

Abstract
Strain Hxd3, an alkane-degrading sulfate reducer previously isolated and described by Aeckersberg et al. (F. Aeckersberg, F. Bak, and F. Widdel, Arch. Microbiol. 156:5-14, 1991), was studied for its alkane degradation mechanism by using deuterium and 13C-labeled compounds. Deuterated fatty acids with even numbers of C atoms (C-even) and 13C-labeled fatty acids with odd numbers of C atoms (C-odd) were recovered from cultures of Hxd3 grown on perdeuterated pentadecane and [1,2-13C2]hexadecane, respectively, underscoring evidence that C-odd alkanes are transformed to C-even fatty acids and vice versa. When Hxd3 was grown on unlabeled hexadecane in the presence of [13C]bicarbonate, the resulting 15:0 fatty acid, which was one carbon shorter than the alkane, incorporated a 13C label to form its carboxyl group. The same results were observed when tetradecane, pentadecane, and perdeuterated pentadecane were used as the substrates. These observations indicate that the initial attack of alkanes includes both carboxylation with inorganic bicarbonate and the removal of two carbon atoms from the alkane chain terminus, resulting in a fatty acid one carbon shorter than the original alkane. The removal of two terminal carbon atoms is further evidenced by the observation that the [1,2-13C2]hexadecane-derived fatty acids contained either two 13C labels located exclusively at their acyl chain termini or none at all. Furthermore, when perdeuterated pentadecane was used as the substrate, the 14:0 and 16:0 fatty acids formed both carried the same numbers of deuterium labels, while the latter was not deuterated at its carboxyl end. These observations provide further evidence that the 14:0 fatty acid was initially formed from perdeuterated pentadecane, while the 16:0 fatty acid was produced after chain elongation of the former fatty acid with nondeuterated carbon atoms. We propose that strain Hxd3 anaerobically transforms an alkane to a fatty acid through a mechanism which includes subterminal carboxylation at the C-3 position of the alkane and elimination of the two adjacent terminal carbon atoms.
Alkanes are commonly found in petroleum-contaminated environments, and the biodegradation of these compounds under aerobic conditions has been well studied (6, 42, 43). On the other hand, while the occurrence of anaerobic alkane biodegradation has been reported (7, 9, 10, 13, 26, 34, 35, 45, 47), still little is known about the diversity of the responsible organisms and the degradation mechanisms. Aeckersberg et al. reported on the first well-described alkane-degrading, sulfate-reducing bacterium, strain Hxd3 (DSM 6200; German Collection of Microorganisms and Cell Cultures), from an oil production plant (1, 2). This strain is able to grow anaerobically on C12 to C20 alkanes and completely oxidize them to carbon dioxide by coupling to sulfate reduction (1, 2). The phenotypic and phylogenetic characteristics of the bacterium indicate that it belongs to the proposed family Desulfobacteriaceae and is closely related to the genus Desulfococcus (1, 3). Recently, So and Young also isolated and characterized a phylogenetically different sulfate reducer, strain AK-01, which can completely oxidize alkanes to CO2 under anaerobic conditions (37, 38). Additionally, three denitrifying alkane degraders were described, but their phylogeny is unclear (14). These organisms were designated HxN1, OcN1, and HdN1. They utilize alkanes with chain lengths from C6 to C8, C8 to C12, and C14 to C20, respectively.
The mechanism of alkane degradation by Hxd3 was examined by Aeckersberg et al., who reported that the chain lengths of alkane substrates had a pronounced effect on the total cellular fatty acid composition of the bacterium. An alkane with an odd number of C atoms (C-odd) yielded predominantly fatty acids with an even number of C atoms (C-even), and a C-even alkane yielded C-odd fatty acids (3). These observations suggested that an alkane was altered by an odd-numbered carbon(s) during its transformation to fatty acids (3). In contrast, strain AK-01 forms C-even fatty acids when grown on C-even alkanes and vice versa (38). Additional evidence reported by Aeckersberg et al. (1, 3) indicated that 1-alkene was unlikely to be an intermediate, as suggested by earlier literature (8, 29).
In previous investigations, stable isotope-labeled hydrocarbon substrates were used to help delineate the pathways of anaerobic degradation by both pure and mixed cultures. They proved to be useful tools for examining the initial mechanisms of attack on compounds such as benzene, naphthalene, and phenanthrene and alkane degradation by strain AK-01 (30, 38, 39, 48, 49). In this report, we present the results of an investigation of the initial reactions of alkane degradation by strain Hxd3. By using deuterated and 13C-labeled alkanes, we provide direct evidence that alkanes are oxidized to fatty acids by the bacterium. In addition, based on a detailed mass spectral analysis of the isotope-labeled fatty acids derived from the labeled alkanes, a proposed mechanism for the initial attack on alkanes is described.
MATERIALS AND METHODS
Source of bacterium and culture conditions.
A stock culture of strain Hxd3 (DSM 6200) was obtained from the German Collection of Microorganisms and Cell Cultures. Medium no. 517, recommended by the German Collection of Microorganisms and Cell Cultures, was modified according to suggestions from Friedrich Widdel (personal communication) and used for culturing the bacterium. The medium contained, per liter of deionized water, the following: NaCl, 17.6 g; MgCl2 · 6H2O, 3 g; CaCl2 · 2H2O, 0.22 g; NH4Cl, 0.22 g; KH2PO4, 0.18 g; Na2SO4, 2 g; NaHCO3, 2.5 g; Na2S · 9H2O, 0.12 g; vitamin solution, 1 ml; trace element solution, 1 ml; and selenite-tungstate solution, 1 ml. The vitamin solution was modified from that of Widdel and Bak (44) and contained, per liter of deionized water, the following: vitamin B12, 50 mg; d-(+)-biotin, 20 mg; folic acid, 20 mg; nicotinic acid, 50 mg; p-aminobenzoic acid, 50 mg; calcium d-(+)-pantothenate, 50 mg; pyridoxine HCl, 100 mg; riboflavin, 50 mg; thiamine HCl · 2H2O, 50 mg; thioctic acid, 50 mg; Na2HPO4, 0.577 g; and KH2PO4, 0.015 g (final pH, 7.0). The trace element solution was modified from that of Widdel and Bak (44) and contained, per liter of 0.1 N HCl, the following: CoCl2 · 6H2O, 190 mg; CuCl2 · 2H2O, 2 mg; FeCl2 · 4H2O, 4.5 g; H3BO3, 6 mg; MnCl2 · 4H2O, 100 mg; Na2MoO4 · 2H2O, 36 mg; NiCl2 · 6H2O, 24 mg; and ZnCl2, 140 mg. The selenite-tungstate solution contained, per liter of 0.01 N NaOH, the following: Na2SeO3, 2 mg; and Na2WO4 · 1.5H2O, 4 mg. All of the above medium components (except for NaHCO3, Na2S · 9H2O, and the vitamin solution) were added to deionized water and then deoxygenated by bubbling with a stream of N2-CO2 gas (70:30) for 1 h. The medium was dispensed into serum bottles, which were then crimp sealed with butyl rubber stoppers and autoclaved. A filter-sterilized solution containing 1 M NaHCO3, 18 mM Na2S · 9H2O, and 325 mM NaOH was added to the autoclaved medium (3% [vol/vol]). The vitamin solution was also filter sterilized and added to the medium (0.1% [vol/vol]). The final pH of the complete medium was 7.2. All cultures were incubated in the dark without shaking.
Effects of alkanes on cellular fatty acid compositions.
Each bottle of the mineral salt medium (100 ml) was amended with 20 μl of filter-sterilized pentadecane or hexadecane (99% purity) and inoculated with a culture of strain Hxd3 that had been maintained on the corresponding alkane substrate (20% [vol/vol]). For each substrate, two experimental cultures and one sterile control were set up. After incubation for 48 to 80 days, cultures were used for extraction and analysis of total cellular fatty acids.
Metabolism of aliphatic compounds in [13C]bicarbonate.
The mineral salt medium was prepared as described previously, except that N2 gas was used for degassing the medium and sodium [13C]bicarbonate (isotopic enrichment, 99%; Cambridge Isotope Laboratories, Inc., Andover, Ma.) was used instead of unlabeled sodium bicarbonate. The medium (40 ml per bottle) was amended with one of the following substrates: unlabeled tetradecane, pentadecane, or hexadecane, 20 μl; perdeuterated pentadecane, 20 μl; pentadecanoate or hexadecanoate, 0.5 mM; and 2-hexadecanone, 10 mg. Each bottle of complete medium was inoculated with a culture of Hxd3 that had been pregrown on stearate (0.5% [vol/vol]), and two experimental cultures were established with each substrate. Cultures were incubated for the following times, depending on the substrates used: unlabeled alkanes, 60 to 92 days; perdeuterated pentadecane, 97 days; fatty acids, 14 days; and 2-hexadecanone, 28 days. Whole cultures were then used for extraction and analysis of total cellular fatty acids.
Metabolism of [1,2-13C2]hexadecane.
The mineral salt medium was amended with filter-sterilized [1,2-13C2]hexadecane (20 μl per 100 ml of medium; isotopic enrichment, 99%; Isotec Inc., Miamisburg, Ohio) and inoculated with a culture of Hxd3 that had been pregrown on stearate (5% [vol/vol]). Two experimental cultures and one sterile control were prepared. The sterile control was identical to the experimental cultures except that the inoculum was autoclaved before addition. All cultures were incubated for 63 days and then used for extraction and analysis of total cellular fatty acids.
Metabolism of perdeuterated pentadecane.
The mineral salt medium was amended with filter-sterilized perdeuterated pentadecane (20 μl per 50 ml of medium; isotopic enrichment, 98%; Cambridge Isotope Laboratories) and inoculated with a culture of Hxd3 that had been pregrown on stearate (1% [vol/vol]). Two experimental cultures and one sterile control were prepared. The sterile control was identical to the experimental cultures except that the inoculum was autoclaved before addition. All cultures were incubated for 103 days and then used for extraction and analysis of total cellular fatty acids.
Extraction and analysis of total cellular fatty acids.
The extraction of total cellular fatty acids was carried out by a method recommended for The Sherlock Microbial Identification System (MIDI, Inc., Newark, Del.). The method involves filtration of cells, saponification of cellular lipids in a hot NaOH-methanol solution, methylation of fatty acids with hot HCl-methanol, and extraction with hexane-methyl-tert-butyl ether. The methylated fatty acids were analyzed by using a gas chromatograph coupled to a flame ionization detector (model 5890 series II; Hewlett-Packard, Wilmington, Del.) to determine the fatty acid compositions of cells under different growth conditions. A mass spectrum detector (5971 series; Hewlett-Packard) was used to further identify individual fatty acids. These methods were described in detail in an earlier report (38). The various monomethylated and stable isotope-labeled fatty acids were identified by interpretation of their mass spectra based on studies reported in the literature (16, 25, 27, 28, 36), as authentic standards are not available. Locations of deuterium atoms or 13C atoms on the stable isotope-labeled fatty acids were determined by detailed analysis of the related mass spectra based on information from the literature in which fragmentation mechanisms of fatty acid methyl esters are reported (11, 25). The fatty acids in each sample were generally quantified as a percentage of the total recovered fatty acids based on the peak area of each fatty acid appearing in the total ion chromatogram. Preliminary studies of a standard mixture of fatty acid methyl esters showed that they all have similar response ratios based on the total mass of each compound.
Nomenclature of fatty acids.
The nomenclature of fatty acids recommended by the International Union of Pure and Applied Chemistry-International Union of Biochemistry (20) was adopted in this report. An n-saturated hexadecanoic acid is designated 16:0, with the first number representing the number of carbon atoms on the acyl group and the second number representing the number of double bonds present. A branched fatty acid, such as 10-methylhexadecanoic acid, is designated 10-Me-16:0.
RESULTS
Transformation of alkanes to fatty acids.
The cellular fatty acid composition of strain Hxd3 is clearly affected by the chain lengths of the alkane substrates. Figure 1 shows the fatty acid profiles of Hxd3 grown on pentadecane and hexadecane, as determined by gas chromatography with a flame ionization detector. When pentadecane is used as a growth substrate, the 14:0 and 16:0 fatty acids (both C-even) are dominant, while C-odd ones are virtually absent. In contrast, when hexadecane is used, C-odd, n-saturated fatty acids (13:0, 15:0, and 17:0) predominate over C-even ones.
FIG. 1.

Gas chromatograms (gas chromatography with a flame ionization detector) showing the fatty acid profiles of strain Hxd3 grown on pentadecane and hexadecane. The n-saturated fatty acids, identified by their retention times, are annotated. Underlined designations indicate C-odd fatty acids.
The detailed compositions and relative abundance of the total cellular fatty acids of Hxd3 grown on the two alkanes were determined further by gas chromatography-mass spectrometry. As summarized in Fig. 2a and c, pentadecane (C-odd) yields predominantly C-even fatty acids (94.7% of the total), while hexadecane (C-even) produces mostly C-odd fatty acids (79.3% of the total).
FIG. 2.

Relative abundance of the total cellular fatty acids of strain Hxd3 grown on pentadecane, perdeuterated pentadecane, hexadecane, and [1,2-13C2]hexadecane. The unlabeled C-even fatty acids are represented by black bars, and the unlabeled C-odd fatty acids are represented by white bars. For cultures grown on perdeuterated pentadecane and [1,2-13C2]hexadecane, only the deuterated (▨) and 13C-labeled (▧) fatty acids, respectively, are depicted. All fatty acids shown are identifiable by gas chromatography-mass spectrometry and comprise more than 1% of the total. None of these fatty acids was found in the sterile controls.
By using stable isotope-labeled alkanes and analyzing the fatty acids recovered from cultures of Hxd3, we are able to directly demonstrate that C-odd and C-even alkanes are indeed transformed to C-even and C-odd fatty acids, respectively. As shown in Fig. 2b, deuterated 14:0, 10-Me-14:0, 16:0, and 10-Me-16:0 fatty acids, all C-even, are found when perdeuterated pentadecane (C-odd) is used as the substrate. In addition, 13C-labeled 13:0, 15:0, 10-Me-15:0, 17:0, and 10-Me-17:0 fatty acids, all C-odd, are recovered when [1,2-13C2]hexadecane (C-even) is used as the substrate (Fig. 2d). None of these labeled fatty acids is found in the sterile controls. The relationship between the carbon numbers of the alkane substrates and the cellular fatty acids indicates that odd-numbered carbon atoms are added or removed from the alkanes during their transformation to fatty acids (3).
Analysis of alkane-derived fatty acids formed in [13C]bicarbonate-amended media.
Hxd3 was grown on unlabeled hexadecane in the presence of [13C]bicarbonate, and the fatty acids recovered were characterized by mass spectrometry. Figure 3 shows the mass spectra of the methyl esters of the 15:0 fatty acid formed in the presence of unlabeled bicarbonate or [13C]bicarbonate. As shown in Fig. 3a, the molecular ion peak for the methyl ester of the 15:0 fatty acid formed in the presence of unlabeled bicarbonate appears at m/z = 256, representing the molecular mass of the compound. The methyl ester of the 15:0 fatty acid formed in the presence of [13C]bicarbonate, however, has a molecular ion peak shifted up to m/z = 257 (Fig. 3b), suggesting that one 13C atom derived from [13C]bicarbonate is incorporated into this fatty acid.
FIG. 3.

Mass spectra of the methyl esters of the 15:0 fatty acid recovered from cultures of strain Hxd3 grown on hexadecane with unlabeled bicarbonate (a) or [13C]bicarbonate (b). Chemical structures represented by the mass spectra are shown as insets. Key diagnostic ion peaks are annotated in bold type with their m/z values, and the structural compositions of the represented ion fragments are delineated. Intersection with a dotted line indicates a point of bond cleavage, and the ion fragment formed subsequently contains only the part of the molecule to the left of the dotted line.
As depicted in Fig. 3a, the ion peak at m/z = 74 for the 15:0 fatty acid formed with unlabeled bicarbonate is due to an ion fragment which includes the C-1 and C-2 positions of the molecule along with the ester-linked methyl group. This ion fragment is formed by a process known as the McLafferty rearrangement, which involves the migration of a hydrogen from C-4 to the carbonyl oxygen and the cleavage of the C-2—C-3 bond (25). In addition, the ion fragment represented by the peak at m/z = 59 is formed by the cleavage of the C-1—C-2 bond and includes only the carboxyl group (C-1) of the fatty acid (25).
Further examination of the mass spectra indicates that the carboxyl group (C-1) of the 15:0 fatty acid is an exogenously added carbon derived from inorganic bicarbonate. As shown in Fig. 3b, the appearance of a peak at m/z = 75 indicates the presence of one 13C atom located within C-1 and C-2 of the molecule. Furthermore, the appearance of a peak at m/z = 60 indicates that the 13C atom is located at C-1. The results therefore show that the carboxyl group of the alkane-derived 15:0 fatty acid is added exogenously through carboxylation with inorganic bicarbonate.
Carboxylation with inorganic bicarbonate is also observed when other alkanes are used as substrates. Table 1 summarizes the occurrence of 13C on the cellular fatty acids of Hxd3 grown on various alkanes in the presence of [13C]bicarbonate, as determined by mass spectrometry (mass spectra not shown). Upon growth on tetradecane, pentadecane, and hexadecane, the fatty acids which are one carbon shorter than the alkane substrates, namely, 13:0, 14:0, and 15:0, respectively, all contain a 13C atom located at their carboxyl groups. Furthermore, when the organism is grown on perdeuterated pentadecane with [13C]bicarbonate, the deuterated 14:0 fatty acid formed also carries a 13C atom located at its carboxyl group (data not shown).
TABLE 1.
Occurrence of 13C on cellular fatty acids of strain Hxd3 grown on various unlabeled alkanes in the presence of [13C]bicarbonatea

Determined by interpretation of mass spectra based on information from the literature. None of the fatty acids shown was found in the sterile controls.
It should be noted, however, that the carboxylation reaction observed is not likely to be due to fatty acid metabolism by Hxd3. When Hxd3 is grown on pentadecanoic or hexadecanoic acid with [13C]bicarbonate, none of the recovered fatty acids has 13C incorporated in its carboxyl group. Hence, the carboxylation reaction is part of the initial attack for transforming alkanes to fatty acids but is not involved in fatty acid metabolism.
Further examination of other fatty acids formed in the presence of [13C]bicarbonate suggests that those with 13C-labeled carboxyl groups can be further metabolized by chain elongation to become one carbon longer than the parent alkane. As shown in Table 1, while fatty acids which are one carbon shorter than the original alkane substrates all have a 13C atom located at C-1 (carboxyl group), those which are one carbon longer than the alkanes contain a 13C atom located at C-3.
Analysis of [1,2-13C2]hexadecane-derived fatty acids.
Mass spectral analysis of the fatty acids derived from [1,2-13C2]hexadecane provides evidence that the two terminal carbon atoms are removed from the alkane chain during the formation of a fatty acid. Figure 4 shows the mass spectra of the 15:0 fatty acids (as methyl esters) recovered from cultures of Hxd3 grown on unlabeled hexadecane and [1,2-13C2]hexadecane. While the molecular ion peak for the methyl ester of the unlabeled 15:0 fatty acid appears at m/z = 256 (Fig. 4a), that for the 15:0 fatty acid recovered from the [1,2-13C2]hexadecane-grown culture appears as a doublet at m/z = 256 and m/z = 258 with comparable intensities (Fig. 4b). The results indicate that the 15:0 fatty acid derived from the labeled hexadecane occurs as two equally abundant “isotopmers” (chemical compounds having the same formula and structure but containing different isotopes). Isotopmer A contains two 13C labels (m/z = 258), while isotopmer B contains none at all (m/z = 256).
FIG. 4.

Mass spectra of the methyl esters of the 15:0 fatty acid recovered from cultures of strain Hxd3 grown on unlabeled hexadecane (a) and [1,2-13C2]hexadecane (b). Chemical structures represented by the mass spectra are shown as insets. The results shows that the 15:0 fatty acid derived from [1,2-13C2]hexadecane occurs as two different isotopmers, A and B.
Further examination of the mass spectral data shows that the two 13C labels are located at the alkyl chain terminus of the labeled isotopmer. As shown in Fig. 4a, the ion fragment at m/z = 143 includes C-1 to C-7 of the unlabeled 15:0 fatty acid (11, 24). The corresponding ion peak for the 15:0 fatty acid derived from [1,2-13C2]hexadecane remains at m/z = 143 (Fig. 4b), indicating the absence of a 13C label on C-1 to C-7 of either isotopmer A or isotopmer B. Since isotopmer A has two 13C labels (m/z = 258), those 13C labels are therefore located not at C-1 to C-7 but most likely at C-14 and C-15, their original locations on [1,2-13C2]hexadecane.
These observations suggest that (i) two terminal carbon atoms have been removed from [1,2-13C2]hexadecane and (ii) that this event can occur with equal likelihoods at both labeled and unlabeled termini. As a result, an initial attack at the unlabeled end of the hexadecane would produce a fatty acid isotopmer with the two 13C labels retained at its alkyl chain terminus, while an attack at the labeled end would produce an isotopmer with both labels eliminated.
In addition, the 13C-labeled 15:0 fatty acid is sometimes elongated to form a 17:0 fatty acid. Table 2 summarizes the characteristics of the 15:0 and 17:0 fatty acids formed from [1,2-13C2]hexadecane. Like the 15:0 fatty acid, the 17:0 fatty acid also occurs as two isotopmers of comparable abundances. One isotopmer contains two 13C labels most likely located at the alkyl chain terminus (C-16 and C-17), while the other isotopmer contains none at all. As demonstrated earlier by examination of the fatty acids formed from [13C]bicarbonate, these observations indicate that the 13C-labeled 17:0 fatty acid is derived from the 15:0 fatty acid by chain elongation.
TABLE 2.
Characteristics of 15:0 and 17:0 fatty acids recovered from cultures of strain Hxd3 grown on [1,2-13C2]hexadecanea
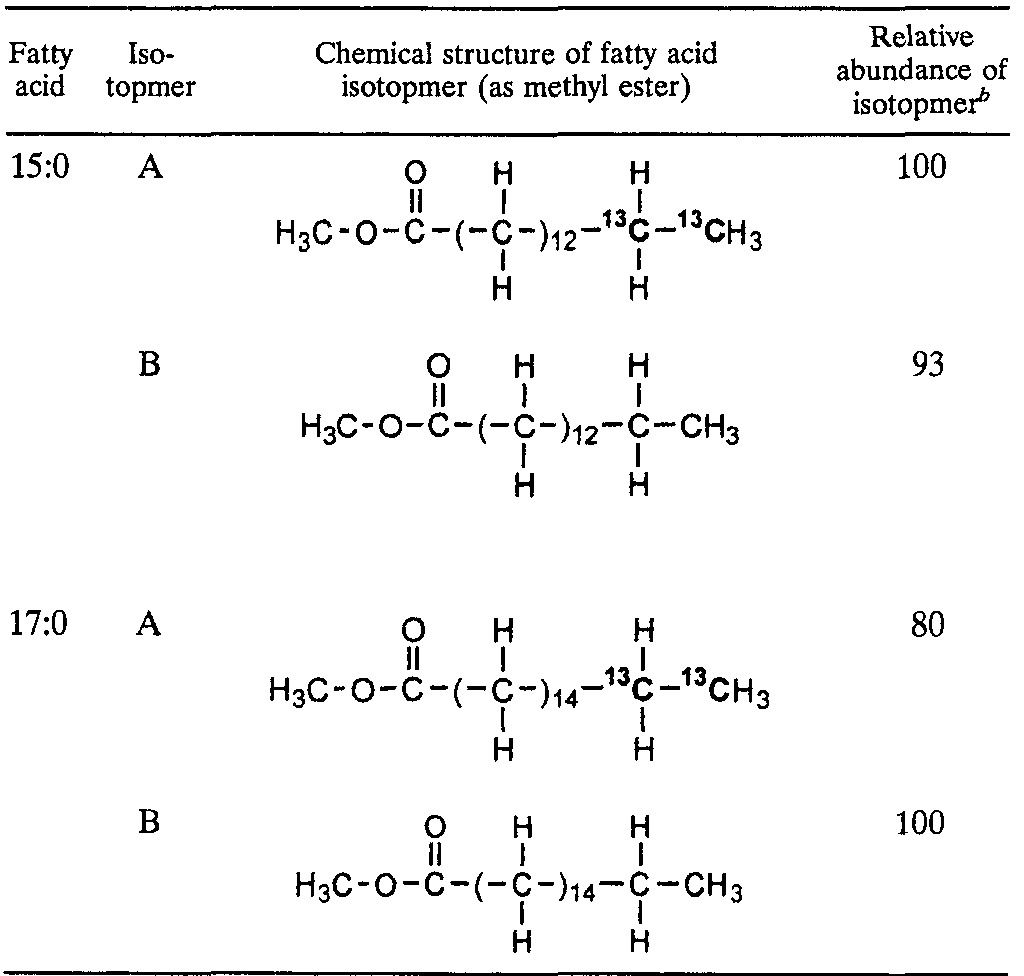
Determined by interpretation of mass spectra based on information from the literature. The 13:0, 10-Me-15:0, and 10-me-17:0 fatty acids not shown here were also 13C labeled and occurred as two isotopmers, like the 15:0 and 17:0 fatty acids. None of the fatty acids shown was found in the sterile controls.
Results are estimations based on the relative intensities of the M+ and (M + 2)+ molecular ion peaks for each fatty acid.
Analysis of perdeuterated pentadecane-derived fatty acids.
An examination of the deuterated fatty acids derived from perdeuterated pentadecane further supports the notion that an alkane is initially transformed to a fatty acid which is one carbon shorter but which subsequently can be elongated. Table 3 summarizes the structural characteristics of the deuterated fatty acids recovered from cultures of Hxd3 grown on perdeuterated pentadecane. Mass spectral analysis of the 14:0 fatty acid shows that it contains a total of 25 deuterium atoms, together with 1 hydrogen atom located at C-2 and another one within C-3 to C-5 of the molecule (exact positions uncertain). (The 14:0 fatty acid would carry 27 deuterium atoms if fully deuterated, i.e., C13D27COOCH3.) The loss of two deuterium atoms near its carboxyl end may be caused by isotope exchange during the initial attack of the alkane. On the other hand, the 16:0 fatty acid also carries a total of 25 deuterium atoms, yet with 2 hydrogen atoms located at C-2 and 4 more within C-3 to C-6 (Table 3). (The 16:0 fatty acid would carry 31 deuterium atoms if fully deuterated, i.e., C15D31COOCH3.). The observations that (i) the deuterated 14:0 and 16:0 fatty acids contain identical numbers of deuterium atoms and that (ii) the 16:0 fatty acid has six hydrogen atoms located at its carboxyl terminus suggest that the 16:0 fatty acid is formed by chain elongation of the deuterated 14:0 fatty acid with two nondeuterated carbon atoms derived from elsewhere (33).
TABLE 3.
Characteristics of deuterated fatty acids recovered from cultures of strain Hxd3 grown on perdeuterated pentadecane (C15D32)a
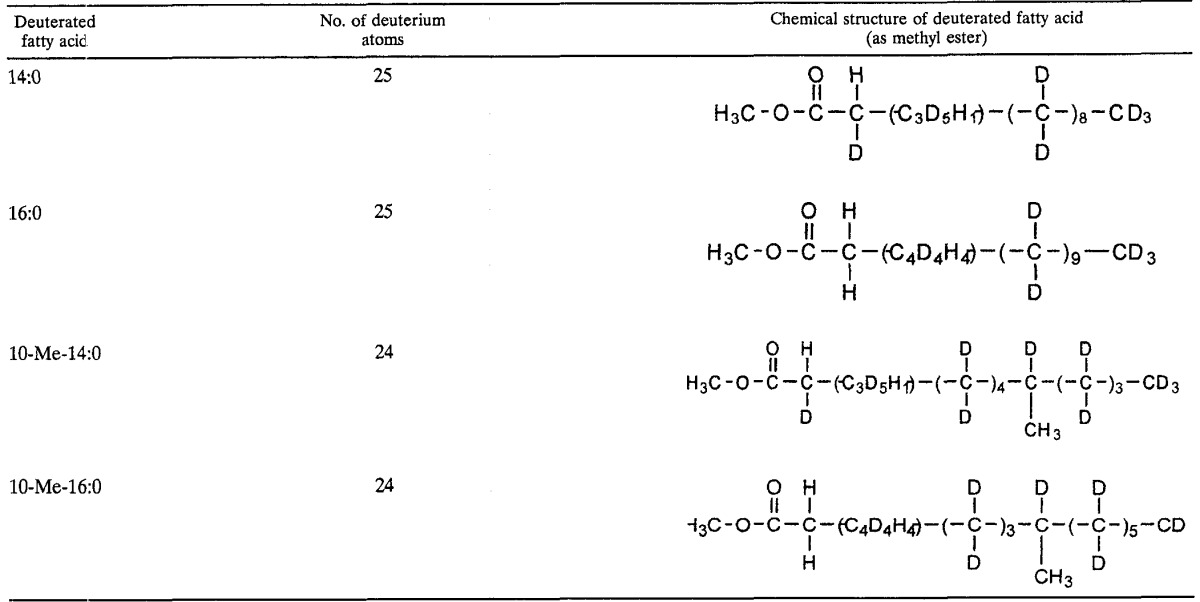
Determined by interpretation of mass spectra based on information from the literature. None of the fatty acids shown was found in the sterile controls.
Examination of the deuterated 10-methylated fatty acids indicates that they are formed by further metabolism of the existing n-saturated fatty acids. As shown in Table 3, the deuterated 10-Me-14:0 and 10-Me-16:0 fatty acids contain the same numbers of hydrogen atoms at their carboxyl termini as the nonmethylated 14:0 and 16:0 fatty acids, respectively. This finding suggests that the two 10-methylated fatty acids are derived from the corresponding n-saturated ones. Also, the methyl groups of the two 10-methylated fatty acids are not deuterated, indicating that they are exogenously added to the acyl chains.
Degradation of 2-hexadecanone in [13C]bicarbonate-amended media.
In order to determine whether 2-ketone is an intermediate formed before carboxylation takes place, Hxd3 was grown on 2-hexadecanone in the presence of [13C]bicarbonate, and the cellular fatty acids recovered from the cultures were characterized by mass spectrometry. Unlike hexadecane (Fig. 2c), 2-hexadecanone yields C-odd and C-even cellular fatty acids of comparable abundances (data not shown). In addition, the mass spectra of the 15:0 fatty acids recovered from cultures grown on the ketone with or without [13C]bicarbonate appear the same, indicating that no 13C is incorporated into the fatty acid. 2-Hexadecanone therefore is unlikely to be an intermediate formed before the carboxylation step during the initial attack of hexadecane.
DISCUSSION
Aeckersberg et al. first reported in 1991 the isolation of strain Hxd3, an alkane-degrading, sulfate-reducing bacterium, and demonstrated quantitatively its ability to oxidize alkanes to carbon dioxide by coupling to sulfate reduction (2). The phenotypic and phylogenetic characteristics of the strain were also described in detail in later studies (1, 3). Upon further investigations into the metabolism of alkanes by Hxd3, Aeckersberg et al. observed a clear impact of the chain lengths of the alkane substrates on the cellular fatty acid composition of the bacterium (3). This observation suggests that the strain anaerobically oxidizes alkanes to fatty acids and that it incorporates them into cellular lipids, a metabolic process observed in many aerobic alkane-degrading organisms (5, 12, 17, 21). The relationship between the carbon numbers of alkane substrates and predominant cellular fatty acids, however, was unusual; namely, a C-odd alkane yielded predominantly C-even fatty acids, and a C-even alkane yielded C-odd fatty acids. It was thus suggested that the initial attack of alkane by Hxd3 involves a novel mechanism in which an odd-numbered carbon(s) is added to or removed from the alkane chain (3).
In this study, we directly confirmed that alkanes are oxidized to fatty acids by strain Hxd3 on the basis of the recovery of 13C-labeled and deuterated fatty acids from cultures grown on the respective labeled alkanes (Fig. 2). Moreover, all deuterated fatty acids derived from perdeuterated pentadecane are C-even, while all 13C-labeled fatty acids derived from [1,2-13C2]hexadecane are C-odd. It is thus established that C-odd and C-even alkanes are indeed transformed to C-even and C-odd fatty acids, respectively, by Hxd3.
As a result of a detailed mass spectral examination of the stable isotope-labeled fatty acids, we conclude that an alkane is first transformed to a fatty acid which is one carbon shorter by the addition of an exogenous carbon atom to and by the removal of two terminal carbon atoms from the alkane chain. As summarized in Fig. 5, we propose that the initial attack of an alkane (A) involves subterminal carboxylation at the C-3 position of the alkyl chain (step I), thus forming intermediate B (hypothetical). The adjacent carbon atoms located at C-1 and C-2 on intermediate B are then removed (step II) to yield a fatty acid (C). This fatty acid (C) therefore contains a carboxyl group exogenously added by the carboxylation reaction, yet it is one carbon shorter than the parent alkane. The initially formed fatty acid (C) can be β-oxidized (step III) and subsequently mineralized to CO2. Alternatively, this fatty acid can undergo transformations such as chain elongation and C-10 methylation (step IV) to form other fatty acids (33).
FIG. 5.

Proposed pathway for the oxidation of alkane to fatty acid by strain Hxd3. An alkane (A) is subterminally carboxylated at C-3 (step I) to form an intermediate (B). Two adjacent terminal carbon atoms are then eliminated (step II) to form a fatty acid one carbon shorter than the original alkane (C). This fatty acid can be beta oxidized (step III) and subsequently mineralized to CO2 or undergo transformation, such as chain elongation and C-10 methylation (step IV). Compound B (in brackets) is only a hypothetical intermediate and has not been observed. Atoms originating from the alkane are shown in bold type.
The notion that carboxylation (Fig. 5, step I) is part of the initial attack of the alkane is supported by the following evidence. First, mass spectral analyses of 13:0, 14:0, and 15:0 fatty acids formed from tetradecane, pentadecane, and hexadecane, respectively, in the presence of [13C]bicarbonate indicate that each fatty acid contains a 13C atom located at its carboxyl group (Fig. 3b and Table 1). Second, the presence of a bicarbonate-derived 13C atom in the carboxyl group of the deuterated 14:0 fatty acid derived from perdeuterated pentadecane further supports the notion that carboxylation occurs during the transformation of alkane to fatty acid. Third, such a carboxylation reaction is not observed in the cellular fatty acids of Hxd3 when it is grown on long-chain fatty acids. Thus, the reaction does not occur when fatty acids are metabolized but is one of the initial reactions in alkane transformation.
Furthermore, the carboxylation reaction (Fig. 5, step I) appears to occur subterminally at C-3 of the alkane carbon chain and is followed by the removal of the two adjacent terminal carbon atoms (step II) to form a fatty acid (C). As shown in Table 1, the fatty acids that have incorporated an exogenous 13C atom in their carboxyl groups are all one carbon shorter than the parent alkanes. Hence, the simplest explanation for the phenomenon is that these alkanes are carboxylated at the C-3 position, and then the two adjacent carbon atoms at the C-1 and C-2 positions are eliminated. The fatty acids subsequently formed therefore contain a 13C-labeled carboxyl group added by carboxylation, yet they are all one carbon shorter than the original alkanes.
The removal of two carbon atoms from the parent alkane (Fig. 5, step II) is further supported by the characteristics of the 13C-labeled fatty acids derived from [1,2-13C2]hexadecane. As shown in Table 2, the two isotopmers of the 15:0 fatty acid derived from [1,2-13C2]hexadecane contain either two 13C labels at the alkyl chain terminus or none at all, but not at the carboxyl end. The results suggest that at least two carbon atoms are removed with equal likelihoods from the labeled and unlabeled termini of [1,2-13C2]hexadecane during its metabolism.
As illustrated in Fig. 5, once the first fatty acid (C) is formed, it can be β-oxidized and subsequently mineralized to CO2. These events are demonstrated by the stoichiometry studies reported by Aeckersberg et al. (1, 2). Alternatively, this fatty acid can be further transformed by chain elongation and C-10 methylation (step IV), as evidenced by the following observations. First, the presence of a 13C atom at the C-3 position of the fatty acids which are one carbon longer than their parent alkanes (Table 1) suggests that they are formed after two-carbon elongation of fatty acids with 13C-labeled carboxyl groups. Second, both deuterated 16:0 and 14:0 fatty acids derived from perdeuterated pentadecane carry the same numbers of deuterium labels, while the former fatty acid contains six hydrogen labels at its carboxyl terminus (Table 3), suggesting that it is derived from the latter fatty acid by chain elongation with nondeuterated carbon atoms. Third, as shown in Table 3, the deuterated, 10-methylated fatty acids and the corresponding n-saturated ones contain identical numbers of hydrogen atoms at their carboxyl termini, suggesting that the former are derived from the latter by the addition of a nondeuterated methyl group at C-10. (10-Methylated fatty acids can also be found in some groups of sulfate-reducing bacteria incapable of alkane degradation [41].)
Strain Hxd3 attacks alkanes by carboxylation, apparently at the C-3 position. The C—H bonds on this subterminal, secondary carbon are weaker than those on a terminal, primary carbon and thus can be more easily broken. (For example, the C—H bond dissociation energies for C-1 and C-2 of propane are 94.5 and 98 kcal/mol, respectively [15].) In addition, the selectivity for a subterminal carbon may also be due to the conformational structure at the active site of the carboxylating enzyme, as suggested by studies on monooxygenase systems which attack predominantly the terminal carbon atoms of alkanes (23, 43).
Although biological systems for the carboxylation of aromatic hydrocarbons (31, 49) and other organic compounds (4, 19, 22, 40) have been documented, the direct addition of a carboxyl group enzymatically to an alkane, a nonpolar, saturated, and chemically inert compound (15), has not yet been reported. A recent study on the anaerobic degradation of 2-hexadecanone by a sulfate-reducing enrichment showed that 2-ketone is transformed to pentadecanoic acid apparently by carboxylation at C-3 and the removal of two terminal carbon atoms at C-1 and C-2, a process very similar to what we observed in the present study (18). (The presence of a carbonyl group at C-2 of the ketone may activate the carbon atoms on the alkane to facilitate subterminal carboxylation.) Although the results of our experiment with 2-hexadecanone showed that the compound is not carboxylated to form fatty acids and thus is unlikely to be an intermediate, we do not discount the possibility that the carboxylation reaction is preceded by another initial step(s) which first activate(s) the alkane.
In contrast, studies of strain AK-01, another alkane-degrading, sulfate-reducing bacterium that was isolated from an estuarine sediment (37), indicate a different mechanism of anaerobic alkane degradation (38; So and Young, Abstr. 98th Gen. Meet. Am. Soc. Microbiol.). The initial attack by strain AK-01 also involves subterminal carbon addition to an alkane. The addition reaction, however, occurs at C-2 instead of C-3 and is not effected by carboxylation. Also, concomitant carbon removal from the alkane chain terminus does not occur (38). In other anaerobic cultures, the mechanism of alkane activation has been identified as a fumarate addition reaction that is analogous to the toluene degradation pathway. This mechanism has been observed for a sulfate-reducing enrichment culture growing on n-dodecane (24) and for denitrifying strain HxN1 growing on hexane (32). For HxN1, additional studies have determined some of the subsequent reactions of the pathway (46). Based on earlier observations (38; So and Young, Abstr. 98th Gen. Meet. Am. Soc. Microbiol.), it appears that strain AK-01 utilizes the same fumarate addition mechanism to degrade hexadecane. We have never observed evidence of a fumarate addition product in Hxd3, despite several analyses (data not shown).
In summary, by using stable isotope-labeled compounds, we confirm that alkanes are anaerobically oxidized to fatty acids by strain Hxd3. We demonstrate that C-odd and C-even alkanes are transformed to C-even and C-odd fatty acids, respectively. The initial reactions include subterminal carboxylation of the alkane with the removal of two adjacent terminal carbon atoms. This mechanism of alkane metabolism is distinct from the fumarate addition reaction that has been reported for other anaerobic organisms. These observations indicate that alkanes can be anaerobically degraded through novel and diverse mechanisms that do not require activation by oxygen.
Acknowledgments
We thank Friedrich Widdel for advice on culturing strain Hxd3; Max Häggblom, Xiaoming Zhang, Amy Callaghan, Mike Logan, and Alfred Boyle for technical advice and editorial help; Ingeborg Bossert and Andreas Naef for translation of German literature; and Maria Rivera and Brian Donovan for technical assistance. We are also thankful for the advice of two anonymous reviewers.
This work was supported in part by the Office of Naval Research, the Defense Advanced Research Project Agency, and the National Science Foundation (EMSI).
REFERENCES
- 1.Aeckersberg, F. 1994. Anaerober Abbau von Alkanen und 1-Alkenen durch sulfatreduzierende Bakterien. Ph.D. dissertation. University of Bremen, Bremen, Germany.
- 2.Aeckersberg, F., F. Bak, and F. Widdel. 1991. Anaerobic oxidation of saturated hydrocarbons to CO2 by a new type of sulfate-reducing bacterium. Arch. Microbiol. 156:5-14. [Google Scholar]
- 3.Aeckersberg, F., F. A. Rainey, and F. Widdel. 1998. Growth, natural relationships, cellular fatty acids and metabolic adaptation of sulfate-reducing bacteria that utilize long-chain alkanes under anoxic conditions. Arch. Microbiol. 170:361-369. [DOI] [PubMed] [Google Scholar]
- 4.Birks, S. J., and D. J. Kelly. 1997. Assay and properties of acetone carboxylase, a novel enzyme involved in acetone-dependent growth and CO2 fixation in Rhodobacter capsulatus and other photosynthetic and denitrifying bacteria. Microbiology 143:755-766. [DOI] [PubMed] [Google Scholar]
- 5.Blasig, R., J. Huth, P. Franke, P. Borneleit, W.-H. Schunck, and H.-G. Müller. 1989. Degradation of long-chain n-alkanes by yeast Candida maltosa. III. Effect of solid n-alkanes on cellular fatty acid composition. Appl. Microbiol. Biotechnol. 31:571-576. [Google Scholar]
- 6.Britton, L. N. 1984. Microbial degradation of aliphatic hydrocarbons, p. 89-129. In D. T. Gibson (ed.), Microbial degradation of organic compounds. Marcel Dekker, Inc., New York, N.Y.
- 7.Caldwell, M. E., R. M. Garrett, R. C. Prince, and J. M. Suflita. 1998. Anaerobic biodegradation of long-chain n-alkanes under sulfate-reducing conditions. Environ. Sci. Technol. 32:2191-2195. [Google Scholar]
- 8.Chouteau, J., E. Azoulay, and J. C. Senez. 1962. Anaerobic formation of n-hept-1-ene from n-heptane by resting cells of Pseudomonas aeruginosa. Nature 194:576-578. [Google Scholar]
- 9.Coates, J. D., J. Woodward, J. Allen, P. Philp, and D. R. Lovley. 1997. Anaerobic degradation of polycyclic aromatic hydrocarbons and alkanes in petroleum-contaminated marine harbor sediments. Appl. Environ. Microbiol. 63:3589-3593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Delaune, R. D., G. A. Hambrick III, and W. H. Patrick, Jr. 1980. Degradation of hydrocarbons in oxidized and reduced sediments. Mar. Pollut. Bull. 11:103-106. [Google Scholar]
- 11.Dinh-Nguyen, N., R. Ryhage, S. Ställberg-Stenhagen, and E. Stenhagen. 1961. Mass spectrometric studies. VIII. A study of the fragmentation of normal long-chain methyl esters and hydrocarbons under electron impact with the aid of deuterium substituted compounds. Ark. Kemi 18:393-399. [Google Scholar]
- 12.Doumenq, P., E. Aries, L. Asia, M. Acquaviva, J. Artaud, M. Gilewicz, G. Mille, and J. C. Bertrand. 2001. Influence of n-alkanes on petroleum on fatty acid composition of a hydrocarbonoclastic bacterium: Marinobacter hydrocarbonoclasticus strain 617. Chemosphere 44:519-528. [DOI] [PubMed] [Google Scholar]
- 13.Ehrenreich, P. 1996. Anaerobes Wachstum neuartiger sulfatreduzierender und nitratreduzierender Bakterien auf n-Alkanen und Erd 216 l. Ph.D. dissertation. University of Bremen, Bremen, Germany.
- 14.Ehrenreich, P., A. Behrends, J. Harder, and F. Widdel. 2000. Anaerobic oxidation of alkanes by newly isolated denitrifying bacteria. Arch. Microbiol. 173:58-64. [DOI] [PubMed] [Google Scholar]
- 15.Fessenden, R. J., and J. S. Fessenden. 1982. Organic chemistry. PWS Publishers, Boston, Mass.
- 16.Graff, G., P. Szczepanik, P. D. Klein, J. R. Chipault, and R. T. Holman. 1970. Identification and characterization of fully deuterated fatty acids isolated from Scenedesmus obliquus cultured in deuterium oxide. Lipids 5:786-792. [DOI] [PubMed] [Google Scholar]
- 17.Hamilton, J. T. G., W. C. McRoberts, M. J. Larkin, and D. B. Harper. 1995. Long-chain haloalkanes are incorporated into fatty acids by Rhodococcus rhodochrous NCIMB 13064. Microbiology 141:2611-2617. [Google Scholar]
- 18.Hirschler, A., J. Rontani, D. Raphel, R. Matheron, and J. Bertrand. 1998. Anaerobic degradation of hexadecan-2-one by a microbial enrichment culture under sulfate-reducing conditions. Appl. Environ. Microbiol. 64:1576-1579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Houwen, F. P., C. Dijkema, A. J. M. Stams, and A. J. B. Zehnder. 1991. Propionate metabolism in anaerobic bacteria; determination of carboxylation reactions with 13C-NMR spectroscopy. Biochim. Biophys. Acta 1056:126-132. [Google Scholar]
- 20.IUPAC-IUB Commission on Biochemical Nomenclature. 1978. The nomenclature of lipids. Biochem. J. 171:21-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.King, D. H., and J. J. Perry. 1975. The origin of fatty acids in the hydrocarbon-utilizing microorganism Mycobacterium vaccae. Can. J. Microbiol. 21:85-89. [DOI] [PubMed] [Google Scholar]
- 22.Knowles, J. R. 1989. The mechanism of biotin-dependent enzymes. Annu. Rev. Biochem. 58:195-221. [DOI] [PubMed] [Google Scholar]
- 23.Krevor, J. V. C. 1990. Alkane oxidation: the oxidation of alkanes by group VIII metal ions, p.19-42. In J. A. Davis, P. L. Watson, J. F. Liebman, and A. Greenberg (ed.), Selective hydrocarbon activation. VCH Publishers, New York, N.Y.
- 24.Kropp, K. G., I. A. Davidova, and J. M. Suflita. 2000. Anaerobic oxidation of n-dodecane by an addition reaction in a sulfate-reducing bacterial enrichment culture. Appl. Environ. Microbiol. 66:5393-5398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McCloskey, J. A. 1970. Mass spectrometry of fatty acid derivatives, p. 369-440. In F. D. Gunstone (ed.), Topics in lipid chemistry, vol. I. John Wiley & Sons, Inc., New York, N.Y.
- 26.Muller, F. M. 1957. On methane fermentation of higher alkanes. Antonie Leeuwenhoek 23:369-384. [DOI] [PubMed] [Google Scholar]
- 27.Oldfield, E. 1972. Gas chromatography-mass spectrometry of biosynthetic 1H-2H hybrid fatty acid methyl esters. J. Chem. Soc. Chem. Commun. 1972:719. [Google Scholar]
- 28.Oldfield, E., D. Chapman, and W. Derbyshire. 1972. Lipid mobility in Acholeplasma membranes using deuteron magnetic resonance. Chem. Phys. Lipids 9:69-81. [DOI] [PubMed] [Google Scholar]
- 29.Parekh, V. R., R. W. Traxler, and J. M. Sobek. 1977. n-Alkane oxidation enzymes of a pseudomonad. Appl. Environ. Microbiol. 33:881-884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Phelps, C. D., X. Zhang, and L. Y. Young. 2001. Use of stable isotopes to identify benzoate as a metabolite of benzene degradation in a sulfidogenic consortium. Environ. Microbiol. 3:600-603. [DOI] [PubMed] [Google Scholar]
- 31.Rabus, R., and J. Heider. 1998. Initial reactions of anaerobic metabolism of alkylbenzenes in denitrifying and sulfate-reducing bacteria. Arch. Microbiol. 170:377-384. [Google Scholar]
- 32.Rabus, R., H. Wilkes, A. Behrends, A. Armstroff, T. Fischer, A. J. Pierik, and F. Widdel. 2001. Anaerobic initial reaction of n-alkanes in a denitrifying bacterium: evidence for (1-methylpentyl)succinate as initial product and for involvement of an organic radical in n-hexane metabolism. J. Bacteriol. 183:1707-1715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rock, C. O., S. Jackowski, and J. E. Cronan, Jr. 1996. Lipid metabolism in prokaryotes, p. 35-74. In D. E. Vance and J. E. Vance (ed.), Biochemistry of lipids, lipoproteins and membranes. Elsevier Science B.V., Amsterdam, The Netherlands.
- 34.Rosenfeld, W. D. 1947. Anaerobic oxidation of hydrocarbons by sulfate-reducing bacteria. J. Bacteriol. 54:664-665. [PMC free article] [PubMed] [Google Scholar]
- 35.Rueter, P., R. Rabus, H. Wilkes, F. Aeckersberg, F. A. Rainey, H. W. Jannasch, and F. Widdel. 1994. Anaerobic oxidation of hydrocarbons in crude oil by new types of sulfate-reducing bacteria. Nature 372:455-458. [DOI] [PubMed] [Google Scholar]
- 36.Ryhage, R., and E. Stenhagen. 1960. Mass spectrometric studies. IV. Esters of monomethyl-substituted long chain carboxylic acids. Ark. Kemi 15:291-304. [Google Scholar]
- 37.So, C. M., and L. Y. Young. 1999. Isolation and characterization of a sulfate-reducing bacterium that anaerobically degrades alkanes. Appl. Environ. Microbiol. 65:2969-2976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.So, C. M., and L. Y. Young. 1999. Initial reactions in anaerobic alkane degradation by a sulfate reducer, strain AK-01. Appl. Environ. Microbiol. 65:5532-5540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sullivan, E. R., C. Phelps, and L. Y. Young. 2001. Anaerobic mineralization of stable isotope-labeled 2-methylnaphthalene. Appl. Environ. Microbiol. 67:4353-4357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Suttie, J. W. 1985. Vitamin K-dependent carboxylase. Annu. Rev. Biochem. 54:459-477. [DOI] [PubMed] [Google Scholar]
- 41.Vainshtein, M., H. Hippe, and R. M. Kroppenstedt. 1992. Cellular fatty acid composition of Desulfovibrio species and its use in classification of sulfate-reducing bacteria. Syst. Appl. Microbiol. 15:554-566. [Google Scholar]
- 42.van Beilen, J. B., M. G. Wubbolts, and B. Witholt. 1994. Genetics of alkane oxidation by Pseudomonas oleovorans. Biodegradation 5:161-174. [DOI] [PubMed] [Google Scholar]
- 43.Watkinson, R. J., and P. Morgan. 1990. Physiology of aliphatic hydrocarbon-degrading microorganisms. Biodegradation 1:79-92. [DOI] [PubMed] [Google Scholar]
- 44.Widdel, F., and F. Bak. 1992. Gram-negative mesophilic sulfate-reducing bacteria, p. 3352-3378. In A. Balows, H. G. Trüper, M. Dworkin, W. Harder, and K.-H. Schleifer (ed.), The prokaryotes, vol. IV. Springer-Verlag, New York, N.Y.
- 45.Widdel, F., and R. Rabus. 2001. Anaerobic biodegradation of saturated and aromatic hydrocarbons. Curr. Opin. Biotechnol. 12:259-276. [DOI] [PubMed] [Google Scholar]
- 46.Wilkes, H., R. Rabus, T. Fischer, A. Armstroff, A. Behrends, and F. Widdel. 2002. Anaerobic degradation of n-hexane in a denitrifying bacterium: further degradation of the initial intermediate (1-methylpentyl)succinate via C-skeleton rearrangement. Arch. Microbiol. 177:235-243. [DOI] [PubMed] [Google Scholar]
- 47.Zengler, K., H. H. Richnow, R. Rosselló-Mora, W. Michaelis, and F. Widdel. 1999. Methane formation from long-chain alkanes by anaerobic microorganisms. Nature 401:266-269. [DOI] [PubMed] [Google Scholar]
- 48.Zhang, X., E. R. Sullivan, and L. Y. Young. 2001. Aromatic ring reduction in the biodegradation of carboxylated naphthalene by a sulfate-reducing consortium. Biodegradation 11:117-124. [DOI] [PubMed] [Google Scholar]
- 49.Zhang, X. M., and L. Y. Young. 1997. Carboxylation as an initial reaction in the anaerobic metabolism of naphthalene and phenanthrene by sulfidogenic consortia. Appl. Environ. Microbiol. 63:4759-4764. [DOI] [PMC free article] [PubMed] [Google Scholar]