macropoma (Forey 1998), and from the Late Cretaceous of North America with Megalocoelacanthus (Schwimmer et al. 1994). Alongside the Latimeria lineage, freshwater mawsoniid coelacanths occurred in the Late Cretaceous of Africa and Madagascar as well as in the Early Cretaceous brackish/freshwater deposits of South America. Here, we provide the first direct evidence, to our knowledge, of a continental coelacanth in the European Late Cretaceous in the form of an angular referable to a mawsoniid coelacanth. Geologically, the new find from France is about 20 million years younger than Macropoma. The specimen shows affinities with the Mawsonia–Axelrodichthys complex and indicates brackish or, more likely, freshwater dispersals between Africa and the European archipelago during the Late Cretaceous.
2. Fossil material
The fossil coelacanth described here was found in 2004 during excavations at the Early Maastrichtian locality of Cruzy (southern France). The vertebrate remains were found in a lens of variegated clays cut by unfossiliferous sandy channels. The sediments of the Cruzy locality were deposited in continental environments, probably a floodplain. The fossil vertebrate assemblage comprises isolated remains of gars, lissamphibians, squamates, turtles, crocodiles, pterosaurs, ornithopod, sauropod and theropod dinosaurs, enantiornithine birds and mammals (Buffetaut et al. 1999).
3. Results
Our identification of the specimen as a coelacanth angular is based on the rectangular outline shape, the pattern of the mandibular sensory canal and the openings from that canal (figure 1a, m.s.c.), the path of the groove for the external mandibular ramus of the facial nerve (figure 1a, gr.VII.m.ext) and the presence of a ridge on the medial side which marks the floor of the adductor fossa (figure 1a, r). The specimen is referred with confidence to the Mawsonia–Axelrodichthys complex because of an ornamentation consisting of coarse ridges radiating from a point in the posterior third of the bone, an inflated lateral surface, the few slit-like ventral openings of the sensory canal (figure 1b, arrows) and the well-marked medial contact surface with the prearticular (figure 1a, con.Part). Coelacanths other than Mawsonia, Axelrodichthys, Lualubaea and Megalocoelacanthus show an ornament upon the angular that consists of enamel-topped tubercles or ridges, or is absent altogether; a mandibular sensory canal that opens through rounded pores; an ill-defined area of contact between the angular and prearticular, and a centre of ossification on the angular that is close to the deepest part of the bone (Forey 1998). Among these coelacanths our specimen may be distinguished from Megalocoelacanthus because that taxon shows an angular with a posteriorly directed opening for a special subopercular branch of the sensory canal, which indicates affinities with the latimeriids (Forey 1998). We are less certain about its distinction from other coelacanths within the Mawsonia–Axelrodichthys complex: for this we would need more material.
Figure 1.
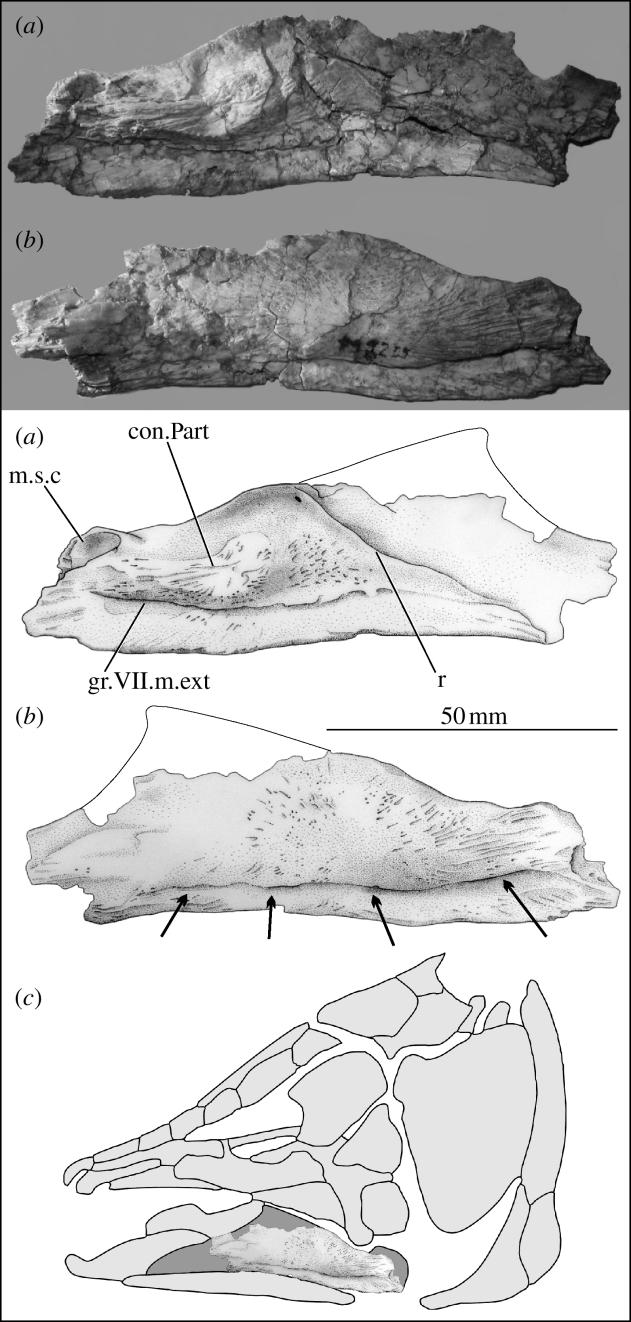
Left angular of a mawsoniid coelacanth from the Early Maastrichtian locality of Cruzy, southern France (M2205, collection Association culturelle, archéologique et paléontologique de l'Ouest biterrois). (a) Internal views, (b) lateral views and (c) location of the bone within the outline of a mawsoniid skull.
4. Discussion
Late Cretaceous coelacanths are represented only by the marine latimeriids and the fresh water mawsoniids. Mawsoniid coelacanths diverged from the latimeriids in the Early Mesozoic. Mawsoniid coelacanths were exclusively Mesozoic and most lived in fresh water. Typical members, such as Mawsonia and Axelrodichthys, were heavy-bodied and showed an unusual pattern of fusion between bones in the skull roof. Some grew to a large size, estimated at over 2 m, and rivalled the size of Latimeria today (Wenz 1981).
Late Cretaceous occurrences of Africa and Madagascar (Gottfried et al. 2004) are exclusively from fresh water. Both Mawsonia and Axelrodichthys are known in the Early and Middle Cretaceous of both sides of the proto South Atlantic (Cavin & Forey 2004) showing a vicariant biogeographic pattern (Maisey 2000). The exclusive occurrences of mawsoniids in fresh waters of Africa (Cavin & Forey 2004) and Madagascar (Gottfried et al. 2004) during the Late Cretaceous strongly suggest that mawsoniids reached the European archipelago by dispersal from Africa. We cannot exclude the possibility that dispersal polarity was from Europe to Africa. However, in the absence of contradictory information, we accept that the earlier and more widespread occurrence on and around the southern continents suggests that this is the more likely starting point. Although reconstructed maps show a wide marine barrier between Africa and the European archipelago in the Late Cretaceous (Hay et al. 1999; Scotese 2003), dispersal events have already been detected for tetrapods (Buffetaut 1989; Le Loeuff 1991) and freshwater fishes (Cavin et al. 1996), probably via a discontinuous route (Rage 1997).
Acknowledgments
We thank all those who took part in the excavations and in the preparation of specimens, in particular the members of the Association culturelle, archéologique et paléontologique de l'Ouest biterrois (ACAP). The excavation works were supported by the ECLIPSE Programme of the Centre National de la Recherche Scientifique (CNRS) and the ACAP. L.C.’s research was supported by a Marie Curie Individual Fellowship funded by the Swiss Federal Office for Education and Science (grant no. 02.0335).
References
- Agassiz L. Recherches sur les poissons fossiles. vol. 2. Published by the author, Petitpierre; Neuchâtel: 1839. [Google Scholar]
- Buffetaut E. Archosaurian reptiles with Gondwanan affinities in the Upper Cretaceous of Europe. Terra Nova. 1989;1:69–74. [Google Scholar]
- Buffetaut E, Le Loeuff J, Tong H, Duffaud S, Cavin L, Garcia G, Ward D, L'association Culturelle, Archéologique Et Paléontologique De Cruzy Un nouveau gisement de vertébrés du Crétacé supérieur à Cruzy (Hérault, Sud de la France) C. R. Acad. Sci. Paris, Sciences de la terre et des planètes. 1999;328:203–208. [Google Scholar]
- Cavin L, Forey P.L. New mawsoniid coelacanth (Sarcopterygii: Actinistia) remains from the Cretaceous of the Kem Kem beds, SE Morocco. In: Tintori A, Arratia G, editors. Mesozoic fishes III. Dr Pfeil Verlag; Munich: 2004. pp. 493–506. [Google Scholar]
- Cavin L, Martin M, Valentin X. Découverte d'Atractosteus africanus (Actinopterygii: Lepisosteidae) dans le Campanien inférieur de Ventabren (Bouches-du-Rhône, France). Implications paléobiogéographiques. Revue de Paléobiologie. 1996;15:1–7. [Google Scholar]
- Erdmann M.V, Caldwell R.L, Kasim Moosa M.K. Indonesian ‘king of the sea’ discovered. Nature. 1998;395:335. [Google Scholar]
- Forey P.L. Chapman & Hall; London: 1998. History of the coelacanth fishes. [Google Scholar]
- Gottfried M.D, Rogers R.R, Curry Rogers K. First record of Late Cretaceous coelacanths from Madagascar. In: Arratia G, Wilson M.V.H, Cloutier R, editors. Recent advances in the origin and early radiation of vertebrates. Dr Pfeil Verlag; Munich: 2004. pp. 687–691. [Google Scholar]
- Hay W.W, et al. An alternative global Cretaceous paleogeography. In: Barrera E, Johnson C.C, editors. Cretaceous ocean/climate systems. Geological Society of America; Boulder, Co: 1999. pp. 1–48. [Google Scholar]
- Le Loeuff J. The Campano–Maastrichtian vertebrate faunas from southern Europe and their relationships with other faunas in the world; palaeobiogeographical implications. Cretaceous Res. 1991;12:93–114. [Google Scholar]
- Maisey J.G. Continental break up and the distribution of fishes of Western Gondwana during the Early Cretaceous. Cretaceous Res. 2000;2000:281–314. [Google Scholar]
- Rage, J. C. 1997. Terrestrial trans-Tethyan dispersals: an overview. Geological Society of Danmark, On Line Series, 1: http://www.2dgf.dk/publikationer/DGF_On_Line/Vol_1/tethys.htm
- Scotese, C. R. 2003 Paleomap project. See www.scotese.com
- Schwimmer D.R, Stewart J.D, Williams G.D. Giant fossil coelacanth of the Late Cretaceous in the eastern United States. Geology. 1994;22:503–506. [Google Scholar]
- Smith J.L.B. A living fish of Mesozoic type. Nature. 1939;143:455–456. [Google Scholar]
- Wenz S. Un coelacanthe géant, Mawsonia lavocati Tabaste, de l'Albien-base du Cénomanien du sud marocain. Annales de Paléontologie (Vertébrés) 1981;67:1–20. [Google Scholar]