

Abstract
Megarachne servinei from the Permo-Carboniferous Bajo de Véliz Formation of San Luis Province, Argentina (32° 17′ S, 65° 25′ E), was described as a giant mygalomorph spider (‘tarantula’) and, with its body length of 339 mm, the largest known spider ever to have lived on Earth. Its identification as a spider was based on interpretations of the shape of the carapace, the position of the eye tubercle, the anterior protrusion of the carapace as a pair of chelicerae, and the posterior circular structure as the abdomen. X-radiography revealed possible morphology hidden in the matrix: cheliceral fangs, sternum, labium and coxae, and so a reconstruction of Megarachne as a giant spider was presented. Difficulties with the interpretation (unusual cuticular ornament, suture dividing the carapace and spade-like anterior border of the chelicera), together with non-preservation of synapomorphies of Araneae, provoked debate about its interpretation as a spider. Now, the holotype and a new specimen have become available for study. Megarachne is shown to be a bizarre eurypterid (‘sea-scorpion’), similar to rare forms known from Carboniferous rocks of Scotland and South Africa, and is the most complete eurypterid so far recorded from Carboniferous strata of South America.
Keywords: Chelicerata, Eurypterida, Arachnida, Permo-Carboniferous, Argentina
1. Introduction
It is a common impression that the largest representative of any group of plants and animals is long extinct, so it was no surprise when Hünicken (1980) described Megarachne servinei from Permo-Carboniferous strata of Argentina as the largest spider that had ever lived. Its estimated 50 cm leg-span greatly exceeds that of the next largest spider (living or extinct), Theraphosa leblondi, with a leg-span of 30 cm. Hünicken (1980) presented a detailed description, illustrations and reconstructive drawings of Megarachne based, to a large extent, on X-ray studies. Plaster casts were eagerly acquired by museums around the world and form the basis of many displays. However, doubt about the interpretation of Megarachne as a spider was expressed by some arachnologists (e.g. Shear et al. 1989; Eskov & Zonshtein 1990). Access to Megarachne for study was difficult until recently because the holotype was deposited in a bank vault, but it has now been accessioned to the Museum of Paleontology, National University of Córdoba, and a new specimen has been discovered in the same locality and horizon.
The original interpretation of Megarachne as a spider was based on the general shape of the carapace and position of the eye tubercle, interpretation of the anteromedian protrusion of the carapace with its median ridge as a pair of spatulate chelicerae, and the circular structure posterior to the first tergite as the abdomen. Curved lines on the X-radiographs were interpreted as structures hidden in the matrix, for example, cheliceral fangs, sternum, labium and coxae. Some difficulties with the spider interpretation were noted by Hünicken: the unusual cuticular ornamentation, the suture dividing the carapace into anterior and posterior areas, and the spatulate chelicerae are all unknown in any other spider. In this new study, these morphological features are interpreted differently and in comparison with other giant chelicerates of the Carboniferous Period. Megarachne is not a spider, but a giant eurypterid akin to Woodwardopterus (Kjellesvig-Waering 1959).
2. Material and methods
The Electronic Appendix contains details of locality, stratigraphy and repository of the fossils. Material was photographed on Fuji Provia 100F slide film with a Minolta Dynax 9 camera, scanned at 3200 dpi on an Epson scanner and manipulated in Adobe Photoshop CS on a Macintosh PowerBook G4. Drawings were made using Adobe Illustrator CS on the same computer.
3. Results
(a) Preservation
Megarachne comes from the upper part of the Pallero Member, the middle of three members constituting the Bajo de Véliz Formation, which also contains well-preserved plants, insects and the trigonotarbid arachnid Gondwanarachne (Pinto & Hünicken 1980). Sedimentary features such as varved clayrocks point to an alluvial/lacustrine environment into which the terrestrial biota have been washed (Hünicken & Pensa 1975). The holotype (figure 1a,b) occurs on dark grey, laminated mudrock. The cuticle is preserved as a carbonized replacement: there is a thin calcite layer beneath this carbon layer, overlying the internal mould, and a slightly thicker one above (see figure 1a: tergite 2). The matrix shows thin sheets of calcite on rock laminae, so the mineralization is secondary. The fossil is part only (dorsal surface), but in places the dorsal cuticle is broken away, revealing the ventral cuticle.
Figure 1.
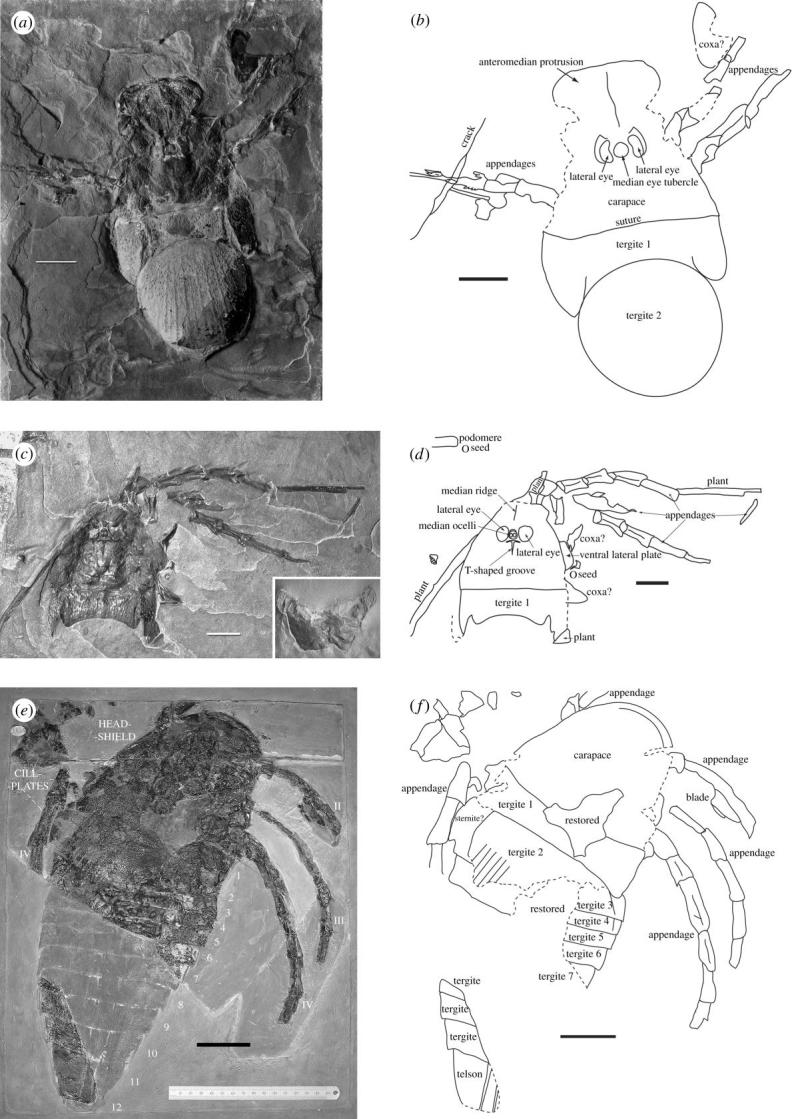
(a) Photograph and (b) camera lucida drawing of holotype specimen of M. servinei (Hünicken 1980); (c) photograph and (d) camera lucida drawing of second specimen of M. servinei (inset to (c) shows ventral side of specimen); (e) photograph and (f) camera lucida drawing of holotype specimen of Woodwardopterus scabrosus (Woodward 1887). Dashed lines mark broken edges. Scale bars, 5 cm.
The new specimen (figure 1c,d) consists of part and counterpart, each on a flagstone of about the same size as the original. The part shows the dorsal surface and its reverse shows ventral structures. The counterpart shows the external mould of the dorsal surface. The new specimen shows no calcite coating to the carbonized cuticle and consists of the carapace and first tergite, but no trace of the second tergite (the supposed spider abdomen).
(b) Morphological interpretation
The Electronic Appendix contains formal systematics including detailed descriptions, but only reinterpretation is considered here.
In the X-radiographs, darker areas correspond to thicker and lighter areas to thinner areas of rock matrix. The curved lines, which were interpreted as morphological features, therefore actually represent the boundaries of thicker and thinner areas—edges where rock laminae have broken away with a curved fracture. Thus, X-radiography provides no morphological information and the identifications of hidden structures should be discounted. The interpretation of the anterior structure of the carapace as digging chelicerae is also erroneous. The anterior border, complete with doublure, is well defined, but the lateral edges are broken and the apparent embayments alongside the eyes are where the carapace edge has been broken away, possibly by being compressed against robust appendages during fossilization (figure 1a,b). There is no articulation at the base of the supposed chelicerae and the cuticular sculpture continues without a break from the anterior area backwards to the main part of the carapace. Similarly, the median line previously interpreted as a cleft between left and right chelicerae is simply a median ridge, and the cuticular ornament can be seen to continue around its anterior and posterior ends. The ‘four stout spines’ on the anterior border of the carapace consist of the broken lateral edges of the carapace (lateral spines) and acute pre-ocular mucrones (median spines). The ocular tubercle bears the median eye-pair; large reniform structures on either side of the tubercle are the true lateral eyes. A large first opisthosomal tergite is sutured onto the rear of the carapace, followed by a second large subcircular plate with its surface covered by mucrones and posteriorly radiating ridges. Remains of three appendages are preserved, two on the right and one on the left, though ghosts of appendages in the matrix can also be seen. Most podomeres are stout, with thickened joints and a posterior longitudinal groove. Blade-like structures can be seen on the podomeres of the left side. An incomplete plate adjacent to the anteriormost preserved appendage on the right side may be part of a coxa or metastoma (median ventral plate in eurypterids).
The carapace of the new specimen shows no division between its main part and the anteromedian protrusion, which is incomplete in this specimen and less prominent than that on the holotype. A pair of large subcircular-reniform lateral eyes lies on either side of the median ocular tubercle, which bears a pair of ocelli with a lunate mound in front. Behind the eyes is a T-shaped groove (figure 1c,d) and in front is the median ridge. Two coxae and the ventral lateral plate can be seen to protrude around the edge of the carapace. The ventral side shows a pair of large coxae with toothed gnathobases (figure 1c, inset), each with two trochanters attached anterolaterally. In front of the coxae lies an ovoid plate that could be part of another coxa or the epistoma (anterior ventral plate in eurypterids).
4. Discussion
Many features of Megarachne indicate its assignment to the Eurypterida; for example, the cuticular sculpture of mucrones and raised lunules are characteristic of eurypterids. Apart from the small, streamlined Adelophthalmidae (Tollerton 1989), Permo-Carboniferous eurypterids are bizarre, giant forms. Table 1 compares Megarachne with these genera, from which it can be seen that Megarachne most closely resembles Woodwardopterus.
Table 1.
Comparison of morphological features of Megarachne with other large Carboniferous eurypterids. (✓, presence; ×, absence; ?, unknown.)
| features | Megarachne (Hünicken 1980) | Woodwardopterus (Kjellesvig-Waering 1959) | Mycterops (Cope 1886) | Cyrtoctenus (Størmer & Waterston 1968) | Hibbertopterus (Kjellesvig-Waering 1959) | Dunsopterus (Waterston 1968) | Vernonopterus (Waterston 1968) |
|---|---|---|---|---|---|---|---|
| carapace length (cm) | 17 | 15 | 5 | 35.5 | 38 | >10 | ? |
| linguoid ornament | scattered | scattered | dense | localized | scattered | scattered | minute |
| anteromedian carapace protrusion | ✓ | × | ? | ✓ | × | ? | ? |
| reniform lateral eyes, median eyes on tubercle | ✓ | ✓ | ✓ | ✓ | ✓ | ? | ? |
| enlarged first opisthosomal tergite | ✓ | ✓ | ✓ | × | × | ? | ? |
| enlarged second opisthosomal tergite | ✓ | ✓ | ? | × | × | ? | ? |
| thickened podomeres | ✓ | ✓ | ? | ✓ | ✓ | ✓ | ? |
| longitudinal grooves on podomeres | ✓ | ✓ | ? | ✓ | × | ✓ | ? |
| blade-like structures on podomeres | ✓ | ✓ | ? | ✓ | ✓ | ? | ? |
| coxal Laden | ✓ | ? | ? | × | ✓ | ? | ? |
| hastate telson with pair of carinae | ? | ✓ | ? | ✓ | ✓ | ? | ? |
Woodwardopterus is known only from the holotype, W. scabrosus (Woodward 1887), detailed description of which is provided in the Electronic Appendix. The first tergite is large and sutured to the carapace. Its lateral edges are obscure but epimera are suggested on the right-hand side. CaCO3 pustules obscure the eye region. Only the anterior part of the second tergite is preserved; it was clearly much larger but is broken around the lateral and posterior edges. Nevertheless, some radiating ridges can be seen (figure 1e,f). Parts of five more posterior tergites are preserved on the right‐hand side, which are delineated by clear tergal boundaries and are much shorter than tergites 1 and 2. A detached piece of rock belonging to the holotype was illustrated by Woodward (1887) and represents parts of the posteriormost tergites and the telson (figure 1e). The telson is a smooth plate with a pair of slight ridges, considerably longer than the preceding tergites, but lacking its posterior end. All features of the preserved portion of the enlarged second tergite of Woodwardopterus agree with the morphology of the second tergite of Megarachne, including the radiating lines not previously noted in Woodwardopterus. The second tergite must have extended beyond and partly covered more posterior tergites, because of the 20° angle between the anterior borders of tergite 1 and 2 and the anterior borders of more posterior tergites (figure 1f). The podomeres of Woodwardopterus are short, with thickened ends and longitudinal grooves as in Megarachne, and a blade-like structure can be seen on the most anterior preserved appendage (figure 1e,f).
We conclude that Megarachne and Woodwardopterus are confamilial (Woodwardopteridae; Kjellesvig-Waering 1959), but there are two differences between them. First, the mucrones on the carapace and enlarged tergites are densely packed in Woodwardopterus, but sparser in Megarachne. This could be a function of ontogeny because Megarachne is larger than Woodwardopterus, and possibly the mucrones became sparser with growth. The generally smaller Mycterops has still more densely packed ornament on the carapace and first tergite, which becomes mucronate and folliculated on other parts (Kjellesvig-Waering 1959), so it could represent a still younger form. Note that the gigantic Cyrtoctenus (Størmer & Waterston 1968) has localized mucrones. Second, the prominent anteromedian carapace protrusion in Megarachne is not seen in Woodwardopterus, but may be taphonomic because it is not as pronounced in the second specimen of Megarachne. It may have been orientated downwards in life and compression during fossilization affected its appearance in the holotype. An anteromedian protrusion occurs in Cyrtoctenus (Waterston et al. 1985), although this animal does not show enlarged tergites. The telson in Woodwardopterus is of a similar morphology to those in Cyrtoctenus (Waterston et al. 1985), Hibbertopterus (Kjellesvig-Waering 1959; Jeram & Selden 1994) and Hastimima (White 1908), which is the only other named eurypterid from the Carboniferous of South America. The blade-like structures on the appendages of Megarachne and Woodwardopterus compare to those in Cyrtoctenus (Waterston et al. 1985), but none of the large, pectinate blades of that genus have been found in association with woodwardopterids. Coxal Laden, seen in Megarachne emerging from beneath the carapace (figure 1c,d), are found in Hibbertopterus. The large coxae of Megarachne (figure 1c, inset) are typical of eurypterids, but Hibbertopterus has a large, triangular ‘median ventral plate’ (metastoma) in this position (Waterston 1957). Waterston illustrated a supposed genital plate beneath the metastoma in one specimen of Hibbertopterus. It is possible that this plate is really the metastoma and that the triangular ‘median ventral plate’ represents (possibly fused) coxae VI. Waterston et al. (1985) distinguished Hibbertopteridae from Cyrtoctenidae on the possession of Laden and ungrooved podomeres in the former, but Jeram & Selden (1994) considered that hibbertopterids could simply represent juvenile cyrtoctenids in which these features had yet to develop.
Figure 2 is a suggested reconstruction of Megarachne. Vestiges of blade-like structures on the anterior appendages suggest a sediment-raking method of feeding, as in Hibbertopterus and Cyrtoctenus, which implies aquatic feeding, but sedimentological evidence points to a non-marine habitat. In addition, the function of the large, circular second opisthosomal tergite remains a mystery. There are many puzzles yet to be solved regarding the functional morphology and mode of life of Megarachne, but its identity as a bizarre eurypterid, rather than a spider, points in the right direction.
Figure 2.
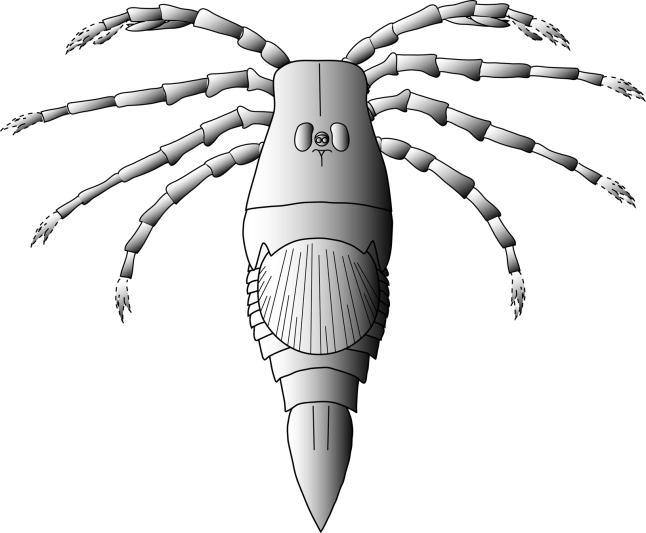
Hypothetical reconstruction of Megarachne based on features of the holotype and second specimen together with Woodwardopterus (metasoma, telson) and comparison with Cyrtoctenus (metasoma, telson). Dashed lines show anatomy reconstructed without direct evidence; chelicerae and palps not known. Total length: ca 54 cm.
Acknowledgments
We thank Guido Pollini (San Luis, Argentina) for facilitating a study of the new specimen of Megarachne, Andrew Ross (Natural History Museum, London) for access to Woodwardopterus, Lyall Anderson (Museum of Scotland, Edinburgh), Simon Braddy (University of Bristol), and an anonymous referee. P.A.S. thanks the Leverhulme Trust and the Natural Environment Research Council, and J.A.C. thanks CONICET (Consejo Nacional de Investigaciones Cientificas y Técnica, Argentina) for their financial support.
Supplementary Material
References
- Cope E.D. An interesting connecting genus of Chordata (Mycterops) Am. Nat. 1886;20:1027–1031. [Google Scholar]
- Eskov K, Zonshtein S. First Mesozoic mygalomorph spiders from the Lower Cretaceous of Siberia and Mongolia, with notes on the system and evolution of the infraorder Mygalomorphae (Chelicerata: Araneae) N. Jb. Geol. Paläontol. Abh. 1990;178:325–368. [Google Scholar]
- Hünicken M.A. A giant fossil spider (Megarachne servinei) from Bajo de Véliz, Upper Carboniferous, Argentina. Bol. Acad. Nac. Ciencias, Córdoba. 1980;53:317–341. [Google Scholar]
- Hünicken M.A, Pensa M.V. Estratigrafía y tectónica de los depósitos gondwánicos del Bajo de Véliz, Pcia. de San Luis, R. Argentina. Rev. Fac. Cs. Ex., Fs. Nat., Sec. C, Geol. 1975;3:1–37. [Google Scholar]
- Jeram A.J, Selden P.A. Eurypterids from the Viséan of East Kirkton, West Lothian, Scotland. Trans. R. Soc. Edinb.: Earth Sci. 1994;84:301–308. [Google Scholar]
- Kjellesvig-Waering E.N. A taxonomic review of some late Paleozoic Eurypterida. J. Paleontol. 1959;33:251–256. [Google Scholar]
- Pinto I.D, Hünicken M.A. Gondwanarachne, a new genus of the order Trigonotarbida (Arachnida) from Argentina. Bol. Acad. Nac. Cienc., Córdoba. 1980;53:307–315. [Google Scholar]
- Shear W.A, Palmer J.M, Coddington J.A, Bonamo P.M. A Devonian spinneret: early evidence of spiders and silk use. Science. 1989;246:479–481. doi: 10.1126/science.246.4929.479. [DOI] [PubMed] [Google Scholar]
- Størmer L, Waterston C.D. Cyrtoctenus gen. nov., a large late Palaeozoic arthropod with pectinate appendages. Trans. R. Soc. Edinb. 1968;68:63–104. [Google Scholar]
- Tollerton V.P. Morphology, taxonomy, and classification of the order Eurypterida Burmeister, 1853. J. Paleontol. 1989;63:642–657. [Google Scholar]
- Waterston C.D. The Scottish Carboniferous Eurypterida. Trans. R. Soc. Edinb. 1957;63:265–288. [Google Scholar]
- Waterston C.D. Further observations on the Scottish Carboniferous eurypterids. Trans. R. Soc. Edinb. 1968;68:1–20. [Google Scholar]
- Waterston C.D, Oelefsen B.W, Oosthuizen R.D.F. Cyrtoctenus wittebergensis sp. nov. (Chelicerata: Eurypterida), a large sweep-feeder from the Carboniferous of South Africa. Trans. R. Soc. Edinb.: Earth Sci. 1985;76:339–358. [Google Scholar]
- White D. Report on the fossil flora of the coal measures of Brazil. In: White J.C, editor. In Final report on the coal measures and associated rocks of South Brazil. National Press; Rio de Janeiro: 1908. pp. 377–607. [Google Scholar]
- Woodward H. On a new species of Eurypterus from the Lower Carboniferous shales of Glencartholm, Eskdale, Scotland. Geol. Mag. 1887;4:481–484. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.