

Abstract
Birds are known to be a group rich in pairs of closely related species that have parapatric or allopatric distributions with relatively narrow contact zones. Here we analyse the geographical distribution of these contact zones for parapatric species pairs of passerine birds of the Palaearctic region. Their contact zones are located mainly in southwestern, northern and central–southern Asia, and in northwestern Africa, with a hotspot in the Middle East. A mid-domain effect null model, where contact zone hotspots are a neutral correlate of continental geometry, had a low explanatory power of 3.8%; the observed distribution of contact zones was not sufficiently predicted. Hypotheses involving range contractions and secondary contact in areas of high topographic and habitat diversity may offer more convincing explanations and offer promising perspectives for future studies.
Keywords: Passeriformes, contact zone, mid-domain effect, parapatry, Palaearctic
1. Introduction
Evolutionary biologists have long recognized the importance of documenting contact zones of related species of organisms (Mayr 1963). The data derived from such studies contribute to the understanding of processes of speciation, dispersal and vicariance. Contact zones usually are defined as the meeting area of pairs of closely related species, implying a degree of congruence in their separate distribution (Short 1969). These areas have been most frequently reported in birds (Dixon 1989), representing major zones of biogeographic discontinuity among western and eastern taxa, and probably current or former barriers (Haffer 1997). A plethora of hypotheses have been suggested to explain the pattern and causes of contact zones, mainly concentrating on historical interpretations such as the refuge, river refuge and disturbance-vicariance hypotheses (Haffer 1977, 1997; Haffer & Prance 2001). Based on cursorial observations, Haffer (1977, 1992) and Vaurie (1954–1965) found that contact zones of parapatric passerine birds (order Passeriformes) appear to be more common than average in the Middle East. The refuge theory (Haffer 1977; Haffer & Prance 2001) explains this pattern by assuming that the species involved survived the preceding cold–arid climatic phase of the last glacial stage in moist refuges, and that the Middle East, with its high diversity of habitat and a fast clinal habitat variation along steep clines, provides ideal opportunities for secondary contact.
A high diversity of species in the central area of a continent, such as the Middle East, is also predicted by the mid-domain effect (MDE), as set out by Colwell & Lees (2000). MDE explains species gradients based solely on geometric constraints on species ranges, independent of all evolutionary and environmental features, leading to a peak in range overlap among species in the centre of a geographical area (Colwell & Hurtt 1994). Such increased range overlap could also influence the distribution of hybrid zones.
Here we used passerines, the most species-rich clade of birds with over 5700 species in 96 families (60% of the world avifauna), as a model group to explore the distribution of contact zones in the Palaearctic. Geographical areas of distributional overlap were calculated from newly compiled distribution maps of parapatric species pairs. Our goals were: (i) to test the hypothesis of increased occurrence of contact zones in the Middle East and (ii) to discover whether the distribution of these zones in the Palaearctic can be explained as a neutral correlate of continental geometry, or whether a historical or ecological explanation needs to be invoked.
2. Material and methods
The study area encompasses the whole of the Palaearctic Region (Europe, North Africa and Asia: 30°–170° W, 86°25′–18°00′ N; land area: 54.1 million km2). Parapatric species pairs were identified as species with contiguous or narrow overlap zones, excluding each other geographically and with no, or restricted, hybridization along their contact zones (Haffer 1992). Lists of parapatric species pairs of passerine birds in this area were available from the works of Haffer (1977, 1992). Records of hybridization among these species were compiled from Bures et al. (2002) and Randler (2002; Electronic Appendix, Appendix 1). Nomenclature and taxonomy follows Roselaar & Shirihai (in press). Separate analyses were carried out for species pairs that do and do not hybridize, respectively, in order to exclude possible artefacts originating from the definition of species pairs, but similar hotspot patterns were recovered in both cases (not shown).
A database of digitized distribution maps was created for the species studied with the help of the computer program Worldmap v. 4.1 (Williams 2000). The geographical distributions were interactively plotted on an equal area map of the Palaearctic (equivalent cylinder projection, equidistant on 55° parallel circle), overlaid by a one degree-wide grid (grid cell area: 4062 km2). For all parapatric passerine species pairs we compiled distribution maps based on various literature sources, supplemented by data obtained from numerous zoological collections (these maps will be published in forthcoming studies). We produced combined maps for the two taxa of each species pair and extracted the overlap of their distribution areas as a new map. The contact zones of all species pairs considered here were subsequently combined, and their geographical distribution analysed.
To examine the effect of the continental edge on the distribution of contact zones in the Palaearctic and Middle East, we used the ‘area corrected’ version of the two-dimensional simulation model (Bokma et al. 2001; Hawkins & Diniz 2002). Based on this stochastic model, the species richness at a point P is a function of its distance to the northern (p), southern (q), western (r) and eastern (t) continental boundaries. The species richness at P is then given by 4pqrtS, where S is the species pool (in this case equal to the 52 contact zones). These values in the null model were estimated from each point to the maximum north-south and east-west axes. Values were only calculated for those grid cells included in one or more hybrid zones in the real dataset; this approach is conservative because any MDE prediction of a contact zone in other grid cells would lower the fit of the model. Coefficients of determination (R2) of linear regression were used to evaluate the relationship between the observed contact zones' richness at each sample point and the prediction obtained by the geometric model.
3. Results
We identified 98 species (52 species pairs) of passerines that make contact in the Palaearctic region (Electronic Appendix, Appendix 1). A relatively large proportion of these species pairs (37%) hybridize in their contact zones. The combined land area of one or more contact zones covers 23% of the Palaearctic. Although contact zones occur over a large part of the Palaearctic, they are not evenly distributed in space (figure 1). The contact zone richness is highest along the mountain chains of the Caucasus, Taurus, Zagros and Alborz (southwest Asia); Atlas (northwest Africa); Hindu Kush, Pamir, Tienshan and Altai (north-central Asia); and Himalaya (south-central Asia). Six hotspots, covering 2% of the Palaearctic, harbour some 55% of all species pairs, whereas the two hottest grid cells, including 8 and 9 species pairs, cover only about 8000 km2 in the Middle East (Kopet Dag, northeast Iran; Electronic Appendix, Appendix 2). Analyses that included or excluded altitudinal or vertical parapatric species pairs (Haffer 1992) showed the same general pattern (results not shown).
Figure 1.
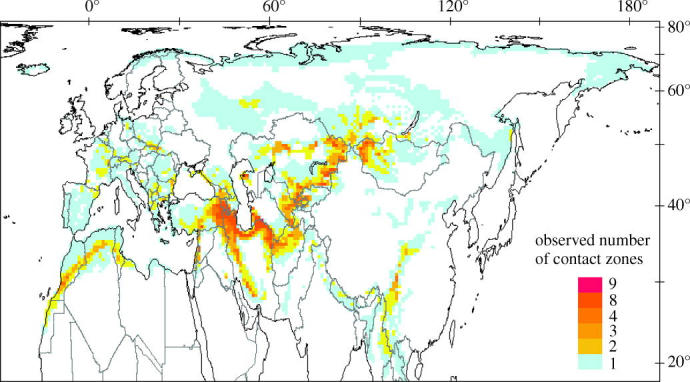
Hotspots of contact zones of species pairs of passerine birds in the Palaearctic region (for further explanation see text).
The mean size of the contact zones is 0.53×106 km2 (±0.08×106 km2, n=52 species pairs). The maximum (3.02×106 km2) and minimum (12.19×103 km2) range of overlap zones are formed by the Common redpoll, Carduelis flammea×the Hoary redpoll, Carduelis hornemanni in Siberia; and the White-throated tit, Aegithalos niveogularis×the Black-browed tit, A. iouschistos species pairs in Nepal, respectively.
The spatial pattern of the contact zones clearly varied from that predicted by geometric models (figure 2), and the coefficient of determination from a standard regression analysis is low (R2=0.038, p<0.0001). The predicted hotspots are in north-central Asia, including Mongolia, Kazakhstan, southern Russia and northern China.
Figure 2.
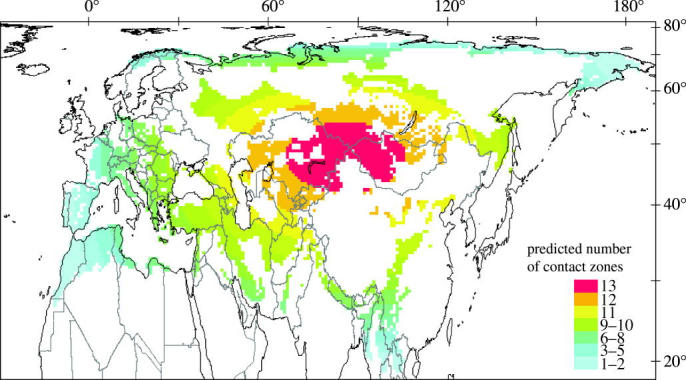
Prediction pattern of contact zones, using the area-corrected bi-dimensional model, for passerine birds in the Palaearctic region (for further explanation see text).
4. Discussion
Our analysis reveals a distinctly uneven geographical distribution of contact zones of passerine birds in the Palaearctic region, and confirms the hypothesis of a contact zone hotspot in the Middle East. The further hotspot areas in central–northern Asia are in apparent agreement with the prediction of the MDE (figure 2), but the geometric model fails to predict the high number of contact zones in southwestern Asia, southern Europe, northwestern Africa, and especially in the Middle East. Despite possessing a statistically significant fit to the real data, the MDE has low explanatory power and accounts for only 3.8% of the variance across all contact zones. This agrees with other bi-dimensional analyses, in which the coefficient of determination ranged from 5 to 26% (Bokma et al. 2001; Jetz & Rahbek 2001; Diniz et al. 2002; Hawkins & Diniz 2002; Rangel & Diniz 2003), whereas other topographical predictors in similar analyses of regional-scale diversity typically generate coefficients of determination greater than 70% (Boone & Krohn 2000; Balmford et al. 2001; Hawkins & Diniz 2002). Hence, we conclude that a random overlap of constrained geographical ranges cannot fully account for the observed pattern of passerine bird contact zones in the Palaearctic.
The refuge theory of Haffer (Haffer 1977; Haffer & Prance 2001) offers an alternative explanation, according to which the distribution history of passerines has been severely influenced by climatic and vegetational fluctuation during the Quaternary period. The successive expansions and contractions of species ranges and the rearrangement of species assemblages would have established secondary contact zones in northwest and southwest Asia in mixed deciduous and coniferous forests (Caucasus, Turkestan, Afghanistan), and juniper woodlands (Iran, Turkmenistan, Tajikistan, China). The question of why these contact zones are concentrated in the Middle East will be explored in future studies by examining possible relationships between the geographical distribution of habitat and topographic diversity on the one hand, and the location of hotspot contact zones on the other hand.
Acknowledgments
We are grateful to F. R. Schram for helpful comments on earlier drafts of this manuscript, to P. H. Williams (Natural History Museum, London) for implementing the Palaearctic map in the Worldmap program, to P.G.M. Mekenkamp (University of Utrecht) for generating our equal area map of the Palaearctic and to two anonymous referees for the helpful comments and critical review of this manuscript.
Supplementary Material
References
- Balmford A, Moore J.L, Brooks T, Burgess N, Hansen L.A, Williams P, Rahbek C. Conservation conflicts across Africa. Science. 2001;291:2616–2629. doi: 10.1126/science.291.5513.2616. [DOI] [PubMed] [Google Scholar]
- Bokma F, Bokma J, Monkkonen M. Random processes and geographic species richness patterns: why so few species in the north? Ecography. 2001;24:43–49. [Google Scholar]
- Boone R.B, Krohn W.B. Partitioning sources of variation in vertebrate species richness. J. Biogeogr. 2000;27:457–470. [Google Scholar]
- Bures S, Nadvornik P, Saetre G.P. Brief report—hybridisation and apparent hybridisation between meadow pipit (Anthus pratensis) and water pipit (A. spinoletta) Hereditas. 2002;136:254–256. doi: 10.1034/j.1601-5223.2002.1360313.x. [DOI] [PubMed] [Google Scholar]
- Colwell R.K, Hurtt G.C. Nonbiological gradients in species richness and a spurious Rapoport effect. Am. Nat. 1994;144:570–595. [Google Scholar]
- Colwell R.K, Lees D.C. The mid-domain effect: geometric constraints on the geography of species richness. Trends Ecol. Evol. 2000;15:70–76. doi: 10.1016/s0169-5347(99)01767-x. [DOI] [PubMed] [Google Scholar]
- Diniz J.A.F, de Sant'Ana C.E.R, de Souza M.C, de Souza M.C, Rangel T.F.L.V.B. Null models and spatial patterns of species richness in South American birds of prey. Ecol. Lett. 2002;5:47–55. [Google Scholar]
- Dixon K.L. Contact zones of avian congeners on the southern Great Plains. Condor. 1989;91:15–22. [Google Scholar]
- Haffer J. Secondary contact zones of birds in northern Iran. Bonn. Zool. Monogr. 1977;10:64. [Google Scholar]
- Haffer J. Parapatric species of birds. Bull. Br. Ornithol. Cl. 1992;112:250–264. [Google Scholar]
- Haffer J. Contact zones between birds of southern Amazonia. Ornithol. Monogr. 1997;48:281–305. [Google Scholar]
- Haffer J, Prance G.T. Climatic forcing of evolution in Amazonia during the Cenozoic: on the refuge theory of biotic differentiation. Amazoniana. 2001;16:579–607. [Google Scholar]
- Hawkins B.A, Diniz J.A.F. The mid-domain effect cannot explain the diversity gradient of Nearctic birds. Global Ecol. Biogeogr. 2002;12:53–64. [Google Scholar]
- Jetz W, Rahbek C. Geometric constraints explain much of the species richness pattern in African birds. Proc. Natl Acad. Sci. USA. 2001;98:5561–5666. doi: 10.1073/pnas.091100998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayr E. Harvard University Press; Cambridge, USA: 1963. Animal species and evolution. [Google Scholar]
- Randler C. Avian hybridisation, mixed pairing and female choice. Anim. Behav. 2002;63:103–119. [Google Scholar]
- Rangel T.F.L.V.D, Diniz J.A.F. Spatial patterns in species richness and the geometric constraint simulation model: a global analysis of mid-domain effect in Falconiformes. Acta Oecol. 2003;24:203–207. [Google Scholar]
- Roselaar, C. S. & Shirihai, H. In press. Handbook of geographical variation and distribution of Palearctic Birds, vol. 1, Passerines. London: A & C Black.
- Short L.L. Taxonomic aspects of avian hybridization. Auk. 1969;86:84–105. [Google Scholar]
- Vaurie, C. 1954–1965 Systematic notes on Palearctic birds. No. 5, 6, 9, 24, 26, 27, 35, 44, 46, 49. Am. Mus. Nov. 1668, 1669, 1685, 1814, 1852, 1863, 1946, 2035, 2039, 2058.
- Williams, P. H. 2000. WORLDMAP: priority areas for biodiversity, Version 4.1. London (privately distributed).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.