

Abstract
To understand the origins and function of the human germ cell lineage and to identify germ cell-specific markers we have isolated a human ortholog of the Drosophila gene vasa. The gene was mapped to human chromosome 5q (near the centromere) by radiation hybrid mapping. We show by Northern analysis of fetal and adult tissues that expression of the human VASA gene is restricted to the ovary and testis and is undetectable in somatic tissues. We generated polyclonal antibodies that bind to the VASA protein in formalin-fixed, paraffin-embedded tissue and characterized VASA protein expression in human germ cells at various stages of development. The VASA protein is cytoplasmic and expressed in migratory primordial germ cells in the region of the gonadal ridge. VASA protein is present in fetal and adult gonadal germ cells in both males and females and is most abundant in spermatocytes and mature oocytes. The gene we have isolated is thus a highly specific marker of germ cells and should be useful for studies of human germ cell determination and function.
Keywords: gametogenesis, DEAD box RNA helicase
Germ cells in most animal species undergo two complex and remarkably different developmental phases. In the first phase, which occurs during early embryogenesis, primordial germ cells (PGCs) are formed and then actively migrate to gonadal anlage believed to contain all of the somatic components of the mature gonad. In the second phase, the germ cells sense appropriate cues from their environment and initiate one of two distinct programs of controlled cell division, meiosis and differentiation-oogenesis or spermatogenesis, to form gametes. The molecular basis of both gametogenesis and early germ cell development is best understood in Drosophila and Caenorhabditis elegans, where systematic genetic screens have identified many of the required genes. Studies in these organisms have shown that the early determination and formation of germ cells is controlled by genes expressed by the mother during oogenesis, whereas later steps depend on genes expressed in the embryo (reviewed in refs. 1 and 2).
The Drosophila gene vasa has a central role in several aspects of germ cell development. It encodes a member of the DEAD (Asp-Glu-Ala-Asp) box family of ATP-dependent RNA helicases (3, 4), and the protein exhibits ATP-dependent RNA helicase activity in vitro (5). The vasa protein appears to regulate the translation of multiple downstream mRNAs and has been shown to interact with a Drosophila homologue of the yeast translation initiation factor 2 (6). Vasa protein is an essential component of germ plasm, a poorly understood complex of RNA and proteins that is required for germ cell determination (2). A null mutation that removes the entire vasa coding region results in female sterility with severe defects in oogenesis, including abnormal germ-line differentiation and oocyte determination (5). Females homozygous for partial loss-of-function alleles produce eggs that can be fertilized, but the resulting embryos lack germ cells (7). Therefore, vasa function is not only required during gametogenesis in the adult but is also essential for the specification of the germ cell lineage during embryogenesis. The vasa protein is localized during Drosophila oogenesis to the posterior of the oocyte (where germ cells will form) and becomes concentrated within early germ cells in the embryo. By gastrulation, vasa protein is detectable only in germ cells, and strong cytoplasmic staining in germ cells is maintained during embryogenesis in both male and females (8).
The vasa gene is conserved in invertebrate and vertebrate species, including C. elegans, Xenopus, zebrafish, mouse, and rat (9–14). Three vasa-like genes have been identified in C. elegans (9) whereas in Drosophila and all vertebrate species to date only one vasa ortholog has been identified. In all species, members of the vasa subfamily are specifically expressed in germ cells throughout development, and in C. elegans and Xenopus they are required for germ cell specification. In zebrafish, studies of vasa have allowed tracing of the germ cell lineage earlier in development than formerly possible. The RNA exhibits a striking and unexpected pattern of localization to the cleavage planes in two- and four-cell embryos and is localized to exactly four putative PGCs at the 32-cell stage (11, 12). Thus, in zebrafish as in Drosophila, vasa appears to be a specific marker of germ cells at their earliest stages of development, and its pattern of expression is consistent with a determinative role in germ cell formation. Mouse and rat vasa genes specifically expressed in the germ cell lineage also have been identified (13, 14), and mice with a targeted mutation of the vasa locus Mvh recently have been described (15).
Here we describe the cloning and characterization of the human VASA gene. The predicted protein is similar throughout its length to previously identified members of the VASA subfamily. We show by Northern analysis and immunohistochemistry that the gene is specifically expressed in germ cells in both sexes, consistent with an evolutionarily conserved role in germ cell development. VASA gene products (both mRNA and protein) should serve as valuable reagents for studies of the human germ cell lineage.
Materials and Methods
Cloning of a Human VASA cDNA.
The primer (oVAS2a) for 5′ rapid amplification of cDNA ends was 5′ CTC TGC ATC AAA ACC ACA GAC TTG AAG G 3′ (28-mer). Marathon human testis cDNA (CLONTECH) was used as a template per the manufacturer's instructions except that the high-fidelity LA Taq DNA polymerase (Takara Shuzo, Kyoto) was used. Parameters for PCR were: 35 cycles of denaturation at 94o for 30 sec, annealing at 56o for 30 sec, and extension at 72o for 4 min. Ten picomoles of oVAS2a and the 5′ rapid amplification of cDNA ends AP1 primer (CLONTECH) were used in a 50-μl reaction. The single discrete 2.2-kb product was gel-purified, subcloned, and sequenced on both strands by primer walking.
Radiation Hybrid Mapping of VASA.
The genebridge 4 human genome radiation hybrid mapping panel (Research Genetics, Huntsville, AL) was screened by PCR, with DNA from the parental hamster cell line 4A23 (Research Genetics) as a negative control and normal human genomic DNA as a positive control. PCRs (50 μl) contained 20 mM Tris⋅HCl (pH 8.4), 50 mM KCl, 1.5 mM MgCl2, 0.2 mM dNTP, 0.2 mM of the VASA primers oVAS3 and oVAS4, and 0.02 unit/μl Taq DNA polymerase (Life Technologies, Gaithersburg, MD). The primer sequences were 5′-ACA GGA TGT TCC TGC ATG GT-3′ (oVAS3, 20-mer) and 5′-TGC CCT TTC TGG TAT CAA CTG-3′ (oVAS4, 20-mer). The PCR protocol entailed 40 cycles of denaturation at 94o for 30 sec, annealing at 52o for 30 sec, and extension at 72o for 1 min. Ten microliters of each PCR was electrophoresed in 2% agarose gels. A reaction was scored positive if a band at 110 bp was visualized by ethidium staining. Results were submitted to the radiation hybrid mapping server at the Whitehead Institute/Massachusetts Institute of Technology Center for Genome Research (http://www-genome.wi.mit.edu/cgi-bin/contig/rhmapper.pl).
Collection of Tissue Samples, RNA Preparation, and Northern Analysis.
Permission for use of discarded tissues was granted by an institutional review board according to local ethical guidelines. Gestational age (equal to the developmental age plus 2 weeks) was estimated based on clinical parameters including last menses and crown-rump length. For RNA preparation, tissues were collected and stored at −70°C. Total RNA was extracted with the Trizol reagent (Life Technologies) per the manufacturer's instructions, subjected to formaldehyde denaturing gel electrophoresis (1.3% agarose), and transferred to a membrane (Micron Separations). A Northern blot of adult poly(A)+ selected RNA (CLONTECH) was hybridized with a VASA cDNA probe labeled by the random primer method. Washes were performed under high stringency conditions (0.1× SSC, 0.1% SDS at 65o for 30 min × 2).
Antibody Generation and Immunohistochemistry.
A region of the VASA protein likely to be antigenic was identified by using the PeptideStructure program (University of Wisconsin Genetics Computer Group). A 30-mer peptide (DHC2) was synthesized and coupled to keyhole limpet hemocyanin. A rabbit was immunized according to standard protocols. Specific antibodies were purified by chromatography using peptide-conjugated Sepharose beads. Sections of paraffin-embedded tissues were dewaxed with xylene. Antigen retrieval was performed by microwave treatment (95o for 30 min) in sodium citrate buffer. Affinity-purified VASA antibodies were used at 1:1,000 to 1:10,000 dilution. A c-kit mAb (Dako) was used at a dilution of 1:150 to 1:300. The EnVision detection system (Dako) was used per the manufacturer's instructions, with the chromogen diaminobenzidine. Sections were counterstained with hematoxylin.
Results
Cloning of a Human VASA cDNA.
Human expressed sequence tags (ESTs) corresponding to the human VASA gene (Genbank accession nos. AA399611, AA398976, and AI217144) were identified with the tblastn program (http://www.ncbi.nlm.nih.gov/BLAST) by using the complete mouse vasa protein sequence (Genbank accession no. 2500525) as the query. These ESTs contained the 3′ untranslated end of the cDNA and encoded a portion of the protein C terminus. They appeared to represent bona fide human VASA ESTs based on the extent of protein sequence similarity and that all were derived from human testis cDNA libraries. To obtain the entire VASA coding sequence, an oligonucleotide corresponding to the 3′ untranslated region was used in a 5′ rapid amplification of cDNA ends reaction with an adult human testis cDNA source. A single 2.2-kb product corresponding to a human VASA cDNA was obtained. This cDNA encodes the full-length protein, based on sequence alignments that show a similar N terminus to other vasa subfamily members (Fig. 1).
Figure 1.

Alignment of VASA protein sequences from human and major experimental organisms, including Mus musculus (Genbank accession no. 2500525), Rattus norvegicus (Genbank accession no. 2500526), Xenopus laevis (Genbank accession no. 2896107), Danio reio (Genbank accession no. 2558535), and Drosophila melanogaster (Genbank accession no. 1352826). Residues conserved in ≥ 50% of the sequences are highlighted in black. The eight conserved homology boxes present in all DEAD box proteins are indicated by I–VIII. * indicate the four copies of the RGG motif in the human protein; all VASA proteins contain multiple copies of this motif in their N termini. A bar overlies the region corresponding to the synthetic peptide used for antibody generation.
Comparison of the Predicted Human VASA Protein Sequence with Other Species.
The cDNA encodes a predicted protein of 724 aa and a molecular mass of 79 kDa. Eight conserved domains (I–VIII) directly involved in RNA helicase activity have been identified in all DEAD box proteins to date (16). All eight of these domains, including domain V, which contains the DEAD motif, are present in the human VASA protein (Fig. 1). The 302-aa sequence encompassing this conserved core region (domains I–VIII including intervening sequences) shows greater similarity to VASA homologues in other species (9–14) than to other known human DEAD box proteins: rat (94% identity), mouse (90%), zebrafish (76%), Xenopus (72%), and Drosophila (56%). The human DEAD box protein exhibiting greatest similarity in this core region was the Y-chromosome encoded DBY, which had lower similarity (70% vs. 75%) and greater overall gap size (4% vs. 2%) than the Drosophila vasa gene. The DBY gene is ubiquitously expressed (17), further arguing it is not a VASA ortholog. Alignment of the VASA protein sequences demonstrates that significant similarity is not confined to the core region (Fig. 1). In contrast, other DEAD box proteins have significant length variation and little or no sequence conservation at their amino and carboxyl termini. The human protein shares with other VASA-related proteins an N terminus rich in glycine, with multiple repeats of an RGG motif (Fig. 1) believed to function in RNA binding (16). Based on these conserved sequence features, we conclude we have isolated a human ortholog of the Drosophila vasa gene.
Mapping of the Human VASA locus.
Radiation hybrid mapping placed VASA in the interval between WI-3071 and AFM138YB8 on chromosome 5. The distance between VASA and WI-3071 was estimated to be 4.71 cR3000 (1 cR3000 ≅ 270 kb), with a logarithm of odds score of 2.00. Fluorescent in situ hybridization using a bacterial artificial chromosome probe spanning the human VASA locus showed hybridization to chromosome 5 band q11.2–12 (data not shown), consistent with the results of radiation hybrid mapping. Notably, this chromosomal region is syntenic to the distal end of mouse chromosome 13, where the mouse vasa homologue Ddx4 resides (18). A search of the Online Mendelian Inheritance in Man database (http://www3.ncbi.nlm.nih.gov/omim/) did not reveal any hereditary infertility syndromes caused by mutations in this chromosomal region.
Human VASA Gene Expression Is Confined to the Testis and Ovary.
VASA gene expression was evaluated in fetal and adult tissues (Fig. 2). A Northern blot of total RNA prepared from tissues obtained at an estimated gestational age of 18 weeks was probed with the VASA cDNA. The predominant VASA transcript of 2.6 kb was present only in testis and ovary. A faint band of greater than 4.0 kb and a high molecular weight smear observed in testis and ovary lanes likely correspond to alternative or incompletely processed transcripts (Fig. 2A). No signals were detected in nongonadal tissues.
Figure 2.
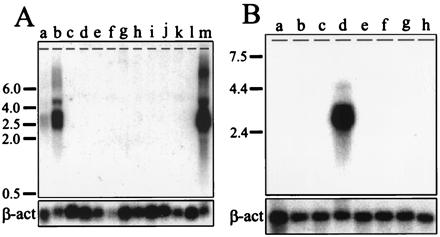
(A) Northern analysis of VASA in fetal tissues (lanes a, testis; b, ovary; c, brain; d, thymus; e, lung; f, liver; g, spleen; h, pancreas; i, small intestine, colon, and stomach; j, kidney; k, adrenal, and l, placenta) and adult testis (lane m). The positions of size standards are indicated on the left. Approximately 5 μg of total RNA was loaded per lane. The blot was reprobed with a human β-actin probe to confirm RNA loading. (B) Northern analysis of VASA in adult tissues: lanes a, spleen; b, thymus; c, prostate; d, testis; e, ovary; f, small intestine; g, colon; and h, peripheral blood leukocytes. Each lane was loaded with 2.5 μg of poly(A)+ RNA from pooled specimens. The ovarian sample was prepared from ovaries of pre- and postmenopausal patients.
In adults, VASA expression was high in the testis and undetectable in nongonadal tissues (Fig. 2). Absence of signal in the ovary lane (Fig. 2B) was likely caused by the low number of oocytes in the adult ovaries from which the RNA was derived. However, as discussed below, VASA protein was easily detectable in the cytoplasm of oocytes in adult ovaries by immunohistochemistry. In summary, VASA expression is gonad-specific in the fetus and adult, consistent with germ cell-specific expression.
Generation of Antibodies and Immunohistochemical Analysis of VASA.
To develop a reagent useful for in situ analyses of human germ cells, antibodies were generated to the VASA protein. Antigen-retrieval conditions were optimized for archival paraffin-embedded, formalin-fixed specimens. Control preimmune serum resulted in no signal in tissue sections of an adult human testis (Fig. 3A), whereas postimmune, affinity-purified antiserum gave a strong reaction in the seminiferous tubules (Fig. 3B). The VASA protein was present in spermatogonia, spermatocytes, and spermatids. Spermatogonia exhibited variable VASA staining, with contiguous groups along the basement membrane showing either weak or intermediate staining (Fig. 3B, arrows and arrowheads). These two groups may correspond to type A and B spermatogonia. Sertoli cells, which serve an accessory function within the seminiferous tubules, and which were seen as a layer immediately above the spermatogonia, exhibited no staining (Fig. 3B, blue arrow). VASA expression was highest in spermatocytes, which had strong cytoplasmic staining that was somewhat granular in appearance. Spermatids also exhibited cytoplasmic staining. Spermatozoa were negative for VASA (Fig. 3B), and no significant staining was observed in sections through epididymis containing numerous spermatozoa (data not shown). No signal was observed in a testicular biopsy from an infertile patient with Sertoli cell only syndrome (Fig. 3C), a condition of varied etiology characterized by azoospermia and the complete absence of germ cells, further confirmation of VASA germ cell-specific expression. In the ovary of a premenopausal female, strong cytoplasmic staining was seen in oocytes (Fig. 3D).
Figure 3.
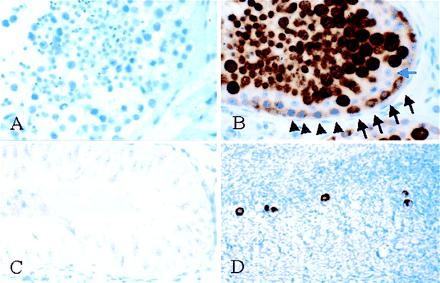
Expression of VASA in adult gonads. (A) Testis, preimmune control. No immunoreactivity was observed. (B) Testis, VASA immunostaining. Strong signal (brown stain) was observed within the seminiferous tubules. The spermatogonia, directly in contact with the basement membrane, exhibited either weak (arrowheads) or intermediate staining (arrows). The Sertoli cells (blue arrow) formed a distinct layer directly above the spermatogonia and showed lack of immunoreactivity. Spermatocytes (large cells) and spermatids (small cells) above the Sertoli layer exhibited strong immunoreactivity. Scattered spermatozoa, identifiable by their small, darkly staining nuclei, were negative for VASA. (C) Testicular biopsy from infertile patient with azoospermia and complete absence of germ cells (Sertoli cell only syndrome). No VASA immunoreactivity was observed. (D) Ovary of premenopausal female. The scattered oocytes in the ovarian cortex were intensely VASA-positive.
Characterization of VASA Expression in Early Development.
We examined the gonadal ridge, a thickened region of mesothelium and underlying mesenchyme that gives rise to the gonad and is adjacent to the mesonephros (part of the primitive excretory system which later degenerates). The interval spanning the arrival of PGCs corresponds to a gestational age of 6–7 weeks (19, 20). At 6 weeks, no VASA-positive cells were identified (Fig. 4A). In contrast, at 7 weeks the mesenchyme of the gonadal ridge was populated by numerous VASA-positive PGCs (Fig. 4 B and C), some of which were present at the coelomic angle apparently migrating toward the gonadal ridge (Fig. 4B). A number of the cells had distinct pseudopodia (Fig. 4C). VASA-positive cells, some undergoing mitosis, also were observed in the gut mesenchyme (Fig. 4D). This is consistent with prior observations that human PGCs increase in number through mitotic activity during their migration to the gonadal ridge (19). Numerous VASA-positive presumptive PGCs also were present at the mesenteric root (data not shown). Based on their morphology and distribution, we conclude that these VASA-positive cells are PGCs. This was further confirmed by immunostaining with c-kit, a membrane receptor expressed in the germ cell lineage, where it regulates migration, proliferation, and apoptosis (21). C-kit immunostaining was observed concurrent with the arrival of PGCs at the gonadal ridge (Fig. 4E and data not shown).
Figure 4.
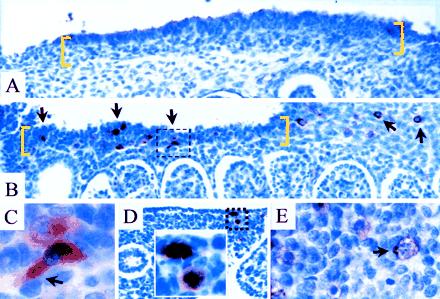
Study of VASA-positive PGCs in the region of the gonadal ridge (sagittal sections). (A) Six weeks (estimated gestational age) before arrival of PGCs at gonadal ridge (bracketed in yellow). No VASA-positive cells were identified. (B–F) Seven weeks during arrival of PGCs at the gonadal ridge. (B) Strongly VASA-positive cells were present in the gonadal ridge (multiple arrows) and also in the mesenchyme at the coelomic angle, apparently migrating toward the ridge (upper right hand corner). A box indicates the area magnified in C. (C) High magnification of a PGC with distinct pseudopodium (arrow). A second VASA-positive PGC, slightly out of focus, is above. (D) Primitive gut. VASA-positive cells, one of which was undergoing mitosis in the gut mesenchyme. (E) C-kit receptor immunoreactivity. Membrane-associated signal was observed in presumptive PGCs; area corresponds to the upper right hand corner in B. A distinct pseudopodium points in the direction of the gonadal ridge (arrow).
VASA Expression in the Early Fetal Gonad.
VASA expression was evaluated at later, sexually dimorphic stages of gonad development. At 17 weeks, ovaries contain many cortical germ cells, larger maturing oocytes, and primordial follicles. All germ cells were VASA-positive, with the undifferentiated cortical cells exhibiting less staining than the more mature forms (Fig. 5A). Testes at 17 weeks contain 1–6 VASA-positive cells per seminiferous tubule cross-section (Fig. 5B), consistent with prior estimates of the number of germ cells in the testis at this developmental stage (20). The greater proportion of germ cells in the fetal ovary relative to the fetal testis paralleled the higher total ovarian VASA RNA levels at a similar stage (Fig. 2A).
Figure 5.
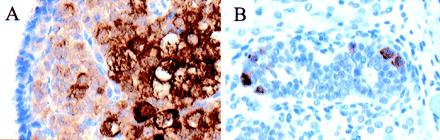
VASA expression in fetal gonads. (A) 17-week (estimated gestational age) ovary. VASA immunostaining was observed in undifferentiated cortical germ cells and maturing oocytes. (B) 17-week testis. Only scattered VASA-positive germ cells were present in the seminiferous tubules at this developmental stage.
Localization of VASA Protein in Oocytes.
At 35 weeks, the ovary consists predominantly of primordial follicles comprised of oocytes surrounded by a single layer of granulosa cells. Many of the oocytes exhibited a striking accentuation of VASA staining within a compact perinuclear body localized to the medial aspect of the eccentrically located oocyte nucleus (Fig. 6A). The location of this body corresponds to the so-called “yolk nucleus” of Balbiani's vitelline body, described ultrastructurally in detail by Hertig and Adams (22) in human primordial follicles. Immunostaining of this body was not observed with preimmune sera or antisera to other ovarian antigens such as inhibin and depended on the presence of primary antibody (data not shown). In adult oocytes observed at high magnification, including the maturing antral follicle shown in Fig. 6B, VASA immunostaining was homogenous throughout the ooplasm, with no distinct subcellular localization.
Figure 6.
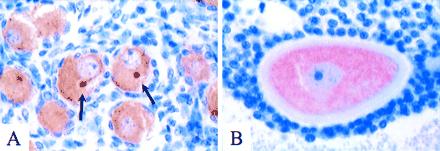
VASA protein localization in fetal and adult ovaries. (A) 35-week (estimated gestational age) ovary with numerous primordial follicles. The oocytes contained VASA-positive cytoplasm. In many oocytes (two in the center of this field), a distinct compact perinuclear structure exhibited accentuated VASA immunostaining (arrows). This structure was always round, of uniform size, and centrally located relative to the nucleus. (B) Maturing antral follicle in adult ovary, consisting of large oocyte with prominent zona pellucida and surrounding granulosa cells. The ooplasm was diffusely VASA-positive, with no distinct subcellular localization.
Discussion
We have shown that in humans, as in other species, VASA is specifically expressed in the germ cell lineage in both sexes. Other markers of the germ cell lineage include Oct-4, a member of the POU family of transcription factors and believed to be a key regulator of totipotency (23, 24). Oct-4 is expressed throughout the preimplantation mouse embryo, later becoming restricted to the germ line. As such, it appears to be a marker of totipotent cells rather than the germ cell lineage per se. Alkaline phosphatase, a marker used in classic studies of germ cell migration, is not reliably expressed in postmigratory germ cells and is not specific for the germ cell lineage (25). Many genes are expressed either during spermatogenesis or oogenesis, such as the mouse DEAD box PL10 gene, which is expressed only in the male germ line and appears to be involved in translational control during spermatogenesis (26). VASA is so far unique in its complete specificity for the germ cell lineage; indeed, in most species VASA expression appears to equate with germ cell fate.
Human homologues exist for several other genes required for germ cell determination in Drosophila. Rodent and human homologues of staufen have been implicated in microtubule-dependent mRNA transport in neuronal dendrites (27, 28). Two distinct cDNAs (Kazusa cDNA project accession nos. D87078 and D43951) with extensive similarity to pumilio have been obtained from a human myeloblast library; both genes are ubiquitously expressed but their biological functions remain unknown (http://www.kazusa.or.jp/huge). Several human proteins contain regions similar to a domain found in multiple copies in tudor (e.g., refs. 29 and 30). None of these human genes has been implicated in embryonic patterning or specification of the germ cell lineage. Mammalian counterparts of nanos, oskar, exuperantia, or valois have not been identified.
The specific expression of VASA in the germ cell lineage during colonization of the gonadal ridge and later in development including gametogenesis suggests that VASA is required for germ cell maintenance and function. Mutational analyses have confirmed such roles in Drosophila and C. elegans. Intriguingly, male mice homozygous for a targeted mutation of the mouse vasa ortholog Mvh are sterile and exhibit severe defects in spermatogenesis, whereas homozygous females are fertile. The lack of an observable oogenesis phenotype may be caused by functional redundancy with other DEAD box family members, although there is no evidence for additional VASA family members in the mouse genome (15). The identification of human VASA gene mutations would be of interest; such mutations also would likely result in male or female sterility.
Morphologically distinct PGCs in mouse and human embryos are first detectable in the yolk sac endoderm at the time of gastrulation, and no germ plasm has been identified in fertilized mouse zygotes that would permit tracing of earlier germ-line precursors. Thus, the germ cell lineage appears to be discontinuous in mammals, although the mechanisms of PGC formation remain obscure. Mice with a targeted (presumed null) Mvh mutation do form PGCs (15). Although these PGCs exhibit decreased proliferative activity and other defects, these findings suggest that VASA is not essential for germ cell specification in mammals (15).
We detected VASA-positive presumptive germ cells in extragonadal locations, including the gut and mesenteric root. In contrast, expression of the mouse Mvh gene was not detected in migratory PGCs before their arrival at the gonadal ridge, but rather was induced after direct interaction with gonadal somatic cells (31). Although the extragonadal VASA-positive cells we observed could be caused by nonspecific cross-reactivity, we do not believe this is the case because the morphology and limited anatomic distribution of these VASA-positive cells is entirely consistent with PGCs. There may be species-specific differences in the precise timing or mechanisms regulating VASA expression.
The increased immunostaining of VASA in a distinct cytoplasmic structure in fetal oocytes is of uncertain significance, and it was not observed in adult oocytes. However, the size and location of this body correspond to the yolk nucleus, a tight aggregate of membranes and vesicles within Balbiani's vitelline body, a poorly understood cytoplasmic complex (including a dense cloud of mitochondria, lamellar aggregates, and electron-dense material) encompassing most of the ooplasm in species ranging from spiders to humans (22, 32). The electron-dense material, which is intimately associated with and typically surrounds the yolk nucleus, corresponds to nuage, which in many species is asymmetrically distributed during embryogenesis and appears to play a determinative role in germ cell formation, whereby cells that inherit germ plasm are destined to become germ cells (1, 2). A structure analogous to the yolk nucleus, termed the sponge body, has been described recently in Drosophila, and proposed to be involved in the assembly and transport of mRNAs such as bicoid in oocytes (33). The apparent localization of VASA protein to an analogous structure in human oocytes is intriguing in light of the fact that VASA, an RNA-binding protein, might be involved in assembly and transport of mRNAs during oogenesis.
In mouse ovaries, Mvh staining is diffusely cytoplasmic in the oocytes of primary follicles but absent in maturing antral follicles (31), whereas we observed decreased but readily detectable VASA staining in human antral follicles (the decrease is likely caused by dilution as the oocyte dramatically increases in volume). No distinct VASA localization was detected in adult mouse oocytes, similar to our findings; VASA localization in mouse fetal ovaries has not been reported. In spermatogenesis, Mvh protein was found to be localized predominantly to a single large cytoplasmic body corresponding to the chromatoid body or nuage, whereas we have not observed localized VASA staining in spermatids. These staining patterns may reflect genuine interspecies differences in protein localization or unique properties of antibody preparations. Although specific expression of VASA in the germ cell lineage appears to be conserved throughout phylogeny, it will be of interest to further explore the differing roles of VASA in germ cell function in mammalian and nonmammalian species.
Acknowledgments
We thank Jonathan Fletcher and members of his laboratory for sharing equipment and supplies. We thank Mel Feany, Todd Kroll, Ivan Moskowitz, Marcus Bosenberg, Larry Saubermann, Dennis McKearin, and Paul Lasko for discussions and technical advice. We acknowledge principal financial support through the Stanley L. Robbins 1999 Research Award (Department of Pathology, Brigham and Women's Hospital) and a Bridge Award (Harvard Medical School).
Abbreviation
- PGC
primordial germ cell
Footnotes
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. AY004154).
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.160274797.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.160274797
References
- 1.Wylie C. Cell. 1999;96:165–174. doi: 10.1016/s0092-8674(00)80557-7. [DOI] [PubMed] [Google Scholar]
- 2.Saffman E E, Lasko P. Cell Mol Life Sci. 1999;55:1141–1163. doi: 10.1007/s000180050363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lasko P F, Ashburner M. Nature (London) 1988;335:611–617. doi: 10.1038/335611a0. [DOI] [PubMed] [Google Scholar]
- 4.Hay B, Jan L Y, Jan Y N. Cell. 1988;55:577–587. doi: 10.1016/0092-8674(88)90216-4. [DOI] [PubMed] [Google Scholar]
- 5.Styhler S, Nakamura A, Swan A, Suter B, Lasko P. Development (Cambridge, UK) 1998;125:1569–1578. doi: 10.1242/dev.125.9.1569. [DOI] [PubMed] [Google Scholar]
- 6.Carrera P, Johnstone O, Nakamura A, Casanova J, Jackle H, Lasko P. Mol Cell. 2000;5:181–187. doi: 10.1016/s1097-2765(00)80414-1. [DOI] [PubMed] [Google Scholar]
- 7.Schupbach T, Wieschaus E. Roux Arch Dev Biol. 1986;195:302–317. doi: 10.1007/BF00376063. [DOI] [PubMed] [Google Scholar]
- 8.Lasko P F, Ashburner M. Genes Dev. 1990;4:905–921. doi: 10.1101/gad.4.6.905. [DOI] [PubMed] [Google Scholar]
- 9.Gruidl M E, Smith P A, Kuznucki K A, McCrone J S, Kirchner J, Rousell D L. Proc Natl Acad Sci USA. 1996;93:13837–13842. doi: 10.1073/pnas.93.24.13837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ikenishi K, Tanaka T S. Dev Growth Differ. 1997;39:625–633. doi: 10.1046/j.1440-169x.1997.t01-4-00010.x. [DOI] [PubMed] [Google Scholar]
- 11.Yoon C, Kawakami K, Hopkins N. Development (Cambridge, UK) 1997;125:3157–3166. doi: 10.1242/dev.124.16.3157. [DOI] [PubMed] [Google Scholar]
- 12.Olsen L C, Aasland R, Anders F. Mech Dev. 1997;66:95–105. doi: 10.1016/s0925-4773(97)00099-3. [DOI] [PubMed] [Google Scholar]
- 13.Fujiwara Y, Komiya T, Kawabata H, Sato M, Fujimoto H, Furusawa M, Noce T. Proc Natl Acad Sci USA. 1994;91:12258–12262. doi: 10.1073/pnas.91.25.12258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Komiya T, Tanigawa Y. Biochem Biophys Res Commun. 1995;207:405–410. doi: 10.1006/bbrc.1995.1202. [DOI] [PubMed] [Google Scholar]
- 15.Tanaka S S, Toyooka Y, Akasu R, Katoh-Fukui Y, Nakahara Y, Suzuki R, Yokoyama M, Noce T. Genes Dev. 2000;14:841–853. [PMC free article] [PubMed] [Google Scholar]
- 16.Lüking A, Stahl U, Schmidt U. Crit Rev Biochem Mol Biol. 1998;33:259–296. doi: 10.1080/10409239891204233. [DOI] [PubMed] [Google Scholar]
- 17.Lahn B T, Page D C. Science. 1997;278:675–680. doi: 10.1126/science.278.5338.675. [DOI] [PubMed] [Google Scholar]
- 18.Abe K, Noce T. Mamm Genome. 1997;8:622–623. doi: 10.1007/s003359900521. [DOI] [PubMed] [Google Scholar]
- 19.Witschi E. Carnegie Inst Washington Publ. 1948;209:67–82. [Google Scholar]
- 20.van Wagenen G, Simpson M E. Embryology of the Ovary and Testis Homo sapiens and Macaca mulatta. New Haven: Yale Univ. Press; 1965. pp. 47–55. [Google Scholar]
- 21.Ashman L K. Int J Biochem Cell Biol. 1999;31:1037–1051. doi: 10.1016/s1357-2725(99)00076-x. [DOI] [PubMed] [Google Scholar]
- 22.Hertig A, Adams E C. J Cell Biol. 1967;34:647–675. doi: 10.1083/jcb.34.2.647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nichols J, Zevnik B, Anastassiadis K, Niwa H, Klewe-Nebenius D, Chambers I, Scholer H, Smith A. Cell. 1998;95:379–391. doi: 10.1016/s0092-8674(00)81769-9. [DOI] [PubMed] [Google Scholar]
- 24.Scholer H R, Ruppert S, Suzuki N, Chowdhury K, Gruss P. Nature (London) 1990;344:435–439. doi: 10.1038/344435a0. [DOI] [PubMed] [Google Scholar]
- 25.Hustin J, Colette J, Franchimont P. Int J Androl. 1987;10:29–35. doi: 10.1111/j.1365-2605.1987.tb00162.x. [DOI] [PubMed] [Google Scholar]
- 26.Leroy P, Alzari P, Sassoon D, Wolgemuth D, Fellous M. Cell. 1989;57:549–559. doi: 10.1016/0092-8674(89)90125-6. [DOI] [PubMed] [Google Scholar]
- 27.Wickham L, Duchaine T, Luo M, Nabi I R, DesGroseillers L. Mol Cell Biol. 1999;19:2220–2230. doi: 10.1128/mcb.19.3.2220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kohrmann M, Luo M, Kaether C, DesGroseillers L, Dotti C G, Kiebler M A. Mol Biol Cell. 1999;10:2945–2953. doi: 10.1091/mbc.10.9.2945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Talbot K, Miguel-Aliaga I, Mohaghegh P, Ponting C P, Davies K E. Hum Mol Genet. 1998;7:2149–2156. doi: 10.1093/hmg/7.13.2149. [DOI] [PubMed] [Google Scholar]
- 30.Porta A, Colonna-Romano S, Callebaut I, Franco A, Marzullo L, Kobayashi G S, Maresca B. Biochem Biophys Res Commun. 1999;3:605–613. doi: 10.1006/bbrc.1998.9894. [DOI] [PubMed] [Google Scholar]
- 31.Toyooka Y, Tsunekawa N, Takahashi Y, Matsui Y, Satoh M, Noce T. Mech Dev. 2000;93:139–149. doi: 10.1016/s0925-4773(00)00283-5. [DOI] [PubMed] [Google Scholar]
- 32.Guraya S S. Int Rev Cytol. 1979;59:249–321. doi: 10.1016/s0074-7696(08)61664-2. [DOI] [PubMed] [Google Scholar]
- 33.Wilsch-Brauninger M, Scharz H, Nusslein-Volhard C. J Cell Biol. 1997;139:817–829. doi: 10.1083/jcb.139.3.817. [DOI] [PMC free article] [PubMed] [Google Scholar]